Introduction
Lactoferrin is an 80-kDa member of the transferrin
family of iron-binding glycoproteins (1,2). This
protein is produced by epithelial cells and is found in mucosal
secretions, including tears, saliva, nasal exudates,
gastrointestinal fluids, and seminal and vaginal fluids (3). It is an important component of the
non-specific immune system, with antimicrobial properties against
bacteria, fungi and several viruses (4–7).
Numerous functions of lactoferrin relate to immune activation and
modulation (8).
Recent evidence indicates that lactoferrin also
possesses potent in vivo activity against cancer cells
(9–11). Subcutaneous administration of
lactoferrin demonstrated an inhibitory effect on both tumor-induced
angiogenesis and tumor growth in mice. In addition, intratumoral
injections of bovine lactoferrin slow the growth of fibrosarcoma
cells subcutaneously (s.c.) injected in mice (10). Furthermore, oral administration of
bovine lactoferrin to rats that had been previously injected with
azoxymethane to promote colon carcinogenesis results in an 83%
reduction in the incidence of colon adenocarcinomas (11). A similar effect was achieved by oral
administration of bovine lactoferrin, which may protect against
colon carcinogenesis, suggesting that lactoferrin may be an
effective therapeutic agent for cancer treatment.
Although the available evidence favors a direct
inhibitory effect of lactoferrin on cancer cell growth and
metastasis, little is known regarding the mechanism by which
lactoferrin exerts its anticancer activity. In this study, we
focused on the effect of human lactoferrin (hLF) overexpression via
adenoviral gene transfer on uterine cervical carcinoma in
vivo. We demonstrated that adenovirus carrying hLF (Ad-hLF)
significantly inhibited U14 solid tumor growth. Natural killer (NK)
cell activity and the number of CD4+ and CD8+
T lymphocyte cells in the peripheral blood of tumor-bearing mice
were increased by Ad-hLF. We also identified that the levels of
interleukin-2 (IL-2), interferon-γ (IFN-γ) and tumor necrosis
factor-α (TNF-α) were increased, and the IL-4 level was decreased
by Ad-hLF. Finally, vascular endothelial growth factor (VEGF)
expression in tumor tissues was downregulated by Ad-hLF. These
results suggest that tumor inhibition induced by lactoferrin is the
result of immunomodulation.
Materials and methods
Cell lines and animals
BJ5183 E. coli was purchased from Invitrogen
(Shanghai, China). The human embryonic kidney (HEK)-293 cell line
was purchased from Microbix Biosystem, Inc. (Toronto, ON, Canada).
The cervical cancer U14 cell line was obtained from the Institute
of Medical Material, Chinese Academy of Medical Sciences (Beijing,
China). The YAC-1 cell line was obtained from the Institute of
Biochemistry and Cell Biology (Shanghai, China). Female Kunming
mice (6–8 weeks old, weighing 18–22 g) were provided by the
Experimental Animal Center of Xiehe Medical University (Beijing,
China). The experimental use of mice was approved by the animal
ethics committee of Xiehe Medical University.
Construction of recombinant adenoviral
vectors
Construction of recombinant adenovirus was performed
as described previously (12).
Briefly, the internal ribosome entrance site (IRES) system was
employed to express hLF and green fluorescent protein (GFP) from
the same cytomegalovirus (CMV) promoter. pAd-hLF and pAd-GFP
plasmid vectors were purified through the BJ5183 E. coli and
then transfected into HEK-293 cells. Ad-hLF was purified by cesium
chloride ultracentrifugation at 80,000 × g for 20 h. A recombinant
adenovirus carrying the GFP protein under the control of the CMV
promoter (Ad-GFP) was used as a control vector. hLF expression in
U14 cells following Ad-hLF transfection was detected.
Western blot analysis
Cell extracts were separated by electrophoresis on
10 SDS-polyacrylamide gels, transferred onto nitrocellulose papers
and probed with a mouse anti-hLF antibody (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). The mouse anti-hLF
antibody was detected with a polyclonal goat anti-mouse Ig coupled
to horseradish peroxidase (HRP) followed by enhanced
chemiluminescence, with SuperSignal ECL western blotting detection
reagents (Pierce Chemical Co., Rockford, IL, USA).
In vivo studies
Mice (six per group) were s.c. injected with
1×107 cells/ml U14 cells into the left axilla and then
monitored daily for tumor growth. When tumors grew to 0.2–0.3
cm3, the mice were intratumorally (i.t.) injected with
Ad-hLF (1×109 pfu) or Ad-GFP (1×109 pfu).
Groups administered intratumorally with 100 μl phosphate-buffered
saline (PBS) and 25 mg/kg cyclophosphamide (CTX) once every other
day, for a total of seven times were used as the negative and
positive controls, respectively. Tumor volumes were calculated
using the equation V (mm3) = ab2/2, where a
is the largest diameter and b is the perpendicular diameter. On day
14, mice were sacrificed and tumor growth was determined.
NK cell activity in response to
Ad-hLF
Spleen cells were collected as effector cells.
Additionally, YAC-1 cells were cultured as target cells.
Subsequently, effector and YAC-1 cells were mixed according to the
ratio of 50:1 and cultured together. There were two control groups:
Natural release group, in which target cells were cultured with
RPMI-1640 medium; and maximum release group, in which target cells
were treated with 1% NP-40 solution. After the cells in each group
were incubated at 37°C for 2 h, the supernatants were collected and
transferred to another well to incubate for 10 min at 37°C. Then,
lactate dehydrogenase (LDH) substrate solution was added and
incubated for 10 min, followed by termination of the enzymatic
reaction with HCl. Finally, optical density in each group was
detected with am iMark 500-nm microplate reader (Bio-Rad, Hercules,
CA, USA) for analyzing NK cell activity.
Flow cytometry assay
To investigate the effect of Ad-hLF on peripheral
blood T-lymphocyte subpopulations of tumor-bearing mice, the
collected anticoagulated blood from the Ad-hLF-treated group was
diluted to 1×106 cells/ml, labeled with anti-mouse
monoclonal antibodies (4A Biotech Co., Ltd., Beijing, China) for 30
min and then incubated for 30 min at 4°C with rabbit anti-mouse
FITC-IgG monoclonal antibodies to CD4+ and
CD8+ (4A Biotech Co., Ltd.). Stained cells were examined
with an EPICS-XL FACSCalibur flow cytometer using EXPO 32 ADC
software (Beckman Coulter, Fullerton, CA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
Prior to sacrifice by cervical dislocation, the
blood of tumor-bearing mice was collected, deposited for 1 h at 4°C
and centrifuged for 15 min at 600 × g. Subsequently, the serum was
collected and IL-4, IL-2, IFN-γ and TNF-α levels were determined
using a commercial ELISA kit (Zhongshan Golden Bridge Biotechnology
Co., Ltd., Beijing, China) according to the manufacturer’s
instructions.
VEGF analyses by
immunohistochemistry
According to SP kit descriptions (Zhongshan Golden
Bridge Biotechnology Co., Ltd.), the paraffin sections were
deparaffinized and then dipped into gradient ethyl alcohol. After
the antigens were fixed, the sections were incubated for 10 min
with 3% H2O2 at room temperature and then the
rabbit anti-VEGF antibody (Santa Cruz Biotechnology, Inc.) was
added. Following incubation overnight at 4°C, a polyclonal goat
anti-rabbit Ig coupled to HRP secondary antibody (Santa Cruz
Biotechnology, Inc.) was added to the sections and incubated for 30
min at room temperature. Finally, the sections were dyed with
diaminobenzidine (DAB) and hematoxylin, and then observed under an
Olympus fluorescence microscope (Shanghai Lai Electronic Technology
Co., Ltd., Shanghai, China). The nuclei of positive cells
containing VEGF were dyed brown and the nuclei of negative cells
were dyed blue. Twenty fields of view were randomly selected. The
total number of cells was counted in each field to calculate the
percentage of positive cells.
Statistical analysis
Statistical analysis was performed using Student’s
t-test. Different letter superscripts between values in the
histograms indicate a significant difference (P<0.05) and same
letter superscripts between values indicate no significant
difference (P>0.05).
Results
hLF expression in U14 cells
It was reported that the IRES sequence allows the
expression of two different genes at high levels (13). We utilized the IRES system to
express hLF and GFP from the same CMV promoter. The GFP and hLF
genes were respectively cloned downstream and upstream of the IRES
region under the control of a CMV promoter. After the recombinant
adenoviral vector was constructed and transfected into HEK-293
cells, GFP proteins were clearly observed under a fluorescence
microscope, suggesting that these vectors were normally transfected
and proliferated in the HEK-293 cells. Subsequently, after a number
of the recombinant adenoviral vectors were obtained from the
HEK-293 cells, they were in turn transfected into the U14 cells,
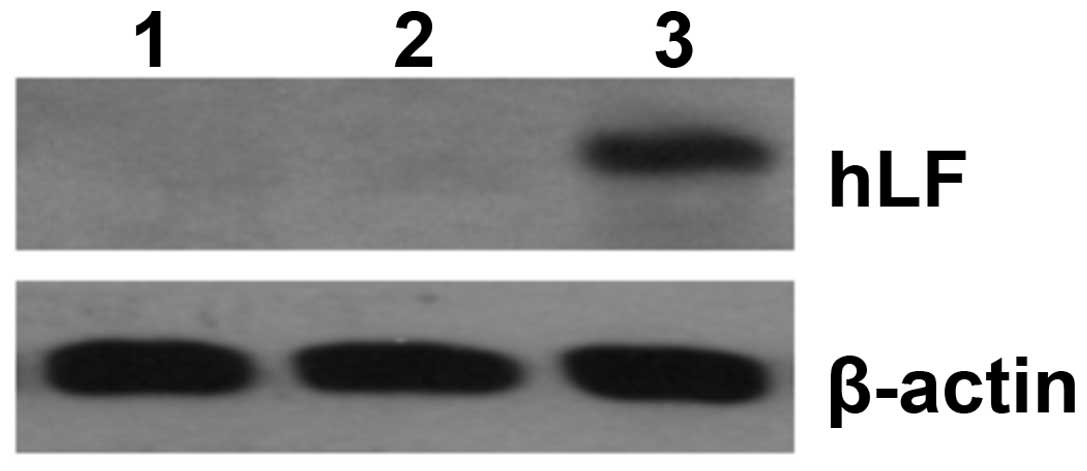
and western blotting revealed that the expression of hLF was
significantly increased in U14 cells (Fig. 1), which greatly inhibited the growth
of U14 cells.
Effects of Ad-hLF on tumor growth in
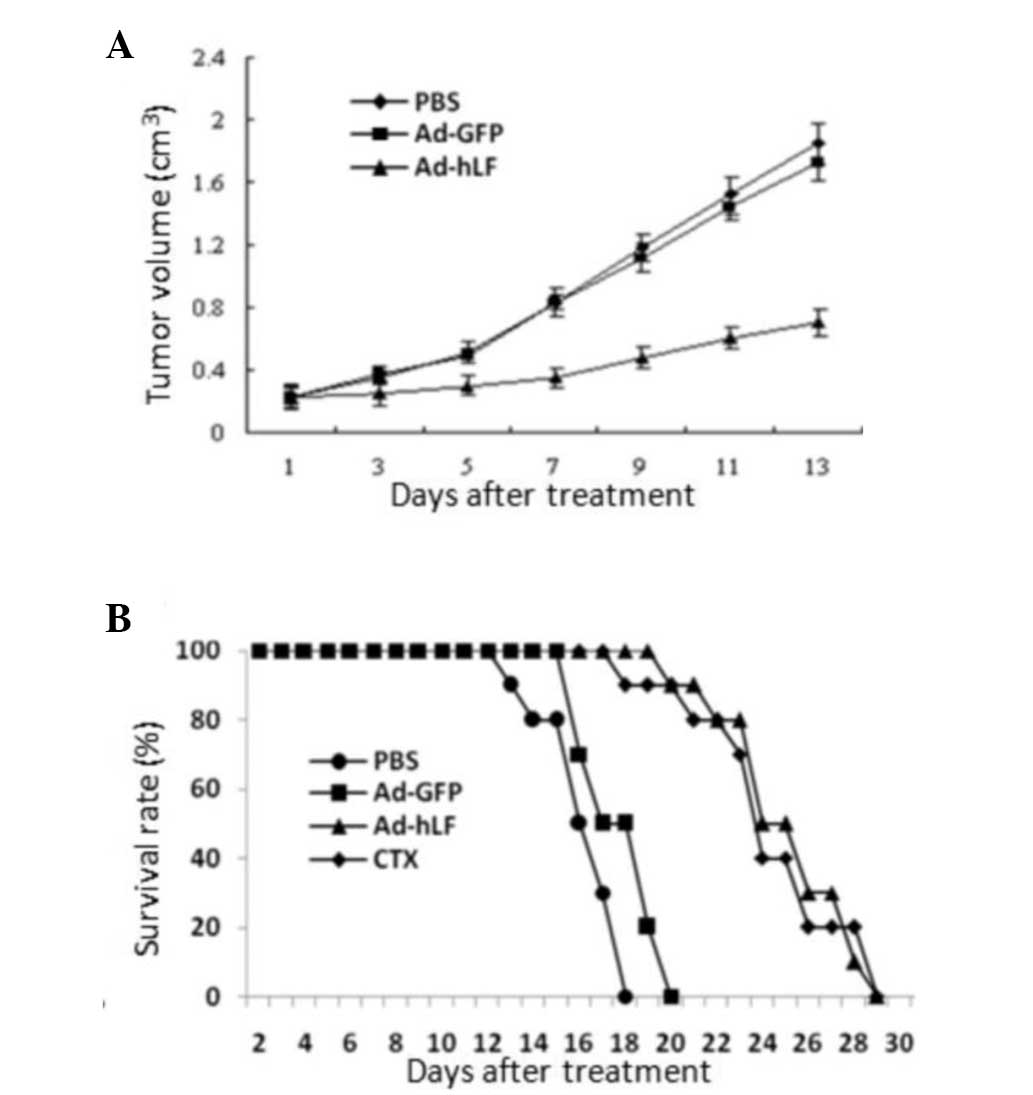
vivo
To evaluate the in vivo antitumor activity of
Ad-hLF, the U14 cells were i.t. injected with Ad-hLF
(1×109 pfu) or Ad-GFP (1×109 pfu) into
tumor-bearing mice, once every other day as described. Tumor growth
was monitored daily. The PBS- and Ad-GFP-treated tumors kept
growing uninterruptedly. The tumors in the Ad-hLF-treated group
were markedly smaller compared with those of the Ad-GFP-treated
group, when measured 14 days after seven injections (Fig. 2A). Moreover, survival of mice
treated with Ad-hLF treatment was significantly prolonged compared
with that of PBS- or Ad-GFP-treated mice (Fig. 2B). The lifespan of PBS- or
Ad-GFP-treated mice was ~20 days, while the mice treated with
Ad-hLF were still alive at the same time-point and their lifespans
were prolonged to ~30 days, close to the CTX-treated group. These
results indicate that Ad-hLF may suppress the growth of U14 solid
tumors in vivo.
NK cell activity in response to
Ad-hLF
NK cells act as cytolytic effector lymphocytes. NK
cells often lack antigen-specific cell surface receptors and are
therefore involved in innate immunity, i.e. they are able to react
immediately with no prior exposure to the pathogen (14). In mice and humans, NK cells play a
role in tumor immunosurveillance by directly inducing tumor cell
death (15,16). In the present study, we analyzed NK
cell activity through the use of LDH. Fig. 3 shows that the percentage of NK cell
activity in the Ad-hLF-treated group was significantly higher
compared with that of the PBS- or Ad-GFP-treated control groups
(P<0.05), which indicates that the activity of NK cells in
killing the tumor is increased by Ad-hLF.
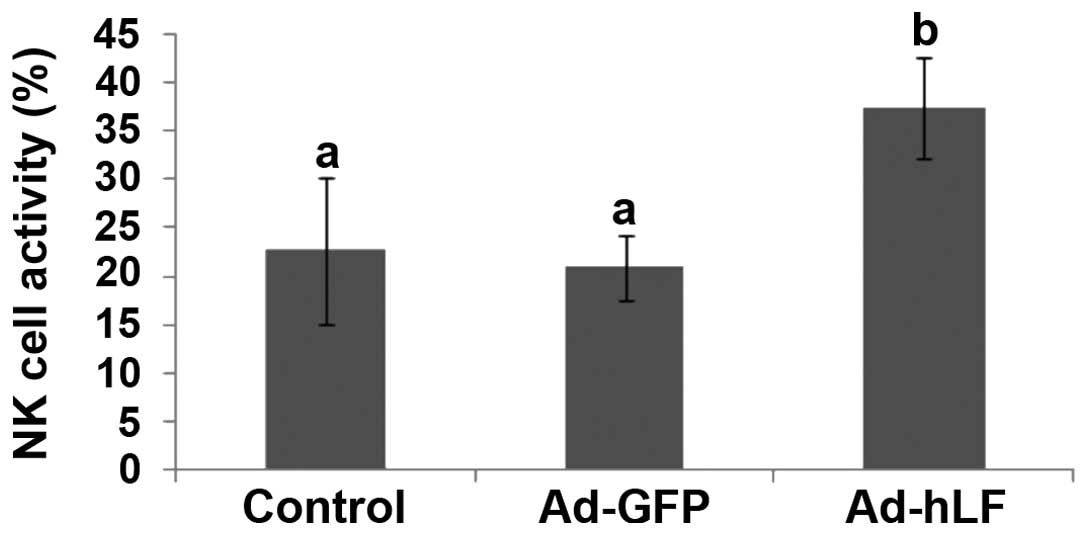 | Figure 3Spleen cells were the effector cells
and YAC-1 cells were the target cells. Effector and target cells
were mixed and cultured together. The two control groups were a
natural release group and a maximum release group. After the cells
were incubated, the supernatants were collected and then lactate
dehydrogenase substrate solution was added. Finally, the OD was
detected in each group with a 500-nm microplate reader. NK cell
activity was determined through the following formula: X =
(x−y)/(z−y), where X is NK cell activity, x is OD value of studied
group, y is OD value of natural release group and z is OD value of
maximum release group. NK cell activities in phosphate-buffered
saline-, Ad-GFP- and Ad-hLF-treated groups were 22.53±7.62,
20.85±3.45 and 37.38±5.12%, respectively. Different letter
superscripts between values indicate a significant difference
(P<0.05) and same letter superscripts between values indicate no
significant difference (P>0.05). OD, optical density; NK,
natural killer; Ad-GFP, adenovirus carrying green fluorescent
protein; Ad-hLF, adenovirus carrying human lactoferrin. |
Effect of Ad-hLF on peripheral blood T
lymphocyte subpopulations
The development and progression of tumors is greatly
affected by the function of T lymphocytes (17). Mature T lymphocytes mainly
differentiate into CD4+ and CD8+ cells.
CD8+ cells activated by CD4+ cells destroy
virally infected cells and tumor cells. The aforementioned two
subpopulations in tumor-bearing mice were quantified by flow
cytometry. The results demonstrated that the number of
CD4+ and CD8+ T lymphocyte cells in the
peripheral blood of tumor-bearing mice treated with Ad-hLF
significantly increased compared with that of the control group
(P<0.05) and was near to that of normal mice (P>0.05)
(Fig. 4). These results demonstrate
that Ad-hLF promotes the development of CD4+ and
CD8+ cells in tumor-bearing mice.
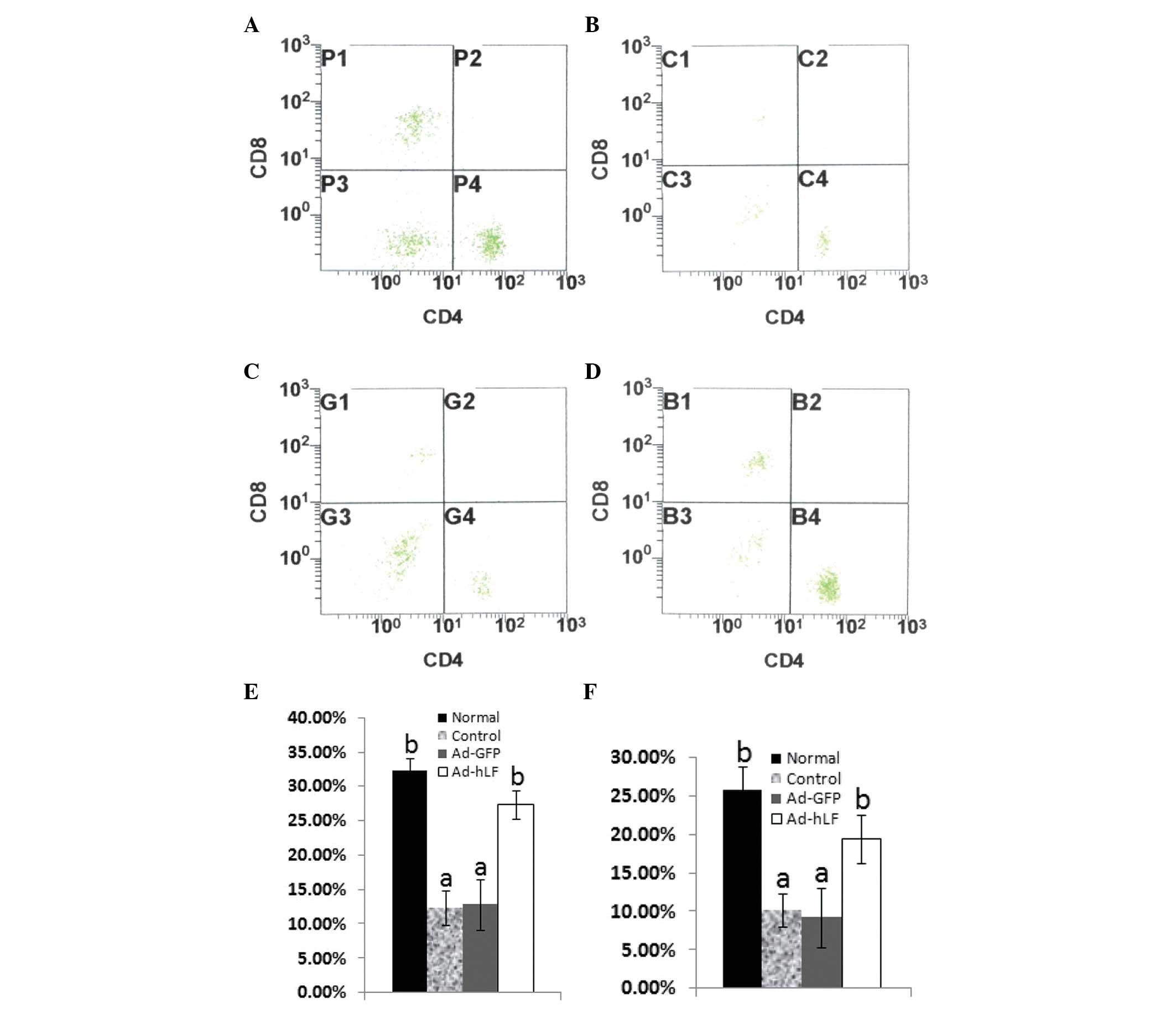 | Figure 4Collected anticoagulated blood from
each group was diluted to 1×106 cell/ml, labeled with
anti-mouse monoclonal antibodies and then incubated for 30 min with
rabbit anti-mouse FITC-IgG monoclonal antibodies to CD4+
and CD8+. Stained cells were examined with an EPICS-XL
FACSCalibur flow cytometer using Expo 32 ADC software. (A) Normal
mice; (B) PBS-treated mice; (C) Ad-GFP-treated mice; (D)
Ad-hLF-treated mice. (E) The percentages of CD4+ T cells
in PBS-, Ad-GFP- and Ad-hLF-treated mice and normal mice were
12.39±2.53, 12.85±3.62, 27.38±2.12 and 32.25±1.87%, respectively.
(F) The percentages of CD8+ T cells in PBS-, Ad-GFP- and
Ad-hLF-treated mice and normal mice were 10.24±2.17, 9.25±3.95,
19.52±3.14 and 25.79±3.14%, respectively. Different letter
superscripts between values indicate a significant difference
(P<0.05) and same letter superscripts between values indicate no
significant difference (P>0.05). PBS, phosphate-buffered saline;
Ad-GFP, adenovirus carrying green fluorescent protein; Ad-hLF,
adenovirus carrying human lactoferrin. |
Effects of Ad-hLF on IL-2, IL-4, IFN-γ
and TNF-α
After T helper (Th) cells are activated, they divide
rapidly and secrete cytokines, including IL-2, IL-4, IFN-γ and
TNF-α, which regulate or assist in the active immune response
(18). To determine the effects of
Ad-hLF on the secretion of cytokines, the serum levels of IL-2,
IL-4, IFN-γ and TNF-α in tumor-bearing mice were analyzed by ELISA.
When Ad-hLF was administered to the tumor-bearing mice, the levels
of serum IFN-γ and IL-2 were significantly increased and were close
to the levels in the normal group (P<0.05) (Fig. 5A and B). Compared with the other
three groups, the level of serum IL-4 was significantly decreased,
but the levels of serum TNF-α were markedly increased in the
Ad-hLF-treated group (P<0.05) (Fig.
5C and D).
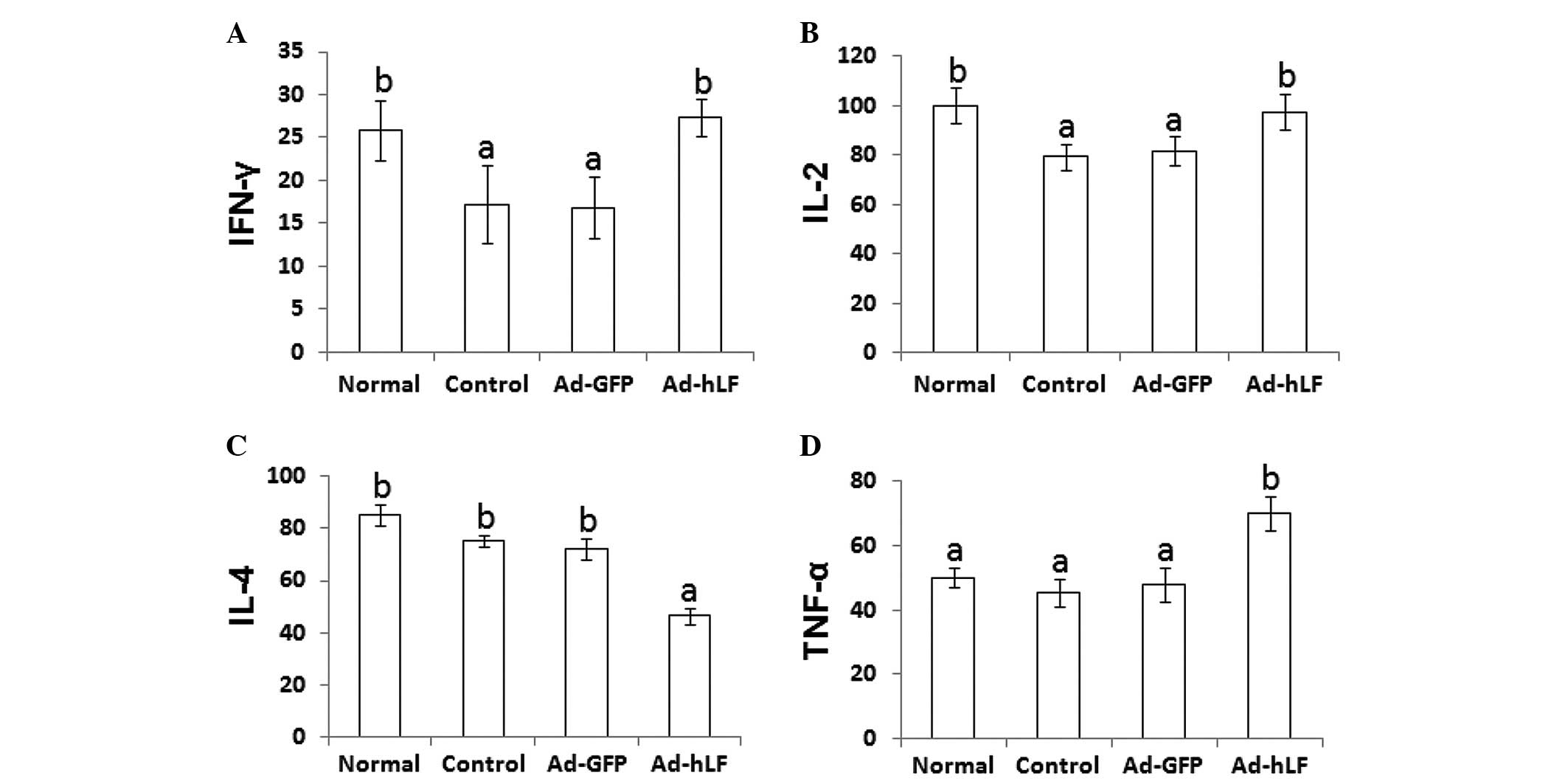 | Figure 5Blood from tumor-bearing mice was
collected, deposited for 1 h at 4°C and centrifuged for 15 min.
Subsequently, the serum was collected and IFN-γ, IL-2, IL-4 and
TNF-α levels were determined using an enzyme-linked immunosorbent
assay kit. (A) IFN-γ levels in phosphate-buffered saline (PBS)-,
Ad-GFP- and Ad-hLF-treated mice and normal mice were 17.25±4.53,
16.85±3.62, 27.38±2.12 and 25.86±3.43 pg/ml, respectively. (B) IL-2
levels in PBS-, Ad-GFP- and Ad-hLF-treated mice and normal mice
were 79.37±5.16, 81.75±6.15, 97.45±7.12, 100.13±7.27 pg/ml,
respectively. (C) IL-4 levels in PBS-, Ad-GFP- and Ad-hLF-treated
mice and normal mice were 75.24±2.17, 72.25±3.95, 46.52±3.14 and
85.13±4.18 pg/ml, respectively. (D) TNF-α levels in PBS-, Ad-GFP-
and Ad-hLF-treated mice and normal mice were 45.32±4.35,
48.03±5.14, 70.12±5.15 and 50.21±3.12 pg/ml, respectively.
Different letter superscripts between values indicate a significant
difference (P<0.05) and same letter superscripts between values
indicate no significant difference (P>0.05). IFN-γ,
interferon-γ; IL, interleukin; TNF-α, tumor necrosis factor-α;
Ad-GFP, adenovirus carrying green fluorescent protein; Ad-hLF,
adenovirus carrying human lactoferrin. |
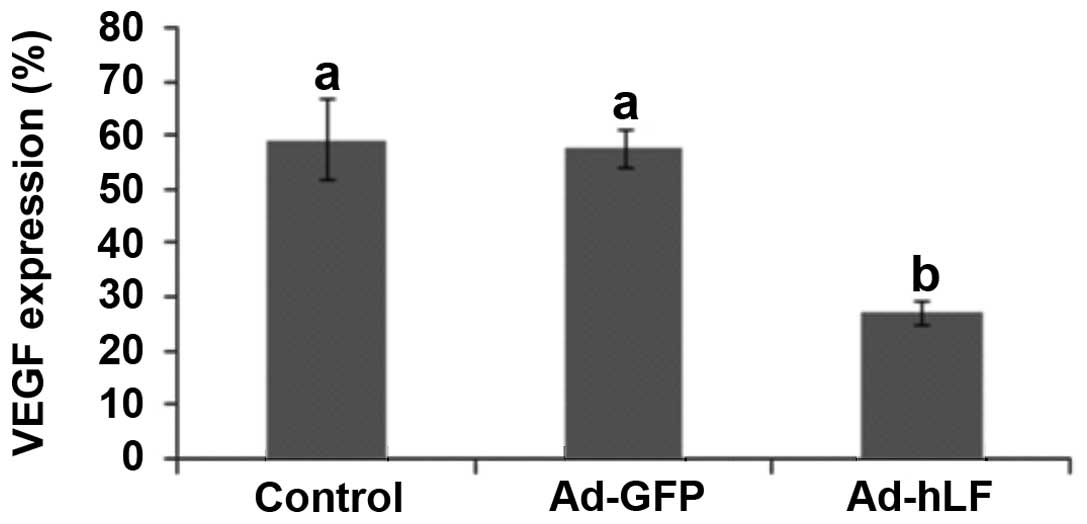
VEGF expression in tumor tissue was
decreased by Ad-hLF
VEGF is a signal protein produced by cells that
stimulates vasculogenesis and angiogenesis. When VEGF is
overexpressed, it contributes to disease. Cancers that express VEGF
are able to grow and metastasize (19). After mice were treated with Ad-hLF,
we detected VEGF expression in tumor tissues using
immunohistochemistry. Brown nuclei indicated VEGF-positive cells
and blue nuclei indicated VEGF-negative cells. Our results
demonstrated that numerous cell nuclei were brown, suggesting that
VEGF was greatly expressed in the PBS- and Ad-GFP-treated control
groups (data not shown). However, few cell nuclei were brown, but a
number were blue in the Ad-hLF-treated group (data not shown),
suggesting that VEGF was slightly expressed. Statistical analysis
certified that the VEGF expression level in the Ad-hLF-treated
group was lower compared with that of the other two groups
(P<0.05) (Fig. 6). These results
show that VEGF expression in tumor tissue is blocked by hLF.
Discussion
Lactoferrin is becoming an increasingly important
strategy for inhibiting carcinogenesis and tumor growth. Direct
inhibition of cellular growth is one mechanism by which lactoferrin
may inhibit the growth of numerous cancers. Lactoferrin treatment
reduces colonic carcinogenesis in rats and decreases solid tumor
growth and metastases in mice (11,20).
hLF acts against the growth of solid tumors and the development of
metastases in mice (21). Bovine
lactoferrin inhibits lung metastasis of B16 melanoma and colon-26
tumor cells in mice and is protective against tongue, esophagus,
intestinal, lung and bladder carcinogenesis in rats (9,22–25).
The growth of cervical cancer is inhibited by Ad-hLF through the
regulation of apoptotic factors (26). The present study extended the
observation to cervical cancer and demonstrated the effects of
Ad-hLF on the immune response of U14 cervical carcinoma-bearing
mice. Our results indicated that the inhibitory roles of Ad-hLF on
cervical cancer may be related to its upregulation of the immune
response against tumors.
NK cells are a type of cytotoxic lymphocyte critical
to the innate immune system. The role NK cells play is analogous to
that of cytotoxic T cells in the vertebrate adaptive immune
response (27). NK cells provide
rapid responses to virally infected cells and respond to tumor
formation, acting approximately three days after infection
(28). After NK cell activity is
promoted by hLF, NK cells release full granzymes to kill tumor
cells. Small granules in the NK cell cytoplasm contain proteins,
such as perforin and proteases known as granzymes. Upon release in
close proximity to a cell marked for cell death, perforin forms
pores in the cell membrane of the target cell, creating an aqueous
channel through which the granzymes and associated molecules enter,
inducing either apoptosis or osmotic cell lysis (29).
Cytokines play a crucial role in NK cell activation.
Cytokines involved in NK activation include IL-12, IL-15, IL-18,
IL-2 and CCL5 (29–31). Based on this, we investigated the
level of serum IL-2 to determine if it was affected by hLF. In the
present study, in which tumor-bearing mice were treated with
Ad-hLF, the level of serum IL-2 was increased, which indicates that
hLF activates NK cells. Once activated, NK cells work to control
viral infections by secreting IFN-γ and TNF-α. IFN-γ activates
macrophages for phagocytosis and lysis, and TNF-α promotes direct
killing of tumor cells by NK cells (30,32).
The increased levels of IFN-γ and TNF-α indicate that hLF is
involved in NK cell activation. NK cells promote the expression of
Fas on cancer cells. Fas is not normally expressed on tumor cells
and therefore NK cells aided Fas-dependent apoptosis upon binding
with Fas ligand (FasL)-expressing NK cells (33).
Aside from the inhibition of tumor cell growth by NK
cells, T cells play an important role in tumor inhibition. Mature T
lymphocytes mainly differentiate into CD4+ and
CD8+ cells. CD4+ and CD8+ T cells
strongly prevent tumor development (34,35).
CD4+ T cells, which are a type of Th cell, induce the
maturation of B cells into plasma cells and the activation of
cytotoxic T cells, including CD8+ cells (36). CD8+ T cells destroy
virally infected cells and tumor cells (37). We found that the number of
CD4+ and CD8+ T cells in the peripheral blood
of tumor-bearing mice treated with Ad-hLF significantly increased
compared with that of the control group, which suggests that Ad-hLF
increases the expression of CD4+ and CD8+ T
cells and may activate them. Through the help of activated
CD4+ T cells, CD8+ T cells are activated and
migrate to the tumor site, producing a specific cytotoxic effect.
Perforin and granzymes are released from CD8+ T cells.
Perforin forms pores in the cell membrane of the target cell,
creating an aqueous channel through which the granzymes enter,
which leads to cell apoptosis via degradation of DNA of the target
cell or stimulation of the FasL/Fas pathway.
Additionally, Th cells include two subsets with
different functions: Th1 and Th2 cells. Once activated, these Th
cells divide rapidly and secrete cytokines that regulate or assist
in the active immune response. Th1 cells mainly secrete IL-2,
IL-12, IFN-γ and TNF-α, which are involved in cellular immunity and
graft rejection (38,39). Th2 cells mainly secrete IL-4, IL-5,
IL-6 and IL-10, which are involved in humoral immune and allergic
reactions (40). IL-2 or IFN-γ may
induce NK cell-induced tumor death and IFN-γ directly causes DNA
fragmentation of tumor cells, which demonstrates that Th1 cells
present a powerful status against tumors (41,42).
However, IL-4 promotes IL-10 in tumor tissue and the latter
interferes with the expression of inflammatory factors, including
IL-12 and IFN-γ, blocks the activation of NK cells and reduces the
antigen formation of tumor cells in order to escape cytoimmunity
against tumor growth (43–45). Therefore, a Thl and Th2 cell balance
maintains a normal state of the body by secreting cytokines. If
these cytokines in Thl and Th2 cells are dysregulated, the
cytoimmunity is destroyed. In the present study, the levels of IL-2
and IFN-γ were significantly increased, but the levels of IL-4 were
reduced by hLF in tumor-bearing mice. This caused the Th1/Th2
balance to favor Th1 progression, promoting the antitumor response.
It was also demonstrated that TNF-α levels were increased by hLF
and increased the antitumor effect. The aforementioned results
demonstrated that hLF may inhibit the growth of cervical cancer by
rescuing the balance between Th1 and Th2 cells and strongly
activating Th1 cells in tumor-bearing mice.
Although we determined that cervical cancer was
inhibited through the cytoimmunity initiated by hLF, it remains
unclear how to directly inhibit the tumor by hLF. Due to the
inhibitory role of IFN-γ with regard to tumor vascular formation,
the expression of VEGF in tumor tissue was investigated (46). VEGF is a signal protein produced by
cells and it stimulates vasculogenesis and angiogenesis (47). When VEGF is overexpressed, it
contributes to disease (48). Solid
cancers, including cervical cancer, do not grow beyond a limited
size without an adequate blood supply. Once released, VEGF may
elicit several responses. It may cause a cell to survive, move or
further differentiate (49). After
Ad-hLF was i.t. injected, VEGF expression in tumor tissue was
significantly decreased, which suggests that Ad-hLF may directly
downregulate VEGF so that the vascular components in the tumor are
not formed, causing inhibition of cervical cancer.
In conclusion, the immune response of tumor-bearing
mice was upregulated by hLF. hLF increased the activity of NK cells
and rescued the balance of serum cytokines in tumor tissue.
Furthermore, hLF upregulated the function of T cells and blocked
the expression of VEGF. Due to the initiation of cytoimmunity
against tumor cells by hLF, cervical cancer was suppressed.
Acknowledgements
This study was supported by the grants ‘Hpa
expression and its relationship with MMP in tumor cells’
(2011BSJJ004) and ‘Study of EGF-mediated signal pathways in cell
mitosis’ (QN2011010) from the Northwest A&F University,
Yangling, China.
References
|
1
|
Metz-Boutigue MH, Jollès J, Mazurier J,
Schoentgen F, Legrand D, Spik G, et al: Human lactotransferrin:
amino acid sequence and structural comparisons with other
transferrins. Eur J Biochem. 145:659–676. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanyshkova TG, Buneva VN and Nevinsky GA:
Lactoferrin and its biological functions. Biochemistry (Mosc).
66:1–7. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nuijens JH, van Berkel PH and Schanbacher
FL: Structure and biological actions of lactoferrin. J Mammary
Gland Biol Neoplasia. 1:285–295. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levay PF and Viljoen M: Lactoferrin: a
general review. Haematologica. 80:252–267. 1995.PubMed/NCBI
|
|
5
|
Vorland LH: Lactoferrin: a multifunctional
glycoprotein. APMIS. 107:971–981. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu T, Zhang YZ and Wu XF: High level
expression of functionally active human lactoferrin in silkworm
larvae. J Biotechnol. 118:246–256. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van der Strate BW, Beljaars L, Molema G,
Harmsen MC and Meijer DK: Antiviral activities of lactoferrin.
Antiviral Res. 52:225–239. 2001.
|
|
8
|
Cumberbatch M, Dearman RJ, Uribe-Luna S,
Headon DR, Ward PP, Conneely OM and Kimber I: Regulation of
epidermal Langerhans cell migration by lactoferrin. Immunology.
100:21–28. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoo YC, Watanabe S, Watanabe R, Hata K,
Shimazaki K and Azuma I: Bovine lactoferrin and lactoferricin, a
peptide derived from bovine lactoferrin, inhibit tumor metastasis
in mice. Jpn J Cancer Res. 88:184–190. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cho E, Smith-Warner SA, Spiegelman D,
Beeson WL, van den Brandt PA, Colditz GA, et al: Dairy foods,
calcium, and colorectal cancer: a pooled analysis of 10 cohort
studies. J Natl Cancer Inst. 96:1015–1022. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsuda H, Sekine K, Nakamura J, Ushida Y,
Kuhara T, Takasuka N, et al: Inhibition of azoxymethane initiated
colon tumor and aberrant crypt foci development by bovine
lactoferrin administration in F344 rats. Adv Exp Med Biol.
443:273–284. 1998. View Article : Google Scholar
|
|
12
|
He TC, Zhou S, da Costa LT, Yu J, Kinzler
KW and Vogelstein B: A simplified system for generating recombinant
adenoviruses. Proc Natl Acad Sci USA. 95:2509–2514. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rees S, Coote J, Stables J, Goodson S,
Harris S and Lee MG: Bicistronic vector for the creation of stable
mammalian cell lines that predisposes all antibiotic-resistant
cells to express recombinant protein. Biotechniques. 20:102–104.
106108–110. 1996.
|
|
14
|
Cheng M, Zhang J, Jiang W, Chen Y and Tian
Z: Natural killer cell lines in tumor immunotherapy. Front Med.
6:56–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kiessling R, Klein E and Wigzell H:
‘Natural’ killer cells in the mouse. I Cytotoxic cells with
specificity for mouse Moloney leukemia cells Specificity and
distribution according to genotype. Eur J Immunol. 5:112–117.
1975.
|
|
16
|
Glässner A, Eisenhardt M, Krämer B, Körner
C, Coenen M, Sauerbruch T, et al: NK cells from HCV-infected
patients effectively induce apoptosis of activated primary human
hepatic stellate cells in a TRAIL-, FasL- and NKG2D-dependent
manner. Lab Invest. 92:967–977. 2012.
|
|
17
|
Kuss I, Hathaway B, Ferris RL, Gooding W
and Whiteside TL: Decreased absolute counts of T lymphocyte subsets
and their relation to disease in squamous cell carcinoma of the
head and neck. Clin Cancer Res. 10:3755–3762. 2004. View Article : Google Scholar
|
|
18
|
Tembhre MK and Sharma VK: T-helper and
regulatory T-cell cytokines in the peripheral blood of patients
with active alopecia areata. Br J Dermatol. 169:543–548. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tie J and Desai J: Antiangiogenic
therapies targeting the vascular endothelial growth factor
signaling system. Crit Rev Oncog. 17:51–67. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masuda C, Wanibuchi H, Sekine K, Yano Y,
Otani S, Kishimoto T, et al: Chemopreventive effects of bovine
lactoferrin on N-butyl-N-(4-hydroxybutyl)nitrosamine-induced rat
bladder carcinogenesis. Jpn J Cancer Res. 91:582–588. 2000.
View Article : Google Scholar
|
|
21
|
Bezault J, Bhimani R, Wiprovnick J and
Furmanski P: Human lactoferrin inhibits growth of solid tumors and
development of experimental metastases in mice. Cancer Res.
54:2310–2012. 1994.PubMed/NCBI
|
|
22
|
Iigo M, Kuhara T, Ushida Y, Sekine K,
Moore MA and Tsuda H: Inhibitory effects of bovine lactoferrin on
colon carcinoma 26 lung metastasis in mice. Clin Exp Metastasis.
17:35–40. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ushida Y, Sekine K, Kuhara T, Takasuka N,
Iigo M and Tsuda H: Inhibitory effects of bovine lactoferrin on
intestinal polyposis in the Apc(Min) mouse. Cancer Lett.
134:141–145. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ushida Y, Sekine K, Kuhara T, Takasuka N,
Iigo M, Maeda M and Tsuda H: Possible chemopreventive effects of
bovine lactoferrin on esophagus and lung carcinogenesis in the rat.
Jpn J Cancer Res. 90:262–267. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tanaka T, Kawabata K, Kohno H, Honjo S,
Murakami M, Ota T and Tsuda H: Chemopreventive effect of bovine
lactoferrin on 4-nitroquinoline 1-oxide-induced tongue
carcinogenesis in male F344 rats. Jpn J Cancer Res. 91:25–33. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li WY, Li QW, Han ZS, Jiang ZL, Yang H, Li
J and Zhang XB: Growth suppression effects of recombinant
adenovirus expressing human lactoferrin on cervical cancer in vitro
and in vivo. Cancer Biother Radiopharm. 26:477–483. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arina A, Murillo O, Dubrot J, Azpilikueta
A, Alfaro C, Pérez-Gracia JL, et al: Cellular liaisons of natural
killer lymphocytes in immunology and immunotherapy of cancer.
Expert Opin Biol Ther. 7:599–615. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Trapani JA and Smyth MJ: Functional
significance of the perforin/granzyme cell death pathway. Nat Rev
Immunol. 2:735–747. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kannan Y, Yu J, Raices RM, Seshadri S, Wei
M, Caligiuri MA and Wewers MD: IκBζ augments IL-12- and
IL-18-mediated IFN-γ production in human NK cells. Blood.
117:2855–2863. 2011.
|
|
30
|
Raja A: Immunology of tuberculosis. Indian
J Med Res. 120:213–232. 2004.
|
|
31
|
Van Elssen CH, Vanderlocht J, Oth T,
Senden-Gijsbers BL, Germeraad WT and Bos GM:
Inflammation-restraining effects of prostaglandin E2 on natural
killer-dendritic cell (NK-DC) interaction are imprinted during DC
maturation. Blood. 118:2473–2482. 2011.
|
|
32
|
Reefman E, Kay JG, Wood SM, Offenhäuser C,
Brown DL, Roy S, et al: Cytokine secretion is distinct from
secretion of cytotoxic granules in NK cells. J Immunol.
184:4852–4862. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Smyth MJ, Hayakawa Y, Takeda K and Yagita
H: New aspects of natural-killer-cell surveillance and therapy of
cancer. Nat Rev Cancer. 2:850–861. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shanker A, Buferne M and Schmitt-Verhulst
AM: Cooperative action of CD8 T lymphocytes and natural killer
cells controls tumour growth under conditions of restricted T-cell
receptor diversity. Immunology. 129:41–54. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jenq RR, Curran MA, Goldberg GL, Liu C,
Allison JP and van den Brink MR: Repertoire enhancement with
adoptively transferred female lymphocytes controls the growth of
pre-implanted murine prostate cancer. PLoS One. 7:e352222012.
View Article : Google Scholar
|
|
36
|
Banchereau J and Steinman RM: Dendritic
cells and the control of immunity. Nature. 392:245–252. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Blanchard DK, Wei S, Duan C, Pericle F,
Diaz JI and Djeu JY: Role of extracellular adenosine triphosphate
in the cytotoxic T-lymphocyte-mediated lysis of antigen presenting
cells. Blood. 85:3173–3182. 1995.PubMed/NCBI
|
|
38
|
Hamza T, Barnett JB and Li B: Interleukin
12 a key immunoregulatory cytokine in infection applications. Int J
Mol Sci. 11:789–806. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Seger J, Zorzella-Pezavento SF, Pelizon
AC, Martins DR, Domingues A and Sartori A: Decreased production of
TNF-alpha by lymph node cells indicates experimental autoimmune
encephalomyelitis remission in Lewis rats. Mem Inst Oswaldo Cruz.
105:263–268. 2010. View Article : Google Scholar
|
|
40
|
Hartenstein B, Teurich S, Hess J, Schenkel
J, Schorpp-Kistner M and Angel P: Th2 cell-specific cytokine
expression and allergen-induced airway inflammation depend on JunB.
EMBO J. 21:6321–6329. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Metelitsa LS, Naidenko OV, Kant A, Wu HW,
Loza MJ, Perussia B, et al: Human NKT cells mediate antitumor
cytotoxicity directly by recognizing target cell CD1d with bound
ligand or indirectly by producing IL-2 to activate NK cells. J
Immunol. 167:3114–3122. 2001. View Article : Google Scholar
|
|
42
|
Xu Z, Hurchla MA, Deng H, Uluçkan O, Bu F,
Berdy A, et al: Interferon-gamma targets cancer cells and
osteoclasts to prevent tumor-associated bone loss and bone
metastases. J Biol Chem. 284:4658–4666. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Koch F, Stanzl U, Jennewein P, Janke K,
Heufler C, Kämpgen E, et al: High level IL-12 production by murine
dendritic cells: upregulation via MHC class II and CD40 molecules
and downregulation by IL-4 and IL-10. J Exp Med. 184:741–746.
1996.PubMed/NCBI
|
|
44
|
Flesch IE, Hess JH, Oswald IP and Kaufmann
SH: Growth inhibition of Mycobacterium bovis by IFN-gamma
stimulated macrophages: regulation by endogenous tumor necrosis
factor-alpha and by IL-10. Int Immunol. 6:693–700. 1994.
|
|
45
|
Alcami A and Koszinowski UH: Viral
mechanisms of immune evasion. Mol Med Today. 6:365–372. 2000.
View Article : Google Scholar
|
|
46
|
Hayakawa Y, Takeda K, Yagita H, Smyth MJ,
Van Kaer L, Okumura K and Saiki I: IFN-gamma-mediated inhibition of
tumor angiogenesis by natural killer T-cell ligand,
alpha-galactosylceramide. Blood. 100:1728–1733. 2002.PubMed/NCBI
|
|
47
|
Davis-Smyth T, Chen H, Park J, Presta LG
and Ferrara N: The second immunoglobulin-like domain of the VEGF
tyrosine kinase receptor Flt-1 determines ligand binding and may
initiate a signal transduction cascade. EMBO J. 15:4919–4927.
1996.
|
|
48
|
Carmeliet P: Angiogenesis in health and
disease. Nat Med. 9:653–660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sukhramani PS and Suthar MP: VEGF
inhibitors for cancer therapy. IJPSDR. 2:1–11. 2010.
|