Introduction
Breast cancer is the most common type of malignant
tumor and the second highest mortality among females with cancer
(1). Survival rates for breast
cancer have greatly increased with significant improvement in
surgical technology and therapy regimens over the last three
decades, particularly for early-stage breast cancer. However, no
effective treatments currently exist for metastatic breast cancer
(2,3). As cancer development largely results
from the uncontrolled growth of malignant cells, in which cell
proliferation surpasses cell death, deregulation of apoptosis,
which occurs frequently in a vast majority of cancer types, has
become a non-negligible target for anticancer strategies (4,5).
Proapoptotic compounds, derived from synthetic chemistry or natural
sources, are also under active investigation for their therapeutic
effects and for their mode of actions against various types of
cancer. Over the past decades, considerable effort has been
directed to using natural products as a source of novel anticancer
drugs in the fight against the challenge that a number of types of
cancer remain incurable by currently available therapeutic
approaches.
Morinda citrifolia L. (Rubiaceae), commonly
known as noni, is a small evergreen tree or shrub that is widely
distributed throughout the pacific islands, Southeast Asia and
other tropical and semitropical areas. It has been widely used in
therapeutic preparations for centuries, owing to its
anti-inflammatory, antibacterial, antiviral, antifungal and
antitumor properties (6–9). Damnacanthal, an anthraquinone
compound, was isolated from the roots of Morinda citrifolia
L. and identified as a potent inhibitor of different types of
human tumor cells, including human T-lymphoblastic and acute
promyelocytic leukemia (10) and
breast carcinoma (11). These
anticancer effects of damnacanthal were identified through
interfering with the cell cycle, inducing apoptosis and inhibiting
the invasive potential of cancer (12–14).
However, the underlying molecular mechanisms of these anticancer
effects remain under investigation. In the present study, the human
breast cancer MCF-7 cell line was used to investigate the effects
of damnacanthal on the breast cancer cell proliferation, apoptosis,
cell cycle arrest and apoptotic gene expression.
Materials and methods
Compound
Damnacanthal was obtained from the Faculty of
Applied Sciences, Mara University of Technology (Shah Alam,
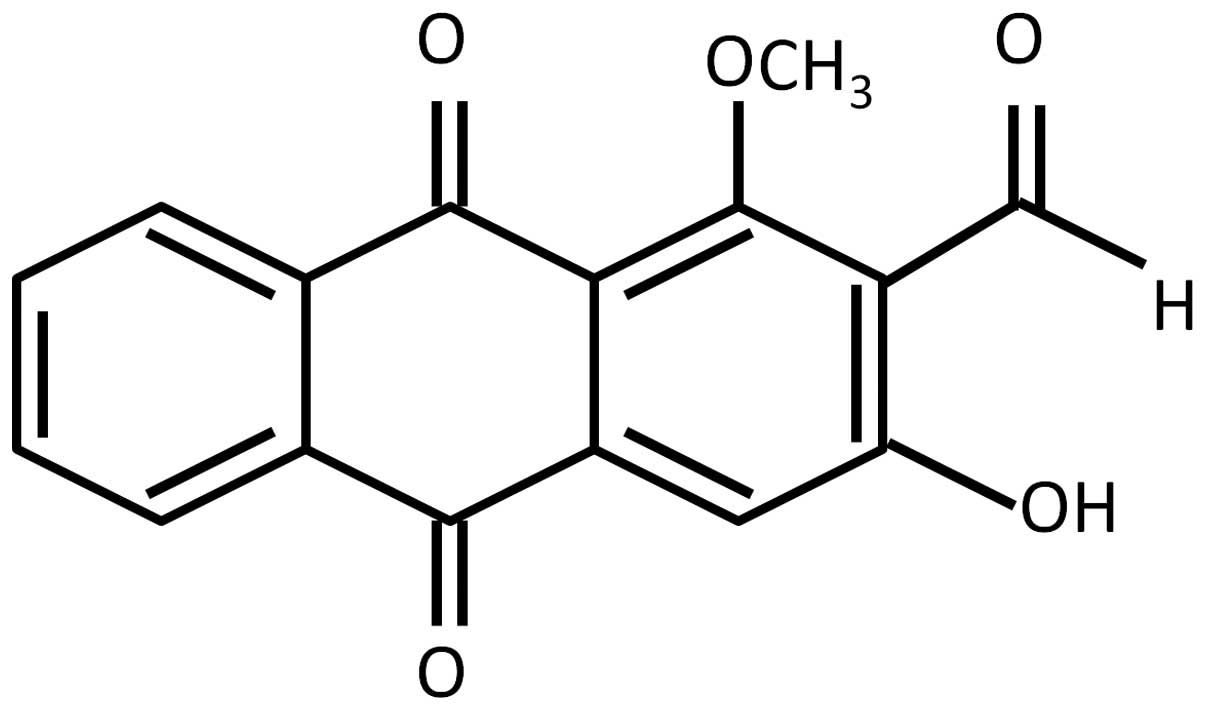
Malaysia) and used as received. The structure of damnacanthal is
shown in Fig. 1.
Cell culture
MCF-7 cells, a human breast cancer cell line
obtained from the American Type Culture Collection (Rockville, MD,
USA), were cultured as monolayers in RPMI-1640 medium
(Sigma-Aldrich, St. Louis, MO, USA) containing 10% (vol/vol) fetal
bovine serum (Gibco, Grand Island, NY, USA), 100 U/ml penicillin
(Sigma-Aldrich) and 100 μg/ml streptomycin (Sigma-Aldrich), at 37°C
in a humidified environment containing 5% CO2 and 95%
air.
Cytotoxicity assay
The MCF-7 cells were seeded in 96-well microtiter
plates (5,000 cells/well). Following 24 h of incubation in the
appropriate medium, cells were treated with various concentrations
of damnacanthal for an additional 72 h of culture. Next, 20 μl
stock MTT solution (Calbiochem, Darmstadt, Germany) was added to
each well (final concentration, 0.5 mg/ml) for an additional 4 h of
incubation (37°C; 5% CO2). Then, 200 μl
dimethylsulfoxide (Sigma-Aldrich)was added to each well and the
absorbance at 570 nm was determined. By MTT method, cell numbers
were obtained as absorbance values at 570 nm. The results were
expressed as viability compared with that of vehicle-treated cells.
Each treatment had three independent plates and the representative
results were reproducible in three independent experiments. The
selectivity of the cytotoxicity of damnacanthal on the MCF-7 cell
line was also evaluated.
Quantification of apoptosis
Damnacanthal-induced cell death in MCF-7 cells was
quantified using propidium iodide (PI) (Sigma-Aldrich) and
acridine-orange (AO, Sigma-Aldrich) double staining according to
standard procedures and examined under fluorescence microscope
(Eclipse Ti, Nikon, Melville, NY, USA). Briefly, treatment was
performed in a 25-ml culture flask. MCF-7 cells were plated at a
concentration of 1×106 cells/ml and treated with
damnacanthal at IC50 concentration. Flasks were
incubated in an atmosphere of 5% CO2 at 37°C for 72 h.
The cells were then spun down at 1,000 × g for 10 min. Supernatant
was discarded and the cells were washed twice using
phosphate-buffered saline (PBS) following centrifugation at 1,000 ×
g for 10 min to remove the remaining media. In total, 10 μl
fluorescent dyes, AO (10 μg/ml) and PI (10 μg/ml), were added into
the cellular pellet at equal volumes. Freshly stained cell
suspension was dropped onto a glass slide and covered by a
coverslip. Slides were observed under ultraviolet (UV)-fluorescence
microscope within 30 min prior to the fading of the fluorescence
color. All the treatments and time points were performed in three
individual experiments. AO and PI are intercalating nucleic
acid-specific fluorochromes that emit green and orange
fluorescence, respectively, when bound to DNA. Of the two, only AO
crosses the plasma membrane of viable and early apoptotic cells.
Viewed by fluorescence microscopy, viable cells appeared to exhibit
green nuclei with intact structures, while apoptotic cells
exhibited bright-green nuclei showing condensation of the chromatin
as dense green areas. Late apoptotic cells and necrotic cells were
stained with AO and PI. Comparatively, PI produces the highest
intensity emission. Thus, late apoptotic cells exhibited orange
nuclei showing condensation of the chromatin, whilst necrotic cells
exhibited orange nuclei with intact structures.
Annexin V and PI staining for cell death
analysis
For assaying the influence of damnacanthal on the
induction of cellular apoptosis and cell death, MCF-7 cells in
exponential growth were treated with IC50 concentrations
of damnacanthal for 72 h. Assessment of apoptotic and dead cells
was performed by Annexin V and PI staining. Following incubation,
cells were washed twice with PBS and the cell pellet was
resuspended with Annexin V binding buffer followed by incubation
with Annexin V conjugated to fluorescein isothiocyanate (FITC) and
PI staining solution (BD Pharmingen, San Diego, CA, USA) for 5 min
at room temperature in the dark. Cells were then analyzed by flow
cytometry within 1 h. Flow cytometry was performed on FACScan (BD
Biosciences, San Jose, CA, USA).
Cell cycle analysis
For cell cycle analysis, 5 ml MCF-7 cells
(5×104 cells/ml) were cultured in 25-cm2
flasks, with or without damnacanthal at IC50
concentration for 72 h. Cells were then harvested, washed in PBS,
centrifuged and resuspended in 1 ml sodium citrate (0.1%)
containing 0.05 mg PI and 50 μg RNase for 30 min at room
temperature in the dark. Flow cytometry was performed on FACScan
(BD Biosciences), with collection and analysis of the results
performed using CellQuest software (BD Biosciences).
Total RNA preparation and reverse
transcription-polymerase chain reaction (RT-PCR) analysis
The apoptosis-related genes were analyzed following
the treatment of the MCF-7 cells with or without damnacanthal at
IC50 concentration for 72 h. The cells were trypsinized
and washed twice with PBS. Total RNA was prepared using a Qiagen
RNA extraction kit (Qiagen, Hilden, Germany). The RNA concentration
was determined by reading the absorbance at 260 and 280 nm with a
UV spectrophotometer (DU730; Beckman Coulter, Petaling Jaya,
Selangor, Malaysia). The total RNA was transcribed to cDNA using
the GenomeLab™ GeXP start kit (Beckman Coulter, Miami, FL, USA) for
RT-PCR, according to the manufacturer’s instructions. The following
primers were designed from the known sequences:
5′-CCCTTTTGCTTCAGGGTTTC-3′ and 5′-ACAAAGTAGAAAAGGGCGACAA-3′ for
Bax; 5′-TGTGGACCTGTCACTGTCTTG-3′ and 5′-TAGGGCTTCCTCTTGGAGAA-3′ for
p21Cip1; 5′-CAGACCGGTCCTCGTTTGTA-3′ and 5′-ACCTCG
GCATCTTTGTCTGTT-3′ for caspase-7; and 5′-AAGGTG AAGGTCGGAGTCAA-3′
and 5′-AGATCTCGCTCCTGG AAGATG-3′ for GAPDH. The amplification
profile was as follows: Denaturing at 94°C for 30 sec; annealing at
55°C for 30 sec; and extension at 68°C for 1 min. The cDNA was
amplified using MJ Research PTC-225 analyzer (MJ Research Inc., St.
Bruno, QC, Canada) for 35 cycles, followed by a step of 10 min at
72°C to extend the partially amplified products. These cycling
conditions were established empirically to provide a linear
increase in product intensity proportional to the amount of
template. The PCR products with fluorescently labeled fragments
were separated by capillary gel electrophoresis (One Capillary
array, BD Biosciences, Franklin Lake, NJ, USA), at 6.0 kV and 50°C
for 35 min, according to their product size, and the results were
analyzed using GenomeLab GeXP system software (Beckman Coulter).
GADPH was selected as the reference gene for normalizing all
results of the targeted genes.
Analysis of Bcl2, p53, estrogen receptor
(ER)-α and X-linked inhibitor of apoptosis protein (XIAP)
expression by flow cytometry
Cells were seeded at a density of 5×105
cells/well in six-well culture plates. Following treatment with
IC50 concentrations of damnacanthal for 72 h, cells were
harvested and fixed by BD Cytofix/Cytoperm™
Fixation/Permeabilization (BD Biosciences). The cells were stained
with fluorochrome-conjugated monoclonal anti-mouse p53 and Bcl-2
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), XIAP (BD
Biosciences) and ER-α (Abcam, Cambridge, MA, USA). The cells were
washed twice with PBS to remove non-specific binding stained with
secondary fluorochrome-conjugated monoclonal antibody mouse
anti-p53, -Bcl2, -XIAP and -ER-α (Abcam). The cells were washed
twice and analyzed by BD FACSCalibur multicolor flow cytometer (BD
Biosciences).
Statistical analysis
Data are presented as the mean ± standard deviation.
P-values were determined by analysis of variance followed by
Student-Newman-Keuls test for multiple comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of damnacanthal on the viability
of MCF-7 cells
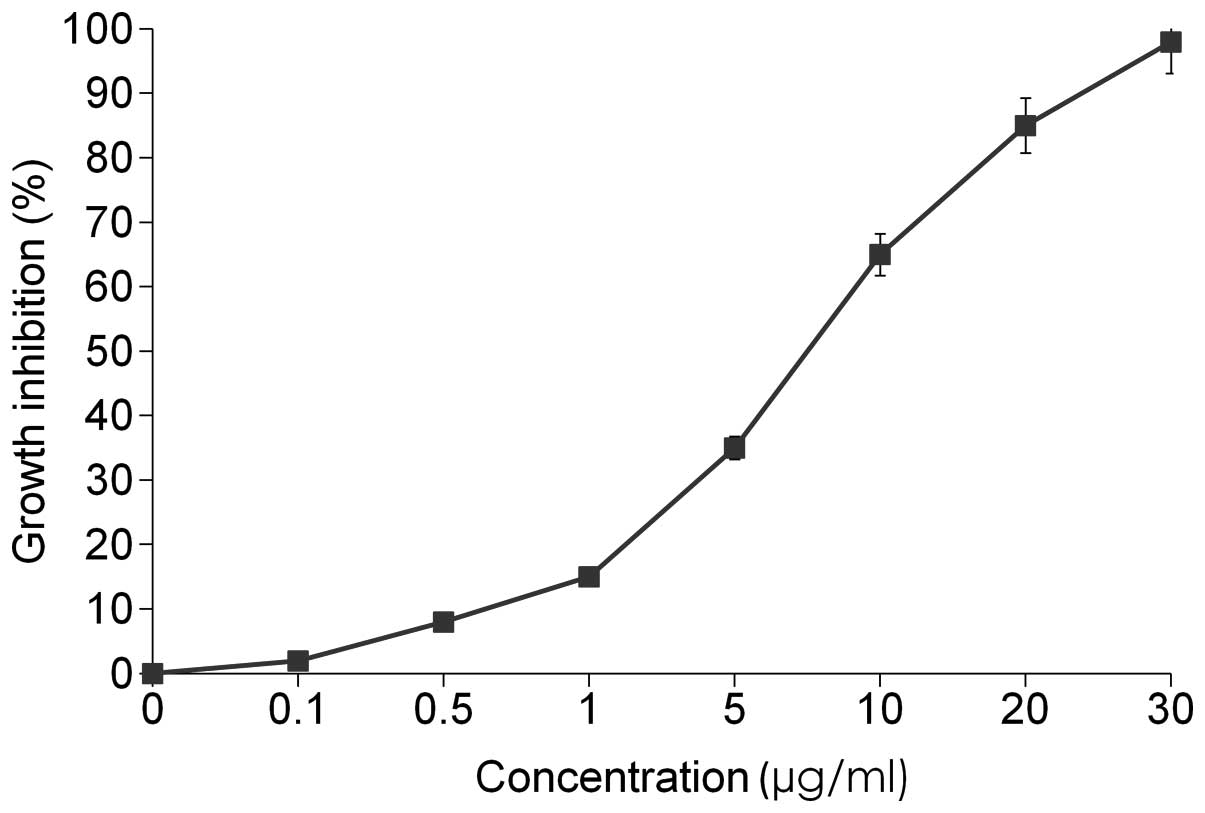
To examine the effects of damnacanthal on MCF-7 cell
viability, MTT assay was performed. MCF-7 cells were plated onto
96-well plates and treated with various concentrations of
damnacanthal (0–30 μg/ml) for 72 h. As shown in Fig. 2, damnacanthal dose-dependently
(P<0.05) inhibited cell viability. These inhibitory effects were
observed following incubation with 8.2 μg/ml damnacanthal, reducing
cell growth by 50% (IC50).
Damnacanthal induced apoptotic cell death
in MCF-7
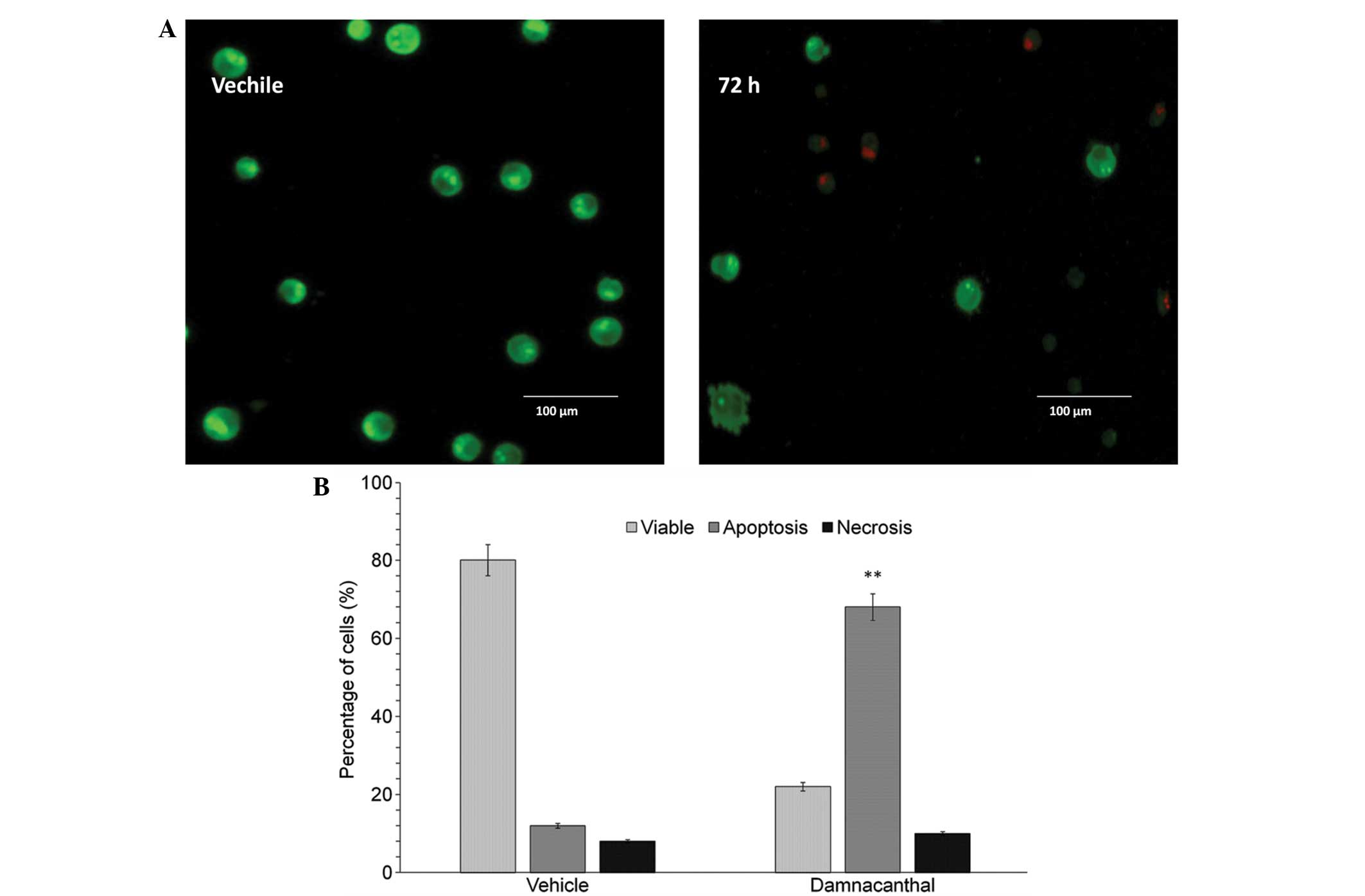
AO and PI dyes were used to differentiate viable,
apoptotic and necrotic cells under fluorescence microscope.
Fig. 3A shows the intact viable
cells and apoptotic and necrotic cells following the treatment of
MCF-7 cells with damnacanthal at IC50 concentration for
72 h. The apoptotic event of damnacanthal-treated cells was
increased significantly: ≤4-fold higher than the vehicle-treated
cells (Fig. 3B). A fraction of
necrotic cells were also detected in the treatment group.
Induction of apoptosis by damnacanthal in
MCF-7 cells
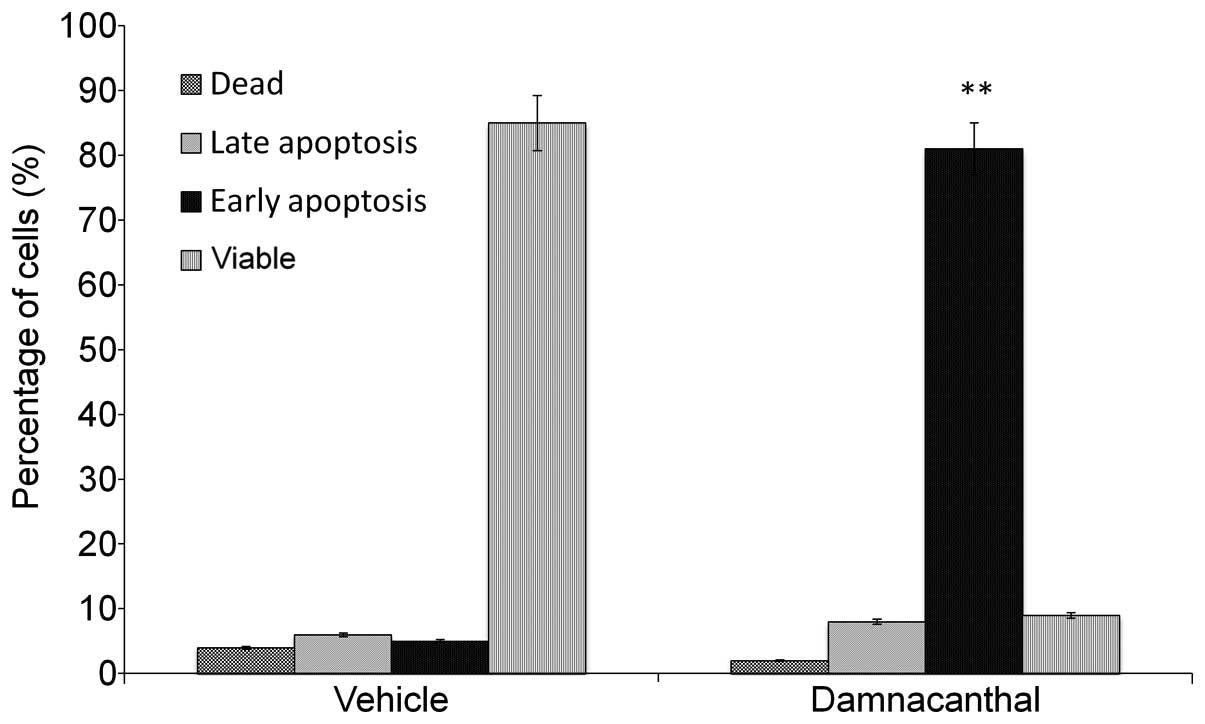
To determine whether the loss of cell viability
induced by damnacanthal was associated with apoptosis, Annexin
V-FITC/PI binding assay was performed. The assay evaluates
phosphatidylserine turnover from the inner to the outer lipid layer
of the plasma membrane, an event typically associated with
apoptosis. Flow cytometric analysis revealed that the percentage of
apoptotic cells with Annexin V-positive but PI-negative cells
increased gradually with concentration in damnacanthal-treated
cells. As shown in Fig. 4,
following 72 h of treatment with damnacanthal at IC50
concentrations, the numbers of apoptotic MCF-7 cells, as revealed
by Annexin V binding, increased in a dose-dependent manner,
indicating a proapoptotic activity of damnacanthal. The proportion
of MCF-7 cells in early apoptosis was 80.6% after 72 h (P<0.05,
vs. vehicle-treated cells at the same time). For cells that were in
the late apoptosis, the proportion was 8.1% at 72 h, while the
proportion of the cells in necrosis was 2% at 72 h (P<0.05, vs.
vehicle-treated cells at the same time). For the vehicle-treated
MCF-7 cells, the percentage of cells that underwent necrosis was
<4% at 72 h incubation time (Fig.
4).
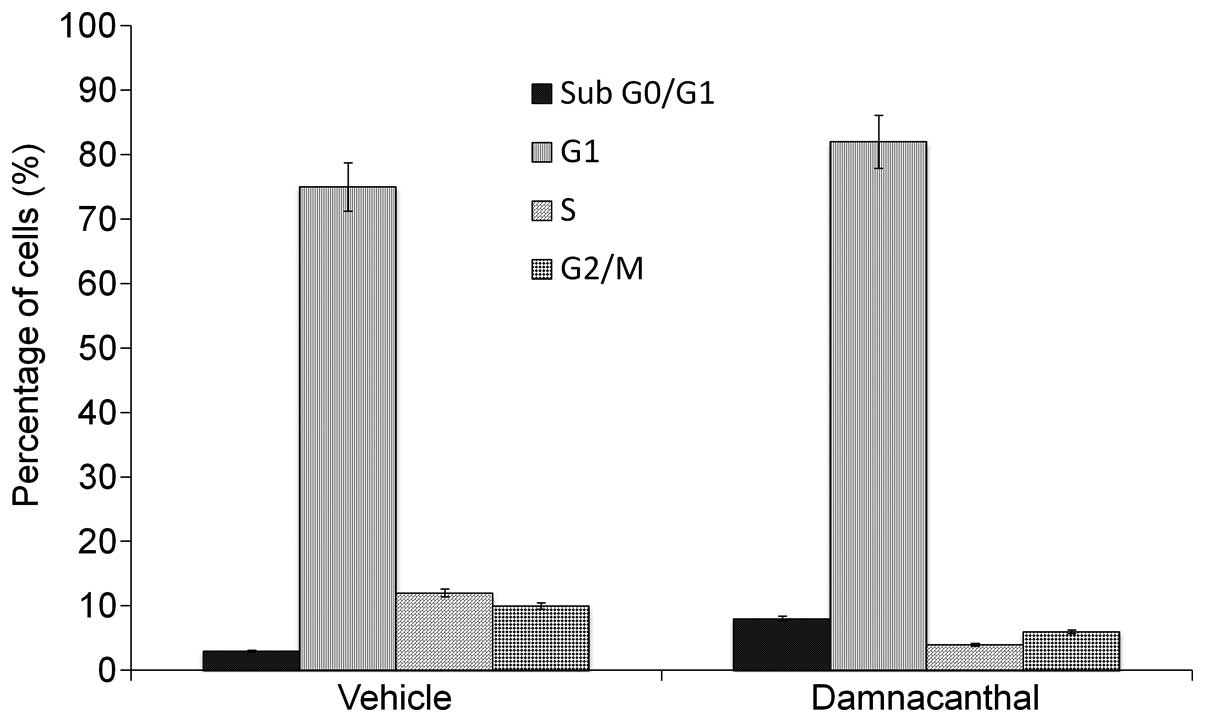
Damnacanthal induces G1 cell cycle
arrest
Following the treatment of MCF-7 cells with 8.2
μg/ml damnacanthal for 72 h, the cell population in G1 phase
increased to 80% which was accompanied with a decrease in the S
(5%) and G2 (8%) phases (Fig. 5).
These results clearly indicated that damnacanthal induces post
G1-arrest and apoptosis among the treated cells.
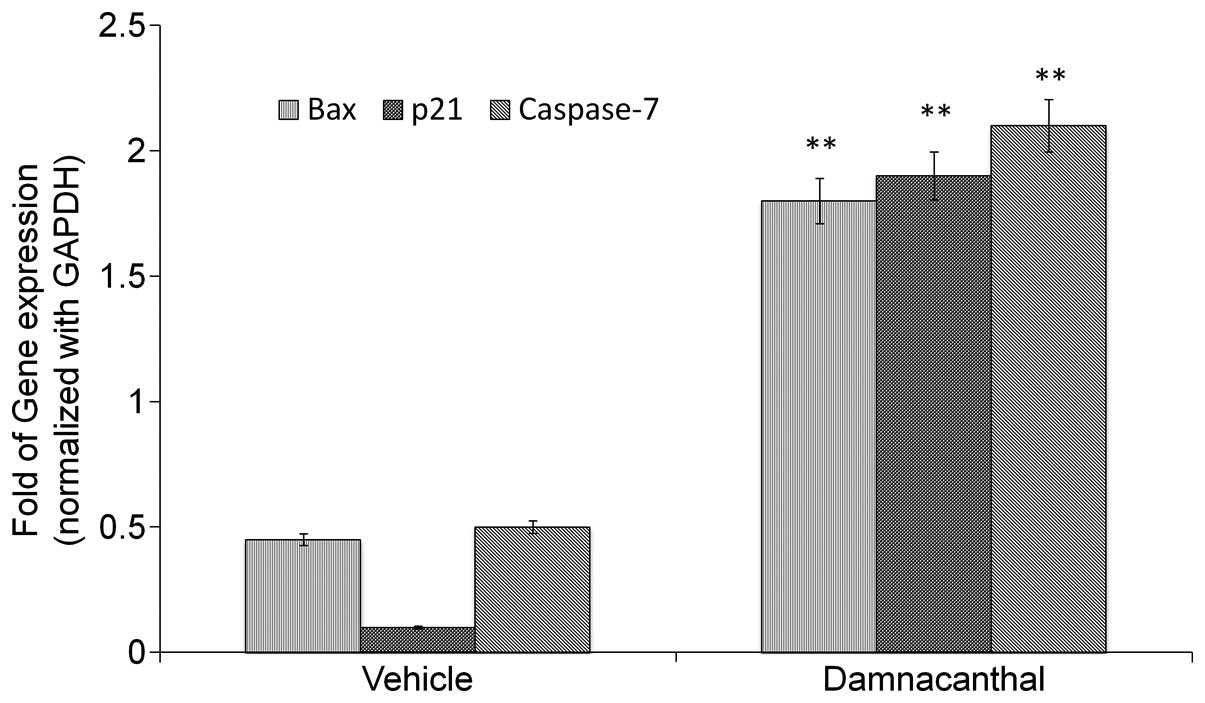
Activation of apoptotic genes by
damnacanthal
The expression of proapoptotic genes was analyzed in
MCF-7 cells treated with or without damnacanthal for 72 h. The
damnacanthal-treated cells showed significant changes in the
expression levels of BAX, p21 and caspase-7 genes (Fig. 6). For the p21 gene, all treatments
exhibited a significant increase (P<0.05) compared with the
control.
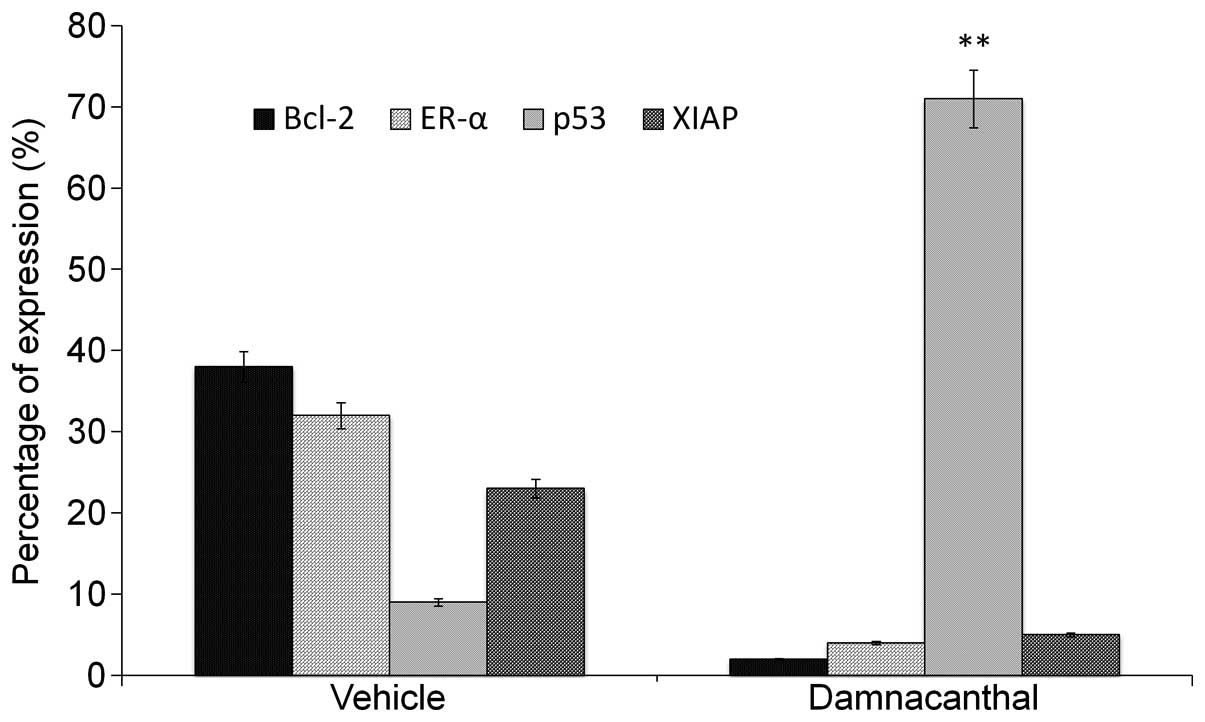
Involvement of apoptotic proteins in
damnacanthal-induced apoptosis
Expression levels of proteins that are involved in
pro- and anti-apoptosis, such as Bcl-2, p53, ER-α and XIAP, were
evaluated using a multicolor flow cytometer. Fig. 7 shows the percentage changes on the
expression level of protein following the treatment of MCF-7 cells
with damnacanthal at 8.2 μg/ml for 72 h. The percentage expression
level of p53 protein increased significantly (P<0.05) in
damnacanthal-treated MCF-7 cells compared with the control.
Simultaneously, percentage expression levels of Bcl-2, XIAP and
ER-α protein were suppressed significantly (P<0.05).
Discussion
Cancer has become an increasing public health issue
for its high rates of morbidity and mortality. In the current
study, the anticancer activity of damnacanthal, an anthraquinone
which is extracted from the noni plant, was investigated.
Considering that little is known concerning the anticancer
activities and related mechanisms of damnacanthal, the current
study performed further investigations to elucidate the antitumor
activities of damnacanthal in the human breast cancer MCF-7 cell
line and the possible mechanisms involved. The experimental results
showed that damnacanthal exhibits potent cytotoxicity to MCF-7
cells, with an IC50 of 8.2 μg/ml (Fig. 2). Apoptosis assay showed that
apoptotic cells induced by damnacanthal exhibited cellular
alterations, determined by counting chromatin condensation and
nuclear fragmentation by AO/PI (Fig. 3A
and C). Annexin V/PI double-staining assay further confirmed
the results of the AO/PI staining by showing the important membrane
alterations associated with apoptosis in MCF-7 cells and the
percentage of apoptosis increase in damnacanthal treatment
(Fig. 4). The cell cycle results
demonstrated that damnacanthal induced G1 arrest and apoptosis
among the damnacanthal-treated cells (Fig. 5). All these results showed that
damnacanthal increases antitumorigenic activity by increasing the
expression of p53, followed by p21. It has been previously reported
that damnacanthal induces apoptosis in a number of cells, including
HL-60, MOLT-4, CEM-SS, HT-29, Hela, 3T3 and peripheral blood
mononucleated cells (10,15,16).
Proposed mechanisms for the proapoptotic effect include activation
of caspases, induction of cytochrome c release, regulation
of protein kinase C isoform expression, inhibition of NF-κB and
suppression of activator protein 1 (17–19).
The results of the current study were consistent with our previous
study, as damnacanthal induced apoptosis in HL-60 and Wehi-3B cells
(10). Further investigations were
performed to highlight the apoptotic pathways involved in the
apoptosis induced by damnacanthal in MCF-7 cells.
Previous studies have revealed that caspases are
critical in executing apoptosis (20). In order to gain further insight into
the mechanism of the signaling cascade, the present study examined
the molecular sequence of events in damnacanthal-induced apoptosis.
Apoptosis may occur via two fundamental pathways: i) death receptor
or extrinsic pathway; and ii) mitochondrial or intrinsic pathway.
The present study demonstrated the considerable role of the
mitochondrial apoptotic pathways in apoptosis induced by
damnacanthal in MCF-7 cells. Damnacanthal-mediated activation of
Bax, p21 and caspase-7 was identified in MCF-7 cells. The
activation of p21 and caspase genes stimulates p53 phosphorylation
(21). Although multiple pathways
contribute to the modulation of p53 (22), the current study investigated the
expression of p21 as one of the upstream molecules of p53. The
results demonstrated that p21-p53 signaling is one of the key
pathways in mediating damnacanthal-induced apoptosis. In addition,
the role of p21 in the transcription of the p53-regulated Bax gene
is likely to involve p53 phosphorylation (23). The increased damnacanthal-dependent
p53 protein levels are consistent with the damnacanthal-dependent
transcriptional induction of Bax. Extensive analyses of
damnacanthal-dependent modifications of p53 are in progress to link
p21 activity with p53 function in damnacanthal-mediated apoptosis.
Although modulation of p21 and p53 signaling is common, the current
study established connections between well-known proapoptotic
molecules in the damnacanthal-induced apoptosis.
In conclusion, damnacanthal, a bioactive compound
from noni roots, enhanced the expression of p21 and caspase-7.
Overexpression of p21 directly activates transcription and
expression of p53 and, subsequently, increases apoptosis in human
breast cancer MCF-7 cells. These results are likely to highlight
the potential benefits of damnacanthal for further preclinical or
clinical practice and damnacanthal may be a useful cancer
prevention/therapeutic agent in human breast carcinoma.
Acknowledgements
The authors would like to thank the Ministry of
Higher Education (Putrajaya, Malaysia) for financial assistance
through the Fundamental Grant Research Scheme (no.
03-10-10-964FR).
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar
|
|
2
|
Jovanovic J, Rønneberg JA, Tost J and
Kristensen V: The epigenetics of breast cancer. Mol Oncol.
4:242–254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wickerham DL: Breast cancer
chemoprevention: progress and controversy. Surg Oncol Clin NA.
19:463–473. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hatcher H, Planalp R, Cho J, Torti FM and
Torti SV: Curcumin: from ancient medicine to current clinical
trials. Cell Mol Life Sci. 65:1631–1652. 2008. View Article : Google Scholar
|
|
7
|
Kawamori T, Lubet R, Steele VE, Kelloff
GJ, Kaskey RB, Rao CV and Reddy BS: Chemopreventive effect of
curcumin, a naturally occurring anti-inflammatory agent, during the
promotion/progression stages of colon cancer. Cancer Res.
59:597–601. 1999.
|
|
8
|
Wang MY, West BJ, Jensen CJ, Nowicki D, Su
C, Palu AK and Anderson G: Morinda citrifolia (Noni): a
literature review and recent advances in Noni research. Acta
Pharmacol Sin. 23:1127–1141. 2002.
|
|
9
|
Kamiya K, Tanaka Y, Endang H, Umar M and
Satake T: New anthraquinone and iridoid from the fruits of
Morinda citrifolia. Chem Pharmaceut Bull (Tokyo).
53:1597–1599. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Alitheen NB, Mashitoh AR, Yeap SK,
Shuhaimi M, Manaf AA and Nordin L: Cytotoxic effect of
damnacanthal, nordamnacanthal, zerumbone and betulinic acid
isolated from Malaysian plant sources. Int Food Res J. 17:711–719.
2010.
|
|
11
|
Ali AM, Ismail NH, Mackeen MM, Yazan LS,
Mohamed SM, Ho AS and Lajis NH: Antiviral, cytotoxic and
antimicrobial activities of antharaquinones isolated from the roots
of morinda elliptica. Pharm Biol. 38:298–301. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Di Salvo J, Nelson SR and Kaplan N:
Protein tyrosine phosphorylation in smooth muscle: a potential
coupling mechanism between receptor activation and intracellular
calcium. Pro Soc Exp Biol Med. 214:285–301. 1997.
|
|
13
|
Hiwasa T, Arase Y, Chen Z, Kita K, Umezawa
K, Ito H and Suzuki N: Stimulation of ultraviolet-induced apoptosis
of human fibroblast UVr-1 cells by tyrosine kinase inhibitors. FEBS
Lett. 444:173–176. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tosa H, Iinuma M, Asai F, et al:
Anthraquinones from Neonauclea calycina and their inhibitory
activity against topoisomerase II. Biol Pharm Bull. 21:641–642.
1998. View Article : Google Scholar
|
|
15
|
Norsyafini I, Latifah SY and Nordin HL:
Nordamnacanthal induced apoptosis and mitotic G2/M arrest with down
regulation of bcl-2 in the human breast cancer cell line (MCF-7).
Med Health Sci J. 2:27–38. 2010. View Article : Google Scholar
|
|
16
|
Latifah SY, Norsyafini I and Nordin HL:
BCL-2 was Down regulated in G2/M-Arrest Breast Cancer Cells
MCF-7-Treated with Nordamnacanthal. J Pharm Sci Resl. 2:197–207.
2010.
|
|
17
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Strasser A, O’Connor L and Dixit VM:
Apoptosis signaling. Ann Rev Biochem. 69:217–245. 2000. View Article : Google Scholar
|
|
19
|
Suzuki A and Shiraki K: Tumor cell ‘dead
or alive’: caspase and surviving regulate cell death, cell cycle
and cell survival. Histol Histopathol. 16:583–593. 2001.
|
|
20
|
Green DR and Reed JC: Mitochondria and
apoptosis. Science. 281:1309–1312. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mirzayans R, Andrais B, Scott A and Murray
D: New insights into p53 signaling and cancer cell response to DNA
damage: implications for cancer therapy. J Biomed Biotechnol.
2012:1703252012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Macleod KF, Sherry N, Hannon G, et al:
p53-dependent and independent expression of p21 during cell growth,
differentiation, and DNA damage. Genes Dev. 9:935–944. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
el-Deiry WS, Tokino T, Velculescu VE, et
al: WAF1, a potential mediator of p53 tumor suppression. Cell.
75:817–825. 1993. View Article : Google Scholar : PubMed/NCBI
|