Introduction
Lung cancer has become one of the leading causes of
cancer-related mortality worldwide and non-small cell lung cancer
(NSCLC) represents 80% of lung cancers (1,2). The
epidermal growth factor (EGFR) is a significant therapeutic target
in NSCLC (3). Individuals with
somatic mutations of the kinase domain of EGFR often respond to
tyrosine kinase inhibitor (TKI) therapy, however, usually exhibit
progressive disease following 6–8 months of therapy (4,5).
Radiotherapy is extremely important for patients with NSCLC who are
not eligible for surgery and patients that have experienced
chemotherapy or TKI therapy failure. However, NSCLC cells are
generally less sensitive to radiotherapy compared with SCLC cells,
which results in radiotherapy failure (6). Although radiotherapy with increased
irradiation dosage may delay tumor development, it leads to serious
side-effects, including irradiation pneumonitis and a repressed
hemopoietic system. Therefore, it is significant to use
radiosensitizers to raise the therapeutic effect at a normal
irradiation dosage. In the last few decades, increased attention
has been focused on identifying biologically active cancer
therapeutic agents derived from natural resources (7).
Rhizoma paridis is the root of Paris
polyphylla Smith var. chinensis (Franch) Hara and Paris
polyphylla Smith var. yunnanensis (Franch) Hand-Mazz.
Preclinical studies have shown that Paris saponins (PS) have
emerged as promising anticancer agents (8–12), and
PSI exerts a wide range of pharmacological activities, including
cytotoxic activity against certain malignancies, such as NSCLC
(13–17). Therefore, PSI has been approved for
cancer therapy due to its potential involvement in the suppression
of tumor growth. However, PSI inhibition of signaling pathways and
its radiosensitization in NSCLC-TKI resistance has not been
identified. The present study focused on examining the
radiosensitization effects of PSI on NSCLCs with acquired gefitinib
resistance in vitro and to further verify the possible
mechanisms.
Materials and methods
Drugs and reagents
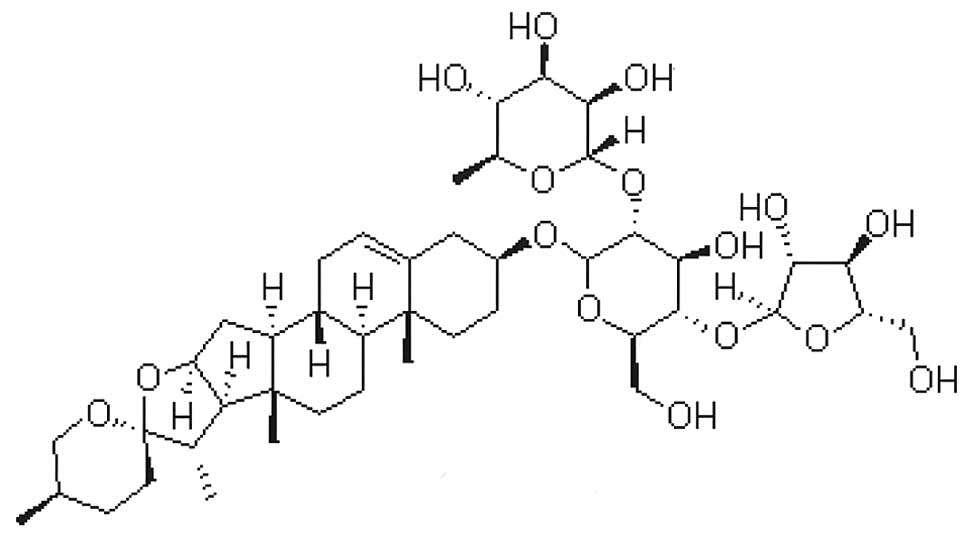
PSI (C44H70O16) was
obtained from Zhejiang Meidikang Ltd. (Zhejiang, China) and the
structure of the compound is shown in Fig. 1. PSI was prepared as a 20-mmol/l
stock in dimethyl sulfoxide (DMSO) and stored at −20°C. PSI was
diluted with cell culture medium to concentrations of 0.5, 1.0,
2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0 and 9.0 μg/ml, with a final DMSO
concentration of 0.25% (v/v). Dulbecco’s modified Eagle’s medium
(DMEM) was purchased from Gibco-BRL (Carlsbad, CA, USA), the Cycle
Test™ Plus DNA reagent and Annexin V-FITC &
Propidium Iodide (PI) Apoptosis Detection kits were obtained from
Becton Dickinson and Co., (Franklin Lakes, NJ, USA), Hoechst 33258
was purchased from BYT Co., (Nanjing, China), mouse and rabbit
antibodies against caspase-3, Bcl-2-like protein 4 (Bax), B-cell
lymphoma 2 (Bcl-2) and cyclin-dependent kinase inhibitor 1
(P21waf1/cip1) were obtained from Cell Signaling
Technology (Danvers, MA, USA) and β-actin from Santa Cruz
Biotechnology, Inc., (Santa Cruz, CA, USA). This study was approved
by the Ethics Committee of Zhejiang Hospital and was performed
according to the Declaration of Helsinki. Written informed consent
was obtained from the family of the patients.
Cell culture
PC-9-ZD (18), an
NSCLC cell line resistant to gefitinib following long-term exposure
to the drug, was obtained from the Laboratory of Biochemistry and
Molecular Biology, Tongji University (Shanghai, China). The PC-9-ZD
cells were grown in DMEM supplemented with 10% fetal bovine serum,
100 μg/ml penicillin and 100 μg/ml streptomycin at 37°C in a 5%
CO2 humidified atmosphere.
Cell proliferation assay
The cell proliferation assays were performed using
the MTT method, according to the manufacturer’s instructions. The
cells were seeded in 96-well plates (Costar; Corning Life Sciences,
Cambridge, MA, USA) with 5,000 cells/well. Subsequent to an
overnight incubation, triplicate wells were treated with various
concentrations of PSI for 24 h. Following this, 20-μl MTT solutions
(5 mg/ml in phosphate-buffered saline; PBS) were added to each well
and incubated for 4 h at 37°C. The MTT formazan was dissolved in
150 μl DMSO and the absorbance was measured with a microplate
reader (Multiskan MK3; Thermo Labsystem, Waltham, MA, USA) at a
wavelength of 570 nm. The drug-cell inhibition curve used the drug
concentration as the abscissa axis and the inhibition ratio of the
drug as the vertical axis. The 50% growth inhibition
(IC50) was subsequently calculated according to the
curve.
Determination of cell
radiosensitivity
The growing cells were exposed to PSI with a
concentration of 20% of the IC50 for 3 h, and irradiated
at 0, 1, 2, 4, 6, 8 or 10 Gy with a 6-MV X-ray. After 24 h, the
cells were trypsinized, counted and seeded at various dilutions
according to the irradiation dose and cultured for 14 days; the
colonies were fixed, stained with crystal violet and counted. Only
the colonies containing 50 cells were scored and the experiments
were performed in triplicate. The cell-survival curve used the
irradiation dosage as the abscissa axis and the survival fraction
(SF) as the vertical axis. The average lethal dosage of cells
(D0) and the quasi-field dosage (Dq), which indicates
the repair ability of cells to sublethal injury, and extrapolation
number (N) values were calculated according to the curve. The
sensitization enhancement ratio (SER) was calculated according to
the following equations: SF =
1-(1-exp[−D/D0])N; Dq =
InN/(1/D0); and SER = control group D0
value/treatment group D0 value.
Cell-cycle distribution
The experimental groups were the control, radiation
and PSI + radiation groups. The radiation group received a 2-Gy
treatment and the PSI + radiation group received a 2-Gy and a PSI
treatment using a concentration that was 20% of the
IC50. Cells were harvested at 12, 24 and 48 h and were
fixed with 70% ethanol and stored overnight at −20°C. The cells
were centrifuged using a Heraeus Labofuge 400 centrifuge (Thermo
Fisher Scientific, Waltham, MA, USA) at 300 × g and washed twice
with PBS. They were labeled with 50 mg/ml PI and protected from
light for 30 min prior to analyses by flow cytometry that were
conducted with a multi-cycle system software package (CellQuest
version 3.1; Beckman Coulter, Inc., Brea, CA, USA). Experiments
were performed in triplicate.
Apoptosis measurement
Apoptosis was measured by PI/Annexin V double and
Hoechst staining. The experimental groups were the control,
radiation and the PSI + radiation groups. The radiation group
received a 2-Gy treatment and the PSI + radiation group received a
2-Gy and PSI treatment using a concentration that was 20% of the
IC50. The cells were harvested at 24 h following
treatment and stained with PI and Annexin V. The apoptotic fraction
was detected by flow cytometry (BD FACSCalibur; BD Biosciences,
Franklin Lakes, NJ, US). The cells were washed in PBS, stained with
Hoechst 33528 (5 μg/ml in PBS) for 15 min at room temperature and
observed under a fluorescence microscope (Olympus BX-60, Olympus
Optical Co., Ltd., Tokyo, Japan) equipped with 356-nm excitation
and 492-nm emission band-pass filters.
Western blot analysis
The cells were exposed to a PSI treatment of 20% of
the IC50 for 3 h and subsequently irradiated at a dose
of 2 Gy and incubated for 24 h. Total cell lysates were separated
by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred to a polyvinylidene fluoride membrane (Seebio Biotech,
Inc., Shanghai, China). The membranes were incubated overnight with
primary antibodies [1:1,000; caspase-3 mouse monoclonal antibody
(mAb), Bax rabbit mAb, Bcl-2 rabbit mAb and p21 Waf1/Clip1 rabbit
mAb] at 4°C with gentle agitation (Wave-SI slim shaker; TAITEC
Corporation, Koshigaya, Japan). The membranes were incubated for 2
h with a horseradish peroxidase-labeled secondary antibodies
(1:2,000; affinity purified goat antimouse IgG and goat antirabbit
IgG) at room temperature. All membranes were detected using the ECL
system (Santa Cruz Biotechnology Inc.).
Statistical analysis
The t-test (mean comparison in two samples) and
single-factor variance analysis (mean comparison in multiple
samples) were assessed using SPSS 17.0 (SPSS, Inc., Chicago, IL,
USA) and experimental data are indicated by the mean ± SD P<0.05
was considered to indicate a statistically significant
difference.
Results
PSI inhibits the proliferation of PC-9-ZD
cells
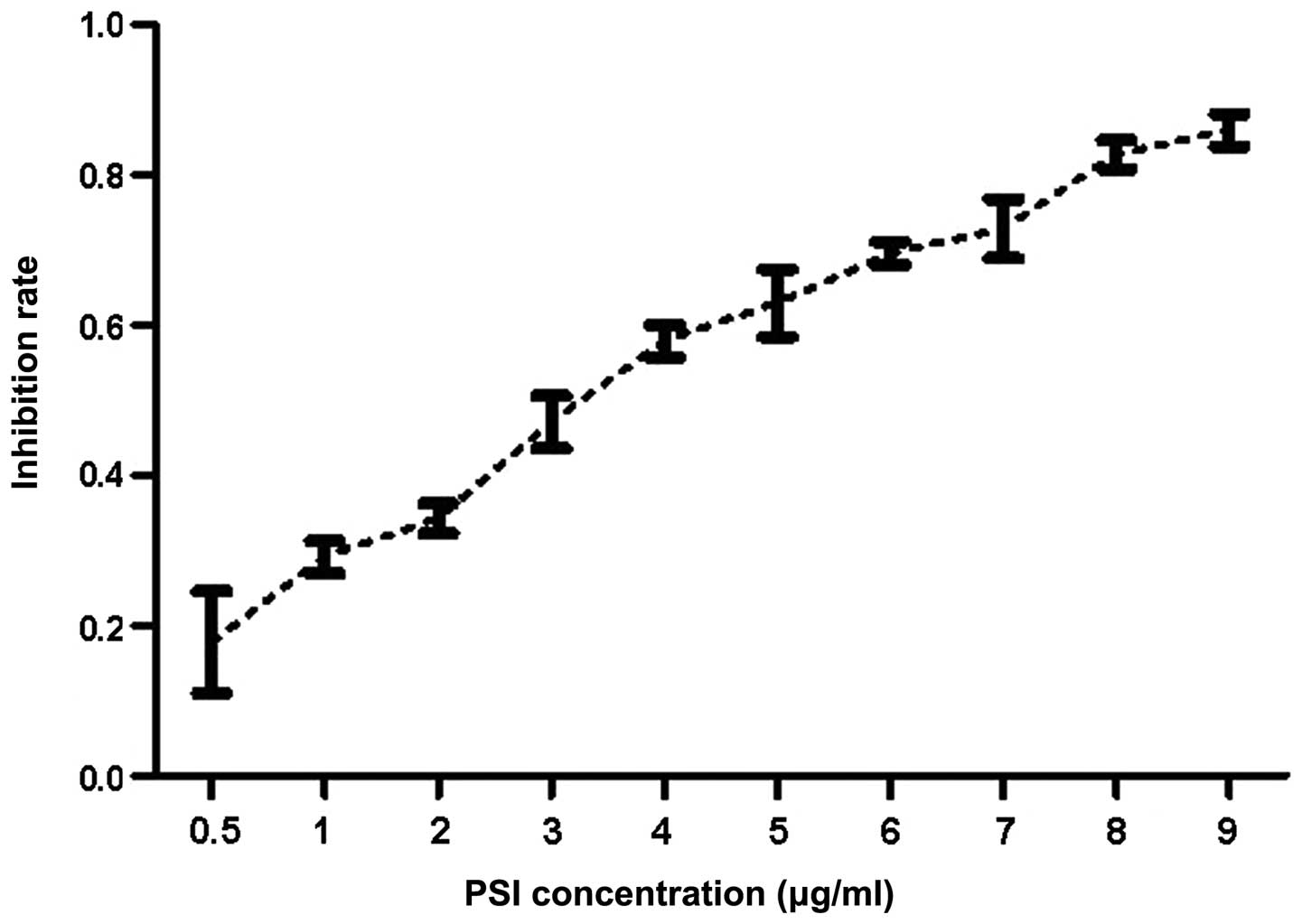
The MTT assay (Fig.
2) showed that PSI treatment inhibited cell proliferation in a
dose-dependent manner. The concentration required to achieve
IC50 was estimated to be 2.5132 μg/ml at 24 h.
PSI enhances the radiosensitivity of
PC-9-ZD cells
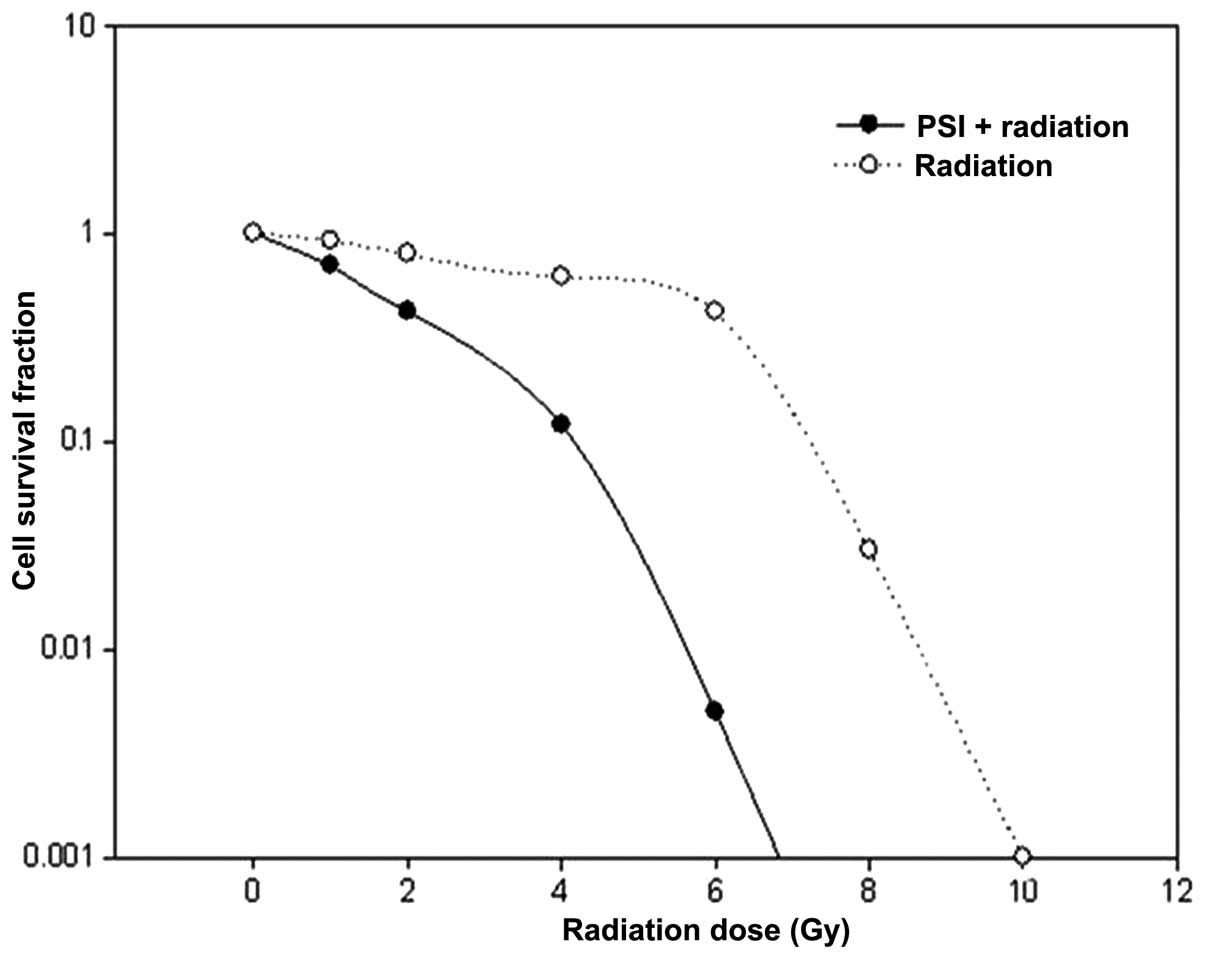
0.5 μg/ml PSI and 20% of the IC50 served
as the experiment concentration. A multi-target click mathematical
model (Fig. 3) simulated the cell
SF curve, through which an associated equation and radioactivity
parameters, D0 and Dq (Table
I) were obtained. The results show a declined SF2, a decreased
Dq and the shoulder of the survival curve is decreased, with an SER
value of 1.77, based on D0.
 | Table IParameters of multi-target click
mathematical model. |
Table I
Parameters of multi-target click
mathematical model.
| Group | D0 | Dq | N | SF2 (%) | SER
D0 |
|---|
| Radiation | 0.5105 | 0.1636 | 1.3777 | 37.23 | - |
| PSI + radiation | 0.2887 | 0.0598 | 1.2302 | 17.43 | 1.77 |
PSI induces the G2/M arrest of irradiated
PC-9-ZD cells
To identify whether the radiosensitivity of PSI was
due to cell cycle arrest, the influence of PSI treatment (0.5
μg/ml; 20% of IC50) on cell cycle distribution was
observed (Table II). The result
showed that irradiation alone induced G2/M arrest in a
time-dependent manner with an increased cell density at the G2/M
phase from 16.56 to 24.07% compared with the control group
(P<0.05). However, PSI altered the cycle distribution of
irradiated cells significantly, leading to cell cycle arrest at the
G2/M phase in a time-dependent manner with an increased cell
density at the G2/M phase from 27.63 to 39.30% compared with the
radiation group (P<0.01).
| Table IIEffect of PSI on the G2/M phase of
irradiated PC-9-ZD cells. |
Table II
Effect of PSI on the G2/M phase of
irradiated PC-9-ZD cells.
| Cell density (%, mean
± standard deviation) |
|---|
|
|
|---|
| Group | 12 h | 24 h | 48 h |
|---|
| Control | 7.18±1.44 | 9.45±2.51 | 10.89±2.72 |
| Radiationa | 16.56±1.35 | 21.67±2.25 | 24.07±2.47 |
| PSI +
radiationb,c | 27.63±2.16 | 35.88±2.14 | 39.30±2.53 |
PSI induces apoptosis of irradiated
PC-9-ZD cells
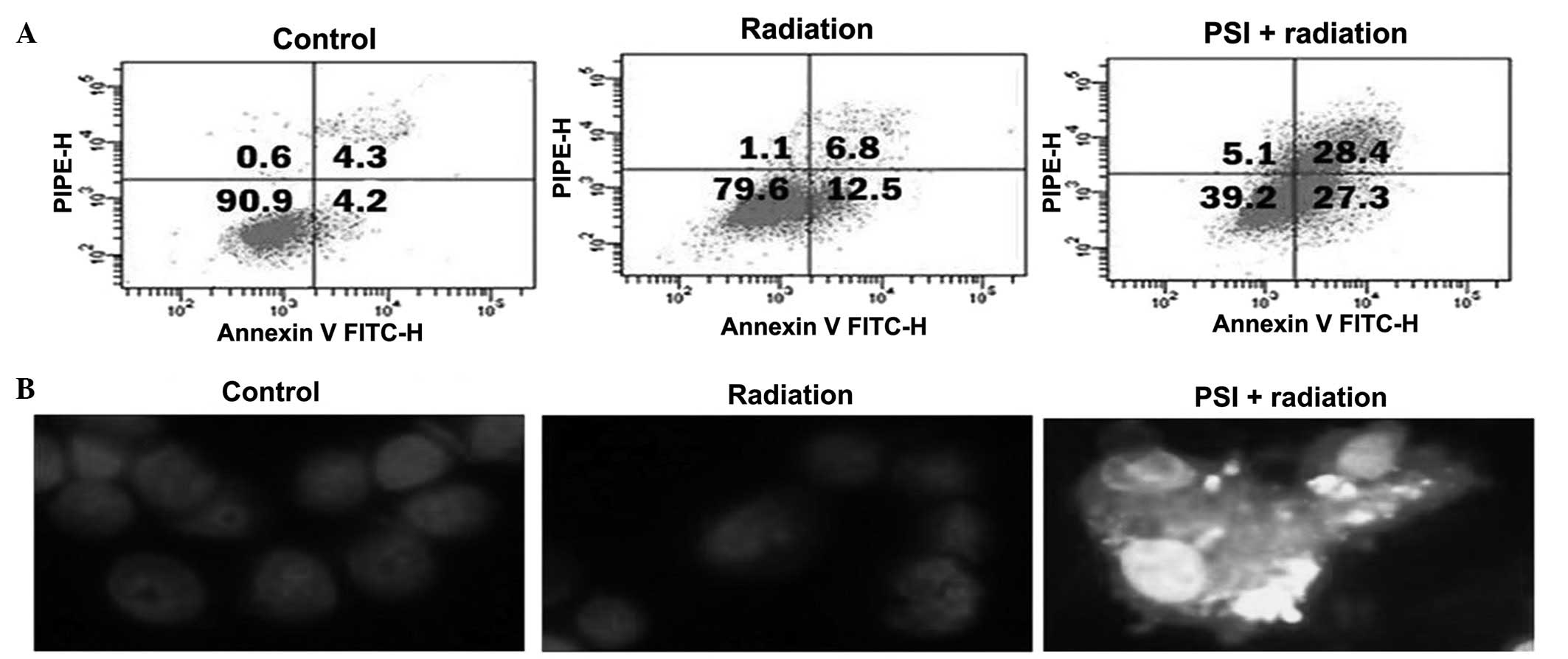
In order to investigate the radiosensitivity
mechanism of PSI, the influence of PSI (0.5 μg/ml; 20% of
IC50) on cell apoptosis was observed by Annexin V/PI
double and Hoechst staining assays. Irradiation increased apoptosis
at 24 h, however, the PSI combination treatment further increased
the apoptosis ratio up to the higher level (P<0.01; Fig. 4A). Fig.
4B demonstrates that the control and irradiated cells were
morphologically normal, and the nuclei were regularly shaped and
evenly stained. However, typical morphological changes of
apoptosis, including nuclear shrinkage, DNA condensation and
chromatin fragmentation were identified in the PSI + radiation
group. This indicates that PSI treatment can further increase
apoptosis that is induced by radioactive rays.
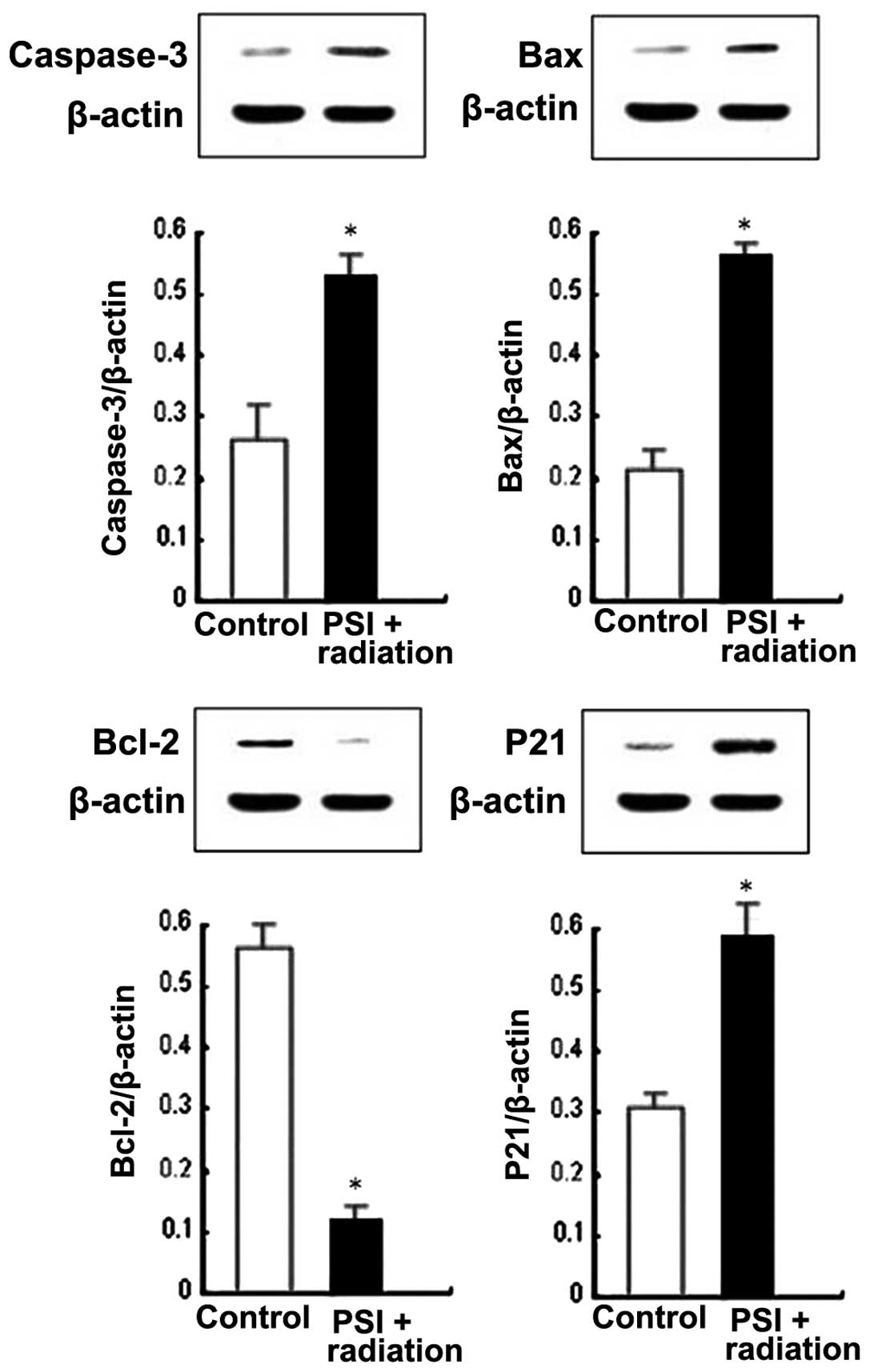
PSI upregulates P21waf1/cip1,
caspase-3 and Bax, and downregulates Bcl-2 protein expression of
irradiated PC-9-ZD cells
In order to confirm which modulating molecules were
involved in the PSI treatment on cell cycle arrest and apoptosis of
irradiated PC-9-ZD cells, P21waf1/cip1, the most
significant regulator in the cell cycle checkpoint, and caspase-3,
Bax and Bcl-2, which are significant apoptosis regulators, were
investigated. The results showed that PSI significantly increased
the expression of P21waf1/cip1, caspase-3 and Bax in
irradiated PC-9-ZD cells. Notably, PSI markedly decreased Bcl-2
expression in the irradiated cells (Fig. 5). This incidates that increased
P21waf1/cip1 expression contributes to G2/M arrest, and
increased caspase-3 and Bax expression levels; however, a decreased
Bcl-2 expression contributes to apoptosis, which is induced by PSI
treatment in irradiated cells.
Discussion
In the present study, simulating the comparison for
the irradiation-survival curve of PC-9-ZD cells using a
multi-target click mathematical model formula reveals that the
D0, Dq and N values of cells is decreased following a
PSI treatment of irradiated cells. PSI enhances the
radiosensitivity effect of PC-9-ZD cells with an SER of 1.77. These
results indicate that, following treatment with PSI, the average
lethal dosage of the PSI + radiation group is decreased compared
with the control group, the shoulder is clearly decreased, and the
repair ability of cell sublethal injury is markedly decreased. The
radiosensitivity effect of PSI treatment is evidently increased in
the PC-9-ZD cells, the average lethal dosage is decreased with
irradiation, the decrease of shoulder is more significant and the
repair ability of cell sublethal injury is clearly decreased.
Further studies have been conducted to investigate
the radiosensitivity mechanism of PSI treatment. The results from
assessing the cell cycle and apoptosis in the present study
indicate that PSI predominantly induces G2/M phase arrest and
apoptosis. Apoptosis was the primary reason for cell death induced
by PSI in the irradiated cells. It was shown that a PSI combination
treatment advanced the apoptosis ratio up to a higher level. In
addition, PSI further increases apoptosis that is induced by
radioactive rays. Caspases are essential mediators of apoptosis.
Among them, caspase-3 is a frequently activated death protease,
catalyzing the specific cleavage of numerous key cellular proteins
(19). The Bcl-2 family, which
comprises of anti-apoptotic (including Bcl-2 and Bcl-xl) and
proapoptotic (including Bax and Bak) members, is the predominant
controller and mediator of cell apoptosis (20,21).
Particularly, the high Bcl-2/Bax ratio is considered to be a
crucial factor of cell resistance to apoptosis (22,23).
To investigate the role of PSI in the irradiation-induced apoptosis
pathway in gefitinib-resistant PC-9-ZD cells, the Bcl-2 family
proteins and the caspase-3 protein were analyzed in the present
study. The results indicated that Bcl-2 was decreased, and Bax and
caspase-3 were increased as a result of PSI treatment. Thus, PSI
promotes the irradiation-induced apoptosis via the association
between Bcl-2 and Bax, and caspase-3, eventually leading to
enhanced radiosensitivity.
Furthermore cell cycle arrest was the major reason
for cell death, which was induced by PSI in the irradiated cells.
Cell cycle regulation was significant for cell proliferation and
the cells exhibited varied radiosensitivity in various phases of
the cell cycle. Cells were most sensitive to irradiation during the
G2/M phase, less sensitive during G1, and least sensitive near the
end of the S phase (24). It was
shown in the present study that PSI treatment significantly altered
the cycle distribution of the irradiated cells, leading to cell
cycle arrest at the G2/M phase in a time-dependent manner, with an
increased cell density at the G2/M phase from 27.63 to 39.30%.
Previously, P21waf1/cip1 was considered to be the most
significant cell cycle checkpoint protein (25–27).
In the present study it was shown that PSI significantly increased
the expression of P21waf1/cip1, which resulted in
cell-cycle progression via G2/M arrest in the PC-9-ZD cells. This
indicates that P21waf1/cip1 is significant in mediating
cell growth through G2/M arrest in gefitinib-resistant cell
lines.
In conclusion, PSI exhibited potent radiosensitivity
against gefitinib-resistant PC-9-ZD cells in vitro. This
radiosensitivity was associated with the cell cycle arrest at the
G2/M phase and apoptosis via increased caspase-3, Bax and
P21waf1/cip1 and decreased Bcl-2 production. Therefore,
PSI may have the potential to be a radiosensitizer, however, this
requires further investigation.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81303274),
Wujieping Foundation of China (grant no. 320.6700.09035) and
Zhejiang traditional medicine Project (grant no. 2011ZZ011).
References
|
1
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinib sensitizing EGFR mutations in lung cancer
activate anti-apoptotic pathways. Science. 305:1163–1167. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Workman P: Altered states: selectively
drugging the Hsp90 cancer chaperone. Trends Mol Med. 10:47–51.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kobayashi N, Toyooka S, Soh J, et al: The
anti-proliferative effect of heat shock protein 90 inhibitor,
17-DMAG, on non-small-cell lung cancers being resistant to EGFR
tyrosine kinase inhibitor. Lung Cancer. 75:161–166. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jackman DM, Yeap BY, Sequist LV, et al:
Exon 19 deletion mutations of epidermal growth factor receptor are
associated with prolonged survival in non-small cell lung cancer
patients treated with gefitinib or erlotinib. Clin Cancer Res.
12:3908–3914. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Riely GJ, Pao W, Pham D, et al: Clinical
course of patients with non-small cell lung cancer and epidermal
growth factor receptor exon 19 and exon 21 mutations treated with
gefitinib or erlotinib. Clin Cancer Res. 12:839–844. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Giaccone G: Clinical impact of novel
treatment strategies. Oncogene. 21:6970–6981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grabley S and Thiericke R: Bioactive
agents from natural sources: trends in discovery and application.
Adv Biochem Eng Biotechnol. 64:101–154. 1999.PubMed/NCBI
|
|
8
|
Wang Y, Zhang YJ, Gao WY and Yan LL:
Anti-tumor constituents from Paris polyphylla var. yunnanensis.
Zhongguo Zhong Yao Za Zhi. 32:1425–1428. 2007.(In Chinese).
|
|
9
|
Sun J, Liu BR, Hu WJ, et al: In vitro
anticancer activity of aqueous extracts and ethanol extracts of
fifteen traditional Chinese medicines on human digestive tumor cell
lines. Phytother Res. 21:1102–1104. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee MS, Yuet-Wa JC, Kong SK, et al:
Effects of polyphyllin D, a steroidal saponin in Paris polyphylla,
in growth inhibition of human breast cancer cells and in xenograft.
Cancer Biol Ther. 4:1248–1254. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheung JY, Ong RC, Suen YK, et al:
Polyphyllin D is a potent apoptosis inducer in drug-resistant HepG2
cells. Cancer Lett. 217:203–211. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Siu FM, Ma DL, Cheung YW, et al: Proteomic
and transcriptomic study on the action of a cytotoxic saponin
(Polyphyllin D): induction of endoplasmic reticulum stress and
mitochondria-mediated apoptotic pathways. Proteomics. 8:3105–3117.
2008. View Article : Google Scholar
|
|
13
|
Jiang H, Su D and Ma SL: The effect of
Chonglou Saponin I on proliferation and apoptosis in lung
adenocarcinoma cell line PC9. J Chinese Oncol. 18:166–169.
2012.
|
|
14
|
Hua YH, Ma SL, Fu ZF, et al: Effect of
Polyphyllin I on radiosensitivity in nasopharyngeal carcinoma cell
line CNE-2 in vitro. Chinese Archives of Traditional Chinese
Medicine. 29:1387–1390. 2011.(In Chinese).
|
|
15
|
Xiao M, Dai X, He X, et al: Paris saponin
I induces G2/M cell cycle arrest and apoptosis in human
gastric carcinoma SGC7901 cells. J Huazhong Univ Sci Technol Med
Sci. 31:768–772. 2011.PubMed/NCBI
|
|
16
|
Xiao X, Bai P, Bui Nguyen TM, et al: The
antitumoral effect of Paris Saponin I associated with the induction
of apoptosis through the mitochondrial pathway. Mol Cancer Ther.
8:1179–1188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan LL, Zhang YJ, Gao WY, et al: In vitro
and in vivo anticancer activity of steroid saponins of Paris
polyphylla var. yunnanensis. Exp Oncol. 31:27–32. 2009.PubMed/NCBI
|
|
18
|
Ji Y, Ma SL, Zhang YP, et al: Combined
treatment with TNF-alpha/gefitinib alleviates the resistance to
gefitinib in PC-9 cells. Anticancer Drugs. 20:832–837. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shroff EH, Snyder C and Chandel NS: Bcl-2
family members regulate anoxia-induced cell death. Antioxid Redox
Signal. 9:1405–1409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Reed JC, Miyashita T, Takayama S, et al:
BCL-2 family proteins: regulators of cell death involved in the
pathogenesis of cancer and resistance to therapy. J Cell Biochem.
60:23–32. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sedlak TW, Oltvai ZN, Yang E, et al:
Multiple Bcl-2 family members demonstrate selective dimerizations
with Bax. Proc Natl Acad Sci USA. 92:7834–7838. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zha L, Qiao T, Yuan S, et al: Enhancement
of radiosensitivity by CpG-oligodeoxyribonucleotide-7909 in human
non-small cell lung cancer A549 cells. Cancer Biother Radiopharm.
25:165–170. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma S, Tang J, Feng J, et al: Induction of
p21 by p65 in p53 null cells treated with Doxorubicin. Biochim
Biophys Acta. 1783:935–940. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tang JJ, Shen C and Lu YJ: Requirement for
pre-existing of p21 to prevent doxorubicin-induced apoptosis
through inhibition of caspase-3 activation. Mol Cell Biochem.
291:139–144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiao GH, Beeser A, Chernoff J and Testa
JR: p21-activated kinase links Rac/Cdc42 signaling to merlin. J
Biol Chem. 277:883–886. 2002. View Article : Google Scholar : PubMed/NCBI
|