Introduction
Lung cancer is one of the most common malignant
tumors in the world. Clinical trials have shown that
cisplatin-based chemotherapy can significantly improve the survival
rate of inoperable non-small cell lung cancer patients (1), but resistance to cisplatin limits its
wider use in clinical applications. At present, the overall
five-year survival rate of lung cancer patients is <15%
(2). Cisplatin resistance is the
main reason for the failure of cancer chemotherapy, which
contributes to the difficulty in providing a lung cancer cure and a
poor long-term survival rate. Therefore, finding an effective
method or medicine to reverse cisplatin resistance is a reasonable
strategy to solve the drug resistance problem in lung cancer. To
this end, the study of the mechanism of cisplatin resistance in
lung cancer is key. During chemotherapy, cisplatin takes effect
through the induction of lung cancer cell apoptosis. The resistance
to cisplatin-induced cell apoptosis is considered as one of the
important mechanisms of drug resistance in lung cancer (3).
Metformin (Met) has been widely used in hypoglycemic
therapy in patients with type 2 diabetes (4). It has been shown in clinical
observations that Met could reduce the incidence of tumors and the
mortality of cancer patients (5).
In recent years, numerous studies have confirmed that Met has
anti-cancer effects (5–10). However the mechanism with regard to
how Met affects the proliferation of lung cancer is not clear. It
is also unclear whether Met could increase the sensitivity of
cancer cells to cisplatin treatment. The present study investigated
the effect of Met on the proliferation of different lung cancer
cells and its effect on the cisplatin resistance. The study also
provides a discussion on the mechanism of Met action, and conducts
a preliminary evaluation of Met as an anticancer drug.
Materials and methods
Cell culture
The human lung cancer A549, cisplatin-resistant lung
cancer A549/CDDP, SPCA and H23 cell lines were kept in the
Institute of Biochemistry and Molecular Biology in Guangdong
Medical College (Zhanjiang, Guangdong, China). The cell lines were
cultured in the Dulbecco’s modified Eagle’s medium (Gibco BRL,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum
(Sijiqing Laboratories, Hangzhou, Zhejiang, China), 100 μg/ml
penicillin and 100 μg/ml streptomycin. The cells were incubated at
37°C in a humidified atmosphere with 5% CO2.
Reagents
Met and MTT were purchased from Sigma-Aldrich (St.
Louis, MO, USA). Rabbit and goat polyclonal antibodies against
cytochrome c, BH3 interacting-domain death agonist (Bid),
poly (ADP-ribose) polymerase (PARP), caspase-3, caspase-8, β-actin
and SB203580 were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The antibodies against phosphorylated forms
of p38 MAPK-Thr180/Tyr182 and against p38 MAPK were purchased from
Cell Signaling Technology (Beverly, MA, USA).
Cell viability assay
The A549, A549/CDDP, SPCA and H23 cells were treated
with different concentrations of Met (0, 0.5, 1, 2,4 and 8 mmol/l)
for various times (24, 48 and 72 h) and the cell viability was
determined by MTT assay, as previously described (11).
Analysis of cell apoptosis by
fluorescence staining
The A549/CDDP cells were treated with 4 mM Met for
the different time periods at 37°C. Priot to the cells being
examined under a fluorescence microscope, they were incubated in
Hoechst 33258 (10 mg/l) solution at 37°C for 20 min. Apoptosis was
evaluated by the uptake of Hoechst 33342 (12). The apoptotic index was determined by
dividing the number of apoptotic nuclei by the number of total
nuclei.
Analysis of the cell cycle by flow
cytometry
The cells were collected by centrifugation at 1,000
× g following treatment with 4 mM Met for the different time
periods at 37°C. The cells were then washed twice with
phosphate-buffered saline (PBS) and fixed with ice-cold 70% ethanol
overnight. Prior to the flow cytometry analysis for cell cycle
distribution, the fixed cells were washed once with PBS
(Sigma-Aldrich) and incubated with 100 μg/ml propidium iodide
(Sigma-Aldrich) plus 200 μg/ml RNase (Sigma-Aldrich).
Treatment with p38 MAPK inhibitors
The ells were preincubated with the MAPK inhibitor,
SB203580 (10 μM), for 2 h and then treated with Met (4 mM) for 24
h. Cell viability was determined using an MTT assay and the protein
levels were measured by western blotting.
Western blotting
The cells were lysed with cell lysis buffer (pH 8.0)
containing 50 mM Tris-HCl (Sigma-Aldrich), 150 mM NaCl (Guangzhou
Chemical Reagent Factory, Guangzhou, China), 5 mM EDTA (Guangzhou
Chemical Reagent Factory), 1% NP40 (Sigma-Aldrich), 0.05%
phenylmethanesulfonyl fluoride (Sigma-Aldrich), 2 μg/ml aprotinin
(Sigma-Aldrich) and 2 μg/ml leupeptin (Sigma-Aldrich). The protein
levels were determined by western blotting, as described previously
(4).
Treatment of A549/CDDP cells with Met and
cisplatin
The A549/CDDP cells were treated with the different
concentrations of Met and cisplatin for 24 h at 37°C. Cell
viability was determined using an MTT assay.
Statistical analysis
Results are presented as the mean ± standard
deviation. Statistical analysis was performed using a one-way
analysis of variance with a least significant difference test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of Met on the growth of different
lung cancer cell lines
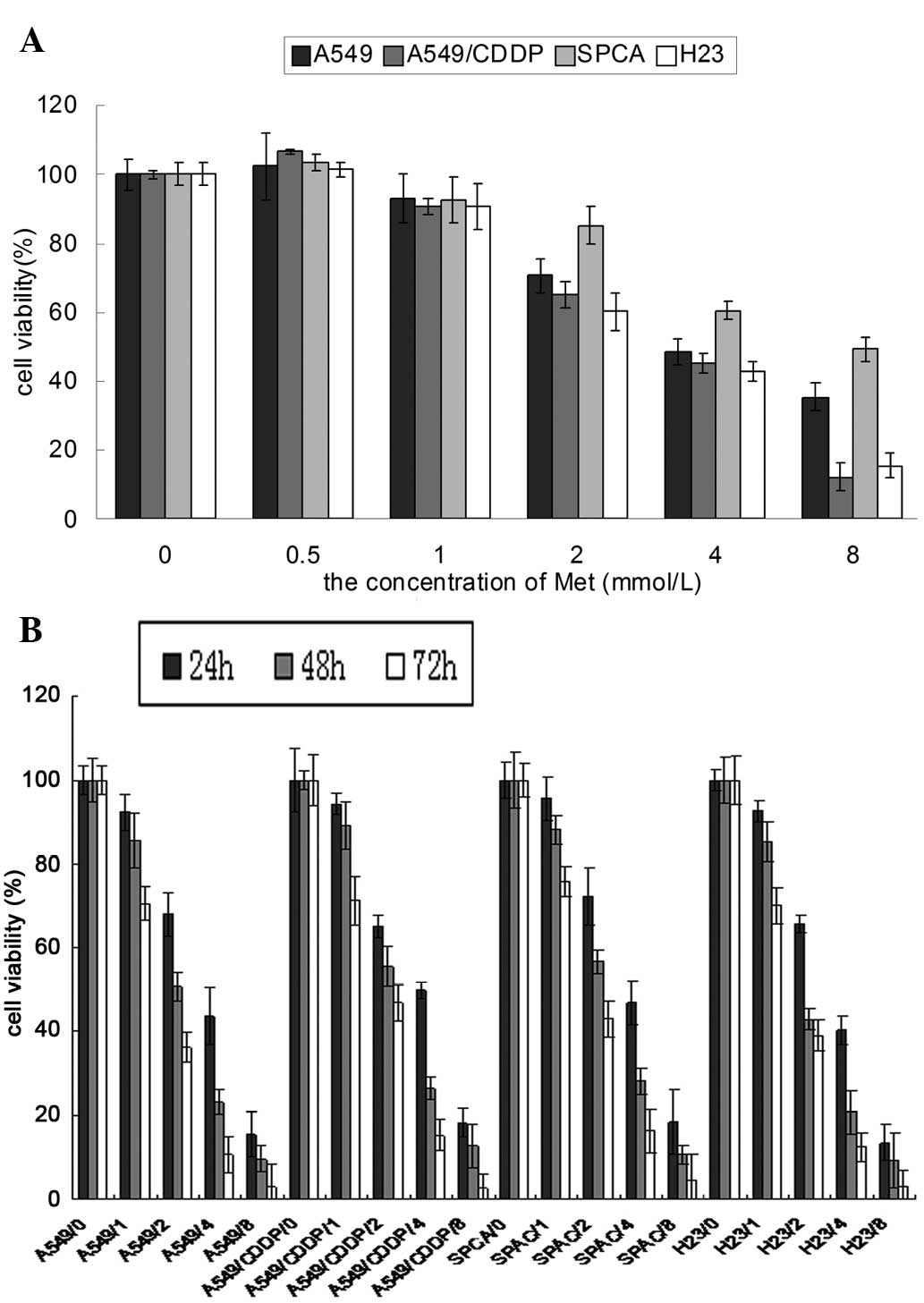
Fig. 1 showed that
Met inhibited the proliferation of the lung cancer cells in a
concentration- and time-dependent manner. The proliferation of the
lung cancer cells was significantly inhibited by 24 h of Met
treatment at concentrations of 2–8 mmol/l (P<0.05). The survival
of the lung cancer cells decreased significantly compared with the
control group following 48 h of treatment with 1 mmol/l Met
(P<0.05). It was also shown that the response of the A549/CDDP
cells was similar to its parent cell line, A549.
Met inhibits the A549/CDDP cell cycle and
induces apoptosis
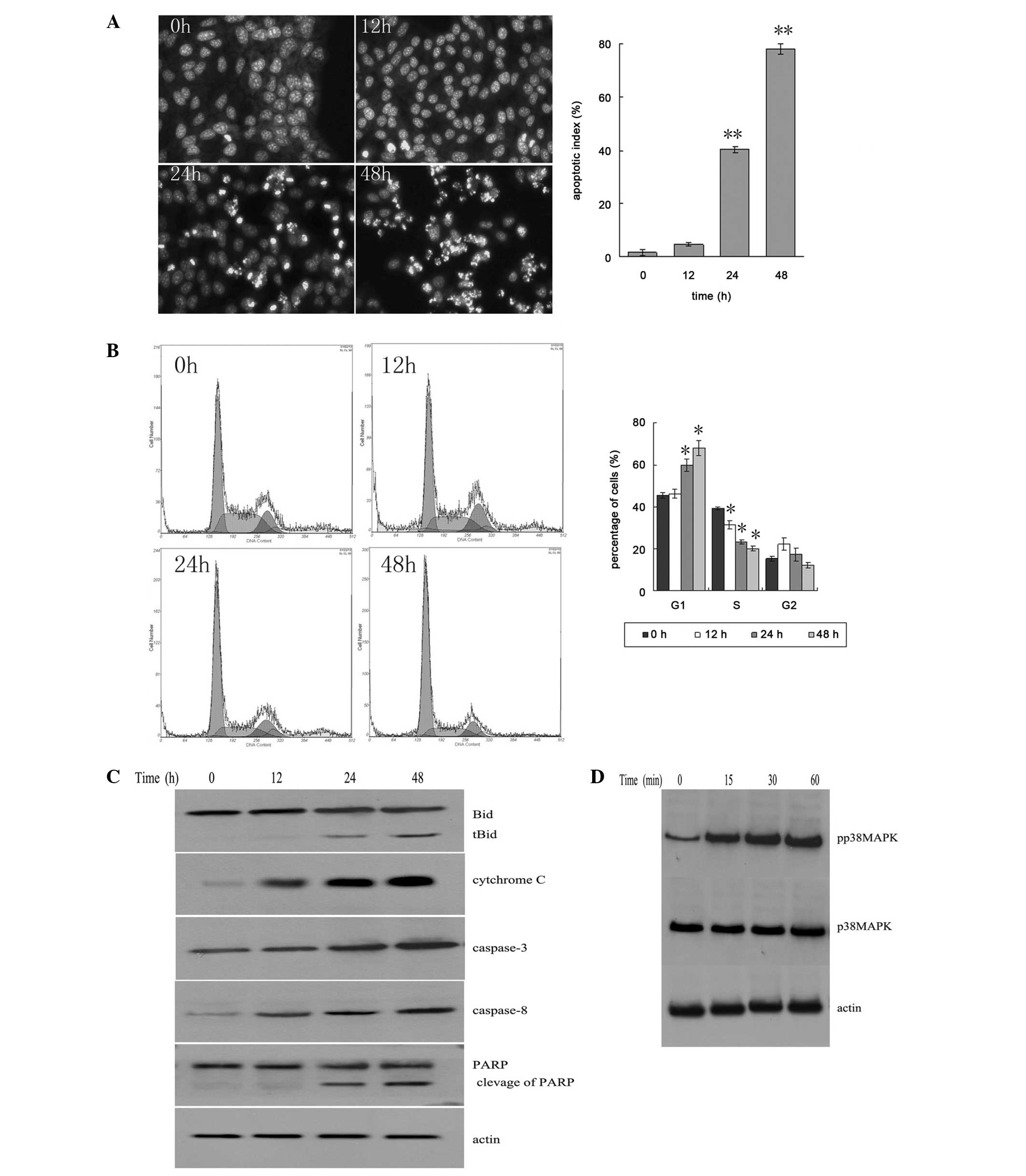
The results showed that a large number of apoptotic
cells appeared following 24–48 h of Met treatment (Fig. 2A). This was demonstrated by the
apoptotic bodies in the cells stained with Hoechst 33258 (Fig. 2B). With Met treatment, the number of
cells in the G1 phase was increased, and the number of
cells in the S phase and G2 phase was decreased
(Fig. 2C).
Met induces the cleavage of Bid and
PARP
The fragments of Bid and PARP increased with the
increase in incubation time. The levels of caspase-3, caspase-8 and
cytochrome c increased in the cytosol (Fig. 2C). It was shown that the
phosphorylation level of p38 MAPK was increased following 15 min of
Met treatment, indicating that Met could activate the p38 MAPK
pathway (Fig. 2D).
Reduction of Met-induced cell apoptosis
by inhibition of p38 MAPK activity
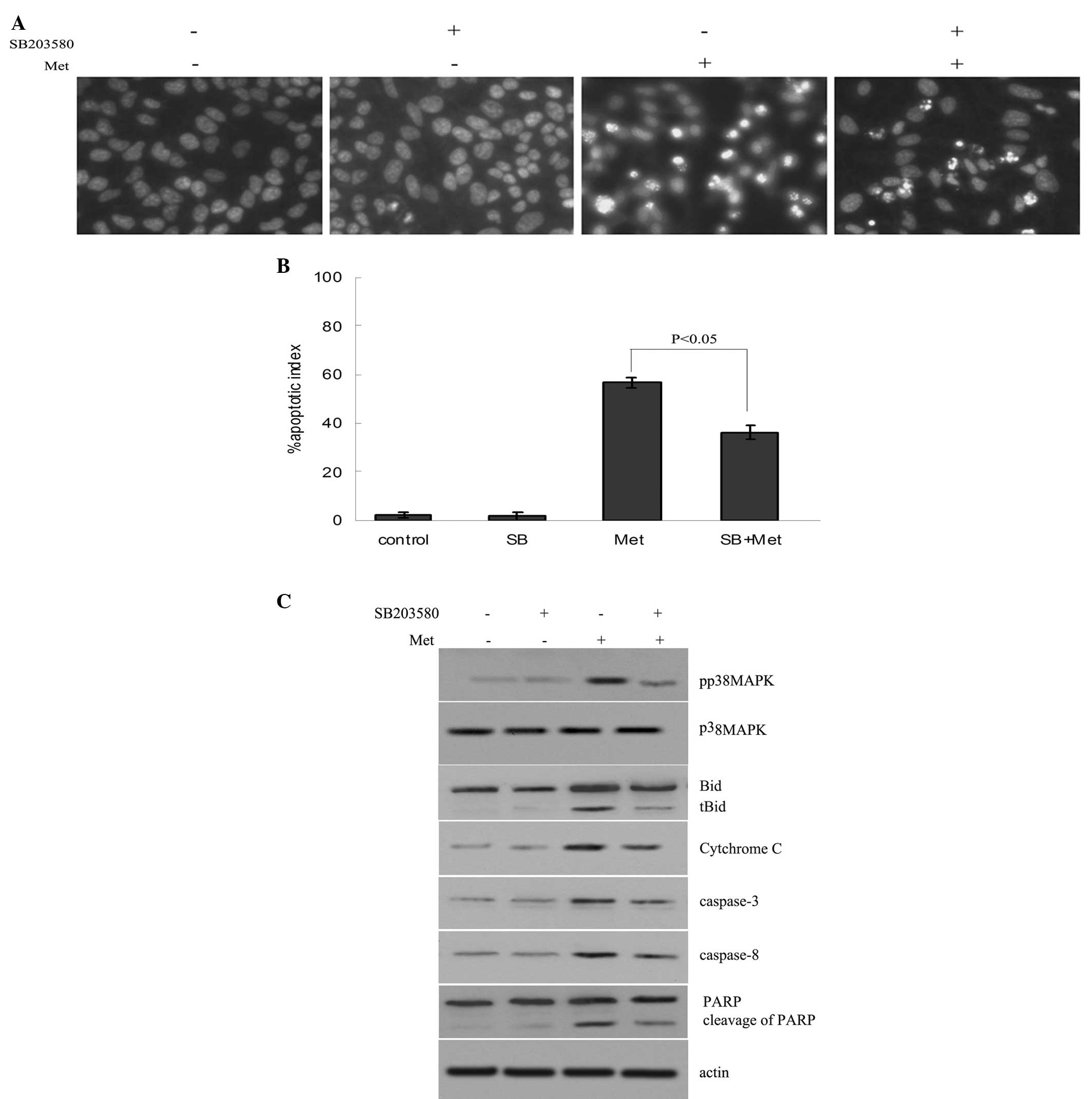
The A549/CDDP cells were preincubated with 10 μM
SB203580 (a p38 MAPK inhibitor) for 2 h, and then treated with 4 mM
Met for 24 h. The results showed that Met activated p38 MAPK and
induced the apoptosis of the A549/CDDP cells. Met-induced apoptosis
was inhibited by SB203580 (Fig. 3A and
B). The data also showed that SB203580 decreased the levels of
Bid and PARP cleavage fragments, and the caspase-3, caspase-8 and
cytosolic cytochrome c levels (Fig. 3C).
Met enhances the sensitivity of A549/CDDP
cells to cisplatin
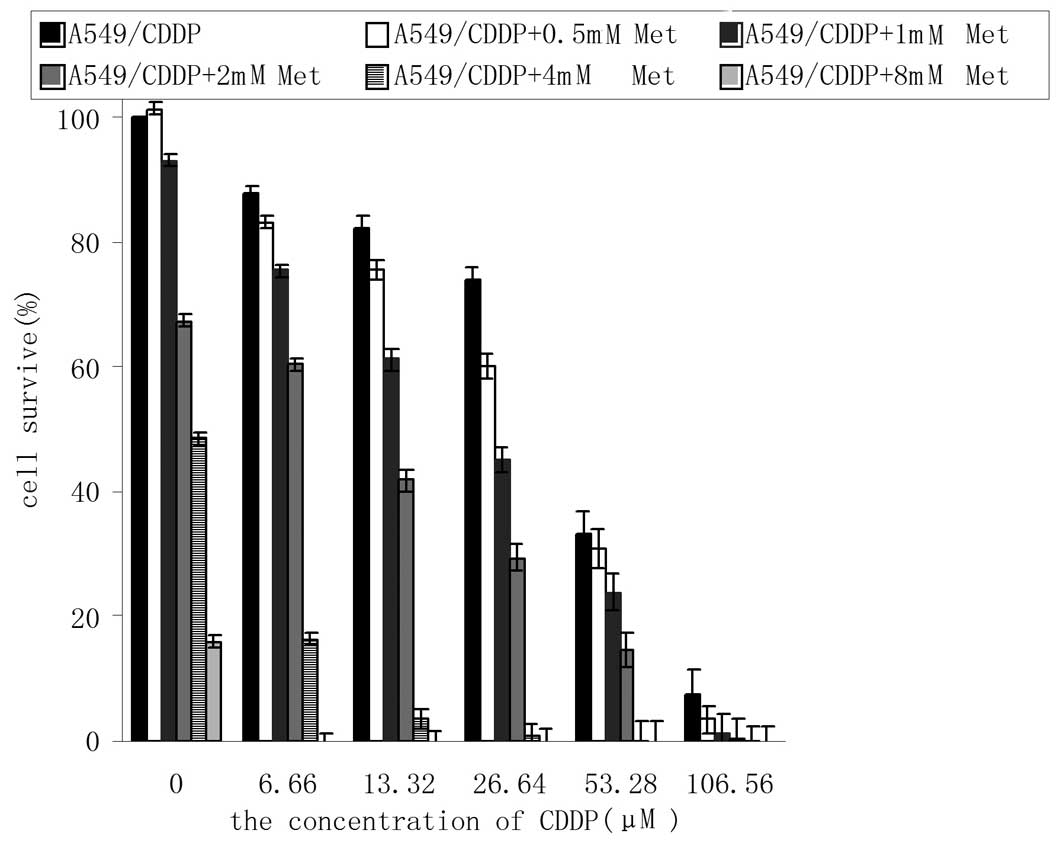
Consistent with our previous study (12), the results showed that the
IC50 values of cisplatin for the A549 cells and
A549/CDDP cells were 2.36±0.10 and 30.27±1.50 μmol/l, respectively.
The difference between the cisplatin-resistant and parent cells was
12.8-fold.
In this experiment, it was shown that the
IC50 of cisplatin for the A549/CDDP cells was 32.18±1.15
μmol/ in the absence of Met, while the IC50 values
decreased to 27.45±2.38, 19.56±1.60, 9.65±1.38, 0.06±0.07 and
0.00±0.05 μmol/l respectively in the presence of 0.5, 1, 2, 4 and 8
mmol/l Met (P<0.05 vs. control) (Fig. 4). Met evidently increased the
cytotoxicity of cisplatin.
Discussion
Every year, 1,200,000 patients are diagnosed with
lung cancer, and ~25% of all cancer mortalities are patients with
lung cancer, representing a large threat to human health (13,14).
Chemotherapy and radiotherapy are the major methods for the
treatment of patients with lung cancer. However, chemotherapy drugs
have toxicity to normal tissue, and may cause serious side-effects
in the clinic. Furthermore, the chemotherapy and radiotherapy drugs
are expensive. Therefore it is of great import to search for
effective antitumor drugs with a low toxicity and cost, which can
enhance the effect of chemotherapy and improve the prognosis of
lung cancer patients.
Met is an effective antihyperglycemic agent, which
has been widely used in the treatment of patients with type 2
diabetes for decades. Certain studies have shown that Met not only
inhibits breast cancer growth in HER-2/neu transgenic mice
(6), but that it also inhibits
pancreatic cancer induced by feeding of a high-fat diet in hamsters
(7). Clinical data has also
revealed that Met has anti-tumor activity. Evans et al
(5) reported that the risk for
tumorigenesis in type 2 diabetes patients administered Met
treatment is 23% lower than those subjected to sulfonylurea
treatment. Furthermore, after following 10,309 patients with newly
diagnosed type 2 diabetes for approximately five years, Bowker
et al (8) concluded that
treatment with Met could be associated with lower mortality for
malignancies in comparison with sulfonylureas in type 2 diabetes
patients. Buzzai et al (9)
demonstrated that Met has toxicity towards p53-mutated colorectal
cancer cells. Ben Sahra et al (10) showed that Met inhibits the
proliferation of prostate cancer cells, but not normal prostate
epithelial cells. The present study data also showed that Met
inhibited lung cancer cell growth and proliferation in a time- and
concentration-dependent manner, which is consistent with previous
results (5–9). In addition, the A549/CDDP cells were
sensitive to Met. These findings suggest that Met may have a wide
range of antitumor effects.
Cell proliferation is mainly affected by two
factors, the cell cycle and apoptosis. Ben Sahra et al
(10) showed that Met suppresses
prostate cancer cell proliferation by arresting the cell cycle in
the G1 phase. However, it has no evident effect on cell
apoptosis. However the present results demonstrated that Met not
only induced A549/CDDP cell arrest at the G1 phase, but
that it also promoted cell apoptosis.
Cisplatin is the most commonly used non-specific
anticancer drug, which binds to tumor cell DNA and interferes with
its function (15). The present
study found no significant difference in the sensitivity to Met
between the A549/CDDP cells and its parent cell line, A549.
Furthermore, Met enhanced the sensitivity of the A549/CDDP cells to
cisplatin when the cells were treated with Met and cisplatin
together. The results suggest that Met may be used to enhance the
cisplatin toxicity in cisplatin-resistant lung cancer cells and to
improve the effectiveness of cisplatin-based chemotherapy in lung
cancer patients.
Cisplatin has been used to treat tumors based on its
ability to induce the apoptosis of tumor cells. One important
mechanism of drug resistance in lung cancers is the tolerance of
cancer cells to cisplatin-induced apoptosis. Apoptosis pathways
include mainly the death receptor, mitochondrial and endoplasmic
reticulum stress pathways. p38 MAPK and caspase family proteins
play key roles in these apoptotic signal pathways.
The activation of p38 MAPK can induce apoptosis in
various tumor cells. Jiang et al (16) reported that selenite activated p38
MAPK activity and promoted apoptosis in Jurkat cells. Inhibiting
the p38 MAPK pathway can effectively reduce the release of
cytochrome c and inhibit Caspase-3 activation and PARP
cleavage, resulting in the decrease of apoptosis cells (17). Mandal et al (18) showed that the p38 MAPK pathway is
indispensable in the apoptosis of leukemia cells induced by
Withaferin A. The p38 MAPK signaling pathway plays a key role in
promoting the apoptosis of cells, a process associated with the
activation of p53 and caspases, and the translocation of Bax
(19,20). A study by Khan et al
(21) demonstrated that the
activation of the p38 MAPK pathway can induce the activation of
caspases, and that the inhibition of p38 MAPK can reduce the
expression levels of caspase activated protein. The study suggested
that p38 MAPK is an upstream regulator of the caspase-dependent
signal transduction pathway of apoptosis (21). The present study showed that Met
induced the phosphorylation of the p38 MAPK protein and increased
the expression of caspase-8 and caspase-3, along with the cleavage
of Bid and PARP, and cytochrome c release in the cytoplasm.
These results suggest that Met may promote the expression of
caspase-8 by activating p38 MAPK phosphorylation. Bid was cleaved
by activated caspase-8 to produce tBid, which migrated to the
mitochondrial membrane, resulting in the change of mitochondrial
membrane permeability, cytochrome c release, caspase-3
activation and cleavage of PARP to promote apoptosis. Inhibition of
p38 MAPK significantly reduced the Met-induced apoptosis of the
A549/CDDP cells. These results indicate that the p38 MAPK-caspase
pathway may be involved in the regulation of A549/CDDP cell
apoptosis induced by Met. Although the inhibition of the p38 MAPK
signal pathway reduced the level of Met-induced A549/CDDP cell
apoptosis, it did not completely inhibit the apoptosis of the
A549/CDDP cells. This result suggests that other factors may
participate in the regulation of A549/CDDP cell apoptosis induced
by Met.
Results of the present study support the hypothesis
that the proliferation of A549/CDDP cells can be inhibited by Met,
which may be used as an adjuvant therapy to improve the clinical
treatment effect of cisplatin in cisplatin-resistant lung cancers.
Met is widely used as an insulin sensitizer and has shown few toxic
side-effects in a number of years of clinical practice. Further
studies are required to explore the other clinical applications of
Met.
Acknowledgements
This project was supported by the Medical Scientific
Research Foundation of Guangdong Province (A2011439), the
Administration of Traditional Chinese Medicine of Guangdong
Province (No.20111239) and the Startup Foundation for Doctors of
The Affiliated Hospital of Guangdong Medical College (no.
10301B010014).
Abbreviations:
|
Met
|
metformin
|
|
MAPK
|
mitogen-activated protein kinase
|
References
|
1
|
Winton T, Livingston R, Johnson D, Rigas
J, Johnston M, Butts C, et al; National Cancer Institute of Canada
Clinical Trials Group; National Cancer Institute of the United
States Intergroup JBR. 10 Trial Investigators. Vinorelbine plus
cisplatin vs observation in resected non-small-cell lung cancer. N
Engl J Med. 352:2589–2597. 2005.
|
|
2
|
Risch A and Plass C: Lung cancer
epigenetics and genetics. Int J Cancer. 123:1–7. 2008.
|
|
3
|
Stewart DJ: Mechanisms of resistance to
cisplatin and carboplatin. Crit Rev Oncol Hematol. 163:12–31.
2007.
|
|
4
|
Gallwitz B and Bretzel RG: How do we
continue treatment in patients with type 2 diabetes when
therapeutic goals are not reached with oral antidiabetes agents and
lifestyle?. Incretin versus insulin treatment. Diabetes Care.
36:S180–S189. 2013.
|
|
5
|
Evans JM, Donnelly LA, Emslie-Smith AM,
Alessi DR and Morris AD: Metformin and reduced risk of cancer in
diabetic patients. BMJ. 330:1304–1305. 2005.
|
|
6
|
Schneider MB, Matsuzaki H, Haorah J,
Ulrich A, Standop J, Ding XZ, Adrian TE and Pour PM: Prevention of
pancreatic cancer induction in hamsters by metformin.
Gastroenterology. 120:1263–1270. 2001.
|
|
7
|
Anisimov VN, Berstein LM, Egormin PA,
Piskunova TS, Popovich IG, Zabezhinski MA, Kovalenko IG, Poroshina
TE, Semenchenko AV, Provinciali M, Re F and Franceschi C: Effect of
metformin on life span and on the development of spontaneousmammary
tumors in HER-2/neu transgenic mice. Exp Gerontol. 40:685–693.
2005.
|
|
8
|
Bowker SL, Majumdar SR, Veugelers P and
Johnson JA: Increased cancer-related mortality for patients with
type 2 diabetes who use sulfonylureas or insulin. Diabetes Care.
29:254–258. 2006.
|
|
9
|
Buzzai M, Jones RG, Amaravadi RK, Lum JJ,
DeBerardinis RJ, Zhao F, Viollet B and Thompson CB: Systemic
treatment with the antidiabetic drug metformin selectively impairs
p53-deficient tumor cell growth. Cancer Res. 67:6745–6752.
2007.
|
|
10
|
Ben Sahra I, Laurent K, Loubat A,
Giorgetti-Peraldi S, Colosetti P, Auberger P, Tanti JF, Le
Marchand-Brustel Y and Bost F: The antidiabetic drug metformin
exerts an antitumoral effect in vitro and in vivo through a
decrease of cyclin D1 level. Oncogene. 27:3576–3586. 2008.
|
|
11
|
Zhang HT, Wu J, Wen M, Su LJ and Luo H:
Galangin induces apoptosis in hepatocellular carcinoma cells
through the caspase 8/t-Bid mitochondrial pathway. J Asian Nat Prod
Res. 14:626–633. 2012.
|
|
12
|
Wu J, Hu CP, Gu QH, Li YP and Song M:
Trichostatin A sensitizes cisplatin-resistant A549 cells to
apoptosis by up-regulating death-associated protein kinase. Acta
Pharmacol Sin. 31:93–101. 2010.
|
|
13
|
Jemal A, Murray T, Ward E, Samuels A,
Tiwari RC, Ghafoor A, Feuer EJ and Thun MJ: Cancer statistics,
2005. CA Cancer J Clin. 55:10–30. 2005.
|
|
14
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007.
|
|
15
|
Florea AM and Büsselberg D: Cisplatin as
an anti-tumor drug: cellular mechanisms of activity, drug
resistance and induced side effects. Cancers (Basel). 15:1351–1371.
2011.
|
|
16
|
Jiang Q, Li F, Shi K, Wu P, An J, Yang Y
and Xu C: ATF4 activation by the p38MAPK-eIF4E axis mediates
apoptosis and autophagy induced by selenite in Jurkat cells. FEBS
Lett. 587:2420–2429. 2013.
|
|
17
|
Neoh CA, Wang RY, Din ZH, Su JH, Chen YK,
Tsai FJ, Weng SH and Wu YJ: Induction of apoptosis by sinulariolide
from soft coral through mitochondrial-related and p38 MAPK pathways
on human bladder carcinoma cells. Mar Drugs. 10:2893–2911.
2012.
|
|
18
|
Mandal C, Dutta A, Mallick A, Chandra S,
Misra L, Sangwan RS and Mandal C: Withaferin A induces apoptosis by
activating p38 mitogen-activated protein kinase signaling cascade
in leukemic cells of lymphoid and myeloid origin through
mitochondrial death cascade. Apoptosis. 13:1450–1464. 2008.
|
|
19
|
Chang HL, Chen CY, Hsu YF, Kuo WS, Ou G,
Chiu PT, Huang YH and Hsu MJ: Simvastatin induced HCT116 colorectal
cancer cell apoptosis through p38MAPK-p53-survivin signaling
cascade. Biochim Biophys Acta. 1830:4053–4064. 2013.
|
|
20
|
Yang X, Yao J, Luo Y, Han Y, Wang Z and Du
L: P38 MAP kinase mediates apoptosis after genipin treatment in
non-small-cell lung cancer H1299 cells via a mitochondrial
apoptotic cascade. J Pharmacol Sci. 121:272–281. 2013.
|
|
21
|
Khan R, Khan AQ, Qamar W, Lateef A, Tahir
M, Rehman MU, Ali F and Sultana S: Chrysin protects against
cisplatin-induced colon toxicity via amelioration of oxidative
stress and apoptosis: probable role of p38MAPK and p53. Toxicol
Appl Pharmacol. 258:315–329. 2012.
|