1. Discovery and function of caveolin
(Cav)-1
The identification of Cav-1 began with a study into
the morphological observations of the caveolae in the 1950s.
Caveolae are morphologically identifiable plasma membrane
invaginations that can be identified by electron microscopy;
caveolae, which are 50–100 nm in size (1), appear as vesicles at the plasma
membrane. Cav (now termed Cav-1) was initially identified as one of
the four major proteins to be resistant to extraction with
non-ionic detergents and to demonstrate a staining pattern in Rous
sarcoma-transformed chicken embryonic fibroblasts (2). Cav-2 and Cav-3 were subsequently
identified through various experiments (3,4).
Further study has indicated that Cav-1 expression is sufficient and
necessary to drive the formation of morphologically identifiable
caveolae (5), making it the first
true protein marker of caveolae (6). To date, three members of the Cav gene
family have been identified. The gene locus of Cav-1 is on human
chromosome 7q31.1, located adjacent to Cav-2 (~19 kb apart), while
Cav-3 is located on a different chromosome (3p25) (7). Two study groups (8,9)
independently revealed that Cav-1 is identical to the vesicular
integral-membrane protein of 21 kDa and, subsequently, was
confirmed to be the same protein.
As the first member of the Cav family (including
Cav-1,-2 and -3), Cav-1, a 22-kDa protein of 178 amino acids, has
been the most sufficiently investigated by a number of biochemical
studies. Cavs are found predominantly at the plasma membrane,
however, their expression levels vary considerably between tissues.
The highest levels of Cav-1 are found in the terminally
differentiated cells, such as adipocyte, endothelia and smooth
muscle cells, as well as type I pneumocytes. The localization and
expression of Cav-2 is mapped to Cav-1 and is required for the
proper membrane localization of Cav-2, whereas Cav-3 is expressed
predominantly in the muscle cells, including the smooth, skeletal
and cardiac myocyte cells (4).
It was initially identified that Cav-1 is resistant
to extraction with sodium carbonate and high salt concentrations,
which demonstrated that it is an integral membrane protein
(6). It has been suggested that the
amino and carboxyl termini of Cav-1 face the cytoplasm, with a
hydrophobic domain inserted into the membrane via the classical
endoplasmic reticulum machinery. The membrane insertion (residues
102–134) are considered to form a unique hairpin loop configuration
that prevents Cav-1 from completely spanning the plasma membrane in
a traditional double-pass fashion (9). Mutational analysis and domain mapping
experiments have demonstrated the importance of two other regions
of Cav-1 that bind to membrane with high affinity (11–14).
These regions are now known as the NH2-terminal membrane
attachment domain (N-MAD; residues 82–101) and COOH-terminal
membrane attachment domain (residues 135–150). The oligomerization
domain (residues 61–101) of Cav-1 meditates the
homo-oligomerization of 14–16 Cav-1 isoforms (15), which subsequently form high
molecular mass oligomers of ~400 kDa through several stages of
oligomerization. The N-MAD (residues 82–101) are also termed the
Cav scaffolding domain (CSD). Couet et al (16) identified two Cav binding motifs
(CBMs; ϕχϕχχϕ and ϕχχχχϕχχ, where ϕ represents an aromatic amino
acid and χ represents a non-aromatic amino acid) in the majority of
proteins matched to the CSD of Cav-1. Via an interaction between
the CSD of Cav-1 and Cav binding domain. of a given
caveolae-associated protein, a number of specific signaling
molecules may be concentrated and regulated by Cav-1, including
G-protein subunits, receptor or non-receptor tyrosine kinases, and
endothelial nitric oxide (NO) synthase (NOS). As a scaffolding
protein, Cav-1 also serves as a signal transduction molecule that
inhibits or enhances the signal activity of a given
caveolae-associated protein. Furthermore, numerous studies have
suggested that Cav-1 serves as a negative regulator of cell
proliferation or as a tumor suppressor.
2. Cav-1 and cancer-associated fibroblasts
(CAFs)
Prognostic value of the downregulation of
stromal Cav-1 expression
To date, the conception of tumorigenesis has been
exclusively focused on the transformation of cancer cells
themselves to the complex cross-talk between cancer cells and the
tumor microenvironment. Furthermore, CAFs constitute a major
portion of the tumor microenvironmental elements, including the
extracellular matrix (ECM) (1),
pericytes (2), endothelial cells,
immune and inflammatory cells (5)
and secreted diffusible growth factors/cytokines (17). A number of studies have suggested
that CAFs are key in cancer progression. CAFs retain a major role
in ECM remodeling that has been reported to influence the
proliferation, survival and migration of cancer cells (18–21).
In addition, activated CAFs secrete components including collagen
types I and IV, extra domain A-fibronectin, hepatocyte growth
factor, epidermal growth factor, basic fibroblast growth factor,
extracellular matrix components and matrix metalloproteinases
(22,23). CAFs also show an ability to prevent
cancer cell apoptosis and induce the proliferation of surrounding
cancer cells. Compared with normal fibroblasts (NFs) mixed with
epithelial tumor cells, CAFs are more accomplished at enhancing
tumor growth and give rise to highly vascularized tumors. According
to associated studies (24–26), certain biological molecules can be
recognized as biomarkers of CAFs, including α-smooth muscle actin,
fibroblast-specific protein 1, fibroblast activation protein and
PDGFRα/β. However, there is little knowledge regarding the origin
of CAFs and the mechanisms of phenotype transformation from benign
to heterogeneous fibroblasts (such as CAFs).
As a principal component of the protein coat of
caveolae, Koleske et al (27) observed that the level of Cav was
clearly reduced in a NIH 3T3 cell line transformed by the
expression of the v-abl, bcr-abl, H-ras, polyomavirus middle T
antigen or crkl oncogenes, and suggested that the deregulation of
Cav (now termed Cav-1) may promote oncogenic transformation.
However, Cav-1 has been found to be expressed in the plasma
membrane of various types of differentiated cells. The
downregulation of Cav-1 is a major characteristic of CAFs and
existing studies have indicated that CAFs have the ability to
prevent cancer cell apoptosis, enhance the proliferation of cancer
cells and stimulate tumor angiogenesis. It is also implicated that
the downregulation of Cav-1 is one of the mechanisms that mediates
the transformation of fibroblasts.
The majority of these studies have concentrated on
breast CAFs. Mercier et al (28) were the first to demonstrate that the
Cav-1 protein is downregulated in human breast cancer (eight out of
11 patients showed a marked downregulation of Cav-1 protein
expression in CAFs by western blot analysis), and observed that
CAFs are more numerous in human breast cancer, with an elongated
appearance and hyperproliferative when compared with NFs,
suggestive of a transformed phenotype. The treatment of
Cav-1-deficient CAFs with a Cav-1 mimetic peptide can reverse the
hyperproliferation phenotype with a three-fold reduction. Sotgia
et al (29) further
established a direct cause-effect association between stromal
Cav-1-deficient and CAF phenotypes, by creating a Cav-1(−/−)
mammary stromal fibroblast (MSF) cell line. The authors showed that
Cav-1(−/−) MSFs share a number of properties with human CAFs,
including similar gene profiles, the functional inactivation of the
retinoblastoma (RB) tumor suppressor and the functional
characteristics of myofibroblasts. Such initial discoveries lead to
the proposal that Cav-1 may serve as a cancer prognostic factor.
Sloan et al (30) further
analyzed tissue sections specifically for the stromal and tumor
epithelial cell expression of Cav-1 from two cohorts of breast
cancer patients. In total, 103 out of 173 patients (60%) and 31 out
of 429 patients (7%) exhibited unambiguous staining for the stromal
compartment of the tumor and epithelial tumor cells, respectively.
According to El-Gendi et al and Witkiewicz et al
(31,32), the presence of Cav-1 in epithelial
tumor cells positively correlates with lower TNM tumor stage
(P=0.05), but does not predict the cancer-specific survival of more
than five years. By contrast, the loss of stromal Cav-1 has been
found to positively correlate with the previously described
clinical characteristics of breast cancer (30). Together with Witkiewicz et al
(33), these studies separately
revealed an independent prognostic value of the downregulation of
breast tumor stromal Cav-1. The 10-year survival rate for patients
with tumors positive for Cav-1 expression in the stroma is 91%,
when compared with 43% for patients lacking stromal Cav-1
(P=0.0001) (30). Similarly, the
loss of stromal Cav-1 expression predicts poor clinical outcome in
triple negative and basal-like breast cancers (32). The overall survival rate has also
been found to decrease with the deregulation of tumor stromal
Cav-1. Notably, TN patients with high-levels of stromal Cav-1 have
a good clinical outcome, with >50% of the patients surviving the
follow-up period. By contrast, the median survival time for TN
patients with moderate stromal Cav-1 staining is 33.5 months.
Similarly, the median survival time for TN patients with absent
stromal Cav-1 staining is 25.7 months (32). In a combined study of 358 resected
breast cancers cocultured with Cav-1 siRNA-treated fibroblasts and
the MDA-MB-468 cell line, Simpkins et al (34) identified that the loss of Cav-1
expression significantly correlates with decreased breast
cancer-specific and disease-free survival (P=0.01), through
promotion of breast cancer cell invasion. Overall, this
clinicopathological study of breast cancer revealed a significant
correlation between the absence of stromal Cav-1, and larger tumor
size, advanced tumor stage (TNM stage), higher grade, lymph node
metastasis, poor tumor prognosis and short overall survival
time.
Similar clinical values of decreased stromal Cav-1
levels have also been found in gastric cancer (GC) (35,36),
prostate cancer (PC) (37,38) and malignant melanoma (39). Additionally, Zhao et al
(35) found that positive rates of
epithelial Cav-1 expression in gastritis without intestinal
metaplasia (IM), gastritis with IM and GC showed a decreasing trend
compared with gastritis without IM, gastritis with IM and GC
(P=0.012). Furthermore, no significant correlation was identified
between tumor cells and CAF Cav-1 expression (P=0.751). The
expression of Cav-1 in CAFs was also found to significantly
correlate with disease-free survival (P=0.029) and overall survival
(P=0.013) (35). In addition, the
downregulation of stromal Cav-1 was found to predict poor survival,
early recurrence and a lower cumulative five-year survival rate for
GC patients. Furthermore, multivariate analysis (COX
proportional-hazard regression model) revealed (35) that CAF Cav-1 expression is an
independent predictor of recurrence and survival in GC patients,
consistent with the study by He et al (36). No correlation has been identified
between the expression stature of stromal Cav-1 and the typical
clinicopathological parameters of GC, such as T stage, TNM stage
and Lauren classification. Notably, no correlation has been
identified between Cav-1 expression in tumor cells, and the
prognosis and clinicopathological parameters of GC. The decreased
trend of stromal Cav-1 in patients with benign prostatic
hypertrophy, primary PCs and PC metastases have also been
identified (38). Furthermore, a
large cohort of 724 PC patients demonstrated a significant
correlation between decreased levels of stromal Cav-1, and
increased Gleason score (P=0.012) and reduced relapse-free survival
(P=0.009) (37). Studies (37,38)
have also found a correlation between the loss of stromal Cav-1 and
upregulation of Akt phosphorylation, suggesting that the loss of
Cav-1 in the tumor microenvironment contributes to the metastatic
behavior of tumor cells by a mechanism that involves the
upregulation of transforming growth factor (TGF)-β1 and SNCG
through Akt activation. In malignant melanoma, the positive
correlation between the loss of stromal Cav-1 and poor overall
survival rate has been clarified by Wu et al (39). The authors identified that a low
stromal Cav-1 expression correlates with shorter survival when
compared with the high stromal Cav-1 expression group (median
survival, 252 days vs. 3,508 days, respectively; P=0.0054).
Conceivable mechanisms
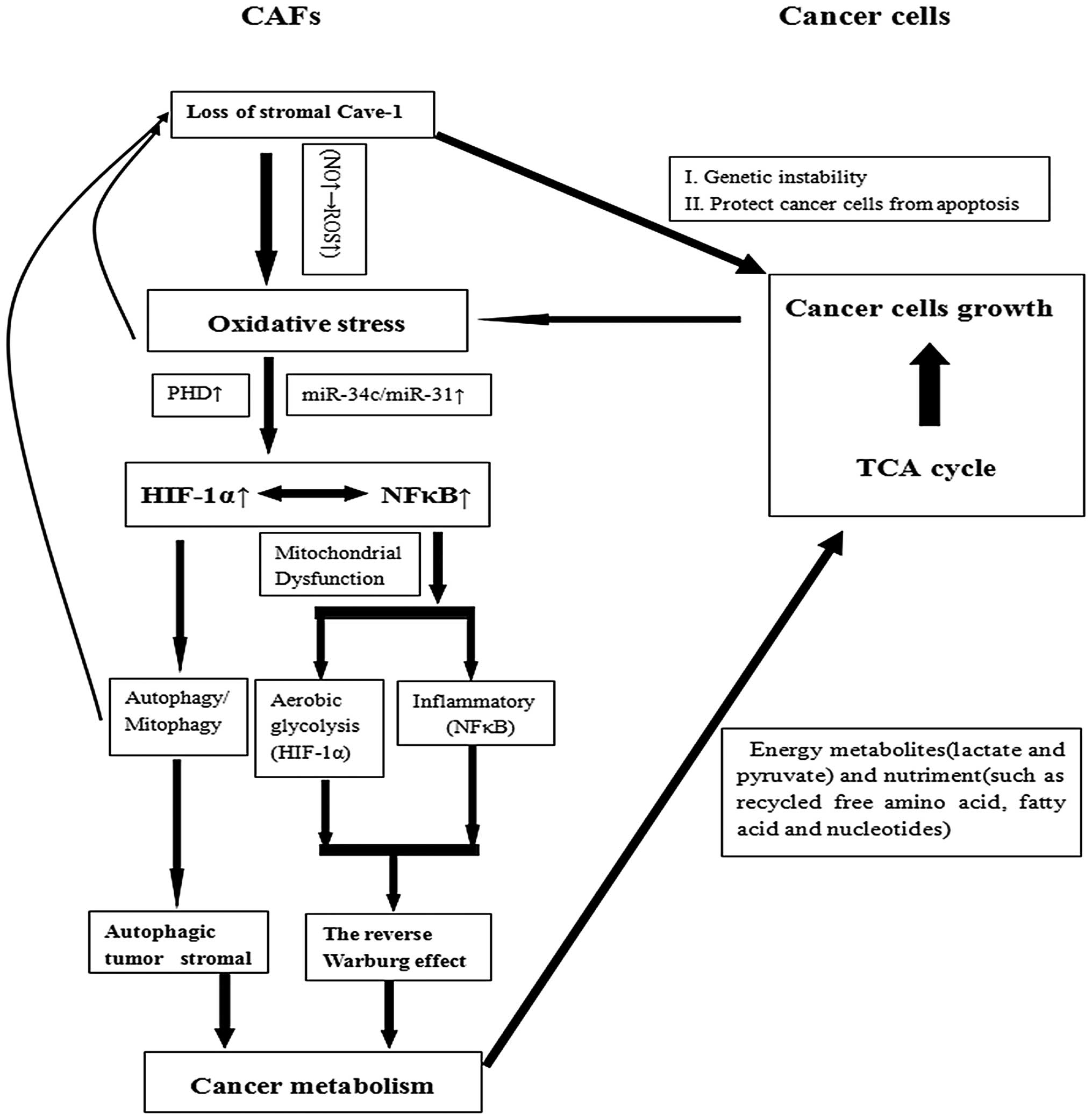
Cav-1-deficient CAFs may predict tumor prognosis
(Fig. 1). Numerous studies
(28,29,40,41)
support that the deregulation of stromal Cav-1 serves as a
functional marker of CAF phenotype. Sotgia et al (29) established a Cav-1(−/−) MSF cell line
that shares numerous characteristics with human CAFs, such as an
almost identical profile of RB/E2F-regulated genes that are
upregulated in human CAFs. The phenotype of the Cav-1-deficient
CAFs may be reversed by treatment with a Cav-1 mimetic peptide
(28). According to these previous
studies, we hypothesize at least three possible mechanisms of Cav-1
deregulation as follows. Firstly, the activation of oncogenes
(H-ras, v-abl, brc-abl and TGF) or inactivation of tumor suppress
genes (p53) may result in a loss of Cav-1 expression in CAFs in
culture (42). Secondly, similar to
the activation of fibroblasts in wound healing, the activation of
the TGF-β signaling pathway may also result in the Cav-1
deregulation of CAFs. Furthermore, CAFs have been shown to secrete
a number of growth factors, including TGFβ (43). Finally, surrounding cancer cells may
downregulate Cav-1 in adjacent NFs via oxidative stress to the
tumor microenvironment (40).
Martinez-Outschoorn et al (44) observed that the coculture of
immortalized human fibroblasts or primary cultures of normal human
fibroblasts with a human breast cancer cell line (MCF7) leads to
Cav-1 downregulation in fibroblasts, acquiring a CAF phenotype. In
addition, the transcription levels of Cav-1 in CAFs have been found
to increase by 2.3- to 2.4-fold or remain unchanged (28), suggesting that the downregulation of
stromal Cav-1 occurs at the post-transcriptional or -translational
level. In accordance with similar studies, we suggest that the
prognostic value of the downregulation of the stromal Cav-1 is
predominantly associated with the metabolism of the tumor
microenvironment, which is carefully discussed in the present
review, including the autophagic tumor stroma model of cancer
metabolism and the reverse Warburg effect. In addition to the
induction of the hyperproliferative phenotype of CAFs through the
RB/E2F pathway.
Several studies have indicated that the decrease of
stromal Cav-1 is accompanied by the activation of the RB/E2F
pathway (28,33). Using the gene profiling method, the
authors identified 118 gene transcripts involved in cell cycle
control that were upregulated. Among them, 44 gene transcripts were
involved in the RB/E2F gene signature, associated with RB
functional inactivation. RB is normally hypophosphorylated in
quiescent or differentiated cells, and prevents the transcription
of genes essential for cell cycle progression by suppressing the
activity of the E2F family (45).
The downregulation of stromal Cav-1 upregulates the phosphorylation
of RB and releases the activity of E2F, increasing downstream
target molecules, such as proliferating cell nuclear antigen (PCNA)
and minichromosome maintenance protein (MCM7), which account for
the hyperproliferative phenotype of CAFs. PCNA is a transcription
factor which helps the DNA polymerase δ to bind to the DNA, while
MCM7 serves as an inhibitor of DNA replication when bound to
hypophosphorylated RB. In addition, certain reports have prompted
that RB is a downstream molecule of mTOR in adipocytes, prostate
and ovarian cancer cells (46–48).
Although other reports have found that mTOR is activated in Cav-1
knock out CAFs (49) and keloids
(50), the newly identified axis in
CAFs (Cav-1 to mTOR to RB) (49)
requires further clarification.
Autophagic tumor stroma model of cancer
metabolism
The theory of ‘the autophagic tumor stroma model of
cancer metabolism’ is a newly established model to understand the
prognostic value of the downregulation of stromal Cav-1.
Martinez-Outschoorn et al (51) initially demonstrated this new
paradigm, which further confirmed the ‘autophagy paradox’ that the
role of autophagy in tumorigenesis is controversial.
Mechanistically, the authors also demonstrated that the state of
oxidative stress in adjacent CAFs results in the
autophagic/lysosomal deregulation of stromal Cav-1 via elevated
hypoxia inducible factor (HIF)-1α and nuclear factor κB (NFκB)
(44,52). Therefore, a positive feedback exists
between oxidative stress and the loss of stromal Cav-1. Although
the detailed mechanisms in which the loss of stromal Cav-1 causes
oxidative stress remain undistinguishable, this review summarizes a
possible mechanism based on previous studies.
As a potent inhibitor of NOS, Cav-1 binds to and
inhibits NOS activity in NFs, thus dampening NO release in a toxic
manner (53). However, studies
(41,54) have found that NOS productions are
transcriptionally overexpressed in human tumor and Cav-1(−/−)
stromal cells. This indicates that the loss of stromal Cav-1 is
deprived of its ability to inhibit NOS activity and to induce the
overexpression of NO. Besides resulting in DNA damage, the
accumulation of NO also induces mitochondrial uncoupling and
increased reactive oxygen species (ROS). As the mitochondrial
respiratory chain is a major source of intracellular ROS. The
mitochondrial uncoupling induced by the overexpression of NO
results in the dysfunction of the mitochondrial respiratory chain
and largely increases the levels of ROS. Finally, oxidative stress
is generated with the upregulation of ROS. Concomitantly, it has
been identified that the subunits of the respiratory chain
complexes (complex I, IV and V) are significantly decreased in
Cav-1 knockdown fibroblasts, and ROS is markedly upregulated in
human telomerase reverse transcriptase (hTERT)-fibroblasts treated
with Cav-1 siRNA (52). Notably,
compared with the homotypic cultures of fibroblasts, ROS levels are
significantly increased in fibroblasts (immortalized with hTERT)
when cocultured with human breast cancer cells (MCF7). Taken
together, the downregulation of Cav-1 and ROS levels are under
‘positive feed-forward control’ in CAFs, where an accumulation of
ROS induces the downregulation of stromal Cav-1, which results in
the subsequent generation of ROS (52).
The state of oxidative stress in the tumor
microenvironment is triggered by lateral epithelial cancer cells
and sustained through positive feed-forward control with the
downregulation of stromal Cav-1. Studies have already demonstrated
stromal oxidative stress based on the methods of proteomic and/or
transcriptional gene profiling. Witkiewicz et al (40) identified the upregulation of 238
gene transcripts and the downregulation of 232 gene transcripts in
the Cav-1-deficient tumor stroma. The gene set enrichment analysis
illustrated that the upregulation of gene transcripts is associated
with myofibroblast differentiation, oxidative stress, mitochondrial
dysfunction and DNA damage. Trimmer et al (55) also identified the upregulation of 21
gene products associated with oxidative stress and hypoxia,
including glycolytic enzymes (LDHA and GAPDH), mitochondrial
components involved in ROS production, enzymes acting as
antioxidants (PRDX1, PRDX4 and TXNDC5) and factors that are
involved in oxidative stress-induced DNA repair (XRCC6BP1).
Pavlides et al (54)
provided evidence of stromal oxidative stress, having identified
~100 metabolites (the two most significant metabolites being
asymmetric dimethylarginine and 3-hydroxybutyrate), which were
associated with the onset of the oxidative stress phenotype, that
were elevated in Cav-1 (−/−) null mammary fat pads.
Studies have demonstrated that oxidative stress in
adjacent CAFs induced by epithelial cancer cells result in
autophagy/mitophagy in the tumor microenvironment (44,51,52,54).
Combined with the stromal oxidative stress, studies have also
identified the upregulation of numerous molecules in adjacent CAFs,
which have been specifically associated with autophagy/mitophagy,
as well as mitochondrial dysfunction (40,54,55).
In addition, a previous study observed that the expression of
autophagy markers is markedly elevated following acute Cav-1
knockdown in fibroblasts, including Beclin 1, BNIP3, BNIP3L, HIF-1α
and NFκB (44). This observation
demonstrated that the loss of stromal Cav-1 is sufficient to induce
autophagy/mitophagy in CAFs. Given that the downregulation of
stromal Cav-1 results in autophagy, which is induced by the
oxidative stress and hypoxia of the tumor microenvironment, this
indicates that a feed-forward mechanism exists in the interactive
association between Cav-1 and autophagy/mitophagy in CAFs.
Mechanically, studies have demonstrated that
oxidative stress drives autophagy/mitophagy via the meditated
induction of HIF-1α and NFκB activation in fibroblasts (44,51,56,57).
Therefore, HIF-1α, which is the main transcription factor mediating
the hypoxia response, promotes transcription of angiogenic factors
[such as vascular endothelial growth factor (VEGF)] and leads to
increased autophagy and glycolysis (58). The state of oxidative stress implies
the accumulation of ROS in CAFs. In addition, the activation of
HIF-1α and NFκB require the reduced activation of prolyl
hydroxylase domain-containing protein (PHD). The results of a
previous study (52) has shown that
Cav-1 knockdown decreases the levels of E1α, E1β and E2 subunits of
the PHD complex. Therefore, the reduced activity of PHD mediated by
increased ROS levels results in the reduced hydroxylation of
HIF-1α, leading to HIF-1α stabilization and activation (59–62).
Additionally, the transcriptional activation of HIF1a by miR-31 is
indirectly mediated by FIH-1 (factor inhibiting HIF), which is the
direct target of miR-31 (54).
Capparelli et al (57)
demonstrated that the activation of the TGFβ/CTGF pathway also
regulates the metabolism of CAFs via the elevation of HIF-1α.
Additionally, as a multimeric inducible transcription factor, the
activation of NFκB is determined by the IκB kinase (IκBK) activity,
which is controlled by oxygen-sensitive PHD. The activation of IκBK
induced by the downregulation of PHD meditates the deregulation of
inhibitor of κB (IκB) by phosphorylation. As NFκB subunits are
inhibited and sequestered in the cytoplasm by IκB, the deregulation
of IκB meditates the activation of NFκB. Furthermore, recent
evidence has demonstrated important cross-talk and the
interdependence of HIF-1α and NFκB signaling. Increased HIF-1α has
also been shown to promote NFκB activity (63). Conversely, NFκB acts as a
transcriptional factor of HIF-1α (64).
The reverse Warburg effect
The reverse Warburg effect is an additional model
that has been proposed to understand the Warburg effect in tumor
metabolism. The Warburg effect, known as aerobic glycolysis, was
first formulated by Warburg (65).
Warburg’s original study demonstrated the propensity of cancer
cells to take up high levels of glucose and to secrete lactate and
pyruvate (energy metabolites generated by aerobic glycolysis).
Furthermore, recent evidence has demonstrated that tumor stromal
fibroblasts also exhibit the Warburg effect and secrete energy
metabolites (including lactate and pyruvate). In addition, CAFs
directly feed cancer cells via a type of host-parasite association.
The state of oxidative stress in Cav-1-deficient tumor stroma
induced by adjacent tumor cells not only results in
autophagy/mitophagy and DNA damage, but also causes mitochondrial
dysfunction and aerobic glycolysis (the Warburg effect) (66,67),
which is important for cancer recurrence, lymph node metastasis and
tumor prognosis. Pavlides et al (66) also demonstrated a cause-effect
association between the downregulation of stromal Cav-1 and aerobic
glycolysis, the upregulation of the myofibroblast marker and eight
glycolytic enzymes (including the M2-isoform of pyruvate kinase, as
well as HIF target genes) via unbiased proteomic analysis and the
transcriptional profiling of Cav-1-deficient stromal cells. This
indicated that a loss of stromal Cav-1 may be a novel biomarker for
aerobic glycolysis (the Warburg effect) in the tumor
microenvironment.
To fully understand the mechanism of the reverse
Warburg effect, Pavlides et al (41) performed an unbiased informatics
analysis of the transcriptional profile of Cav-1(−/−)-deficient
mesenchymal stromal cells. The authors identified that
Cav-1-deficient stromal fibroblastic cells show a markedly reduced
mitochondrial reserve capacity and a mitochondrial defect in
Cav-1-deficient stromal cells which may drive oxidative stress,
leading to aerobic glycolysis (HIF-1α) and inflammation (NFκB) in
the tumor microenvironment. The authors also found that genes
associated with NOS production, complexes I and IV and the
generation of ROS were upregulated. However, western blot analysis
(52) showed a significant decrease
in the subunits of complexes (I, IV and V) in Cav-1 knockdown
fibroblasts. This contradiction may imply that the loss of
mitochondrial respiratory chain complexes occurs at the
post-transcriptional or -translational level, or that the
upregulation of associated genes is a compensatory response to
mitochondrial dysfunction. Concomitantly, fibroblast-MCF7
cocultures, where Cav-1 is downregulated in fibroblasts, show a
marked decrease in mitochondrial mass compared with monocultured
fibroblasts (52). Further study
demonstrated that 45 known HIF-target genes and 86 NFκB-target
genes were transcriptionally upregulated in Cav-1(−/−) stromal
cells (41). In addition, the
upregulation of 151 mitochondrial associated genes served as a
compensatory response to mitochondrial dysfunction in Cav-1(−/−)
stromal cells. Furthermore, animal experiments have demonstrated
that Cav-1-deficient mice suffer from a reduced mitochondrial
reserve capacity, and that the lethality of Cav-1 deficiency may be
rescued if Cav-1 null mice are fed glucose (68). Overall, we propose a possible
mechanism (Fig. 1) to summarize the
reverse Warburg effect in tumor stromal fibroblasts.
Several recent studies have markedly suggested that
mitochondrial activity and oxidative phosphorylation is sufficient
to promote tumor growth. In vitro study has shown that MCF7
(human breast cancer cells) cells exhibit extremely high levels of
mitochondrial staining when cocultured with Cav-1 null fibroblasts,
as compared with homotypic cultures of MCF7 cells (52). Furthermore, lactate administration
was found to significantly increase mitochondrial mass in MCF7
cells. This study demonstrated in vitro that CAFs undergoing
aerobic glycolysis generate and secrete lactate and pyruvate,
enhance the mitochondrial respiratory and TCA cycles, and promote
tumor growth. By contrast, loss of function mutations in the TCA
cycle gene, isocitrate dehydrogenase, are found to correlate with
an improved prognosis and survival, suggesting that inactivity of
TCA cycle enzymes does not favor tumor aggressiveness (69). The mitochondrial protein, p32, has
also been found to maintain high levels of mitochondrial oxidative
phosphorylation in human cancer cells and to sustain tumorigenicity
in vivo (70).
Overall, cancer cells trigger oxidative stress in
the tumor microenvironment and activate two pro-autophagic
promoters, HIF-1α and NFκB, in stromal CAFs. As a result, adjacent
stromal fibroblasts undergo autophagy and mitophagy, leading to the
autophagic loss of Cav-1 and mitochondrial dysfunction. A loss of
stromal Cav-1 aggravates oxidative stress and further promotes
autophagy and mitophagy. As a result, stromal aerobic glycolysis
and autophagy/mitophagy generate energy metabolites (lactate and
pyruvate) and building blocks (such as recycled free amino acid,
fatty acid and nucleotides), respectively, that directly utilize
adjacent cancer cells to sustain growth and maintain cell
viability.
3. The role of Cav-1 expression in human
cancer cells
Expression of Cav-1 in human cancer
cells
Cav-1 expression in human cancer cells is not
considered to conform with that in the tumor stroma. Therefore,
contradictory roles of Cav-1 expression in human cancer cells have
been reported. Certain studies insist that Cav-1 is downregulated
and serves as a tumor suppressor in breast cancer (71–73),
GC (74), hepatic cancer (75) and mucoepidermoid carcinoma (MEC) of
the salivary glands (76); while
other studies suggest that the expression levels of Cav-1 are
upregulated, consistent with advanced tumor stage, high
histological type and the metastasis of human cancer cells,
including esophagus (77,78), pancreatic (79), renal (80), prostate (81) and colorectal (82) cancer.
The current review of previous studies lead to the
recognition of a contradictory theory with regard to the expression
of Cav-1 in breast (71–73,83),
gastric (74,84), hepatic (75,85,86)
and oral (76,87) cancer. Sagara et al (73) examined the mRNA and protein
expression levels of Cav-1 in 162 cases of breast cancer and found
that the mRNA and protein expression levels of Cav-1 were
suppressed in breast cancer tissue compared with the corresponding
normal tissues. In addition, reduced Cav-1 was found to
significantly (P=0.041) correlate with tumor size, consistent with
other studies (71,72) However, Savage et al (83) questioned the tumor suppressive
effect of Cav-1 following the immunohistochemical analysis of Cav-1
expression levels in benign lesions, breast cancer precursors and
metaplastic breast carcinomas, in a cohort of 245 invasive breast
carcinomas, and a CAV1 gene amplification assessment of 25 cases.
Despite its variable intensity, Cav-1 was consistently expressed in
MECs of radial scar, sclerosing adenosis, columnar cell lesions and
ductal carcinoma in situ, and significantly associated with
the ‘basal-like’ immunophenotype, with shorter disease-free and
overall survival (83). In GC, a
study (84) found that the positive
staining of Cav-1 was higher in the advance GC group than in the
early GC group (P=0.037), whereas, the progressive downregulation
of Cav-1 in gastric epithelial cells was found to correlate with
gastric carcinogenesis (74).
Additionally, Yan et al (78) identified that the Cav-1 mRNA
expression in hepatitis B virus-related hepatocellular carcinoma
(HCC) cells was found to negatively correlate with the tumor size,
major venous invasion, single or multiple tumors, pTNM staging and
factors associated with the prognosis of HCC, inconsistent with
other studies (85,86). Given the conflicting information on
the expression of Cav-1, at least in breast cancer, GC, hepatic
cancer and oral cancer, further studies analyzing the expression of
Cav-1 in human cancer cells are warranted.
In addition, pancreatic (87), esophagus (77), renal (89) and oral (87) cancer have shown the downregulation
of Cav-1 in cancer cells compared with non-cancerous tissues. By
contrast, breast (83), ovarian
(90), hepatic (75) and lung (91,92)
cancer exhibit an upregulation of Cav-1 in cancer cells compared
with the non-cancerous tissues. This inconsistent phenomenon may be
associated with the cell type-related expression of Cav-1; however,
further experiments are required to demonstrate the mechanism of
the different Cav-1 expression trends in different types of
tissue.
Clinical value of Cav-1 expression in a
variety of human cancer types
Despite the contradictory views of the clinical role
of Cav-1 expression in several types of cancer, the upregulation of
Cav-1 in human cancer cells serves as a tumor promoter role in the
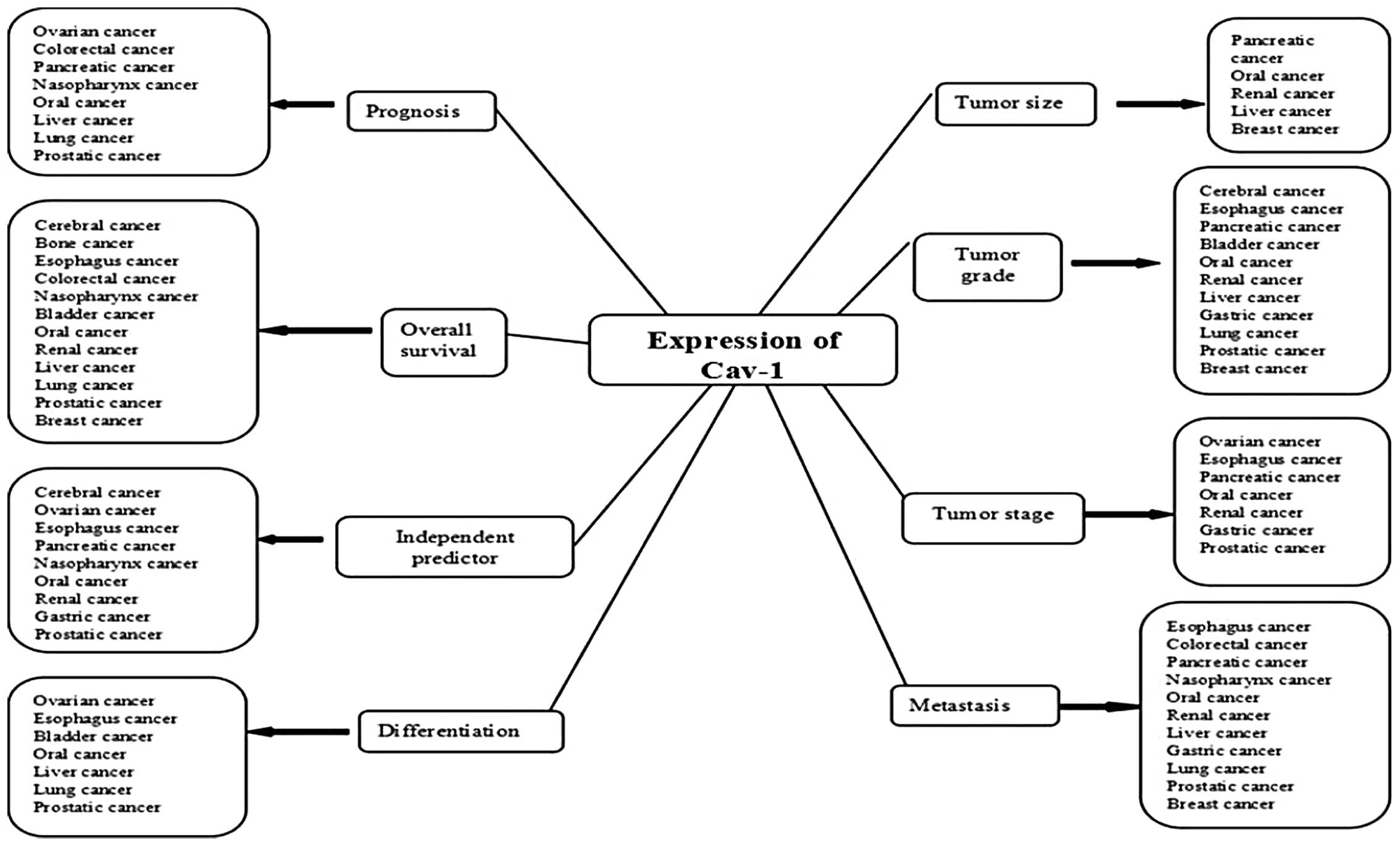
majority of human cancer types. The correlation between the
expression of Cav-1 in various types of cancer cells and clinical
characteristics, including tumor size, differentiation, tumor
grade, tumor stage, hematogenous or lymph node metastasis, tumor
prognosis and overall survival rate, has been clarified (Fig. 2).
In order to target the clinical value of Cav-1
expression in human cancer cells, this study reviewed the progress
of present clinical studies on several types of human cancer.
Pancreatic cancer
In pancreatic cancer, Cav-1 is frequently expressed
in the tumor tissue compared with the little or no staining
identified in chronic pancreatitis specimens, normal ductal
epithelium (88) and peritumoral
tissue (79,93). The prognostic significance of Cav-1
expression in pancreatic carcinoma was initially demonstrated by
Suzuoki et al (88). The
authors found 32 cases among 79 patients (40.5%) with pancreatic
adenocarcinoma showing positive Cav-1 immunostaining. Positive
Cav-1 expression was found to correlate with tumor diameter
(P=0.0079), histopathological grade (P=0.0272) and poor prognosis
(P=0.0008). Finally, the authors suggested that positive Cav-1
expression is an independent negative predictor of survival
(P=0.0358). In a recent study of 34 pancreatic ductal
adenocarcinoma (PDAC) tissue samples, Tanase et al (79) confirmed the prognostic role of the
expression of Cav-1. Furthermore, the expression of Cav-1 was found
to significantly correlate with Ki-67 and p53, as well as serum
levels of CA 19-9. Additionally, a series analyzing pancreatic
precancerous lesions (pancreatic intraepithelial neoplasia) and a
pancreatic cancer survey (93) also
indicated that Cav-1 may be a good candidate prognostic marker,
combined with the upregulation of fatty acid synthase.
Renal cancer
The clinical prognostic value of the upregulation of
Cav-1 in renal cell carcinoma (RCC) has been clarified. Campbell
et al (94) were the first
to interpret a correlation between the cytoplasmic expression of
Cav-1 and the outcome of RCC. The ICC scoring is determined as
follows: 0, no detectable deposit in tumor cells; 1, extremely
light diffuse or focal light deposit in tumor cell cytoplasm; 2,
light diffuse or moderate focal deposit (but may include small
areas of heavy deposit); and 3, tumor containing areas of heavy
deposit in tumor cells. Among 114 consecutive non-metastatic RCC
samples, 50 tumors exhibited ICC scores of 1, 43 of score 2 and 21
of score 3. Statistical analysis revealed that significantly higher
scores combined with larger and higher grade tumors, as well as
tumors with vascular invasion and Cav ICC scores are independent
predictors of poor disease-free survival (82,94,95).
Other study has also demonstrated that tumors with upregulated
Cav-1 exhibit a positive correlation with tumor diameter and tumor
grade/stage (pTNM and pM stages) (96,97).
Increased levels of cytoplasmic Cav-1 (P=0.037) have also been
clarified to correlate with hematogenous metastasis (98). Survival analysis has independently
shown that patients with tumors with increased Cav-1 staining
exhibit a shorter overall survival rate (99). Waalkes et al (89) initially confirmed that Cav-1 mRNA
expression is significantly increased in normal renal tissue
(P=0.0003), clear cell RCC (P=1.48×10−7) and advanced
disease (P=0.019), compared with patients with distant metastasis
at the time of diagnosis (P=0.0058).
Liver cancer
To date, the clinicopathological role of Cav-1
expression in HCC remains contradictory. Certain reports have
demonstrated that the expression of Cav-1 is markedly upregulated
in HCC patients (85,86) or cell lines (86). In addition, a marked increase of
Cav-1 expression has been identified in metastatic HCC cell lines
and tumors compared with normal liver cell lines and all
non-tumorous liver tissues. Following the analysis of a cohort of
HCC samples, Tang et al (85) identified a positive correlation
between the upregulation of tumor Cav-1 and the histological
differentiation, portal or hepatic venous invasion, intrahepatic
metastases and recurrence of HCC. In addition, Cav-1 expression has
been found to positively correlate with VEGF expression,
microvessel density, and unpaired artery (99). Furthermore, Tse et al
(86) identified that the
overexpression of Cav-1 promotes the growth, motility and
invasiveness, as well as tumorigenicity of HCC cells in
vivo. Similar findings have also been observed in metastatic
HCC cells with knockdown of Cav-1. By contrast, studies have
suggested that the upregulation of Cav-1 in HCC may serve as a
tumor suppressor (75,76). Yan et al (75) found that the expression levels of
Cav-1 in HCC tissues were significantly lower than those of the
adjacent non-cancerous tissues (P=0.026), and that the low
expression of Cav-1 is associated with a poor prognosis of HCC.
Lung cancer
A positive correlation exists between the
upregulation of Cav-1 and the clinical features of primary lung
cancer. Although Cav-1 levels in lung tumor tissues are
significantly lower than in tumor-free lung tissues (91,92,100–102), the expression of Cav-1 in lung
tumor tissues is markedly higher in patients with lymph node
metastasis (92,93,100)
and advanced tumor stage (93,100,103). Following the statistical analysis
of Cav-1 immunostaining and the clinical data of several primary
lung cancer cohorts, the expression of Cav-1 was demonstrated to
statistically correlate with poor differentiation, pathological
stage and lymph node metastasis, as well as a predicted poor
prognosis (104). Furthermore, a
multivariate analysis of the Cav-1 ICC results of 95 lung
adenocarcinoma specimens by Chao-Chi et al (101) suggested that Cav-1 is an
independent functional predictor of poor survival in lung
adenocarcinoma. In addition, Ho et al (105) identified that Cav-1 expression
significantly correlates with drug resistance and poor prognosis in
advanced non-small cell lung cancer (NSCLC) patients treated with
gemcitabine-based chemotherapy, by analyzing the immunostaining of
Cav-1 and the clinical response to the chemotherapy of 73 NSCLC
(stages IIIB and IV) patients.
PC
The correlation between the upregulation of Cav-1
and the clinical characteristics of PC has not been completely
clarified. However, a higher incidence of Cav-1 expression has
generally been found in patients with poorly differentiated tumors
(higher Gleason score), positive surgical margins, high tumor
stages (TNM T4), lymph node metastasis and poor tumor prognosis
(82,106–109). Satoh et al (107) further indicated that, in patients
with organ-confined (pT2N0) disease, the positive Cav-1 expression
was a significant predictor of disease recurrence following radical
prostatectomy. An ICC staining analysis of 189 radical
prostatectomy specimens (106)
identified positive Cav-1 immunostaining as an independent
predictor for time to disease progression (P=0.0186). Yang et
al (110) found that Cav-1 was
overexpressed in 41.7% (15 out of 36 patients) of human high-grade
prostatic intraepithelial neoplasia (HGPIN) specimens and further
revealed a highly significant correlation between Cav-1 (+) HGPIN
and Cav-1 (+) PC. Whereas Steiner et al (111) found that the number of caveolae
was significantly reduced in LNCaP and PC3 cells (P<0.0001),
which implied that the downregulation of Cav-1 occurs with the
development of PC, while the downregulation of Cav-1 in PC tissues
conversely correlates with pT category (P=0.006) and Gleason score
(P=0.041).
In addition, the serum Cav-1 levels of PC patients
have also been investigated. Certain studies have shown increased
serum Cav-1 levels in patients with poor prognosis (112,113). Langeberg et al (114) analyzed two case-control (n=1,458
and 1,351, respectively) studies of PC among males in Washington
State, USA; however, no correlation was identified between higher
post-treatment serum levels of Cav-1 and the risk of aggressive or
adverse PC outcome.
GC
The role of Cav-1 expression in GC requires further
clarification. The ICC study of 405 GC tissue specimens (84) revealed the upregulation of Cav-1
expression in the non-neoplastic gastric mucosa (not detectable)
compared with GC [shown in 22 (5.4%) out of 405 cases] tissue. In
addition, the upregulation of Cav-1 expression was found to
significantly correlate with advanced pTNM stage (P=0.027) and
lymph node metastasis (P=0.018). Furthermore, survival analysis
showed that Cav-1 expression is an independent prognostic factor of
poor survival (P=0.028). However, Gao et al (74) analyzed the expression of Cav-1 in 56
GC, 29 non-cancerous mucosa, 11 intestinal metaplasia and seven
atypical hyperplasia specimens. The authors concluded a reverse
expression trend of Cav-1; the positive rate of Cav-1 was
significantly lower in GC than in non-cancerous mucosa, intestinal
metaplasia and atypical hyperplasia (17.9 vs. 84.8, 81.8 and 57.1%,
respectively; P<0.05). The decreased expression of Cav-1 in GC
was found to significantly correlate with differentiation, advanced
GC and lymph node metastases. By contrast, Barresi et al
(115) demonstrated that the role
of Cav-1 in GC is not stage-specific or associated with prognosis,
following ICC analysis of the expression of Cav-1 in a series of
gastric carcinoma and the adjacent normal gastric mucosa.
Breast cancer
Previous studies have not reached a consensus
concerning the role of Cav-1 in human breast cancer. Certain
reports have insisted the tumor suppressive functions of Cav-1 by
knockout of the CAV1 gene in cells with a luminal phenotype
(116). In addition, Sagara et
al (73) quantitatively
examined the mRNA levels of CAV1 in 162 cases of breast cancer
using real-time polymerase chain reaction. Finally, it has also
been identified that reduced CAV1 mRNA levels significantly
correlate with increasing tumor size (P=0.041) and negative
estrogen receptor (ER) status (P=0.021), even though no significant
correlation has been identified with disease-free survival
(P=0.520). By contrast, other studies (72,83)
have identified a positive correlation between the expression of
Cav-1, and high histological grade and lack of steroid hormone
receptor positivity [ER and progesterone receptor (PR)], as well as
the expression of basal markers (basal cytokeratins, p63 and
P-cadherin). Furthermore, Joshi et al (82) identified an independent prognostic
role of Cav-1 expression in human breast cancer, by the
multivariate analysis (Cox regression model) of the Cav-1
immunostaining.
Other types of cancer
The clinical value of Cav-1 expression in other
types of cancer, including bladder, nasopharynx, oral (76,87),
colorectal, esophagus, ovarian (90), bone (117) and cerebral (118) cancer, have also been reported. The
ICC analysis of several cohorts of esophageal squamous cell
carcinoma samples (range, 47–130 samples) (77,78),
has identified that positive Cav-1 immunostaining positively
correlates with pathological stage (pT, pN and pM stages) and
lymphatic or vein invasion, and predicts a significantly shorter
overall survival rate. Notably, no significant correlation has been
identified between CAV1 mRNA expression and clinicopathological
factors (77). Ruan et al
(119) statistically analyzed the
positive expression rates of Cav-1 in primary and recurrent bladder
transitional cell carcinoma (BTCC), and the tumor-free survival
times in groups with and without Cav-1 expression. The authors also
found that the positive expression of Cav-1 predicts a higher
recurrence risk of BTCC and shows a lower disease-free survival
rate (120). The role of the
upregulation of Cav-1 in nasopharyngeal carcinoma (NPC) has been
classified by Du et al (121). The Cav-1 expression levels were
found to significantly correlate with metastasis (P=0.025), a lower
five-year survival rate (P=0.02) and local recurrence (P=0.038).
Multivariate Cox regression analysis indicated that the combination
of high Cav-1 and CD147 expression is a significant, independent
prognostic predictor in patients with NPC (hazard ratio=2.135;
P=0.006). Survival analysis of the Cav-1 expression in colon cancer
(120 samples) and rectal cancer (131 samples) patients (82) has also identified that Cav-1
expression significantly correlates with distant metastasis in
colon cancer and decreased disease-free survival (P=0.005) in
rectal cancer. In addition, Rödel et al (122) demonstrated that local control
rates at five years for patients with tumors showing low Cav-1
expression were significantly improved than for patients with high
Cav-1 expression carcinoma cells.
Possible mechanisms associated with the
clinical values of tumor cell Cav-1 expression
The role of the tumor Cav-1 gene
(CAV1)
Contrasting functions of Cav-1 have been
demonstrated; a tumor suppressor function and an oncogenic role.
Firstly, several epidemiological studies have revealed a
correlation between the Cav-1 gene and the risk of several types of
cancer. In addition, a number of case-control studies have revealed
a correlation between the polymorphism of Cav-1 (CAV1) T29107A
(rs7804372) and the risk of PC (113,123) and NPC (124). These studies have independently
obtained parallel results, in which a significant difference exists
between PC or NPC and the control groups in the distributions of
their genotypes and allelic frequencies in the CAV1 T29107A
(rs7804372) polymorphism. However, no significant correlations have
been identified between this polymorphism and the
clinicopathological characteristics which have been declassified
(113). More recently, studies
have concentrated on the gene expression of Cav-1 in cancer cells.
Syeed et al (125)
investigated 130 breast cancer samples and demonstrated that the
gene encoding Cav-1 is associated with the development and
progression of breast cancer. Furthermore, the authors revealed
that promoter hypermethylation and the loss of expression of the
CAV-1 gene is an important alternative mechanism for the
inactivation of CAV-1 leading to complete gene silencing (125). In addition, an animal study has
identified that low Cav-1 expression is associated with increased
cell proliferation, and ERα expression and reduced apoptosis
(126).
Metastasis
The role of Cav-1 in cell migration is
controversial. Evidence is available indicating that Cav-1 promotes
migration in a variety of cells, including fibroblasts, endothelial
cells and tumor-derived cell lines. Alternatively, the inhibition
of migration has been observed in endothelial, pancreatic carcinoma
and metastatic breast cancer cells. In pancreatic cancer cells, the
Rho protein (RhoC) has a promoting role in tumor metastasis and
growth. Lin et al (127)
demonstrated that high Cav-1 expression may regulate RhoC activity,
thus limiting cell migration and promoting growth. In addition, a
reciprocal correlation has been identified between Cav-1 expression
and p42/p44 Erk activation with PC cell migration, invasion, RhoC
GTPase and p38 MAPK activation. Thomas et al (128) further demonstrated the phenocopy
effect of Cav-1 depletion and the reduced UMUC-3 lung metastasis of
bladder cancer in vivo, by treatment with a ROCK inhibitor.
Arpaia et al (129) also
demonstrated that the interaction between Cav-1 and Rho-GTPases
(most likely RhoC but not RhoA) promotes metastasis. By regulating
the overexpression of an activated form of Stat3, Chiu et al
(130) revealed that the Cav-1
promoter activity and gene expression were increased, preventing
the formation of brain metastases. Furthermore, the pathological
analysis of a cohort of head and neck squamous cell carcinoma
patients suggested that Cav-1 may have an inhibitory function in
tumorigenesis and lung metastasis by regulating integrin β1- and
Src-mediated cell-cell and cell-matrix interactions (131).
Motility and focal adhesion (FA)
The theory that Cav-1 promotes the motility of
tumor cells is well established. Following the transfection of a
wild-type CVA1 gene, an NSCLC cell line was found to exhibit an
enlarged cell shape with filopodia (132). Cav-1 and Rho/ROCK signaling is
known to promote the migration and metastasis of tumor cells by
regulating FA dynamics through the tyrosine (Y14) phosphorylation
of Cav-1. Joshi et al (82)
further defined a feedback loop between Rho/ROCK, Src and
phosphorylated Cav-1 in tumor cell protrusions. The authors
demonstrated that phosphorylated Cav-1 expression stimulates Rho
activation, stabilizes FAK association with FAs, and promotes cell
migration and invasion. However, increased levels of phosphorylated
Cav-1 were also associated with elevated Src kinase and Rho/ROCK
signaling. The Src family of kinase inhibitors can also reduce
Cav-1 phosphorylation on tyrosine-14 and cell migration in
vitro (133). The Rh/ROCK
signaling pathway has also been identified in pancreatic
adenocarcinoma cells. Mark et al (134) were the first to demonstrate that
FA is dependent on c-Src kinase activation, for which Cav-1 is
required, in the CEACAM6-overexpressing PDAC cell line, BxPC3.
However, Cantiani et al (116) found that c-Src and c-Met tyrosine
kinases are activated in osteosarcoma and inhibited with Cav-1
overexpression.
Antiapoptosis
Meyer et al (135) identified that Cav-1 is a crucial
hepatocyte fate determinant for TGF-β effects. The knockdown of
Cav-1 was found to markedly reduce TGF-β-mediated AKT
phosphorylation and, thus, sensitized primary murine hepatocytes
for proapoptotic TGF-β signaling. In further study of the
androgen-independent PC DU145 cell line, the colocalization of the
α1A-adrenoceptor with Cav-1 was observed by electron
microscopy (136). These results
showed that the agonist stimulation of the
α1A-adrenoceptor induces resistance to
thapsigargin-induced apoptosis and that Cav-1 (caveolae integrity)
was necessary for this process. By contrast, Rodriguez et al
(137) found that augmented Cav-1
expression in cells with low basal levels of proteins, such as
COX-2 and PGE2, and COX-2 overexpression or PGE2 supplementation,
increases the levels of the inhibitor of apoptosis protein,
survivin, by a transcriptional mechanism. In a study on human colon
and PC cells, it was suggested that Cav-1 regulates the sensitivity
to β-carotene growth-inhibitory and proapoptotic effects (138). The authors found that β-carotene
functions as a growth inhibitory agent in Cav-1(+) cells, and that
the transfection of Cav-1 in Cav-1(−) cells increases cell
sensitivity to β-carotene by inducing apoptosis.
4. Conclusion
As a main structural component of caveolae, which
are plasma membrane invaginations that are involved in vesicular
trafficking and signal transduction events, Cav-1 is important in
the modulation of cellular signaling. Advances in understanding the
contribution of Cav-1 in cancer progression and the clinical
characteristics from stromal and cancer cells are likely to enhance
the awareness and acknowledgement of the reciprocal signaling that
supports and promotes oncogenesis, tumor differentiation, tumor
stage, metastasis and survival. Revealing the essential biological
and pathological mechanisms involved has realized the requirement
for Cav-1-specific therapeutic strategies.
It is already clear that the expression state of
stromal Cav-1 is coincidently downregulated in various types of
human cancer, including breast cancer, compared with non-cancerous
tissues (Table I), and the
mechanisms and clinical role of the deregulation of Cav-1 have been
sufficiently demonstrated. Future studies are required to varify
the role of Cav-1 in other types of CAFs. However, the expression,
clinical roles and associated mechanisms of tumor Cav-1 expression
are upregulated or decreased based on different cancer types or
different experiments of a same cancer type. Cav-1 may have an
oncogenic or tumor suppressor role depending on the cell type;
however, further investigation of Cav-1 expression and the possible
underlying mechanisms are required. In addition, it must be
determined whether correlations exist between Cav-1 expression and
tumor stromal and cancer cells, and the mechanisms understood.
 | Table IComparison of Cav-1 expression
between tumor stromal and human tumor cells in the literature. |
Table I
Comparison of Cav-1 expression
between tumor stromal and human tumor cells in the literature.
| Expression of
Cav-1, na | |
|---|
|
| |
|---|
| Tumor
components | Upregulation | Downregulation | Total reference
counts, n |
|---|
| Tumor cells | 53 | 10 | 63 |
| Stromal cells | 0 | 15 | 15 |
| Total | 53 | 25 | 78 |
Despite a number of contradictory Cav-1 studies,
the majority of reports markedly suggest that Cav-1 represents an
important cancer cell biomarker in carcinogenesis, differentiation,
metastasis and tumor progression, and independently serves as a
predictor of overall survival rate. In addition, through
interaction with other biological molecules, Cav-1 modulates
angiogenesis and correlates with chemotherapeutic resistance. To
succeed in establishing novel diagnostic molecular and targeted
therapies against Cav-1, high-quality, basic and translational
studies are required to further unveil the clinical value of Cav-1
expression in multiple types of cancer and tumor stromal cells.
References
|
1
|
Palade GE: Fine structure of blood
capillaries. J Appl Phys. 24:1424–1436. 1953.
|
|
2
|
Glenney JR Jr and Zokas L: Novel tyrosine
kinase substrates from Rous sarcoma virus-transformed cells are
present in the membrane skeleton. J Cell Biol. 108:2401–2408.
1989.
|
|
3
|
Sowa G, Pypaert M, Fulton D and Sessa WC:
The phosphorylation of caveolin-2 on serines 23 and 36 modulates
caveolin-1-dependent caveolae formation. Proc Natl Acad Sci USA.
100:6511–6516. 2003.
|
|
4
|
Song KS, Scherer PE, Tang Z, et al:
Expression of caveolin-3 in skeletal, cardiac, and smooth muscle
cells. Caveolin-3 is a component of the sarcolemma and
co-fractionates with dystrophin and dystrophin-associated
glycoproteins. J Biol Chem. 271:15160–15165. 1996.
|
|
5
|
Razani B, Combs TP, Wang XB, et al:
Caveolin-1-deficient mice are lean, resistant to diet-induced
obesity, and show hypertriglyceridemia with adipocyte
abnormalities. J Biol Chem. 277:8635–8647. 2002.
|
|
6
|
Rothberg KG, Heuser JE, Donzell WC, et al:
Caveolin, a protein component of caveolae membrane coats. Cell.
68:673–682. 1992.
|
|
7
|
Engelman JA, Zhang XL and Lisanti MP:
Sequence and detailed organization of the human caveolin-1 and -2
genes located near the D7S522 locus (7q31.1). Methylation of a CpG
island in the 5′ promoter region of the caveolin-1 gene in human
breast cancer cell lines. FEBS Lett. 448:221–230. 1999.
|
|
8
|
Kurzchalia T, Dupree P, Parton RG, et al:
VIP 21, A 21-kD membrane protein is an integral component of
trans-Golgi-network-derived transport vesicles. J Cell Biol.
118:1003–1014. 1992.
|
|
9
|
Glenney JR: The sequence of human caveolin
reveals identity with VIP 21, a component of transport vesicles.
FEBS Lett. 314:45–48. 1992.
|
|
10
|
Monier S, Parton RG, Vogel F, et al:
VIP21-caveolin, a membrane protein constituent of the caveolar
coat, oligomerizes in vivo and in vitro. Mol Biol
Cell. 6:911–927. 1995.
|
|
11
|
Arbuzova A, Wang L, Wang J, et al:
Membrane binding of peptides containing both basic and aromatic
residues. Experimental studies with peptides corresponding to the
scaffolding region of caveolin and the effector region of MARCKS.
Biochemistry. 39:10330–10339. 2000.
|
|
12
|
Luetterforst R, Stang E, Zorzi N, et al:
Molecular characterization of caveolin association with the Golgi
complex: identification of a cis-Golgi targeting domain in the
caveolin molecule. J Cell Biol. 145:1443–1459. 1999.
|
|
13
|
Schlegel A and Lisanti MP: A molecular
dissection of caveolin-1 membrane attachment and oligomerization.
Two separate regions of the caveolin-1 C-terminal domain mediate
membrane binding and oligomer/oligomer interactions in vivo.
J Biol Chem. 275:21605–21617. 2000.
|
|
14
|
Schlegel A, Schwab R, Scherer PE, et al: A
role for the caveolin scaffolding domain in mediating the membrane
attachment of caveolin-1. The caveolin scaffolding domain is both
necessary and sufficient for membrane binding in vitro. J
Biol Chem. 274:22660–22667. 1999.
|
|
15
|
Sargiacomo M, Scherer PE, Tang ZL, et al:
Oligomeric structure of caveolin: implications for caveolae
membrane organization. Proc Natl Acad Sci USA. 92:9407–9411.
1995.
|
|
16
|
Couet J, Li S, Okamoto T, et al:
Identification of peptide and protein ligands for the
caveolin-scaffolding domain. Implications for the interaction of
caveolin with caveolaeassociated proteins. J Biol Chem.
272:6525–6533. 1997.
|
|
17
|
Dvorak HF, Weaver VM, Tlsty TD, et al:
Tumor microenvironment and progression. J Surg Oncol. 103:468–474.
2011.
|
|
18
|
Chun TH, Hotary, et al: A pericellular
collagenase directs the 3-dimensional development of while adipose
tissue. Cell. 125:577–591. 2006.
|
|
19
|
Kan S, Konishi E, Arita T, et al:
Podoplanin expression in cancer-associated fibroblasts predicts
aggressive behavior in melanoma. J Cutan Pathol. Mar 3–2014.(Epub
ahead of print).
|
|
20
|
Zhou B, Chen WL, Wang YY, et al: A role
for cancer-associated fibroblasts in inducing the
epithelial-to-mesenchymal transition in human tongue squamous cell
carcinoma. J Oral Pathol Med. Mar 20–2014.(Epub ahead of
print).
|
|
21
|
Yu Y, Lee JS, Xie N, et al: Prostate
stromal cells express the progesterone receptor to control cancer
cell mobility. PLoS One. 9:e927142014.
|
|
22
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006.
|
|
23
|
De Wever O, Demetter P, Mareel M and
Bracke M: Stromal myofibroblasts are drivers of invasive cancer
growth. Int J Cancer. 123:2229–2238. 2008.
|
|
24
|
Sugimoto H, Mundel TM, Kieran MW and
Kalluri R: Identification of fibroblast heterogeneity in the tumor
microenvironment. Cancer Biol Ther. 5:1640–1646. 2006.
|
|
25
|
Erez N, Truitt M, Olson P, et al:
Cancer-associated fibroblasts are activated in incipient neoplasia
to orchestrate tumor-promoting inflammation in an
NF-kappaB-dependent manner. Cancer Cell. 17:135–147. 2010.
|
|
26
|
Ostman A: PDGF receptors-mediators of
autocrine tumor growth and regulators of tumor vasculature and
stroma. Cytokine Growth Factor Rev. 15:275–286. 2004.
|
|
27
|
Koleske AJ, Baltimore D and Lisanti MP:
Reduction of caveolin and caveolae in oncogenically transformed
cells. Proc Natl Acad Sci USA. 92:1381–1385. 1995.
|
|
28
|
Mercier I, Casimiro MC, Wang C, et al:
Human breast cancer-associated fibroblasts (CAFs) show caveolin-1
downregulation and RB tumor suppressor functional inactivation.
Cancer Biol Ther. 7:1212–1225. 2008.
|
|
29
|
Sotgia F, Del Galdo F, Casimiro MC, et al:
Caveolin-1−/− null mammary stromal fibroblasts share
characteristics with human breast cancer-associated fibroblasts. Am
J Pathol. 174:746–761. 2009.
|
|
30
|
Sloan EK, Ciocca DR, Pouliot N, et al:
Stromal cell expression of caveolin-1 predicts outcome in breast
cancer. Am J Pathol. 174:2053–2043. 2009.
|
|
31
|
El-Gendi SM, Mostafa MF and El-Gendi AM:
Stromal caveolin-1 expression in breast carcinoma. Correlation with
early tumor recurrence and clinical outcome. Pathol Oncol Res.
18:459–69. 2012.
|
|
32
|
Witkiewicz AZ, Dasgupta A, Sammons S, et
al: Loss of stromal caveolin-1 expression predicts poor clinical
outcome in triple negative and basal-like breast cancers. Cancer
Biol Ther. 10:135–143. 2010.
|
|
33
|
Witkiewicz AK, Dasgupta A, Sotgia F, et
al: An absence of stromal caveolin-1 expression predicts early
tumor recurrence and poor clinical outcome in human breast cancers.
Am J Pathol. 74:2023–2034. 2009.
|
|
34
|
Simpkins SA, Hanby AM, Holliday DL, et al:
Clinical and functional significance of loss of caveolin-1
expression in breast cancer-associated fibroblasts. J Pathol.
227:490–498. 2012.
|
|
35
|
Zhao X, He Y, Gao J, et al: Caveolin-1
expression level in cancer associated fibroblasts predicts outcome
in gastric cancer. PLoS One. 8:e591022013.
|
|
36
|
He Y, Zhao X, Gao J, et al: Quantum
dots-based immunofluorescent imaging of stromal fibroblasts
caveolin-1 and light chain 3B expression and identification of
their clinical significance in human gastric cancer. Int J Mol Sci.
13:13764–13780. 2012.
|
|
37
|
Ayala G, Morello M, Frolov A, et al: Loss
of caveolin-1 in prostate cancer stroma correlates with reduced
relapse-free survival and is functionally relevant to tumour
progression. J Pathol. 23:77–87. 2013.
|
|
38
|
Di Vizio D, Morello M, Sotgia F, et al: An
absence of stromal caveolin-1 is associated with advanced prostate
cancer, metastatic disease and epithelial Akt activation. Cell
Cycle. 8:2420–2424. 2009.
|
|
39
|
Wu KN, Queenan M, Brody JR, et al: Loss of
stromal caveolin-1 expression in malignant melanoma metastases
predicts poor survival. Cell Cycle. 10:4250–5. 2011.
|
|
40
|
Witkiewicz AK, Kline J, Queenan M, et al:
Molecular profiling of a lethal tumor microenvironment, as defined
by stromal caveolin-1 status in breast cancers. Cell Cycle.
10:1794–1809. 2011.
|
|
41
|
Pavlides S, Tsirigos A, Vera I, et al:
Loss of stromal caveolin-1 leads to oxidative stress, mimics
hypoxia and drives inflammation in the tumor microenvironment,
conferring the ‘reverse Warburg effect’: a transcriptional
informatics analysis with validation. Cell Cycle. 9:2201–2219.
2010.
|
|
42
|
Bist A, Fielding CJ and Fielding PE: p53
regulates caveolin gene transcription, cell cholesterol, and growth
by a novel mechanism. Biochemistry. 39:1966–1972. 2000.
|
|
43
|
Mueller MM and Fusenig NE: Friends or
foes-bipolar effects of the tumour stroma in cancer. Nat Rev
Cancer. 4:839–849. 2004.
|
|
44
|
Martinez-Outschoorn UE, Trimmer C, Lin Z,
et al: Autophagy in cancer associated fibroblasts promotes tumor
cell survival: Role of hypoxia, HIF1 induction and NFκB. Cell
Cycle. 9:3515–3533. 2010.
|
|
45
|
Bartek J, Bartkova J and Lukas J: The
retinoblastoma protein pathway and the restriction point. Curr Opin
Cell Biol. 8:805–14. 1996.
|
|
46
|
Usui I, Haruta T, Iwata M, et al:
Retinoblastoma protein phosphorylation via PI 3-kinase and mTOR
pathway regulates adipocyte differentiation. Biochem Biophys Res
Commun. 275:115–120. 2000.
|
|
47
|
Gao N, Flynn DC, Zhang Z, et al: G1 cell
cycle progression and the expression of G1 cyclins are regulated by
PI3K/AKT/mTOR/p70S6K1 signaling in human ovarian cancer cells. Am J
Physiol Cell Physiol. 287:C281–C291. 2004.
|
|
48
|
Xu Y, Chen SY, Ross KN, et al: Androgens
induce prostate cancer cell proliferation through mammalian target
of rapamycin activation and post-transcriptional increases in
cyclin D proteins. Cancer Res. 66:7783–7792. 2006.
|
|
49
|
Mercier I, Camacho J, Titchen K, et al:
Caveolin-1 and accelerated host aging in the breast tumor
microenvironment: chemoprevention with rapamycin, an mTOR inhibitor
anti-aging drug. Am J Path. 181:278–292. 2012.
|
|
50
|
Ong CT, Khoo YT, Mukhopadhyay A, et al:
mTOR as a potential therapeutic target for treatment of keloids and
excessive scars. Exp Dermatol. 16:394–404. 2007.
|
|
51
|
Martinez-Outschoorn UE, Whitaker-Menezes
D, Pavlides S, et al: The autophagic tumor stroma model of cancer
or ‘battery-operated tumor growth’: A simple solution to the
autophagy paradox. Cell Cycle. 9:4297–4306. 2010.
|
|
52
|
Martinez-Outschoorn UE, Balliet RM,
Rivadeneira DB, et al: Oxidative stress in cancer associated
fibroblasts drives tumor-stroma co-evolution: A new paradigm for
understanding tumor metabolism, the field effect and genomic
instability in cancer cells. Cell Cycle. 9:3256–3276. 2010.
|
|
53
|
Garcia-Cardena G, Martasek P, Masters BS,
et al: Dissecting the interaction between nitric oxide synthase
(NOS) and caveolin. Functional significance of the nos caveolin
binding domain in vivo. J Biol Chem. 272:25437–40. 1997.
|
|
54
|
Pavlides S, Tsirigos A, Migneco G, et al:
The autophagic tumor stroma model of cancer: Role of oxidative
stress and ketone production in feuling tumor cell metabolism. Cell
Cycle. 9:3485–3505. 2010.
|
|
55
|
Trimmer C, Sotgia F, Whitaker-Menezes D,
et al: Caveolin-1 and mitochondrial SOD2 (MnSOD) function as tumor
suppressors in the stromal microenvironment: a new genetically
tractable model for human cancer associated fibroblasts. Cancer
Biol Ther. 11:383–394. 2011.
|
|
56
|
Chiavarina B, Whitaker-Menezes D, Migneco
F, et al: HIF1-alpha functions as a tumor promoter in cancer
associated fibroblasts, and as a tumor suppressor in breast cancer
cells: Autophagy drives compartment-specific oncogenesis. Cell
Cycle. 9:3534–3551. 2010.
|
|
57
|
Capparelli C, Whitaker-Menezes D, Guido C,
et al: CTGF drives autophagy, glycolysis and senescence in
cancer-associated fibroblasts via HIF1 activation, metabolically
promoting tumor growth. Cell Cycle. 11:2272–2284. 2012.
|
|
58
|
Papandreou I, Cairns RA, Fontana L, et al:
HIF-1 mediates adaptation to hypoxia by actively downregulating
mitochondrial oxygen consumption. Cell Metab. 3:187–97. 2006.
|
|
59
|
Salceda S and Caro J: Hypoxia-inducible
factor 1alpha (HIF-1alpha) protein is rapidly degraded by the
ubiquitin-proteasome system under normoxic conditions. Its
stabilization by hypoxia depends on redox-induced changes. J Biol
Chem. 272:22642–22647. 1997.
|
|
60
|
Guzy RD, Hoyos B, Robin E, et al:
Mitochondrial complex III is required for hypoxia-induced ROS
production and cellular oxygen sensing. Cell Metab. 1:401–8.
2005.
|
|
61
|
Schofield CJ and Ratcliffe PJ: Signalling
hypoxia by HIF hydroxylases. Biochem Biophys Res Commun.
338:617–626. 2005.
|
|
62
|
Valko M, Rhodes CJ, Moncol J, et al: Free
radicals, metals and antioxidants in oxidative stress-induced
cancer. Chem Biol Interact. 160:1–40. 2006.
|
|
63
|
Pantuck AJ, An J, Liu H, et al:
NFkappaBdependent plasticity of the epithelial to mesenchymal
transition induced by Von Hippel-Lindau inactivation in renal cell
carcinomas. Cancer Res. 70:752–61. 2010.
|
|
64
|
Jung Y, Isaacs JS, Lee S, et al:
Hypoxia-inducible factor induction by tumour necrosis factor in
normoxic cells requires receptor-interacting protein-dependent
nuclear factor kappaB activation. Biochem J. 370:1011–1017.
2003.
|
|
65
|
Robinson JM and Gibbs M: Photosynthetic
intermediates, the warburg effect, and glycolate synthesis in
isolated spinach chloroplasts. Plant Physiol. 53:790–797. 1974.
|
|
66
|
Pavlides S, Whitaker-Menezes D,
Castello-Cros R, et al: The reverse Warburg effect Aerobic
glycolysis in cancer associated fibroblasts and the tumor stroma.
Cell Cycle. 8:3984–4001. 2009.
|
|
67
|
Witkiewicz AK, Whitaker-Menezes D,
Dasgupta A, et al: Using the ‘reverse Warburg effect’ to identify
high-risk breast cancer patients: stromal MCT4 predicts poor
clinical outcome in triple-negative breast cancers. Cell Cycle.
11:1108–1117. 2012.
|
|
68
|
Fernandez MA, Albor C, Ingelmo-Torres M,
et al: Caveolin-1 is essential for liver regeneration. Science.
313:1628–1632. 2006.
|
|
69
|
Yan H, Parsons DW, Jin G, et al: IDH1 and
IDH2 mutations in gliomas. N Engl J Med. 360:765–773. 2009.
|
|
70
|
Fogal V, Richardson AD, Karmali PP, et al:
Mitochondrial p32 protein is a critical regulator of tumor
metabolism via maintenance of oxidative phosphorylation. Mol Cell
Biol. 30:1303–1318. 2010.
|
|
71
|
Elsheikh SE, Green AR, Rakha EA, et al:
Caveolin 1 and Caveolin 2 are associated with breast cancer
basal-like and triple-negative immunophenotype. Br J Cancer.
99:327–334. 2008.
|
|
72
|
Zuccari DAPC, Castro1 R, Gavioli AF, et
al: Immunohistochemical and molecular analysis of caveolin-1
expression in canine mammary tumors. Genet Mol Res. 11:153–165.
2012.
|
|
73
|
Sagara Y, Mimori K, Yoshinaga K, et al:
Clinical significance of Caveolin-1, Caveolin-2 and HER2/neu mRNA
expression in human breast cancer. Br J Cancer. 91:959–965.
2004.
|
|
74
|
Gao X, Sun Y, Huang L, et al:
Down-regulation of caveolin-1 in gastric carcinoma and its clinical
biological significance. Ai Zheng. 24:311–316. 2005.(In
Chinese).
|
|
75
|
Yan J, Lu Q, Cai L, et al: Expression of
caveolin-1 gene in hepatitis B-related hepatocellular carcinoma and
the clinical significance thereof. Zhonghua Yi Xue Za Zhi.
88:3272–3274. 2008.(In Chinese).
|
|
76
|
Shi L, Chen XM, Wang L, et al: Expression
of caveolin-1 in mucoepidermoid carcinoma of the salivary glands:
correlation with vascular endothelial growth factor, microvessel
density, and clinical outcome. Cancer. 109:1523–1531. 2007.
|
|
77
|
Ando T, Ishiguro H, Kimura M, et al: The
overexpression of caveolin-1 and caveolin-2 correlates with a poor
prognosis and tumor progression in esophageal squamous cell
carcinoma. Oncol Rep. 18:601–609. 2007.
|
|
78
|
Yan J, Lu Q, Cai L, et al: Expression of
caveolin-1 gene in hepatitis B-related hepatocellular carcinoma and
the clinical significance thereof. Zhonghua Yi Xue Za Zhi.
88:3272–3274. 2008.
|
|
79
|
Tanase CP, Dima S, Mihai M, et al:
Caveolin-1 overexpression correlates with tumour progression
markers in pancreatic ductal adenocarcinoma. J Mol Histol.
40:23–29. 2009.
|
|
80
|
Cho DS, Yim H, Cho KS, et al: Impact of
caveolin-1 expression on the prognosis of transitional cell
carcinoma of the upper urinary tract. J Korean Med Sci. 23:296–301.
2008.
|
|
81
|
Yang G, Timme TL, Frolov A, et al:
Combined c-Myc and caveolin-1 expression in human prostate
carcinoma predicts prostate carcinoma progression. Cancer.
103:1186–1194. 2005.
|
|
82
|
Joshi B, Strugnell SS, Goetz JG, et al:
Phosphorylated caveolin-1 regulates Rho/ROCK-dependent focal
adhesion dynamics and tumor cell migration and invasion. Cancer
Res. 68:8210–8220. 2008.
|
|
83
|
Savage K, Lambros MB, Robertson D, et al:
Caveolin 1 is overexpressed and amplified in a subset of basal-like
and metaplastic breast carcinomas: a morphologic ultrastructural,
immunohistochemical, and in situ hybridization analysis. Clin
Cancer Res. 13:90–101. 2007.
|
|
84
|
Nam KH, Lee BL, Park JH, et al: Caveolin 1
expression correlates with poor prognosis and focal adhesion kinase
expression ingastric cancer. Pathobiology. 80:87–94. 2013.
|
|
85
|
Tang Y, Zeng X, He F, et al: Caveolin-1 is
related to invasion, survival, and poor prognosis in hepatocellular
cancer. Med Oncol. 29:977–984. 2012.
|
|
86
|
Tse EY, Ko FC, Tung EK, et al: Caveolin-1
overexpression is associated with hepatocellular carcinoma
tumourigenesis and metastasis. J Pathol. 226:645–653. 2012.
|
|
87
|
Hung KF, Lin SC, Liu CJ, et al: The
biphasic differential expression of the cellular membrane protein,
caveolin-1, in oral carcinogenesis. J Oral Pathol Med. 32:461–467.
2003.
|
|
88
|
Suzuoki M, Miyamoto M, Kato K, et al:
Impact of caveolin-1 expression on prognosis of pancreatic ductal
adenocarcinoma. Br J Cancer. 87:1140–1144. 2002.
|
|
89
|
Waalkes S, Eggers H, Blasig H, et al:
Caveolin 1 mRNA is overexpressed in malignant renal tissue and
might serve as a novel diagnostic marker for renal cancer. Biomark
Med. 5:219–225. 2011.
|
|
90
|
Davidson B, Nesland JM, Goldberg I, et al:
Caveolin-1 expression in advanced-stage ovarian carcinoma-a
clinicopathologic study. Gynecol Oncol. 81:166–171. 2001.
|
|
91
|
Liu HX, Xing LX, Wang HB, et al:
Relationship between expression of caveolin-1 and pERK1/2 and
prognosis in non-small cell lung cancer. Zhonghua Bing Li Xue Za
Zhi. 37:615–619. 2008.(In Chinese).
|
|
92
|
Zhang YX, Chen HL, Ye B, et al: Study on
expression and methylation of caveolin 1 gene in non-small cell
lung cancers. Zhonghua Bing Li Xue Za Zhi. 37:300–304. 2008.(In
Chinese).
|
|
93
|
Witkiewicz AK, Nguyen KH, Dasgupta A, et
al: Co-expression of fatty acid synthase and caveolin-1 in
pancreatic ductal adenocarcinoma: implications for tumor
progression and clinical outcome. Cell Cycle. 7:3021–3025.
2008.
|
|
94
|
Campbell L, Gumbleton M and Griffiths DFR:
Caveolin-1 overexpression predicts poor disease-free survival of
patients with clinically confined renal cell carcinoma. Br J
Cancer. 89:1909–1913. 2003.
|
|
95
|
Steffens Sandra, Schrader Andres J, Blasig
Hanna, et al: Caveolin 1 protein expression in renal cell carcinoma
predicts survival. BMC Urology. 11:252011.
|
|
96
|
Campbell L, Jasani B, Edwards K, et al:
Combined expression of caveolin-1 and an activated AKT/mTOR pathway
predicts reduced disease-free survival in clinically confined renal
cell carcinoma. Br J Cancer. 98:931–940. 2008.
|
|
97
|
Joo HJ, Oh DK, Kim YS, et al: Increased
expression of caveolin-1 and microvessel density correlates with
metastasis and poorprognosis in clear cell renal cell carcinoma.
BJU Int. 93:291–296. 2004.
|
|
98
|
Phuoc NB, Ehara H, Gotoh T, et al:
Immunohistochemical analysis with multiple antibodies in search of
prognostic markers for clear cellrenal cell carcinoma. Urology.
69:843–848. 2007.
|
|
99
|
Zhang ZB, Cai L, Zheng SG, et al:
Overexpression of caveolin-1 in hepatocellular carcinoma with
metastasis and worse prognosis: correlation with vascular
endothelial growth factor, microvessel density and unpaired artery.
Pathol Oncol Res. 15:495–502. 2009.
|
|
100
|
Yu JH, Wei Q, Qi FJ, et al: Significance
of caveolin-1 expression in primary lung cancer. Zhonghua Bing Li
Xue Za Zhi. 35:664–668. 2006.
|
|
101
|
Ho Chao-Chi, Huang Pei-Hsin, Huang
Hsin-Yi, et al: Up-regulated caveolin-1 accentuates the metastasis
capability of lung adenocarcinoma by inducing filopodia formation.
Am J Pathol. 161:1647–1656. 2002.
|
|
102
|
Zhan P, Shen XK, Qian Q, et al: Expression
of caveolin-1 is correlated with disease stage and survival in lung
adenocarcinomas. Oncol Rep. 27:1072–1078. 2012.
|
|
103
|
Yoo SH, Park YS, Kim HR, et al: Expression
of caveolin-1 is associated with poor prognosis of patients with
squamous cell carcinoma of the lung. Lung Cancer. 42:195–202.
2003.
|
|
104
|
Moon KC, Lee GK, Yoo SH, et al: Expression
of caveolin-1 in pleomorphic carcinoma of the lung is correlated
with a poor prognosis. Anticancer Res. 25:4631–467. 2005.
|
|
105
|
Ho CC, Kuo SH, Huang PH, et al: Caveolin-1
expression is significantly associated with drug resistance and
poor prognosis in advanced non-small cell lung cancer patients
treated with gemcitabine-based chemotherapy. Lung Cancer.
59:105–110. 2008.
|
|
106
|
Yang G, Truong LD, Wheeler TM and Thompson
TC: Caveolin-1 expression in clinically confined human prostate
cancer: a novel prognostic marker. Cancer Res. 59:5719–5723.
1999.
|
|
107
|
Satoh T, Yang G, Egawa S, et al:
Caveolin-1 expression is a predictor of recurrence-free survival in
pT2N0 prostate carcinoma diagnosed in Japanese patients. Cancer.
97:1225–1233. 2003.
|
|
108
|
Gumulec J, Sochor J, Hlavna M, et al:
Caveolin-1 as a potential high-risk prostate cancer biomarker.
Oncol Rep. 27:831–841. 2012.
|
|
109
|
Karam JA, Lotan Y, Roehrborn CG, et al:
Caveolin-1 overexpression is associated with aggressive prostate
cancer recurrence. Prostate. 67:614–22. 2007.
|
|
110
|
Yang G, Goltsov AA, Ren C, et al:
Caveolin-1 upregulation contributes to c-Myc-induced high-grade
prostatic intraepithelial neoplasia and prostate cancer. Mol Cancer
Res. 10:218–229. 2012.
|
|
111
|
Steiner I, Jung K, Miller K, et al:
Expression of endothelial factors in prostate cancer: a possible
role of caveolin-1 for tumour progression. Oncol Rep. 27:389–395.
2012.
|
|
112
|
Tahir SA, Ren C, Timme TL, et al:
Development of an immunoassay for serum caveolin-1: a novel
biomarker for prostate cancer. Clin Cancer Res. 9:3653–3659.
2003.
|
|
113
|
Sugie S, Tsukino H, Yamauchi T, et al:
Functional polymorphism in the CAV1 T29107A gene and its
association with prostate cancer risk among Japanese men.
Anticancer Res. 33:1023–1027. 2013.
|
|
114
|
Langeberg WJ, Tahir SA, Feng Z, et al:
Association of caveolin-1 and -2 genetic variants and
post-treatment serum caveolin-1 with prostate cancer risk and
outcomes. Prostate. 70:1020–1035. 2010.
|
|
115
|
Barresi V, Giuffre’ G, Vitarelli E, et al:
Caveolin-1 immuno-expression in human gastric cancer:
histopathogenetic hypotheses. Virchows Arch. 453:571–578. 2008.
|
|
116
|
Sotgia F, Williams TM, Schubert W, et al:
Caveolin-1 deficiency (−/−) conveys premalignant alterations in
mammary epithelia, with abnormal lumen formation, growth factor
independence, and cell invasiveness. Am J Pathol. 168:292–309.
2006.
|
|
117
|
Cantiani L, Manara MC, Zucchini C, et al:
Caveolin-1 reduces osteosarcoma metastases by inhibiting c-Src
activity and met signaling. Cancer Res. 67:7675–7685. 2007.
|
|
118
|
Senetta R, Miracco C, Lanzafame S, et al:
Epidermal growth factor receptor and caveolin-1 coexpression
identifies adult supratentorial ependymomas with rapid unfavorable
outcomes. Neuro Oncol. 13:176–183. 2011.
|
|
119
|
Ruan J and Weng ZL: Analysis of the
relationship between expression of caveolin-1 and prognosis in
bladder transitional cell carcinoma. Zhonghua Zhong Liu Za Zhi.
32:429–431. 2010.(In Chinese).
|
|
120
|
Qayyum T, Fyffe G, Duncan M, et al: The
interrelationships between Src, Cav-1 and RhoGD12 in transitional
cell carcinoma of the bladder. Br J Cancer. 13; 106:1187–1195.
2012.
|
|
121
|
Du ZM, Hu CF, Shao Q, et al: Upregulation
of caveolin-1 and CD147 expression in nasopharyngeal carcinoma
enhanced tumor cell migration and correlated with poor prognosis of
the patients. Int J Cancer. 125:1832–1841. 2009.
|
|
122
|
Rödel F, Capalbo G, Rödel C, et al:
Caveolin-1 as a prognostic marker for local control after
preoperative chemoradiation therapy inrectal cancer. Int J Radiat
Oncol Biol Phys. 73:846–852. 2009.
|
|
123
|
Wu HC, Chang CH, Tsou YA, et al:
Significant association of caveolin-1 (CAV1) genotypes with
prostate cancer susceptibility in Taiwan. Anticancer Res.
31:745–749. 2011.
|
|
124
|
Tsou YA, Tsai CW, Tsai MH, et al:
Association of caveolin-1 genotypes with nasopharyngeal carcinoma
susceptibility in Taiwan. Anticancer Res. 31:3629–3632. 2011.
|
|
125
|
Syeed N, Hussain F, Husain SA, et al:
5′-CpG island promoter hypermethylation of the CAV-1 gene in breast
cancer patients of Kashmir. Asian Pac J Cancer Prev. 13:371–375.
2012.
|
|
126
|
Shajahan AN, Goel S, de Assis S, et al:
Changes in mammary caveolin-1 signaling pathways are associated
with breast cancer risk in rats exposed to estradiol in utero or
during prepuberty. Horm Mol Biol Clin Investig. 2:227–234.
2010.
|
|
127
|
Lin M, DiVito MM, Merajver SD, et al:
Regulation of pancreatic cancer cell migration and invasion by RhoC
GTPase and caveolin-1. Mol Cancer. 4:212005.
|
|
128
|
Thomas S, Overdevest JB, Nitz MD, et al:
Src and caveolin-1 reciprocally regulate metastasis via a common
downstream signaling pathway in bladdercancer. Cancer Res.
71:832–841. 2011.
|
|
129
|
Arpaia E, Blaser H, Quintela-Fandino M, et
al: The interaction between caveolin-1 and Rho-GTPases promotes
metastasis by controlling the expression of alpha5-integrin and the
activation of Src, Ras and Erk. Oncogene. 31:884–896. 2012.
|
|
130
|
Chiu WT, Lee HT, Huang FJ, et al:
Caveolin-1 upregulation mediates suppression of primary breast
tumor growth and brain metastases by stat3 inhibition. Cancer Res.
71:4932–4943. 2011.
|
|
131
|
Zhang H, Su L, Müller S, et al:
Restoration of caveolin-1 expression suppresses growth and
metastasis of head and neck squamous cellcarcinoma. Br J Cancer.
99:1684–1694. 2008.
|
|
132
|
Yeh D, Chen C, Sun MZ, et al: Caveolin-1
is an important factor for the metastasis and proliferation of
human small cell lung cancer NCI-H446cell. Anat Rec (Hoboken).
292:1584–1592. 2009.
|
|
133
|
Urra H, Torres VA, Ortiz RJ, et al:
Caveolin-1-enhanced motility and focal adhesion turnover require
tyrosine-14 but not accumulation to the rear inmetastatic cancer
cells. PLoS One. 7:e330852012.
|
|
134
|
Duxbury MS, Ito H, Ashley SW, et al:
CEACAM6 cross-linking induces caveolin-1-dependent, Src-mediated
focal adhesion kinase phosphorylation inBxPC3 pancreatic
adenocarcinoma cells. J Biol Chem. 279:23176–82. 2004.
|
|
135
|
Meyer C, Liu Y, Kaul A, et al: Caveolin-1
abrogates TGF-β mediated hepatocyte apoptosis. Cell Death Dis.
4:e4662013.
|
|
136
|
Katsogiannou M, El Boustany C, Gackiere F,
et al: Caveolae contribute to the apoptosis resistance induced by
the alpha(1A)-adrenoceptor in androgen-independent prostate cancer
cells. PLoS One. 4:e70682009.
|
|
137
|
Rodriguez DA, Tapia JC, Fernandez JG, et
al: Caveolin-1-mediated suppression of cyclooxygenase-2 via a
beta-catenin-Tcf/Lef-dependent transcriptional mechanism reduced
prostaglandin E2 production and survivin expression. Mol Biol Cell.
20:2297–2310. 2009.
|
|
138
|
Palozza P, Sestito R, Picci N, et al: The
sensitivity to beta-carotene growth-inhibitory and proapoptotic
effects is regulated by caveolin-1 expression in human colon and
prostate cancer cells. Carcinogenesis. 29:2153–2161. 2008.
|