Introduction
Sorcin, a soluble resistance-related calcium-binding
protein, was initially observed to be overproduced in the
vincristine-resistant DC-3F/VCR-5 Chinese hamster cell line, by
Meyers and Biedler (1). Following
this, it was also identified in drug-resistant mice and human cell
lines (2–4). Sorcin is expressed in numerous tissues
in mammals, such as the liver, lungs and, most abundantly, cardiac
myocytes. Its expression in normal mammalian tissues is highly
conserved.
Sorcin is a cytoplasmic protein that is tightly
associated with free ribosomes, rough endoplasmic reticulum
cisternae, mitochondria, nuclear membrane and microtubules
(5). Biological characteristic
studies of sorcin have confirmed a molecular mass of 22 kDa and
determined that it is part of the penta-EF-hand (PEF) protein
family, with typical calcium-binding sites located in the first
pair of EF-hands (4,6,7).
The overexpression of sorcin has been reported in a
number of tumor-resistant cell lines. Increasingly convincing
evidence has suggested that sorcin is involved in survival
mechanisms responsible for multidrug resistance and is associated
with a poor prognosis during the therapeutic treatment of cancer
patients (8). To date, the
overexpression of sorcin has been observed in a number of
multidrug-resistant (MDR) cell lines and several tumor cell types,
including human colorectal cancer cells, human gastric cancer cells
(9,10), leukemia (11,12),
ovarian and breast cancer cells (13,14)
and lung cancer (15). Our
unpublished data, regarding the correlation between the expression
levels of sorcin and the outcome of neoadjuvant chemotherapy (NAC)
in breast cancer patients, showed that the remission rate was
significantly higher in patients with low expression levels of
sorcin than in patients with high sorcin expression levels, and
that the expression of sorcin was reduced following treatment. It
was hypothesized that the expression level of sorcin in breast
cancer may predict the efficiency of the paclitaxel/epirubicin
regimen in NAC.
To date, laboratory detections of sorcin expression
are predominantly at the mRNA and protein level using methods of
proteomics, including reverse transcription-polymerase chain
reaction, western blot analysis and immunohistochemistry (IHC),
subsequent to siRNA transfection. At the DNA level, sorcin
expression has also been tested using microarray and northern blot
analysis (12,13). Two-dimensional gel electrophoresis
(2-DE) is one of the most commonly used techniques in proteomics.
It is widely used to study protein expression patterns in a variety
of cell lines (11). Over the last
two decades, this approach has been used for profiling expression
patterns in cancer and in cancer cells with multidrug resistance,
which enables the identification of proteins that are involved in
tumorigenesis and multidrug resistance for specific drugs.
Therefore, methods of proteomics and immunohistochemistry were
investigated in this study to assess the role of sorcin in a
phenotype of breast cancer with multidrug resistance.
Understanding this protein may provide targeted
therapeutic applications among cancer patients. Sorcin may be a
potential prognostic marker for a number of malignancies, including
acute leukemia and breast cancer, which is of particular relevance
in the current study. However, the mechanisms whereby sorcin is
interrelated with multidrug resistance may vary in different cancer
cells (16).
Materials and methods
Sample preparation
Serum samples were extracted from 30 stage III and
IV breast cancer patients who received preoperative NAC and were
recruited for a prospective preoperative clinical trial at the
Yantai Yuhuangding Hospital (Yantai, China) between 2008 and 2010.
The patients suffered from locally advanced breast cancer, in which
the purpose of neoadjuvant treatment was to downstage the cancer
for an improved chance of complete resection, or high operative
risks were anticipated due to old age or comorbidities, which
prevented them from undergoing initial surgical treatment. Among
the subjects, 20 patients received two cycles of 175
mg/m2 paclitaxel and 80 mg/m2 epirubicin on
day one, once every three weeks, while 10 patients received two
cycles of 75 mg/m2 docetaxel and 80 mg/m2
epirubicin on day one, once every three weeks. In total, 24
patients responded to the chemotherapy [complete response (CR),
partial response (PR) and stable disease (SD)] and 18 patients
developed progressive disease (PD). All samples were removed with
patient consent.
Blood samples were drawn prior to each chemotherapy
course, including a baseline pretreatment sample (day zero) and a
post-chemotherapy (week six), but prior to the third course of
chemotherapy or surgery, sample. Blood samples were subsequently
centrifuged at 1,200 × g for 20 min at 4°C, and the supernatant was
aliquoted and stored at −80°C until use. This biomarker study was
approved by the institutional review board at Yantai Yuhuangding
Hospital and a waiver of informed consent was granted.
Serum preparation (extraction of serum
from human subject whole blood)
Serum samples were prepared according to the
manufacturer’s instructions in the Proteoprep kit, Sigma-Aldrich
(St. Louis, MO, USA). In total, 300 μl of the protein extraction
reagent, mixed with high purity water, was added to 400 μl of the
equilibration buffer, and then subjected to centrifugation at 5,000
× g for 5–10 sec at −4°C and repeated once. Following this, 25–50
μl of each serum sample was diluted to 100 μl with the
equilibration buffer and mixed thoroughly. This sample was then
added to the top of the column and incubated for 5–10 min at room
temperature, and subsequently centrifuged at 8,000 × g for 60 sec
at −4°C and repeated once. In the final step, 125 μl of
equilibration buffer was added prior to storage of the samples at
−20°C.
Extraction of salt with acetone and
protein solubilization
Serum samples were initially precipitated with
acetone. Cold acetone (Fluka Biochemika, Buchs, Switzerland) was
added to a serum sample at a ratio of 1:4 and stored at −20°C for 2
h, following which, centrifugation was conducted at 12,000 × g for
30 min. The supernatant was discarded and the pellet was dissolved
in 200 μl of the lysis buffer containing 40 mM Tris, 7 M urea, 2 M
thiourea, 2% 3-[(3-cholamidopropyl)
dimethylammonio]-1propanesulfonate (CHAPS), 65 mM dithiothreitol
(DTT) and 1% IPG Buffer (GE Healthcare, Little Chalfont, UK).
2-DE analysis
Protein concentration was determined using the
2D-Quant Kit (Amersham Pharmacia Biotech, Amersham, UK), according
to the manufacturer’s instructions. In total, 500 μg of each pooled
protein sample was diluted in the rehydration buffer (7 M urea, 18
mM DTT, 4% CHAPS, 0.5% IPG buffer and 0.002% bromophenol blue). The
isoelectric focusing was performed on the Ettan IPGphor II (GE
Healthcare Bio-Sciences, Piscataway, NJ, USA). The IPG strips were
initially rehydrated at 30 V for 12 h, and subsequently focused at
500 V for 1 h, 1,000 V for 1 h, 3,000 V for 3 h and 5,000 V for 3
h, and then maintained at 8,000 V until a total of 50,000 V/hr was
achieved. Following isoelectric focusing, the IPG strips were
equilibrated with 1.5 M Tris-HCl (pH 8.8), 6 M urea, 87% glycerol,
2% sodium dodecyl sulfate (SDS) and 0.2% bromophenol blue. The IPG
strips were initially treated with 15 ml 1% DTT for 10 min with
constant shaking, followed by alkylation with 15 ml 2.5%
indole-3-acetic acid for 15 min.
The equilibrated strips were transferred to 12.5%
SDS polyacrylamide gel electrophoresis (SDS-PAGE) on the Ettan DALT
twelve system (GE Healthcare) with constant power (0.2 W/gel, 1 h;
0.4 W/gel, 1 h; 250V, 4.5 h). The gels were stained with Coomassie
blue R350 (GE Healthcare), and scanned using a PowerLook 2100 XL
scanner system (Umax Technologies, Taipei, Taiwan).
Spots of interest were excised from gels stained by
Coomassie Blue R350, and were digested with sequencing grade
modified trypsin (Promega Corporation, Madison, WI, USA) using the
following protocol: The colloidal particle was soaked in decoloring
working liquid (50% acetonitrile/50 mmol/l
NH4HCO3) in 37°C water bath until the blue
faded away. The decolored blue dots were dehydrated for 15 min with
100% acetonitrile, and subsequently spun briefly. Trypsin solution
was added to the dried colloidal particle (10 μg/μl trypsin, 100
mmol/l NH4HCO3 and 125 mmol/l
CaCl2) at 4°C and bulged for 30 min. The excessive
trypsin epispastics were removed and 10 μl 25 mmol/l
NH4HCO3 enzyme solution was added and the
resulting mixture was incubated at 37°C overnight. Following the
completion of the enzyme solution preparation, the extract liquor
[acetonitrile (ACN) 50% + 5% trifluoroacetic acid (TFA)] was used
to extract the peptides and the solution was heated at 37°C for 1
h. The liquid supernatant was extracted to a new microcentrifuge
tube, enriched and dried by centrifugation. The dissolved and dried
peptides were subsequently used with 3 μl diluent (ACN 30% +1 %
TFA) for mass spectrometry detection.
Mass spectrum identification
Subsequent protein identification was conducted on
the ABI 4700 Proteomic Analyzer MALDI-TOF-MS/MS mass spectrometer
(Life Technologies, Carlsbad, CA, USA) in the reflective mode. The
peptide mass fingerprint (PMF) was acquired between 800–3,500 Da.
In total, 24 peaks from the PMF were selected to obtain the MS/MS
spectra. The PMF and MS/MS results were then searched against a
human subset of the Swiss-Prot database using the GPS explorer
software (Life Technologies).
Western blot analysis
In total, 100 μg serum sample was diluted with an
equal amount of loading buffer (12% SDS; 135 mM Tris-HCl, pH 6.8;
20% glycerol; 0.02% bromophenol blue and 10% 3-mercaptoethanol).
The samples were subjected to polyacrylamide gel electrophoresis
and transferred onto a polyvinylidene difluoride membrane
(Millipore, Massachusetts, CA, USA). The membranes were treated
with Tris-buffered saline with 0.1% Tween-20 containing 5% dried
non-fat milk at 4°C overnight. The membranes were then
immunoblotted with the monoclonal mouse anti-human sorcin antibody
(sc-100859; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
and the monoclonal rabbit anti-avian β-actin antibody) sc-47778;
Santa Cruz Biotechnology, Inc.) at 4°C overnight. The following
day, the membranes were washed three times with phosphate-buffered
saline containing 0.05% Tween-20 and incubated with a polyclonal
goat anti-mouse peroxidase-conjugated secondary antibody (ZSGB-BIO,
Beijing, China) at room temperature for 1 h. The immunoreactive
blots were identified using the chemiluminescence detection ECL
plus kit (GE healthcare, Buckinghamshire, UK) and the LAS 3000
Lumino-image analyzer (Fujifilm, Tokyo, Japan). The band intensity
was analyzed by using the NIH image analysis software (NIH,
Bethesda, ML, USA).
Statistical analysis
The χ2 test was used to establish the
statistical significance between the expression levels of sorcin
and the number of patients who developed resistance to NAC.
P<0.01 was considered to indicate a statistically significant
difference (Table I).
 | Table IPatient responses, exhibiting an up-
or downregulation in sorcin levels among patients who were
NAC-sensitive or -resistant, respectively. |
Table I
Patient responses, exhibiting an up-
or downregulation in sorcin levels among patients who were
NAC-sensitive or -resistant, respectively.
| Subgroups | Total, n | Upregulated sorcin,
n | Downregulated sorcin,
n | χ2 | P-value* |
|---|
| NAC-sensitive | 29 | 6 | 23 | 12.87 | <0.01 |
| MDR | 21 | 15 | 6 | | |
Results
Sorcin upregulation in MDR patients
According to the Response Evaluation Criteria in
Solid Tumors (17), the subjects
were grouped into CR, PR, SD and PD. In the current study, patients
that were evaulated as CR, PR and SD were predicted to be
responsive to NAC treatment. Therefore, among the 50 breast cancer
patients included in this study, 29 responded well to the NAC
(58%), whereas 21 patients developed multidrug resistance (42%)
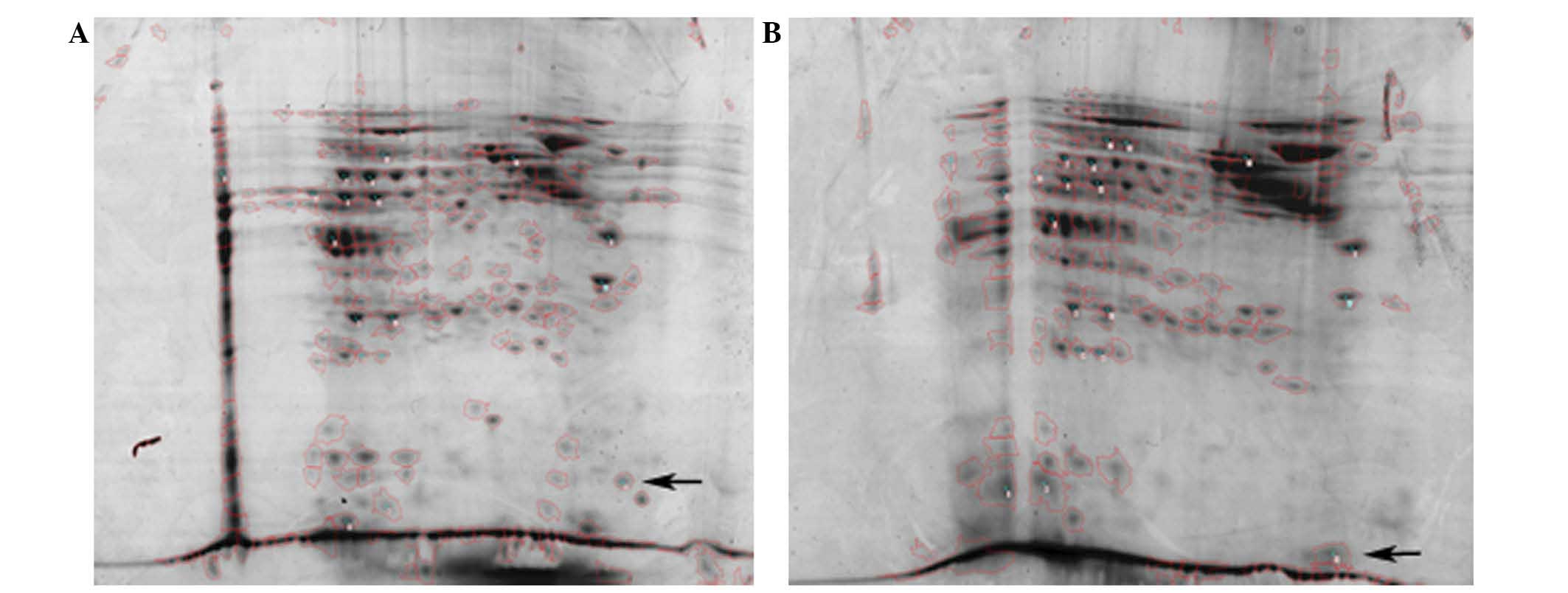
(Table I). The 2-DE analysis of the
serum samples revealed that sorcin was upregulated in six out of 29
(20.7%) NAC-sensitive patients and, in those who developed
multidrug resistance, sorcin was upregulated in 15 out of 21
patients (71.4%).
The sample pools of NAC-sensitive and MDR patients
were run separately using 2-DE analysis; a protein spot of sorcin
was identified on the 2-DE map loaded with serum samples from the
NAC-resistant patients (Fig. 1B) as
well as a further 19 distinguishable protein spots (results not
shown). Each pool was analyzed three times.
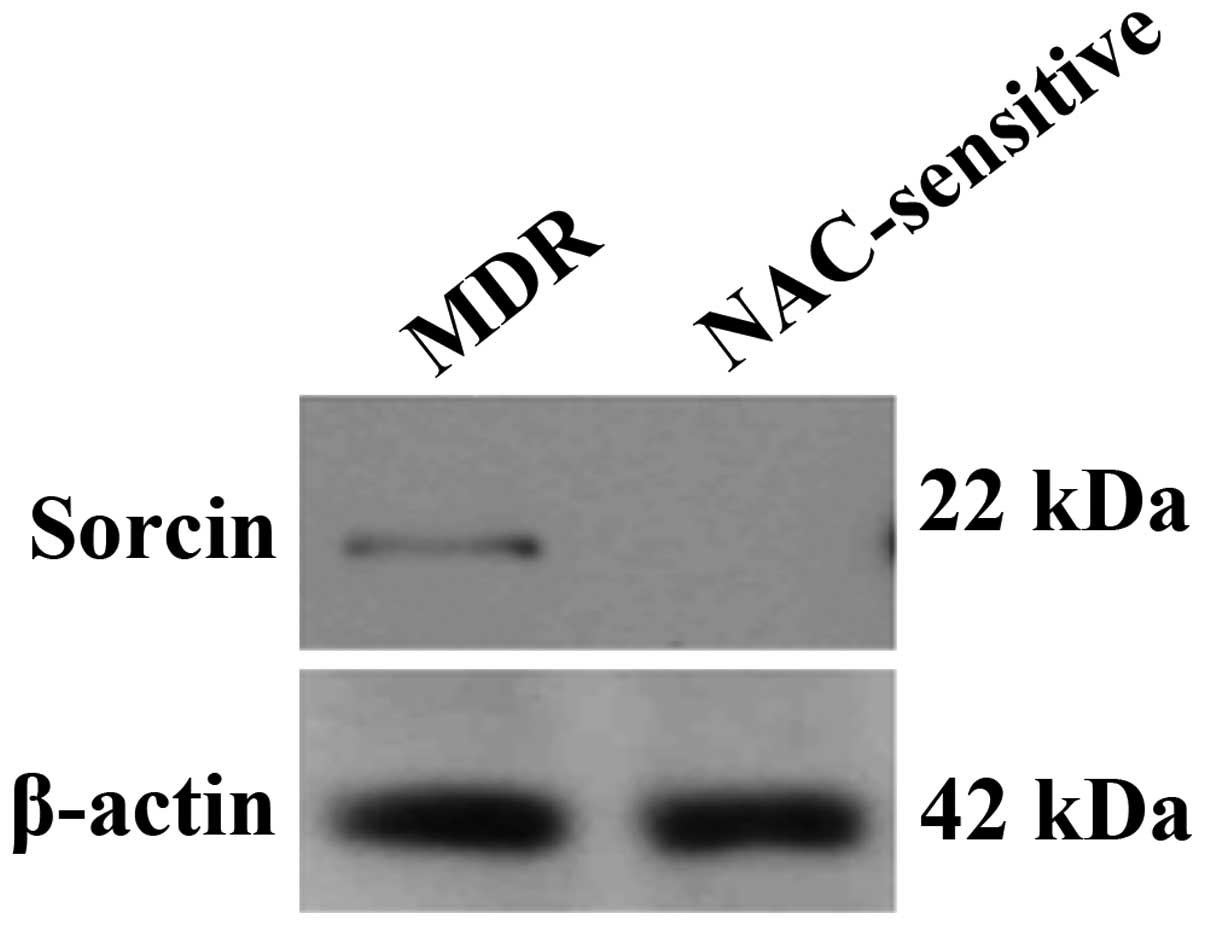
Western blot analysis of MDR
patients
Western blot analysis using anti-sorcin antibodies
revealed a specific band in the sample pool of MDR serum, whereas
no band was visible for the NAC-sensitive group (Fig. 2). Anti-β-actin antibodies were used
as a control for the analysis.
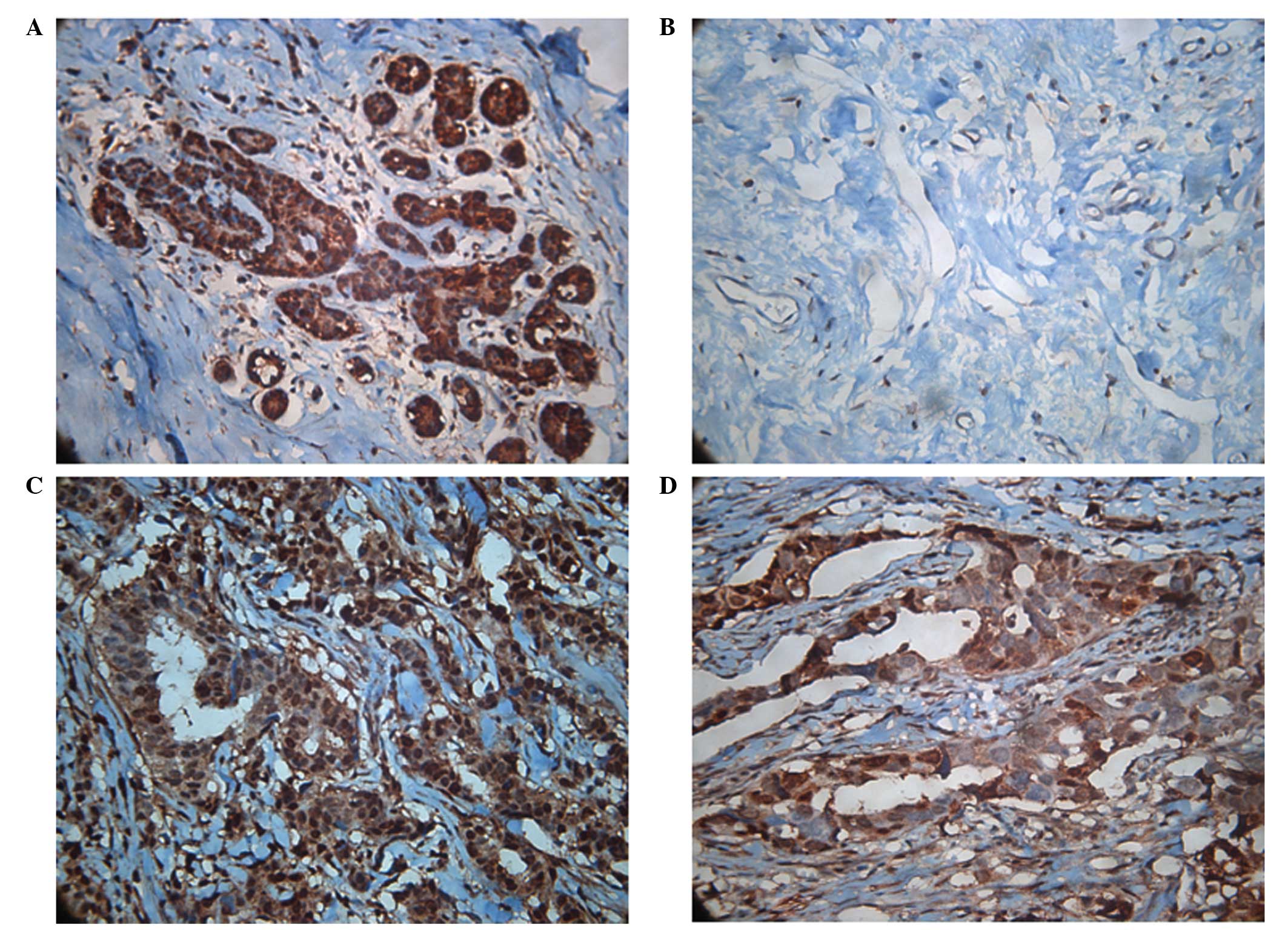
IHC
Furthermore, as predicted, IHC of the infiltrating
ductal breast cancerous tissue revealed heavy staining of sorcin in
tissues obtained from patients with developed resistance to NAC.
The results from two cases are shown in Fig. 3, where staining is particularly
evident in the cytoplasm.
Discussion
This study provides evidence of the involvement of
sorcin in the development of drug resistance in breast cancer.
Using 2-DE and western blot analysis, sorcin was identified in the
blood serum of human breast cancer subjects and new insights into
the manner by which the expression levels of sorcin affect the
outcome of NAC have been presented. The 2-DE analysis of the pooled
sample serum of breast cancer patients revealed the upregulation of
sorcin in >70% of all participants who did not respond to NAC
(those with PD). Subsequent western blot analysis confirmed a
positive band for patients who developed multidrug resistance, in
contrast to those who were responsive to NAC. Furthermore, IHC
staining of the cancerous tissue biopsy confirmed the sorcin
upregulation. To the best of our knowledge, this is one of few
studies that uses human serum as the test sample, as the majority
of studies investigating the sorcin expression in tumors use
tissues or cell lines. IHC staining of the breast tissue of
patients from biopsies prior to and following NAC indicate that
patients who responded well to NAC have significantly reduced
sorcin expression in the cytosol, following NAC.
The majority of studies investigating sorcin
expression have employed cell lines that are commercially available
and engineered using cDNA cloning or siRNA (14,18).
Additionally, by examining the resistance of particular cancer
cells in apoptosis, the mechanism of sorcin in the prognosis of
cancer maybe disclosed (11).
Proteomic studies of organelle compartmentalization of sorcin have
also been thoroughly investigated, predominantly through direct
cell fractionation or following the treatment of target cell lines
with antiblastic agents, such gemcitabine (15) and 5-fluorouracil (19). The current study, however, utilized
serum samples acquired directly from breast cancer patients, and
compared the sorcin levels between patients who presented with a
resistant phenotype for chemotherapy and those who were sensitive
to NAC. To the best of our knowledge, studies analyzing sorcin
levels using human serum are limited. To date, the mechanisms of
the occurrence, development, metastasis or resistance of breast
cancer are not fully understood. Research is ongoing to locate
suitable markers at the molecular level; sorcin is one of the
numerous protein markers that is being systematically studied
(20), and we hypothesize that the
role of sorcin in predicting the chemotherapeutic response and
aggressiveness of the malignancy is closely associated with its
causative effect in multidrug resistance.
Breast cancer is one of the leading causes of
cancer-related mortality in females worldwide (21); it is associated with high morbidity,
poor prognosis and high metastatic rates (22). Predominantly, breast cancer cell
resistance to antiblastic cells is most likely a causative factor
in therapeutic failure (23). As
sorcin is considered a pivotal breast cancer resistance-related
protein, understanding the mechanisms of sorcin at a molecular
level may have a significant impact on the clinical management of
breast carcinoma. IHC has been used to study the expression of
sorcin in breast cancer tissue (24). Liu et al (25) demonstrated that 85.1% (40/47) of
postoperative samples from breast cancer patients positively
express sorcin. This may be partially associated with the presence
of the progesterone receptor, overall survival rate and
disease-free survival, but is not likely to be associated with the
prognosis or clinical manifestation.
However, another study demonstrated that sorcin was
only involved in the development of low-level paclitaxel resistance
when full-length sorcin cDNA was transfected into MCF-7 human
breast cancer cells, which are estrogen receptor-positive, and
MDA-MB435S (parental MCF-7) cells (14). However, the overexpression of
P-glycoprotein (P-gp) did not correlate with the degree of
resistance in the paclitaxel-resistant human ovarian carcinoma
subline, MCF-7. Therefore, it was speculated that sorcin may cause
paclitaxel resistance in breast cancer, and may be dependent on the
presence of estrogen receptors. Additionally, Kawakami et al
(16) demonstrated that if sorcin
was knocked down from an MDR1/P-gp-overexpressing MDR subline
established from the human cervical carcinoma cell line, HeLa, the
level of MDR1, which modulates the MDR1/P-gp transporter, was
increased. Together with the increased level of caspase-3, it was
hypothesized that the downregulation of sorcin may elevate the
intracellular levels of calcium via the upregulation of MDR1 and
thus, activated caspase-3 may induce apoptosis. Furthermore, by
examining 25 breast cancer patient samples for P-gp expression,
Zhao et al (26)
demonstrated that minimal MDR1 mRNA expression may also lead to a
MDR phenotype. P-gp is not expressed in normal breast tissue;
however, it may be observed in cancerous breast tissue and normal
peritumoral tissue, a common phenomenon that can be applied to the
majority of malignancies (27).
To conclude, the upregulation of sorcin in the serum
of breast cancer patients may be partially responsible for the
development of multidrug resistance. NAC moderately reduces sorcin
expression; however this does not occur in all breast cancer cases.
The mechanism by which sorcin affects the development of multidrug
resistance and patient response to NAC remains unknown. Although
sorcin may be a potential prognostic marker for predicting the
treatment outcome in breast cancer patients and possibly further
malignancies, the mechanism of sorcin and the association with
multidrug resistance may differ across cancer cell types. Gaining
an improved understanding of this protein may provide targeted
therapeutic applications among cancer patients.
Acknowledgements
This study was supported by Yantai Science and
Technology Program (grant no. 2009155-5).
Reference
|
1
|
Meyers MB and Biedler JL: Increased
synthesis of a low molecular weight protein in
vincristine-resistant cells. Biochem Biophys Res Commun.
99:228–235. 1981.
|
|
2
|
Beyer-Sehlmeyer G, Hiddemann W, Wörmann B
and Bertram J: Suppressive subtractive hybridisation reveals
differential expression of serglycin, sorcin, bone marrow
proteoglycan and prostate-tumour-inducing gene I (PTI-1) in
drug-resistant and sensitive tumour cell lines of haematopoetic
origin. Eur J Cancer. 35:1735–1742. 1999.
|
|
3
|
Hamada H, Okochi E, Oh-hara T and Tsuruo
T: Purification of the Mr 22,000 calcium-binding protein (sorcin)
associated with multidrug resistance and its detection with
monoclonal antibodies. Cancer Res. 48:3173–3178. 1988.
|
|
4
|
Wang SL, Tam MF, Ho YS, Pai SH and Kao MC:
Isolation and molecular cloning of human sorcin a calcium-binding
protein in vincristine-resistant HOB1 lymphoma cells. Biochim
Biophys Acta. 1260:285–293. 1995.
|
|
5
|
Lalioti VS, Ilari A, O’Connell DJ, Poser
E, Sandoval IV and Colotti G: Sorcin links calcium signaling to
vesicle trafficking, regulates Polo-like kinase 1 and is necessary
for mitosis. PLoS One. 9:e854382014.
|
|
6
|
Van der Bliek AM, Meyers MB, Biedler JL,
Hes E and Borst P: A 22-kd protein (sorcin/V19) encoded by an
amplified gene in multidrug-resistant cells, is homologous to the
calcium-binding light chain of calpain. EMBO J. 5:3201–3208.
1986.
|
|
7
|
Xie X, Dwyer MD, Swenson L, Parker MH and
Botfield MC: Crystal structure of calcium-free human sorcin: a
member of the penta-EF-hand protein family. Protein Sci.
10:2419–2425. 2001.
|
|
8
|
Maddalena F, Laudiero G, Piscazzi A, et
al: Sorcin induces a drug-resistant phenotype in human colorectal
cancer by modulating Ca(2+) homeostasis. Cancer Res. 71:7659–7669.
2011.
|
|
9
|
Deng L, Su T, Leng A, et al: Upregulation
of soluble resistance-related calcium-binding protein (sorcin) in
gastric cancer. Med Oncol. 27:1102–1108. 2010.
|
|
10
|
He Q, Zhang G, Hou D, et al:
Overexpression of sorcin results in multidrug resistance in gastric
cancer cells with up-regulation of P-gp. Oncol Rep. 25:237–243.
2011.
|
|
11
|
Qi J, Liu N, Zhou Y, et al: Overexpression
of sorcin in multidrug resistant human leukemia cells and its role
in regulating cell apoptosis. Biochem Biophys Res Commun.
349:303–309. 2006.
|
|
12
|
Tan Y, Li G, Zhao C, et al: Expression of
sorcin predicts poor outcome in acute myeloid leukemia. Leuk Res.
27:125–131. 2003.
|
|
13
|
Hu Y, Cheng X, Li S, et al: Inhibition of
sorcin reverses multidrug resistance of K562/A02 cells and
MCF-7/A02 cells via regulating apoptosis-related proteins. Cancer
Chemother Pharmacol. 72:789–798. 2013.
|
|
14
|
Parekh HK, Deng HB, Choudhary K, Houser SR
and Simpkins H: Overexpression of sorcin, a calcium-binding
protein, induces a low level of paclitaxel resistance in human
ovarian and breast cancer cells. Biochem Pharmacol. 63:1149–1158.
2002.
|
|
15
|
Qu Y, Yang Y, Liu B and Xiao W:
Comprarative proteomic profiling identified sorcin being associated
with gemcitabine resistance in non-small cell lung cancer. Med
Oncol. 27:1303–1308. 2010.
|
|
16
|
Kawakami M, Nakamura T, Okamura N, et al:
Knock-down of sorcin induces up-regulation of MDR1 in HeLa cells.
Biol Pharm Bull. 30:1065–1073. 2007.
|
|
17
|
Ding Q, Cheng X, Yang L, et al: PET/CT
evaluation of response to chemotherapy in non-small cell lung
cancer: PET response criteria in solid tumors (PERCIST) versus
response evaluation criteria in solid tumors (RECIST). J Thorac
Dis. 6:677–683. 2014.
|
|
18
|
Hu Y, Li S, Yang M, et al: Sorcin
silencing inhibits epithelial-to-mesenchymal transition and
suppresses breast cancer metastasis in vivo. Breat Cancer Res
Treat. 143:287–299. 2014.
|
|
19
|
Landriscina M, Laudiero G, Maddalena F, et
al: Mitochondrial chaperone Trap1 and the calcium binding protein
Sorcin interact and protect cells against apoptosis induced by
antiblastic agents. Cancer Res. 70:6577–6586. 2010.
|
|
20
|
Zhu L, Zhang L, Li G, et al: detection and
clinical significance of sorcin gene in relapse or refractory ALL
patients. Zhong Hua Zhong Liu Fang Zhi Za Zhi. 14:1568–1570.
2007.
|
|
21
|
Banerji S, Cibulskis K, Rangel-Escareno C,
et al: Sequence analysis of mutations and translocations across
breast cancer subtypes. Nature. 486:405–409. 2012.
|
|
22
|
Glackin CA: Targeting the twist and Wnt
signaling pathways in metastatic breast cancer. Maturitas.
79:48–51. 2014.
|
|
23
|
Paplomata E and O’Regan R: The
PI3K/AKT/mTOR pathway in breast cancer: targets, trails and
biomarkers. Ther Adv Med Oncol. 6:154–166. 2014.
|
|
24
|
Ward S, Scope A, Rafia R, et al: Gene
expression profiling and expanded immunohistochemistry tests to
guide the use of adjuvant chemotherapy in breast cancer management:
a systematic review and cost-effectiveness analysis. Health Technol
Assess. 17:1–302. 2013.
|
|
25
|
Liu Y, Wang S, Sun Y, et al: Prognostic
significance of soluble resistance-related calcium binding protein
expression in primary breast carcinoma. Xian Dai Yi Yao Wei Sheng.
12:1769–1771. 2006.(In Chinese).
|
|
26
|
Zhao F, Jiang J, Fan LJ, Yang XH and Zhang
Y: P-glycoprotein expression in the development of primary
multidrug resistance in breast carcinoma. Yi Xue Zheng Ming.
16:1513–1516. 2005.(In Chinese).
|
|
27
|
Zhang BB, WU Y and Li QW: Soluble
resistance-related calcium binding protein. Zhong Guo Sheng Wu Hua
Xue Yu Fen Zi Sheng Wu Xue Bao. 12:942–946. 2006.(In Chinese).
|