Introduction
5-Fluorouracil (5-FU) is a key chemotherapy drug for
colorectal cancer, and therefore, the inherent or acquired
resistance to 5-FU is a serious problem. A number of studies have
reported the association between the response to or toxicity of the
drug and 5-FU metabolism-associated factors, including thymidylate
synthase, dihydropyrimidine dehydrogenase, folate cofactors and
orotate phosphoribosyltransferase (1–4).
However, at present no reliable biomarkers for the sensitivity or
resistance to 5-FU chemotherapy have been identified.
Mammalian heat shock proteins (HSPs) such as HSP90,
HSP70, HSP60 and small HSPs (15–30 kDa), including HSP27, are
present in numerous organs and are considered as molecular
chaperones in protein-protein interactions, including folding,
unfolding and assembly, as well as anti-apoptotic proteins and
contributors to cell survival (5,6).
Numerous studies have reported that HSP27 expression contributes to
the malignant properties of cancer cells, including the resistance
to treatment, tumorigenicity and the inhibition of apoptosis
(7–11). In colorectal cancer, certain studies
have reported that the expression of HSP27 is involved in
doxorubicin or irinotecan resistance in vitro (12,13),
and recent clinicopathological studies have revealed HSP27 to be a
prognostic marker (14–16). Our previous studies also indicated
that the protein levels of HSP27 expression contributed to the
degree of resistance to 5-FU in studies performed in vitro
and in vivo using a xenograft model (17,18).
A variety of stimuli induce the phosphorylation of
serine residues 15, 78 and 82 in HSP27 using various protein
kinases, such as mitogen-activated protein kinase (MAPK) activated
protein kinase 2 (MAPKAPK-2), via p38 MAPK (19–24).
This post-translational modification affects a number of the
cellular functions of HSP27. As a consequence of the functional
importance of HSP27 phosphorylation, aberrant HSP27 phosphorylation
has been linked to several clinical conditions, including cancer
progression and the malignant behavior of a number of cancer types
(19). Pathogenic conditions
associated with aberrant HSP27 phosphorylation, as well as
potential therapeutic strategies aimed at modulating HSP27
phosphorylation, are expected to be developed in the future.
The present study aimed to clarify whether HSP27
expression or phosphorylation was modulated by 5-FU exposure and to
investigate whether the inhibition of HSP27 phosphorylation by a
specific kinase inhibitor affected the sensitivity of colorectal
cancer cells to 5-FU.
Materials and methods
Drug, cell lines and cell culture
conditions
The anticancer drug 5-FU was purchased from Kyowa
Hakko Bio Co., Ltd. (Tokyo, Japan). A selective p38 MAPK inhibitor,
SB203580 (20), was purchased from
Promega Corporation (Madison, WI, USA). The human colon cancer cell
lines, HCT116, HCT15 and HT29, were obtained from the American Type
Culture Collection (Manassas, VA, USA). The cells were grown in
RPMI-1640 medium (Gibco-BRL, Carlsbad, CA, USA). Each culture was
supplemented with 10% heat-inactivated fetal bovine serum (FBS; CSL
Ltd., Melbourne, Australia) and 1% penicillin/streptomycin (1 ml)
in a humidified 5% CO2 incubator at 37°C. Prior to the
experiments, the cells were incubated without FBS for 24 h.
Western blot analysis for HSP27 and
phosphorylated HSP27
Subconfluent cells were exposed or not exposed to
5-FU (1.28 μg/ml) with SB203580 treatment (0, 1 or 10 μM) in
culture medium for 48 h. The total cell lysates were then extracted
using lysis buffer [20 mM Tris/HCL (pH 7.5), 150 mM NaCl, 1 mM
EDTA, 1 mM EGTA, 1% TritonX-100, 2.5 mM sodium pyrophosphate, 50 mM
NaF, 50 mM HEPES, 1 mM Na3VO4 and 2 mM
phenylmethylsulfonyl fluoride; Cell Signaling Technology, Inc.,
Danvers, MA, USA]. The quantity of cell lysates was determined
using a Bio-Rad DC Protein Assay kit (Bio-Rad, Hercules, CA, USA),
and a total of 20 μg lysates were resolved in Ready Gel (Bio-Rad)
and transferred to an Immuno-Blot polyvinylidene fluoride membrane
(Bio-Rad). The membrane was blocked with phosphate-buffered saline
(PBS; Gibco-BRL) containing 5% skimmed milk powder for 2 h at room
temperature, then incubated at 4°C overnight with anti-human HSP27
mouse monoclonal antibody (1:2,000; G3.1; Lab Vision Corporation,
Fremont, CA, USA), anti-human phospho-HSP27 (Ser15) rabbit
polyclonal antibody (1:500; Upstate Biotechnology, Inc., Lake
Placid, NY, USA), anti-human phospho-HSP27 (Ser78) mouse monoclonal
antibody (1:2,000; Upstate Biotechnology, Inc.), anti-human
phospho-HSP27 (Ser82) rabbit polyclonal antibody (1:1,000; Upstate
Biotechnology, Inc.) or anti-human β-actin mouse monoclonal
antibody (1:5,000; AC74; Sigma-Aldrich, St. Louis, MO, USA). The
membranes were incubated for 30 min with a horseradish
peroxidase-conjugated anti-mouse immunoglobulin G (IgG) (1:2,500;
Promega Corporation) or anti-rabbit IgG (Promega Corporation).
Bound complexes were detected using the ECL-Plus reagent (GE
Healthcare Life Sciences, Chalfont, UK) according to the
manufacturer’s instructions. Each experiment was performed in
triplicate.
Cell proliferation assay
A cell proliferation assay was performed using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl bromide (MTT) assay, as
described previously (17).
Briefly, 5,000 cells/well in 96-well microtiter plates (Sumilon;
Sumitomo Bakelite Co., Ltd., Tokyo, Japan) were incubated for 24 h
and exposed to various concentrations of 5-FU with SB203580
treatment (0, 1, 10 or 50 μM) for 48 h, followed by a drug-free
medium for an additional 24 h. The absorbance in the wells was
measured using an NJ-2300 microplate spectrophotometer at
wavelengths of 540 and 630 nm (Immuno Reader; Nalge Nunc
International, Rochester, NY, USA). The inhibition rate was
calculated using the following formula: Inhibition rate (%) = [1 -
(mean absorbance of drug wells / mean absorbance of control wells)]
× 100. The absorbance of each well was adjusted using the mean
absorbance of the blank wells. 5-FU sensitivity was evaluated using
the half maximal inhibitory concentration (IC50) value,
which corresponded to the concentration of the drug required to
inhibit cell growth by 50% relative to the untreated cells.
A cell count of viable cells was also performed to
evaluate cell proliferation. Viable cells were counted four days
after plating in triplicate dishes in various concentrations of
SB203580 (0, 1, 10 or 50 μM) in RPMI-1640 containing 5% FBS.
Statistical analysis
Data are expressed as the mean ± standard deviation.
The statistical analysis was performed using Student’s t-test or
the Mann-Whitney U test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Upregulation of HSP27 phosphorylation
following exposure to 5-FU in colorectal cancer cells
The effect of 5-FU exposure on HSP27 expression or
phosphorylation was examined in colorectal cancer cells. Colorectal
cancer cells expressing high levels of HSP27 with a low sensitivity
to 5-FU (HCT116 and HCT15) and those expressing a low level of
HSP27 with a high sensitivity to 5-FU (HT29) (17) were used. Although the exposure of
the HCT116 and HCT15 cells to 5-FU exhibited a minimal effect on
HSP27 expression (Fig. 1A and B),
the exposure of the HT29 cells to 5-FU caused a marginal increase
in HSP27 expression (Fig. 1C).
Furthermore, exposure to 5-FU mainly upregulated the
phosphorylation of HSP27 at Ser78 in the HCT116 and HCT15 cells
(Fig. 1).
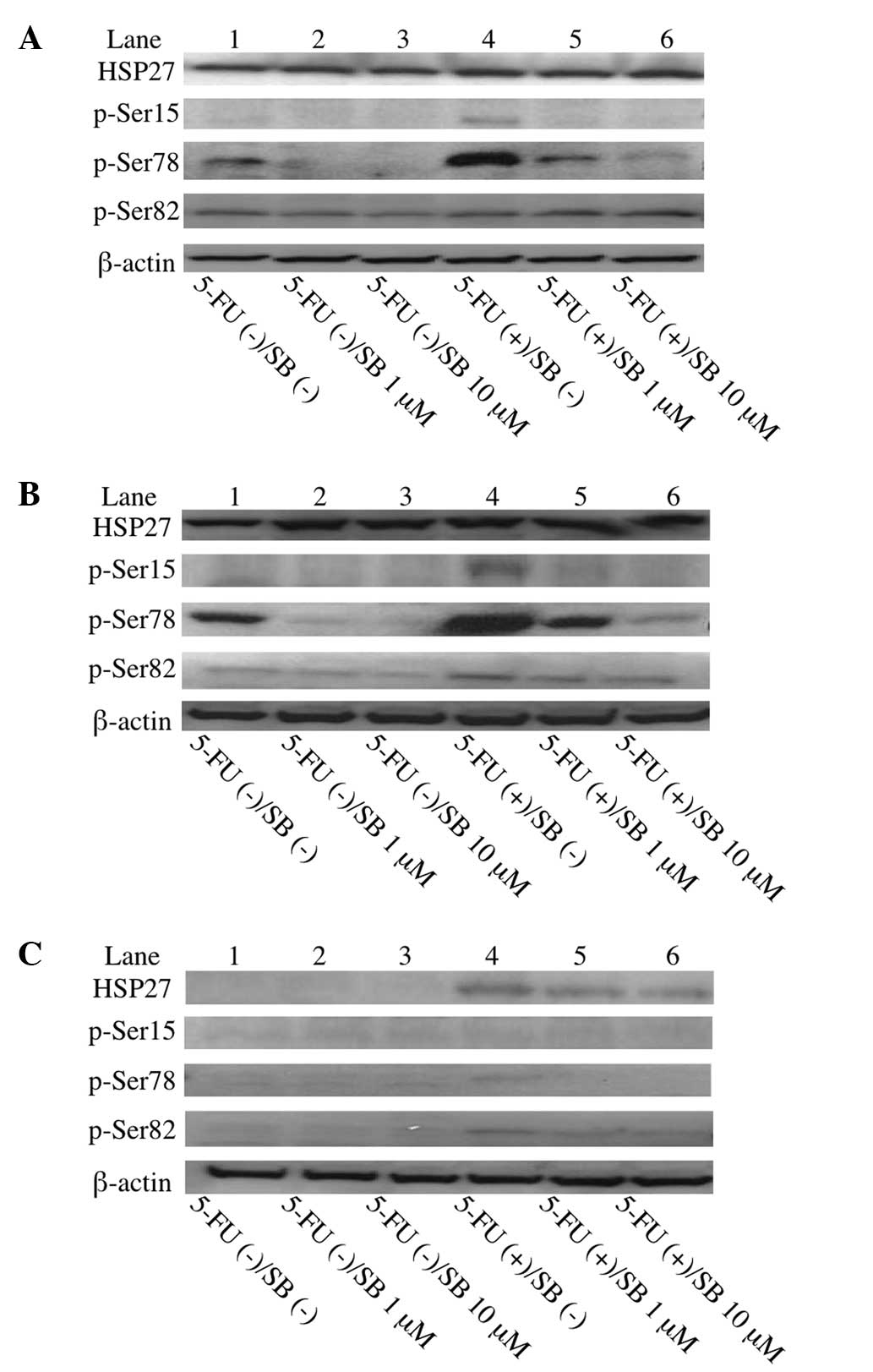 | Figure 1Western blot analysis of
phosphorylated HSP27 expression in colorectal cancer cells. 5-FU
exposure upregulated HSP27 phosphorylation, particularly Ser78, in
HCT116 and HCT15 cells with high levels of HSP27 expression, but
not in HT29 cells with a low level of HSP27 expression. SB203580
treatment dose-dependently inhibited HSP27 phosphorylation in
HCT116 and HCT15 cells. (A) HCT116; (B) HCT15; and (C) HT29. Lanes
1–3, no 5-FU treatment; lanes 4–6, 5-FU treatment; lanes 2 and 5,
treatment with 1 μM SB203580; and lanes 3 and 6, treatment with 10
μM SB203580. HSP, heat shock protein; 5-FU, 5-fluorouracil; SB,
SB203580. |
Inhibition of HSP27 phosphorylation by a
selective inhibitor of p38 MAPK (SB203580) in colorectal cancer
cells
The effect of SB203580, a p38 MAPK selective
inhibitor (20), on the
phosphorylation of HSP27 was investigated. Treatment with SB203580
attenuated the phosphorylatio of HSP27, particularly at Ser78,
without 5-FU exposure in the HCT116 and HCT15 cells. In addition,
SB203580 dose-dependently suppressed the phosphorylation of HSP27
at Ser78, which had been upregulated following exposure to 5-FU, in
the HCT116 and HCT15 cells (Fig. 1A and
B). These results indicated that the stress of 5-FU exposure
upregulates the phosphorylation of HSP27, particularly at Ser78,
via p38 MAPK in HCT116 and HCT15 cells.
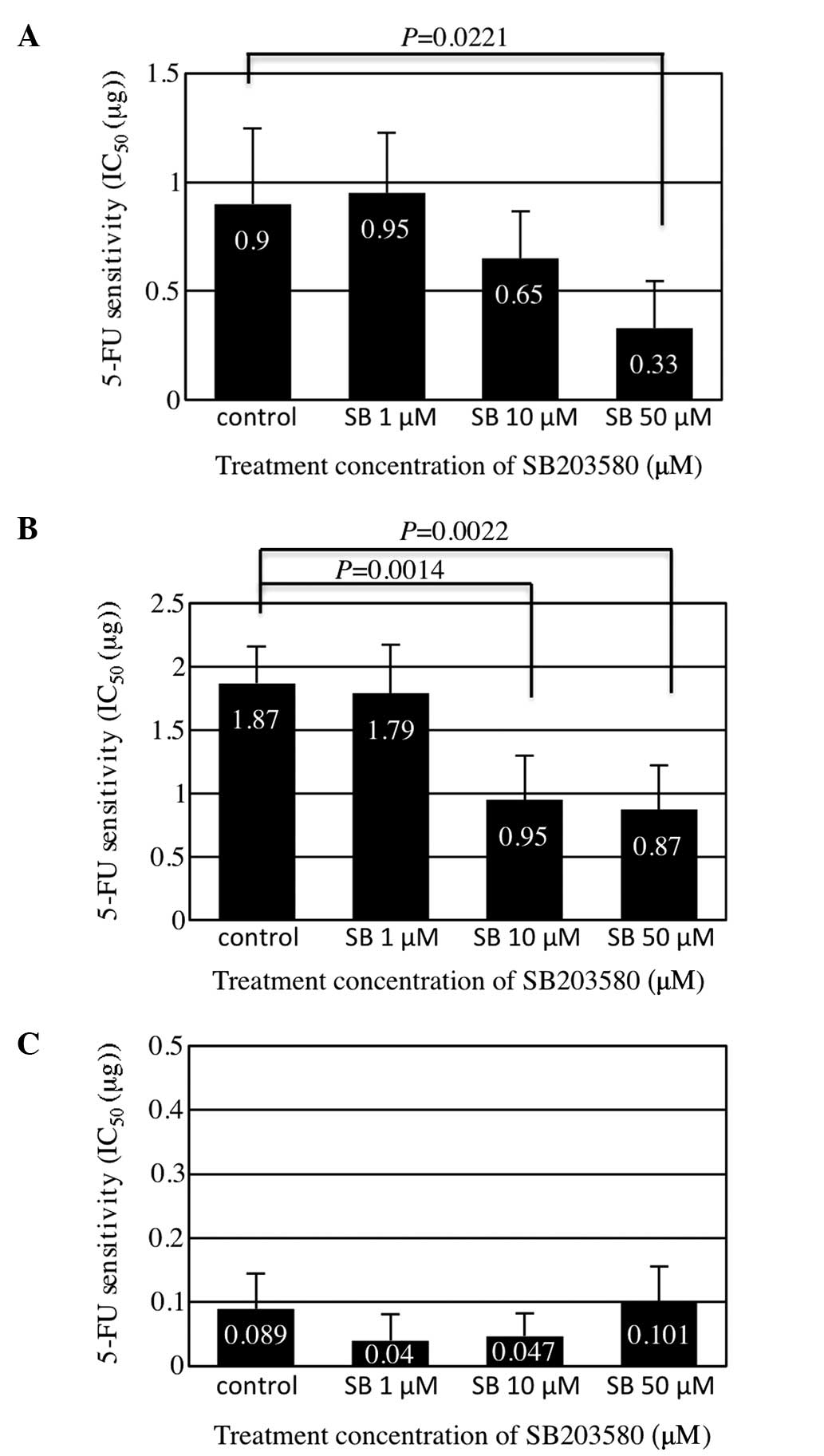
Promotion of 5-FU sensitivity by
inhibition of HSP27 phosphorylation in colorectal cancer cells
The inhibition of cell growth by 5-FU was analyzed
using the MTT assay and 5-FU sensitivity was determined using the
IC50 value. The HCT116 and HCT15 cells expressing high
levels of HSP27 exhibited high IC50 values, while the
HT29 cells expressing a low level of HSP27 exhibited a low
IC50 value (Fig. 2), as
previously reported (17).
Treatment with SB203580 significantly reduced the IC50
values of 5-FU in the HCT116 and HCT15 cells, which exhibited high
levels of HSP27 phosphorylation following 5-FU exposure (Fig. 2). These results indicated that the
inhibition of HSP27 phosphorylation by SB203580 promotes
sensitivity to 5-FU in colorectal cancer cells with high levels of
HSP27 expression.
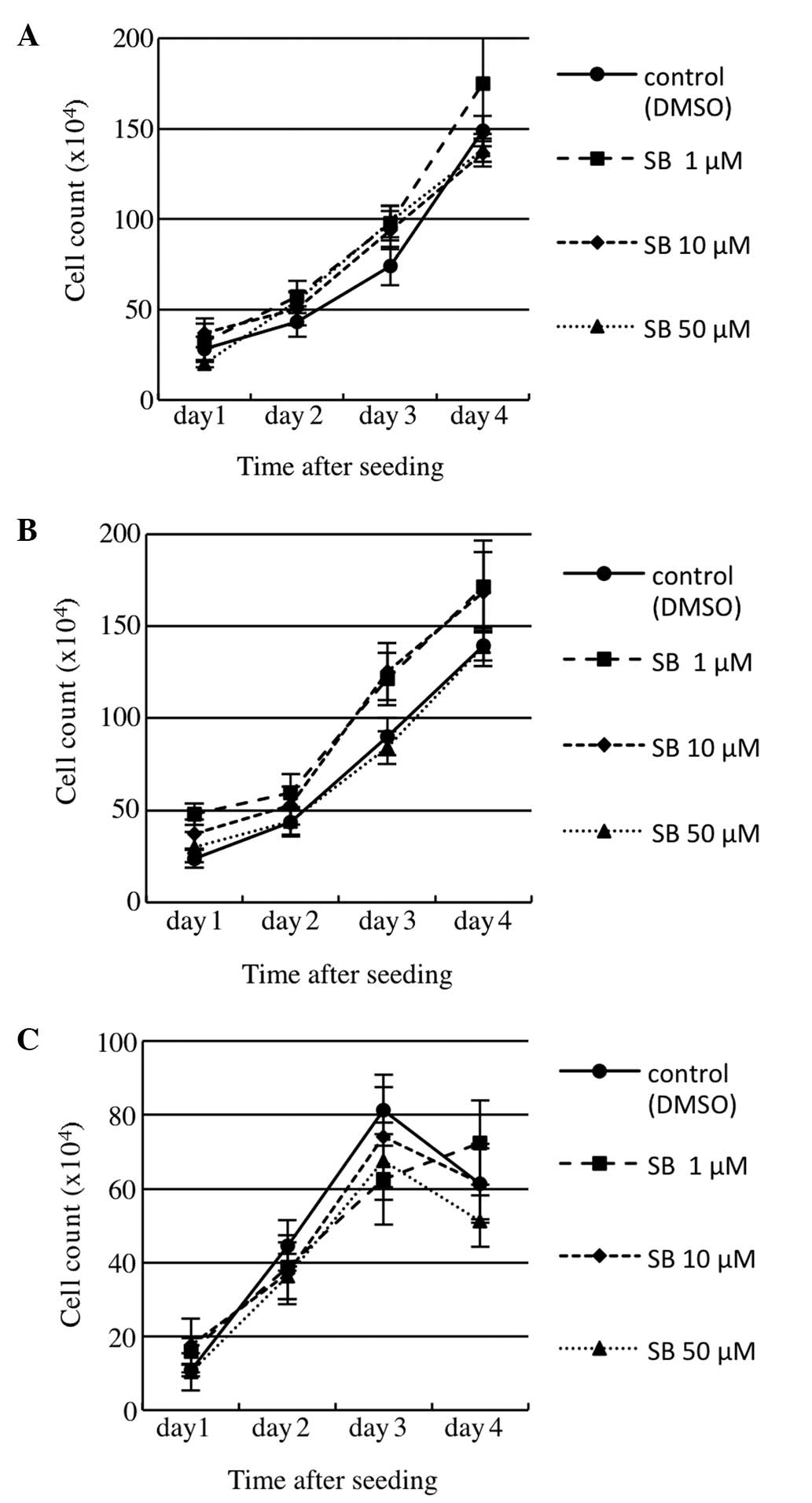
Effect of SB203580, a p38 MAPK selective
inhibitor, on colorectal cancer cell growth
To investigate the effect of SB203580 on cell growth
or survival, a cell growth assay was performed using a count of
viable cells. Three colorectal cancer cell lines were examined
following treatment with various concentrations of SB203580 (0, 1,
10 or 50 μM) in RPMI-1640 containing 5% FBS (Fig. 3). As a result, SB203580 treatment
exhibited no significant effect on cell growth or survival in the
cell lines.
Discussion
HSP27 is well-known as a stress-activated, adenosine
triphosphate-independent cytoprotective chaperone that is
associated with numerous functions, including the resistance to
chemotherapy. HSP27 has been reported to be a clinical prognostic
factor or as a resistant factor for the cytotoxic agent irinotecan
in colorectal cancer studies performed in vitro (13–16).
In addition, using in vitro and in vivo studies
performed with colorectal cancer cells, we previously demonstrated
that the overexpression of HSP27 reduced 5-FU sensitivity, while
the suppression of HSP27 expression promoted 5-FU sensitivity. In
addition, the promotion of 5-FU sensitivity by the suppression of
HSP27 expression resulted in the induction of apoptosis despite the
upregulation of 5-FU metabolism (17,18).
The functions of HSP27 are considered to be
regulated by post-transcriptional modifications, such as
phosphorylation (21,22). Human HSP27 is phosphorylated at
three sites, namely, Ser15, 78 and 82 (21). These serine residues have been
identified as sites for a kinase known as MAPKAPK-2 (23), and p38 MAPK is known to act as a
direct kinase for MAPKAPK-2 (24),
thus, p38 MAPK may phosphorylate HSP27. The phosphorylation of
HSP27 promotes the dissociation of large oligomers, induces nuclear
translocation and finally becomes involved in cellular protection
by HSP27 (25). However, the role
of HSP27 phosphorylation in chemoresistance or sensitivity remains
unclear.
Recently, HSP27 has been identified as a treatment
target for several cancers, and clinical trials using the antisense
oligonucleotide, OGX-427, which inhibits HSP27 expression, have
been performed in patients with prostate, bladder, ovarian, breast
or non-small cell lung cancer, but not colorectal cancer. This
therapy has been reported to be feasible and effective (26,27).
The inhibition of HSP27 expression would also be clinically
effective as a treatment for patients with 5-FU resistant
colorectal cancer, considering the results of our previous studies
(17,18).
In addition, the aim of the present study was to
verify that the inhibition of HSP27 phosphorylation may also
present an alternative target for the treatment of colorectal
cancer to promote 5-FU sensitivity or to reduce 5-FU resistance.
This study demonstrated that the stress of 5-FU exposure
upregulated the phosphorylation of HSP27, particularly at the Ser78
site, in the colorectal cancer cells with high levels of HSP27
expression (HCT116 and HCT15). In the colorectal cancer cells with
a low level of HSP27 expression (HT29), HSP27 expression was
marginally increased, with a small amount of phosphorylation of
HSP27 at the three serine residues. The phosphorylated residues of
HSP27 have been reported to differ in response to different
inducers (28). In these colorectal
cancer cells with high levels of HSP27 expression, Ser78 was
identified as the phosphorylated residue in the major
phosphorylated HSP27 form induced by 5-FU stimulation.
SB203580, a p38 MAPK selective inhibitor, clearly
inhibited the phosphorylation of the Ser78 residue of HSP27 induced
by 5-FU exposure. Furthermore, the inhibition of HSP27
phosphorylation by SB203580 significantly promoted the sensitivity
of the HCT116 and HCT15 cells to 5-FU. This effect may be caused by
a reduced anti-apoptotic effect as a result of the inactivation of
HSP27. In addition, SB203580 had a minimal effect on cell growth or
survival in vitro. This result indicates that the
dephosphorylation of HSP27 via the inhibition of p38 MAPK by
SB203580 exerted little toxicity in these colorectal cancer
cells.
In conclusion, the present study indicated that the
inhibition of HSP27 phosphorylation promotes 5-FU sensitivity in
colorectal cancer cells with high levels of HSP27 expression. If
the agents that specifically inhibit HSP27 phosphorylation exhibit
minimal toxicity, such agents may present a novel strategy for the
treatment of colorectal cancer when combined with current
chemotherapy using 5-FU. Further investigations into HSP27
regulation are important and are required for the development of
novel treatments targeting HSP27 in colorectal cancer.
References
|
1
|
Kinoshita M, Kodera Y, Hibi K, et al: Gene
expression profile of 5-fluorouracil metabolic enzymes in primary
colorectal cancer: potential as predictive parameters for response
to fluorouracil-based chemotherapy. Anticancer Res. 27:851–856.
2007.
|
|
2
|
Okumura K, Shiomi H, Mekata E, et al:
Correlation between chemosensitivity and mRNA expression level of
5-fluorouracil-related metabolic enzymes during liver metastasis of
colorectal cancer. Oncol Rep. 15:875–882. 2006.
|
|
3
|
Aschele C, Debernardis D, Casazza S, et
al: Immunohisto chemical quantitation of thymidylate synthase
expression in colorectal cancer metastases predicts for clinical
outcome to fluorouracil-based chemotherapy. J Clin Oncol.
17:1760–1770. 1999.
|
|
4
|
Soong R, Shah N, Salto-Tellez M, et al:
Prognostic significance of thymidylate synthase, dihydropyrimidine
dehydrogenase and thymidine phosphorylase protein expression in
colorectal cancer patients treated with or without
5-fluorouracil-based chemotherapy. Ann Oncol. 19:915–919. 2008.
|
|
5
|
Bruey JM, Ducasse C, Bonniaud P, et al:
Hsp27 negatively regulates cell death by interacting with
cytochrome c. Nat Cell Biol. 2:645–652. 2000.
|
|
6
|
Mehlen P, Schulze-Osthoff K and Arrigo AP:
Small stress proteins as novel regulators of apoptosis. Heat shock
protein 27 blocks Fas/APO-1-and staurosporine-induced cell death. J
Biol Chem. 271:16510–16514. 1996.
|
|
7
|
Andrieu C, Taieb D, Baylot V, et al: Heat
shock protein 27 confers resistance to androgen ablation and
chemotherapy in prostate cancer cells through eIF4E. Oncogene.
29:1883–1896. 2010.
|
|
8
|
Sarto C, Valsecchi C, Magni F, et al:
Expression of heat shock protein 27 in human renal cell carcinoma.
Proteomics. 4:2252–2260. 2004.
|
|
9
|
Song TF, Zhang ZF, Liu L, Yang T, Jiang J
and Li P: Small interfering RNA-mediated silencing of heat shock
protein 27 (HSP27) Increases chemosensitivity to paclitaxel by
increasing production of reactive oxygen species in human ovarian
cancer cells (HO8910). J Int Med Res. 37:1375–1388. 2009.
|
|
10
|
Ciocca DR, Fuqua SA, Lock-Lim S, Toft DO,
Welch WJ and McGuire WL: Response of human breast cancer cells to
heat shock and chemotherapeutic drugs. Cancer Res. 52:3648–3654.
1992.
|
|
11
|
Vargas-Roig LM, Gago FE, Tello O, Aznar JC
and Ciocca DR: Heat shock protein expression and drug resistance in
breast cancer patients treated with induction chemotherapy. Int J
Cancer. 79:468–475. 1998.
|
|
12
|
Garrido C, Mehlen P, Fromentin A, et al:
Inconstant association between 27-kDa heat-shock protein (Hsp27)
content and doxorubicin resistance in human colon cancer cells. The
doxorubicin-protecting effect of Hsp27. Eur J Biochem. 237:653–659.
1996.
|
|
13
|
Choi DH, Ha JS, Lee WH, et al: Heat shock
protein 27 is associated with irinotecan resistance in human
colorectal cancer cells. FEBS Lett. 581:1649–1656. 2007.
|
|
14
|
Tweedle EM, Khattak I, Ang CW, et al: Low
molecular weight heat shock protein HSP27 is a prognostic indicator
in rectal cancer but not colon cancer. Gut. 59:1501–1510. 2010.
|
|
15
|
Yu Z, Zhi J, Peng X, Zhong X and Xu A:
Clinical significance of HSP27 expression in colorectal cancer. Mol
Med Report. 3:953–958. 2010.
|
|
16
|
Wang F, Zhang P, Shi C, Yang Y and Qin H:
Immunohisto chemical detection of HSP27 and hnRNP K as prognostic
and predictive biomarkers for colorectal cancer. Med Oncol.
29:1780–1788. 2012.
|
|
17
|
Tsuruta M, Nishibori H, Hasegawa H, et al:
Heat shock protein 27, a novel regulator of 5-fluorouracil
resistance in colon cancer. Oncol Rep. 20:1165–1172. 2008.
|
|
18
|
Hayashi R, Ishii Y, Ochiai H, et al:
Suppression of heat shock protein 27 expression promotes
5-fluorouracil sensitivity in colon cancer cells in axenograft
model. Oncol Rep. 28:1269–1274. 2012.
|
|
19
|
Kostenko S and Moens U: Heat shock protein
27 phosphorylation: kinases, phosphatases, functions and pathology.
Cell Mol Life Sci. 66:3289–3307. 2009.
|
|
20
|
Cuenda A, Rouse J, Doza YN, et al:
SB203580 is a specific inhibitor of a MAP kinase homologue which is
stimulated by cellular stress and interleukin-1. FEBS Lett.
364:229–233. 1995.
|
|
21
|
Benjamin IJ and McMillan DR: Stress (heat
shock) proteins: molecular chaperones in cardiovascular biology and
disease. Circ Res. 83:117–132. 1998.
|
|
22
|
Lambert H, Charette SJ, Bernier AF, et al:
HSP27 multimerization mediated by phosphorylation-sensitive
intermolecular interactions at the amino terminus. J Biol Chem.
274:3978–9385. 1999.
|
|
23
|
Landry J, Lambert H, Zhou M, et al: Human
HSP27 is phosphorylated at serines 78 and 82 by heat shock and
mitogen-activated kinases that recognize the same amino acid motif
as S6 kinase II. J Biol Chem. 267:794–803. 1992.
|
|
24
|
Rouse J, Cohen P, Trigon S, et al: A novel
kinase cascade triggered by stress and heat shock that stimulates
MAPKAP kinase-2 and phosphorylation of the small heat shock
proteins. Cell. 78:1027–1037. 1994.
|
|
25
|
Geum D, Son GH and Kim K:
Phosphorylation-dependent cellular localization and
thermoprotective role of heat shock protein 25 in hippocampal
progenitor cells. J Biol Chem. 277:19913–19921. 2002.
|
|
26
|
Hotte SJ, Yu EY, Hirte HW, et al: Phase I
trial of OGX-427, a 2′methoxyethyl antisense oligonucleotide (ASO),
against heat shock protein 27 (Hsp27): Final results. J Clin Oncol.
28(15s): abstr. 3077. 2010.
|
|
27
|
Lamoureux F, Thomas C, Yin MJ, et al:
Suppression of heat shock protein 27 using OGX-427 induces
endoplasmic reticulum stress and potentiates heat shock protein 90
inhibitors to delay castrate-resistant prostate cancer. Eur Urol.
66:145–155. 2014.
|
|
28
|
Paul C, Simon S, Gilbert B, et al: Dynamic
pr Paul C, Simon S, Gilbert B, et al: Dynamic processes that
reflect anti-apoptotic strategies set up by HspB1 (Hsp27). Exp Cell
Res. 316:1535–1552. 2010.
|