Introduction
Neuroblastoma is the most common extracranial solid
cancer in childhood and the most common cancer in infancy, arising
from the developing sympathetic nervous system (1–3). The
prevalence of neuroblastoma is ∼1 case in 7,000 live births
(1–3).
Approximately 40% of neuroblastoma patients present with a
localized tumor, which appears as a large lump in the abdomen,
pelvis, chest or neck, that may press on other parts of the body
(4). The disease exhibits extreme
heterogeneity, and is stratified into three risk categories, low,
intermediate and high risk (3). These
risk catergories are based on clinical and biological features,
including MYCN copy number, histopathology, tumor ploidy in
infants, patient age and tumor stage [according to the
International Neuroblastoma Staging System (5)] (3). The
risk categories are used to predict how likely it is that a child
can be cured (6) and to stratify
patients in clinical trials to determine the appropriate type and
intensity of therapy (5). A
higher-risk group indicates that there is less chance of a cure,
and thus more intensive treatments are often required (5). Approximately 50% of patients exhibit
metastasis to the lymph nodes, bone, bone marrow and liver,
presenting with syndromes including bone pain and bone marrow
failure (4). Occasionally, patients
may exhibit renin-mediated hypertension (4). Notably, metastasis of neuroblastoma
occurs prior to the appearance of any symptoms in 50–60% of all
neuroblastoma cases (7). A rare type
of neuroblastoma, termed stage 4S, that accounts for 5% of
neuroblastoma cases, presents as small localized primary tumors
with metastases to the liver, skin or bone marrow, which usually
regresses spontaneously (8). The
majority of cases of stage 4S neuroblastoma are diagnosed in
children aged <1 year old (5,9). Certain
neuroblastoma patients have an excellent prognosis, such as those
with stage 4S neuroblastoma (10),
however, older children usually exhibit metastatic disease and thus
have a worse prognosis. The overall five-year survival rate for
neuroblastoma patients is 59%, ranging from 47% (Eastern
populations) to 67% (Western populations) (2). The majority of neuroblastomas are
treated using conventional approaches, including surgery, radiation
and chemotherapy (3). Although the
success rate of current therapies for the treatment of patients
with low- and intermediate-risk neuroblastoma is >90 and 70–90%,
respectively, only ∼30% of high-risk neuroblastoma patients have
been successfully treated in the previous two decades (11–13).
Therefore, research into novel agents that are able to more
effectively treat neuroblastoma is necessary. At present,
therapeutic approaches that exhibit less toxicity and increased
specificity are being investigated. For example, 13-cis retinoic
acid, which induces differentiation of neuroblastoma cells, has
become a standard treatment for the management of high-risk
neuroblastoma patients following marrow or stem cell
transplantation (14). Other
potential treatment modalities for neuroblastoma include drugs that
target apoptosis or inhibit angiogenesis, targeted radiation
therapy (3,4) and immunotherapy (12).
Cordycepin, also termed 3′-deoxyadenosine,
originates from Ophiocordyceps sinensis, which is commonly
used in Traditional Chinese Medicine (15). Cordycepin is an adenosine analogue
that differs from adenosine by the deletion of oxygen in the 3′
position of the ribose region (16).
Therefore, certain enzymes are unable to discriminate between the
two. Cordycepin may be phosphorylated to mono-, di- and
triphosphate forms within cells, and participates in a number of
biochemical reactions (16). For
example, triphosphate cordycepin may be incorporated into RNA
molecules, leading to the premature termination of RNA synthesis
and inhibition of transcription elongation (17). Cordycepin is an example of a proposed
new class of anticancer drugs that target RNA synthesis (18).
Previously, cordycepin has been revealed to induce
apoptosis in a number of cancer cell lines, including breast
(19), colon (20) and oral cancers (21), mouse leydig tumor (22), and oral squamous cell carcinoma
(23). Cordycepin also induces
autophagy in breast cancer cells (19) and has been used in a phase I study in
combination with pentostatin to treat refractory acute lymphocytic
or chronic myelogenous leukemia (Study ID, CDR0000065572) (24). However, the effects of cordycepin on
neuroblastoma cells have not yet been examined.
The present study investigates the apoptotic and
autophagic effects of cordycepin on the human neuroblastoma cell
lines SK-N-SH and BE(2)-M17, in order to assess its feasibility as
a drug candidate for neuroblastoma therapy.
Materials and methods
Chemicals
Hirsutella sinensis extract (0.199 g/ml),
which was produced by fermentation and extraction using ethanol,
water and enzyme digestion methods by Huadong Medicine (Hangzhou,
Zhejiang, China), was provided by Dr Shenglang Zhu at Nanshan
Hospital (Shenzhen, Guangdong, China). Hirsutella sinensis
is the anamorphic, mycelial form of Cordyceps sinensis
(25,26). Cordycepin (catalog no., C3394) and
3-methyladenine (catalog no., M9281) were purchased from
Sigma-Aldrich (St. Louis, MO, USA).
Cell lines and cell culture
The human neuroblastoma cell line SK-N-SH was
obtained from the Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Word Federation for Culture Collection
registered no., 793; Shanghai, China). The human neuroblastoma cell
line BE(2)-M17 was from the Cell Resource Center (Institute of
Basic Medical Sciences, Chinese Academy of Medical Sciences &
Peking Union Medical College, Beijing, China). The cells were
cultured using standard methods (27)
in Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, UT,
USA) supplemented with 10% fetal bovine serum (FBS; Gibco,
Carlsbad, CA, USA), 2 mmol/l L-glutamine (Sigma-Aldrich), 100
units/ml penicillin and 100 µg/ml streptomycin (Gibco), at 37°C in
a 5% CO2 atmosphere.
Vectors and transfection
The pEGFP-LC3 plasmid (plasmid no., 21073; Addgene,
Inc., Cambridge, MA, USA) is a pEGFP-C1 plasmid (Clontech
Laboratories, Inc., Mountainview, CA, USA) inserted with a 738-bp
long cDNA sequence from rat microtubule-associated protein 1 light
chain 3 (LC3). This sequence encodes a fusion protein composed of
green fluorescent protein (GFP) at the N-terminus and LC3 at the
C-terminus (28). In total,
1×106 SK-N-SH cells/well were seeded into six-well
plates, 24 h prior to transfection. The SK-N-SH cells were
transfected with pEGFP-LC3 using FuGENE HD Transfection Reagent
(Promega Corporation, Madison, WI, USA) and left for 48 h, followed
by selection with 800 µg/ml G418 (Amresco, Inc., Framingham, MA,
USA) for one week.
Cell morphology study
The SK-N-SH or BE(2)-M17 cells were seeded at a
density of 1×106 cells/well into six-well plates 24 h
prior to treatment. SK-N-SH cells were treated with various
concentrations of Cordyceps sinensis extract. The SK-N-SH
and BE(2)-M17 cells were treated with various concentrations of
cordycepin. The morphology of each cell line was analyzed using an
Axiovert 40 microscope (Zeiss, Oberkochen, Germany).
MTT assay
The SK-N-SH or BE(2)-M17 cells were seeded at a
density of 104 cells/well in 96-well plates, 24 h prior
to treatment. At 48 h subsequent to cordycepin treatment, the
medium was removed and 100 µl of serum free DMEM containing 0.5
mg/ml MTT (Sigma-Aldrich) was added to each well. The cells were
then incubated at 37°C for 4 h. Subsequent to the removal of the
MTT media, 100 µl dimethyl sulfoxide was added to each well and
incubated at 37°C for 20 min. The plate was incubated at room
temperature for an additional 5 min with agitation, and the
absorbance of the solution was subsequently quantified at a
wavelength of 570 nm, which was normalized against a reference
value measured at a wavelength of 690 nm to account for debris,
using the PARADIGM Detection Platform (Beckman Coulter, Brea, CA,
USA).
Fluorescence-activated cell sorting
(FACS)
The SK-N-SH cells were cultured in DMEM supplemented
with 10% FBS, 2 mmol/l L-glutamine, 100 units/ml penicillin and 100
µg/ml streptomycin at 37°C in a 5% CO2 atmosphere. The
SK-N-SH cells (5×105) were seeded into each well of a
six-well plate and grown overnight, which was followed by treatment
with 0, 50, 100 or 200 µmol/l cordycepin for 48 h. For the
experiment measuring the effect of 3-methyladenine on
cordycepin-induced apoptosis, SK-N-SH cells were treated with 0 or
150 µmol/l cordycepin, 5 mmol/l 3-methyladenine, or combination of
the latter two, for 48 h. Untreated cells were used as the negative
control. The free-floating and attached SK-N-SH cells from each
well were harvested and pooled. The cells were washed with
phosphate buffered saline (PBS) and stained using a Dead Cell
Apoptosis kit with Annexin V Alexa Fluor 488 and propidium iodide
(PI) (Invitrogen Life Technologies, Carlsbad, CA, USA), according
to the manufacturer's instructions. The stained cells were analyzed
using a Cytomics FC 500 flow cytometer (Beckman Coulter).
Western blot analysis
The antibodies against caspase-3 (catalog no.,
9665S), poly(adenosine diphosphate-ribose) polymerase 1 (PARP1;
catalog no., 9542S) and LC3 A/B (catalog no., 4108S) were purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA). The
anti-β-actin antibody (catalog no., sc-47778) was purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA), and the
whole-molecule anti-rabbit immunoglobulin (Ig)G-peroxidase (cat.
no. A9169) and whole-molecule anti-mouse IgG-peroxidase antibodies
(cat. no. A9044) were purchased from Sigma-Aldrich.
In total, 1×106 cells were lysed in 150
µl cell lysis buffer (Cell Signaling Technology) containing an
EDTA-free protease inhibitor cocktail (Roche Diagnostics, Basel,
Switzerland). The protein concentrations were determined using a
detergent-compatible protein assay kit (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). For each well, 30 µg of protein lysate was
combined with 2 µl 2X sample buffer, comprising final
concentrations of 60 mmol/l Tris (pH, 6.8), 10% glycerol, 2% sodium
dodecyl sulfate and 5% mercaptoethanol. The lysate was then
incubated at 100°C for 5 min to denature the proteins. The protein
samples were loaded for electrophoresis, and then transferred to
Immobilon-P membranes (Millipore, Billerica, MA, USA) using wet
transfer apparatus. The membrane was blocked in 1% bovine serum
albumin (Sigma-Aldrich) in PBS for 2 h at room temperature. The
membranes were subsequently probed with primary antibodies
overnight at 4°C, followed by secondary antibody incubation at room
temperature for 1.5 h. The protein bands were visualized with
enhanced chemiluminescence prime western blotting detection reagent
(Amersham, GE Healthcare, Shanghai, China) and images were captured
using the MiniChemi professional machine (SageCreation Science,
Co., Ltd., Beijing, China).
Statistical analysis
Student's t-test was used to compare the
viability of SK-N-SH and BE(2)-M17 cells measured by MTT assay.
Statistical analyses were performed using Microsoft Excel 2010
software (Microsoft, Redmond, WA, USA). The results are presented
as the mean ± standard error of the mean. All P-values were
two-tailed and P<0.05 was considered to indicate a statistically
significant difference.
Results
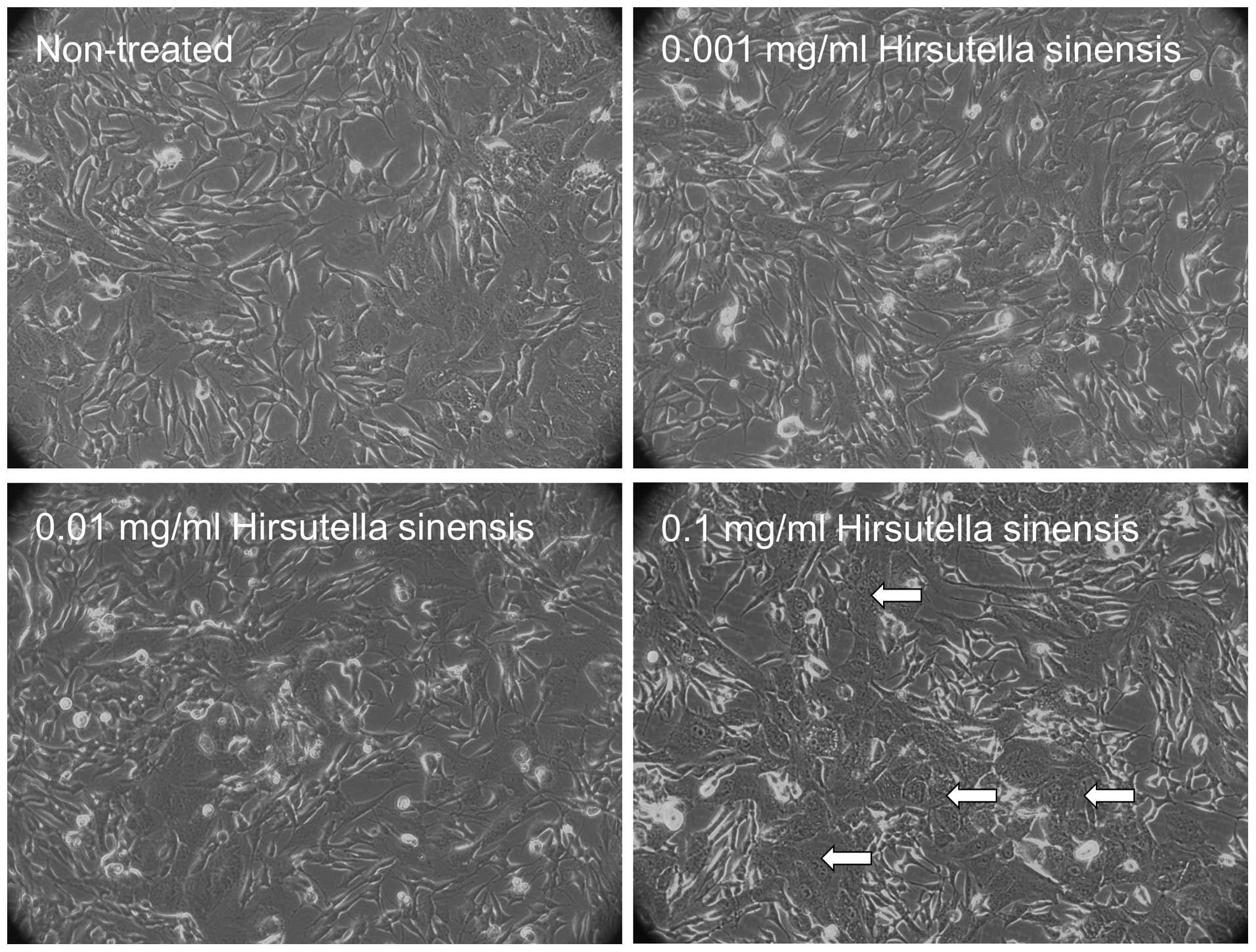
Morphological changes in neuroblastoma
cells treated with Hirsutella sinensis extract
To examine the effects of Hirsutella sinensis
extract on neuroblastoma cells, various concentrations of the
extract, comprising 0, 0.001, 0.01 and 0.1 mg/ml, were incubated
with SK-N-SH cells for 48 h. Administration of 0.1 mg/ml
Hirsutella sinensis extract markedly altered the cell
phenotype, as indicated by an increased proportion of SK-N-SH cells
possessing cell granulations that were visible under a
phase-contrast microscope, compared with the untreated control
(Fig. 1). Administration of 0.001 and
0.01 mg/ml Hirsutella sinensis extract resulted in no
significant effect on the phenotype of the SK-N-SH cells (Fig. 1).
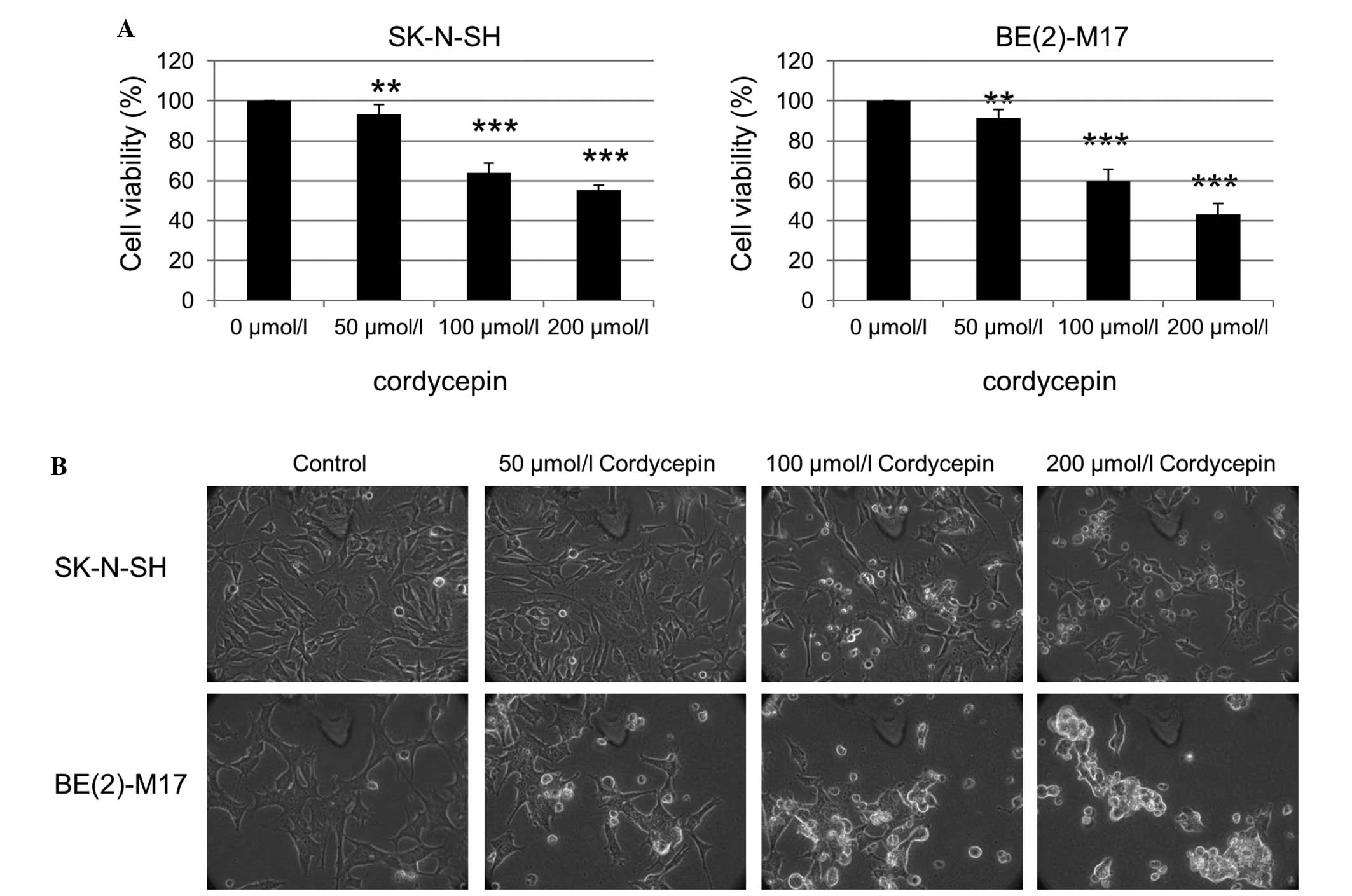
Viability inhibition of neuroblastoma
cells by cordycepin
The effect of the purified cordycepin chemical, one
of the bioactive components of Cordyceps sinensis, on the
neuroblastoma cells was also investigated. The SK-N-SH cells were
treated with 0, 50, 100 and 200 µmol/l cordycepin for 48 h, and the
cell viability was assessed using an MTT assay. The viability of
the SK-N-SH and BE(2)-M17 cells was significantly reduced as the
cordycepin concentration increased (Fig.
2A). Examination of the SK-N-SH and BE(2)-M17 cells by phase
contrast microscopy also revealed an increased number of floating
cells and reduced the number of attached cells following treatment
with cordycepin, compared with that of non-treated cells (Fig. 2B).
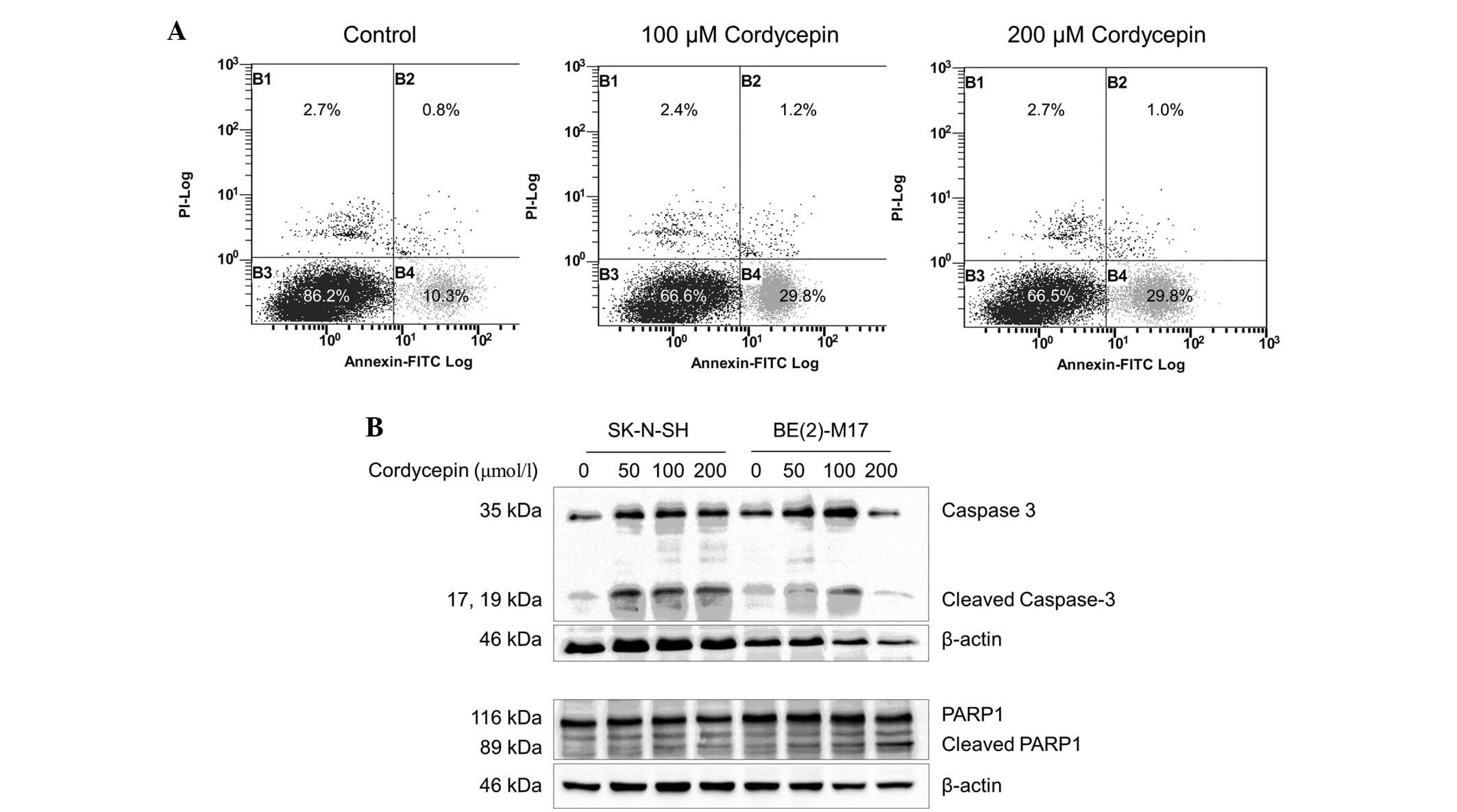
Cordycepin induces apoptosis in
BE(2)-M17 and SK-N-SH cells
In order to investigate whether apoptosis occurs in
cordycepin-treated neuroblastoma cells, SK-N-SH cells were
incubated with or without cordycepin and analyzed by FACS following
staining with Annexin V-PI. The percentage of early-apoptotic cells
in the SK-N-SH cell populations treated with 100 or 200 µmol/l
cordycepin was significantly increased (29.8%), compared with the
percentage of early-apoptotic cells (10.3%) in the negative control
(Fig. 3A). As BE(2)-M17 cells tend to
form clusters and do not separate well, apoptosis in this cell line
was not assessed by FACS.
The expression of the apoptosis markers caspase-3
and PARP1 was examined by western blot analysis (29). As shown in Fig. 3B, the expression of the 35 kDa full
length caspase-3 protein and its cleavage products were upregulated
in the SK-N-SH cells treated with 50, 100 and 200 µmol/l
cordycepin, compared with the expression in the non-treated control
cells. Similar results were obtained from the BE(2)-M17 cells
treated with 50 or 100 µmol/l cordycepin. However, in the BE(2)-M17
cells treated with 200 µmol/l cordycepin, caspase-3 upregulation
and cleavage was decreased.
In the SK-N-SH and BE(2)-M17 cells, expression of
full-length PARP1 was not altered by cordycepin at concentrations
up to 200 µmol/l. A significant increase in cleavage of PARP1 was
observed in SK-N-SH cells treated with 50, 100 or 200 µmol/l
cordycepin for 48 h, compared with untreated cells
(P<0.001)(Fig. 3B). In the
BE(2)-M17 cells treated with 50, 100 or 200 µmol/l cordycepin,
PARP1 cleavage increased in a dose-dependent manner, compared with
the control samples (Fig. 3B).
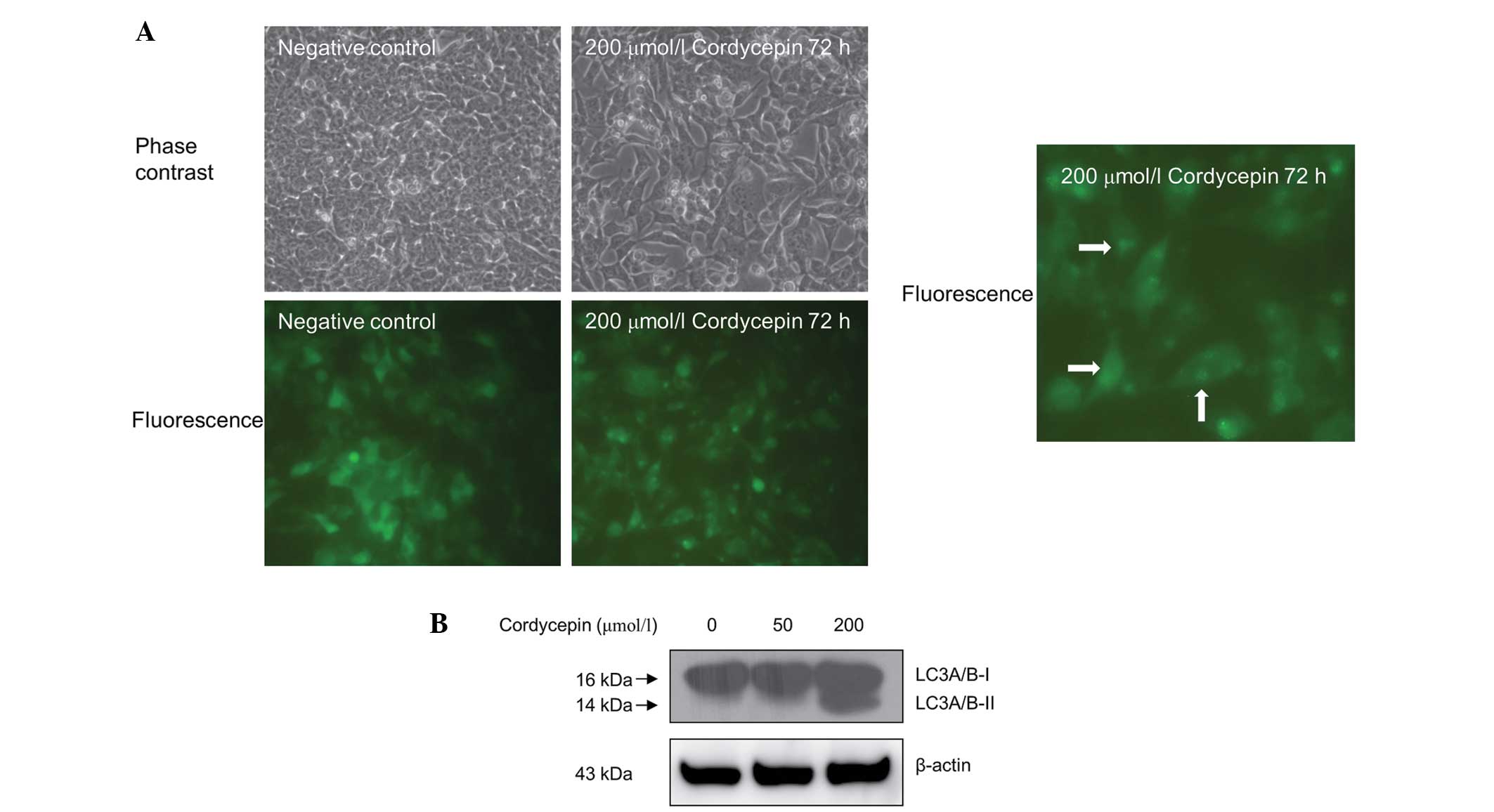
Cordycepin induces autophagy in
SK-N-SH cells
The effect of cordycepin treatment on autophagy in
the neuroblastoma SK-N-SH cells was assessed using fluorescence
microscopy. As shown in Fig. 4A, the
SK-N-SH cells that were stably transfected with pEGFP-LC3 and
treated with 200 µmol/l cordycepin for 72 h exhibited a marked
shift from a diffuse to a punctate pattern of LC3-associated green
fluorescence (Fig. 4A), indicating
that autophagy occurs in cordycepin-treated SK-N-SH cells.
Western blot analysis also revealed increased LC3
A/B protein cleavage in the SK-N-SH cells treated with 200 µmol/l
cordycepin compared with the untreated controls (Fig. 4B). However, treatment with 40 µmol/l
of cordycepin exhibited no significant effect on autophagy in
SK-N-SH cells (Fig. 4B).
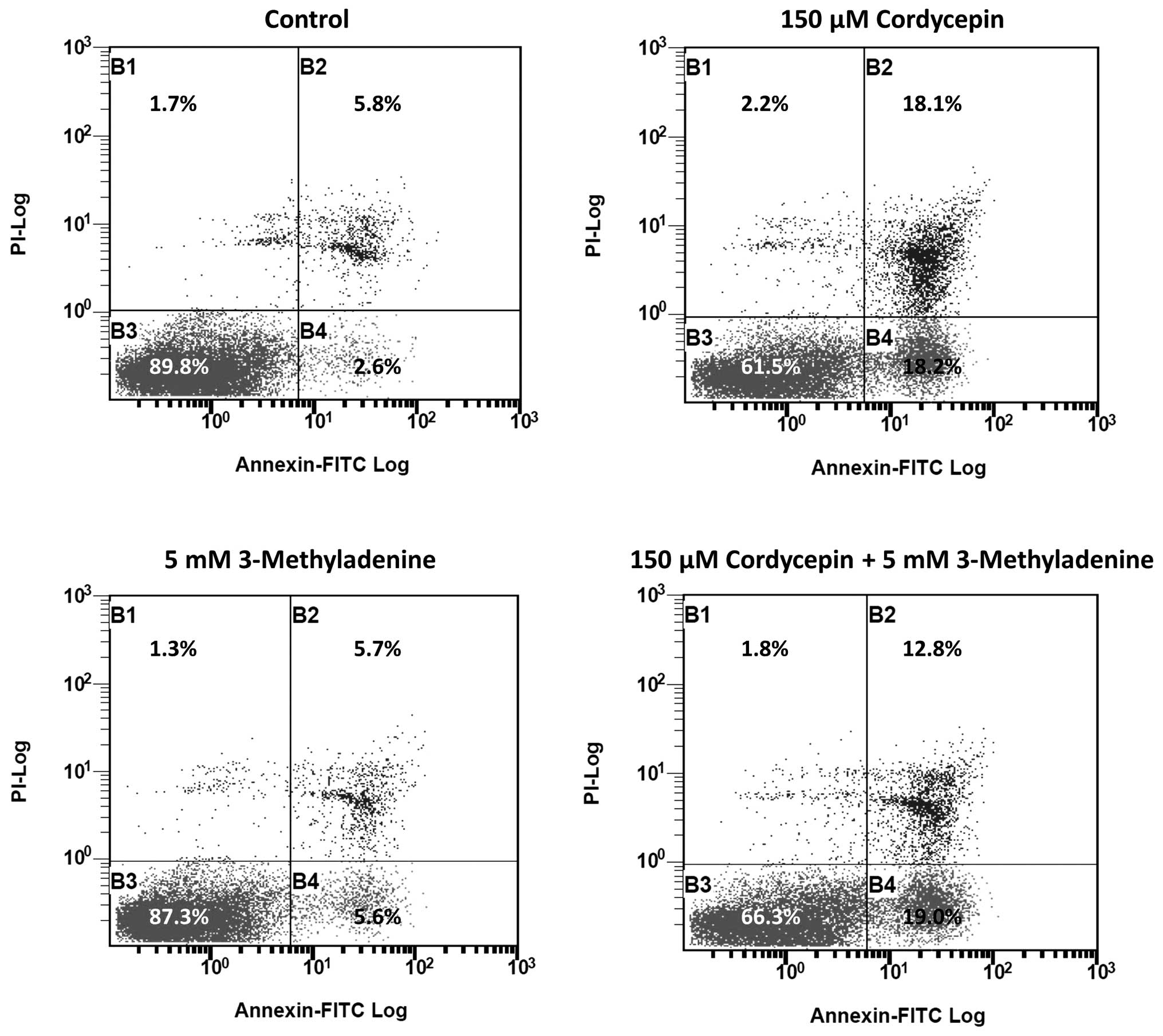
3-methyladenine was previously reported to be an
autophagy inducer under nutrient-rich conditions (13), which may negatively affect apoptosis.
Thus, we investigated the effect of 3-methyladenine on
cordycepin-induced apoptosis. The results of FACS revealed that the
percentage of late-apoptotic cells in SK-N-SH cell populations
treated with 150 µmol/l cordycepin and 5 mM 3-methyladenine was
significantly reduced (12.8%), compared with the percentage of
late-apoptotic cells (18.1%) in SK-N-SH cells treated with 150
µmol/l cordycepin alone (Fig. 5).
Discussion
The results of the present study indicate that
cordycepin, also termed 3′-deoxyadenosine, induces apoptosis in
neuroblastoma cells. Previous studies have demonstrated that
polysaccharide-peptide complexes produced by the submerged mycelial
culture of an entomopathogenic fungus, Cordyceps
sphecocephala, may induce apoptosis in SK-N-SH cells (30). However, the particular components that
induced apoptosis remain unclear. To the best of our knowledge, the
current study presents the first evidence that the cordycepin
chemical alone is able to induce apoptosis in neuroblastoma
cells.
Although cordycepin has been reported to induce
autophagy in breast cancer cells (19), evidence for cordycepin-induced
autophagy in cancer cells remains rare. The present study
demonstrated a shift between diffuse and punctate patterns of
LC3-associated green fluorescence (Fig.
4A), in addition to an increase in LC3 A/B cleavage (Fig. 4B) in cordycepin-treated neuroblastoma
cells compared with the untreated controls. These results indicate
that autophagy occurs in neuroblastoma cells treated with
cordycepin. It has been reported that Cordyceps militaris
and mycelial fermentation induce apoptosis and autophagy in human
glioblastoma cells (31). However,
whether Cordyceps militaris and mycelial fermentation induce
autophagy in neuroblastoma cells remains unclear. In addition, the
particular component of Cordyceps militaris and mycelial
fermentation that induces autophagy has not been previously
reported.
The investigation into the effect of cordycepin on
the cleavage of LC3 (Fig. 4B)
indicated that low concentrations of cordycepin (50 µmol/l) are
able to induce apoptosis but have little effect on autophagy.
However, apoptosis and autophagy occurred in SK-N-SH cells treated
with cordycepin at a high concentration (200 µmol/l), suggesting
that this concentration increases stress in neuroblastoma cells,
which respond by initiating autophagy for survival. Previously,
3-methyladenine has been demonstrated to perform a dual role in
autophagy; 3-methyladenine has been shown to inhibit
starvation-induced autophagy, however, it may also promote
autophagic flux following treatment for a prolonged period under
nutrient-rich conditions, based on the inhibition of PI3K-III
(32). Using FACS analysis, the
present study revealed that 3-methyladenine treatment led to a
reduction in the proportion of SK-N-SH cells undergoing
intermediate-stage cordycepin-induced apoptosis, between 18.1 and
12.8% (Fig. 5). This observation is
consistent with previous studies, which indicated that the
inhibition of autophagy triggers apoptosis (33), and therefore reinforces the hypothesis
that autophagy is a cytoprotective process under physiological
conditions.
The results of the present study showed that
cordycepin induces the apoptosis of neuroblastoma SK-N-SH and
BE(2)-M17 cells. Furthermore, the study revealed that treatment
with cordycepin induces autophagy, as well as apoptosis, in
neuroblastoma cells, in a concentration-dependent manner. This
suggests that autophagy may counteract apoptosis and serve as a
cell survival mechanism. Furthermore, 3-methyladenine, a promoter
of autophagy at full growth medium conditions, inhibited cordycepin
induced apoptosis of SK-N-SH cells, which indicated that apoptosis
and autophagy are inversely correlated. Future studies, which
investigate the inhibition of autophagy following treatment with
cordycepin, may improve the therapeutic efficacy of cordycepin
neuroblastoma treatment.
Acknowledgements
The authors would like to thank Professor Cho Chi
Hin and Dr Lin Zhang at the Chinese University of Hong Kong for
kindly providing the pEGFP-LC3 vector. This study was funded by the
National Natural Science Foundation of China (grant no., 31100943),
Shenzhen City Science and Technology Project (grant no., 201102136)
and Shenzhen Nanshan Science and Technology Research Fund (grant
no., 2010012).
References
|
1
|
Gurney JG, Ross JA, Wall DA, Bleyer WA,
Severson RK and Robison LL: Infant Cancer in the U.S.:
histology-specific incidence and trends, 1973 to 1992. J Pediatr
Hematol Oncol. 19:428–432. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Spix C, Pastore G, Sankila R, Stiller CA
and Steliarova-Foucher E: Neuroblastoma incidence and survival in
European children (1978–1997): report from the Automated Childhood
Cancer Information System project. Eur J Cancer. 42:2081–2091.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brodeur GM: Neuroblastoma: biological
insights into a clinical enigma. Nat Rev Cancer. 3:203–216. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brodeur GM, Pritchard J, Berthold F, et
al: Revisions of the international criteria for neuroblastoma
diagnosis, staging, and response to treatment. J Clin Oncol.
11:1466–1477. 1993.PubMed/NCBI
|
|
6
|
National Cancer Society: Neuroblastoma
risk groups. http://www.cancer.org/cancer/neuroblastoma/detailedguide/neuroblastoma-risk-groupsAccessed.
February 16–2015.
|
|
7
|
Al-Salem AH: An Illustrated Guide to
Pediatric Surgery. Springer International Publishing AG.
Switzerland: 2014. View Article : Google Scholar
|
|
8
|
D'Angio GJ, Evans AE and Koop CE: Special
pattern of widespread neuroblastoma with a favourable prognosis.
Lancet. 1:1046–1049. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brodeur GM, Seeger RC, Barrett A, et al:
International criteria for diagnosis, staging, and response to
treatment in patients with neuroblastoma. J Clin Oncol.
6:1874–1881. 1988.PubMed/NCBI
|
|
10
|
Diede SJ: Spontaneous regression of
metastatic cancer: learning from neuroblastoma. Nat Rev Cancer.
14:71–72. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hallett A and Traunecker H: A review and
update on neuroblastoma. Paediatrics and Child Health. 22:103–107.
2012. View Article : Google Scholar
|
|
12
|
Cheung NK and Dyer MA: Neuroblastoma:
developmental biology, cancer genomics and immunotherapy. Nat Rev
Cancer. 13:397–411. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maris JM: Recent advances in
neuroblastoma. N Engl J Med. 362:2202–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matthay KK, Villablanca JG, Seeger RC, et
al: Treatment of high-risk neuroblastoma with intensive
chemotherapy, radiotherapy, autologous bone marrow transplantation,
and 13-cis-retinoic acid. N Engl J Med. 341:1165–1173. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kondrashov A, Meijer HA, Barthet-Barateig
A, et al: Inhibition of polyadenylation reduces inflammatory gene
induction. RNA. 18:2236–2250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tuli HS, Sharma AK, Sandhu SS and Kashyap
D: Cordycepin: a bioactive metabolite with therapeutic potential.
Life Sci. 93:863–869. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Siev M, Weinberg R and Penman S: The
selective interruption of nucleolar RNA synthesis in HeLa cells by
cordycepin. J Cell Biol. 41:510–520. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hollerer I, Grund K, Hentze MW and Kulozik
AE: mRNA 3′end processing: A tale of the tail reaches the clinic.
EMBO Mol Med. 6:16–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Choi S, Lim MH, Kim KM, Jeon BH, Song WO
and Kim TW: Cordycepin-induced apoptosis and autophagy in breast
cancer cells are independent of the estrogen receptor. Toxicol Appl
Pharmacol. 257:165–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee SY, Debnath T, Kim SK and Lim BO:
Anti-cancer effect and apoptosis induction of cordycepin through
DR3 pathway in the human colonic cancer cell HT-29. Food Chem
Toxicol. 60:439–447. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen YH, Hao LJ, Hung CP, Chen JW, Leu SF
and Huang BM: Apoptotic effect of cisplatin and cordycepin on OC3
human oral cancer cells. Chin J Integr Med. 20:624–632. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jen CY, Lin CY, Huang BM and Leu SF:
Cordycepin Induced Ma-10 Mouse Leydig Tumor Cell Apoptosis through
Caspase-9 Pathway. Evid Based Complement Alternat Med.
2011.9845372011.PubMed/NCBI
|
|
23
|
Wu WC, Hsiao JR, Lian YR, Lin CY and Huang
BM: The apoptotic effect of cordycepin on human OEC-M1 oral cancer
cell line. Cancer Chemother Pharmacol. 60:103–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
National Cancer Institute: Chemotherapy
with cordycepin plus pentostatin in treating patients with
refractory acute lymphocytic or chronic myelogenous leukemia.
http://www.cancer.gov/clinicaltrials/search/view?cdrid=65572&version=healthprofessionalAccessed.
June 10–2014.
|
|
25
|
Huang TT, Chong KY, Ojcius DM, et al:
Hirsutella sinensis mycelium suppresses interleukin-1β and
interleukin-18 secretion by inhibiting both canonical and
non-canonical inflammasomes. Sci Rep. 3:13742013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li C, Li Z, Fan M, et al: The composition
of Hirsutella sinensis, anamorph of Cordyceps sinensis. J Food
Compost Anal. 19:800–805. 2006. View Article : Google Scholar
|
|
27
|
Szemes M, Dallosso AR, Melegh Z, et al:
Control of epigenetic states by WT1 via regulation of de novo DNA
methyltransferase 3A. Hum Mol Genet. 22:74–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kabeya Y, Mizushima N, Ueno T, et al: LC3,
a mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Elmore S: Apoptosis: a review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oh JY, Baek YM, Kim SW, et al: Apoptosis
of human hepatocarcinoma (HepG2) and neuroblastoma (SKN-SH) cells
induced by polysaccharides-peptide complexes produced by submerged
mycelial culture of an entomopathogenic fungus Cordyceps
sphecocephala. J Microbiol Biotechnol. 18:512–519. 2008.PubMed/NCBI
|
|
31
|
Yang CH, Kao YH, Huang KS, Wang CY and Lin
LW: Cordyceps militaris and mycelial fermentation induced apoptosis
and autophagy of human glioblastoma cells. Cell Death Dis.
3:e4312012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu YT, Tan HL, Shui G, et al: Dual role of
3-methyladenine in modulation of autophagy via different temporal
patterns of inhibition on class I and III phosphoinositide
3-kinase. J Biol Chem. 285:10850–10861. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boya P, González-Polo RA, Casares N, et
al: Inhibition of macroautophagy triggers apoptosis. Mol Cell Biol.
25:1025–1040. 2005. View Article : Google Scholar : PubMed/NCBI
|