Introduction
Caveolin-1 (Cav) is an important protein component
of caveolae, a specialized type of membrane lipid raft, which are
involved in multiple cellular processes, such as molecular
transport, cell adhesion and signal transduction (1,2). Recently,
Cav has been shown to be overexpressed in prostate cancer, and to
promote the growth and metastasis of this tumor (3–6). Cav has
been demonstrated to activate oncogenic pathways involving Akt and
to promote the proliferation of prostate cancer, in addition to
stimulating tumor-associated angiogenesis and inflammation
(3–6).
Furthermore, in a large population study involving men with a serum
prostate-specific antigen of >10 ng/ml, high pre-treatment serum
levels of Cav were a prognostic marker for prostate cancer
recurrence (7). Increased circulating
levels of Cav were also shown to be associated with the
histological grade and progression of prostate cancer (8,9). These
observations suggest the involvement of Cav in tumor growth and
aggressiveness of prostate cancer.
Pigment epithelium-derived factor (PEDF) is a
glycoprotein that belongs to the superfamily of serine protease
inhibitors, which exhibit potent neuronal differentiating activity
(10). PEDF has been identified as a
highly effective inhibitor of angiogenesis in cell culture models
as well as in animal models (11,12).
Furthermore, studies have demonstrated that PEDF blocks
cytokine-induced and vascular endothelial growth factor-induced
angiogenesis and inflammatory reactions, inhibits tumor growth and
induces apoptosis in tumors, including prostate cancer (11–24). Our
group has recently demonstrated that PEDF binds to Cav and blocks
its pro-inflammatory effects in endothelial cells (25). It is therefore possible that PEDF may
exert its antitumor effects in prostate cancer by modulating the
actions of Cav. However, it remains unclear whether PEDF inhibits
the tumor-promoting effects of Cav in cultured prostate cancer
cells. The present study examined the effects of PEDF on prostate
cancer cell growth and on the expression of the interleukin-8
(IL-8) gene, which contributes to prostate cancer progression in
PC-3 cells, a human hormone-refractory (HRPC) cell line (17).
Materials and methods
Materials
Polyclonal anti-human rabbit antibodies (Abs)
directed against Cav (cat no. 3238) and polyclonal
biotin-conjugated Abs against PEDF (cat no. BAF 1177) were
purchased from Cell Signaling Technology Japan K.K. (Tokyo, Japan)
and R&D Systems (Minneapolis, MN, USA), respectively.
Monoclonal mouse anti-human Abs raised against GAPDH were obtained
from Santa Cruz Biotechnology, Inc., Dallas, TX, USA (cat no.
sc-32233).
Expression and purification of
Cav
Full-length human Cav (residues 1-178) was amplified
using polymerase chain reaction (PCR), sub-cloned into the NdeI and
XhoI sites of the pET21b vector, incorporating pentahistidine-tag
into the C-terminus of the protein and purified as previously
described (26). Briefly, the PCR
primers for Cav were as follows: Forward,
5′-CCGGTCCATATGTCTGGGGGCAAATAC-3′ and reverse,
5′-CCCCTCGAGTATTTCTTTCTGCAAGTT-3′. PCR reactions were performed
using PrimeSTAR® HS (Takara Bio Inc., Shiga, Japan) and the PCR
conditions were as follows: 35 cycles of 98°C for 10 sec, 55°C for
10 sec and 72°C for 120 sec.
Preparation of PEDF proteins
Hexahistidine-tagged PEDF proteins were purified
from conditioned media using a Ni-NTA spin kit (Qiagen GmbH,
Hilden, Germany), according to the manufacturer's instructions as
described previously (27). SDS-PAGE
analysis of the purified PEDF proteins identified a single band
with a molecular mass of ~50 kDa, which exhibited positive
reactivity with monoclonal mouse Abs against human PEDF (cat. no.
KM037; 1:1,000 dilution; Transgenic, Kumamoto, Japan).
Construction and transfection of small
interfering RNAs (siRNAs)
The following siRNAs to Cav (siCavs) were used:
Sense, 5′-GCUUCCUGAUUGAGAUUCAtt-3′ and antisense,
5′-UGAAUCUCAAUCAGGAAGCtc for siCav#1; and sense,
5′-GCCGUGUCUAUUCCAUCUAtt-3′ and antisense, 5′-UAGAUGGAAUAGACACGGCtg
for siCav#2. The siCavs were obtained from Life Technologies Japan,
Ltd. (Tokyo, Japan). Non-silencing control siRNAs (siCon) were also
obtained from Life Technologies Japan, Ltd. (Silencer Negative
Control #1 siRNA). The siRNA duplexes were transfected into PC-3
cells using Lipofectamine 2000™ (Invitrogen Life Technologies,
Carlsbad, CA, USA), as previously described (28). Following 2 days of transfection, Cav
and GAPDH protein levels were analyzed using western blotting.
Cells
PC-3 human prostate cancer cells (American Type
Culture Collection, Manassas, VA, USA) were cultured in RPMI-1640
(Sigma-Aldrich, St. Louis, MO, USA) with 10% fetal bovine serum and
100 U/ml penicillin/streptomycin (Life Technologies Japan, Ltd.).
PEDF or Cav treatment was conducted in a medium containing 1% fetal
bovine serum.
Reverse transcription-quantitative PCR
(RT-qPCR)
Transfected or non-transfected PC-3 cells were
treated with or without the indicated concentrations of Cav (0.1, 1
or 10 nM), in the presence or absence of 10 nM PEDF for 4 h. Total
RNA was then extracted using a NucleoSpin RNA kit (Takara Bio
Inc.), according to the manufacturer's instructions. qRT-PCR was
performed using Assay-on-Demand and TaqMan 5 fluorogenic nuclease
chemistry (Life Technologies Japan Ltd.), according to the
manufacturer's instructions. The IDs of the primers for human
interleukin-8 (IL-8) and the β-actin gene (Applied Biosystems Life
Technologies, Foster City, CA, USA) were Hs99999034_m1 and
Hs01060665_g1, respectively. RT-qPCR conditions were as follows:
50°C for 2 min and 95°C 10 min, followed by 45 cycles of 95°C for
15 sec and 60°C for 2 min. The results were quantified using the
2−ΔΔCT method (29).
Serum preparation
Serum was obtained from 3 individuals: A healthy 38
year-old male [body mass index (BMI), 21.1]; a 59 year-old diabetic
man with cardiovascular disease (BMI, 31.1; HbA1c, 8.6%), as PEDF
levels have been shown to increase in diabetic patients (24); and a 64 year-old patient with stage II
prostate cancer (BMI unknown). All prostate cancer patient serum
speciemens were purchased from Tissue Solutions Ltd., Glasgow,
UK.
Western blotting analysis
Proteins were extracted from siCon- or
siCav-transfected PC-3 cells, using lysis buffer as previously
described (28). The samples or serum
were then separated by SDS-PAGE and transferred to nitrocellulose
membranes (Life Technologies Japan, Ltd.). Membranes were probed
with Abs against Cav (1:1,000 dilution), PEDF (1:1,000 dilution) or
GAPDH (1:200 dilution) for 12 h at room temperature, and then
incubated with peroxidase-conjugated polyclonal donkey anti-rabbit
IgG Abs (1:20,000 dilution; cat no. NA934-100UL; GE Healthcare UK
Ltd., Little Chalfont, UK), streptavidin-conjugated peroxidase
(1:5,000) or polyclonal sheep anti-mouse Abs (1:20,000 dilution;
cat no. NA931-100UL; GE Healthcare UK Ltd.), respectively. Immune
complexes were visualized using an enhanced chemiluminescence
detection system (Amersham Bioscience, Buckinghamshire, UK). The
protein signals were quantified using ImageJ software (version
1.46; National Institutes of Health, Bethesda, MD, USA). The Cav
protein levels of each sample were normalized to those of
GAPDH.
Measurement of [3H]
thymidine incorporation into PC-3 cells
PC-3 cells were treated with or without 10 nM Cav,
in the presence or absence of 10 nM PEDF for 20 h. Subsequently,
[3H] thymidine was added, to produce a final
concentration of 1 mCi/ml, and cells were incubated for a further 4
h. Following incubation, PC-3 cells were fixed with ice-cold 10%
(w/v) trichroloacetic acid for 20 min. [3H] thymidine
incorporation into PC-3 cells was measured as described previously
(30). Briefly, [3H]
thymidine incorporation into PC-3 cells was measured by processing
the resultant acid-insoluble materials for liquid scintillation
counting (AccuFLEX LSC 7200; Hitachi Aloka Medical, Ltd., Tokyo,
Japan).
Statistical analysis
Data are presented as the mean ± standard error.
Student's t-test was performed for statistical comparisons. All
statistical analyses were performed using PASW Statistics version
18.0 software (SPSS Japan Inc., Tokyo, Japan) and P<0.05 was
considered to indicate a statistically significant difference.
Results
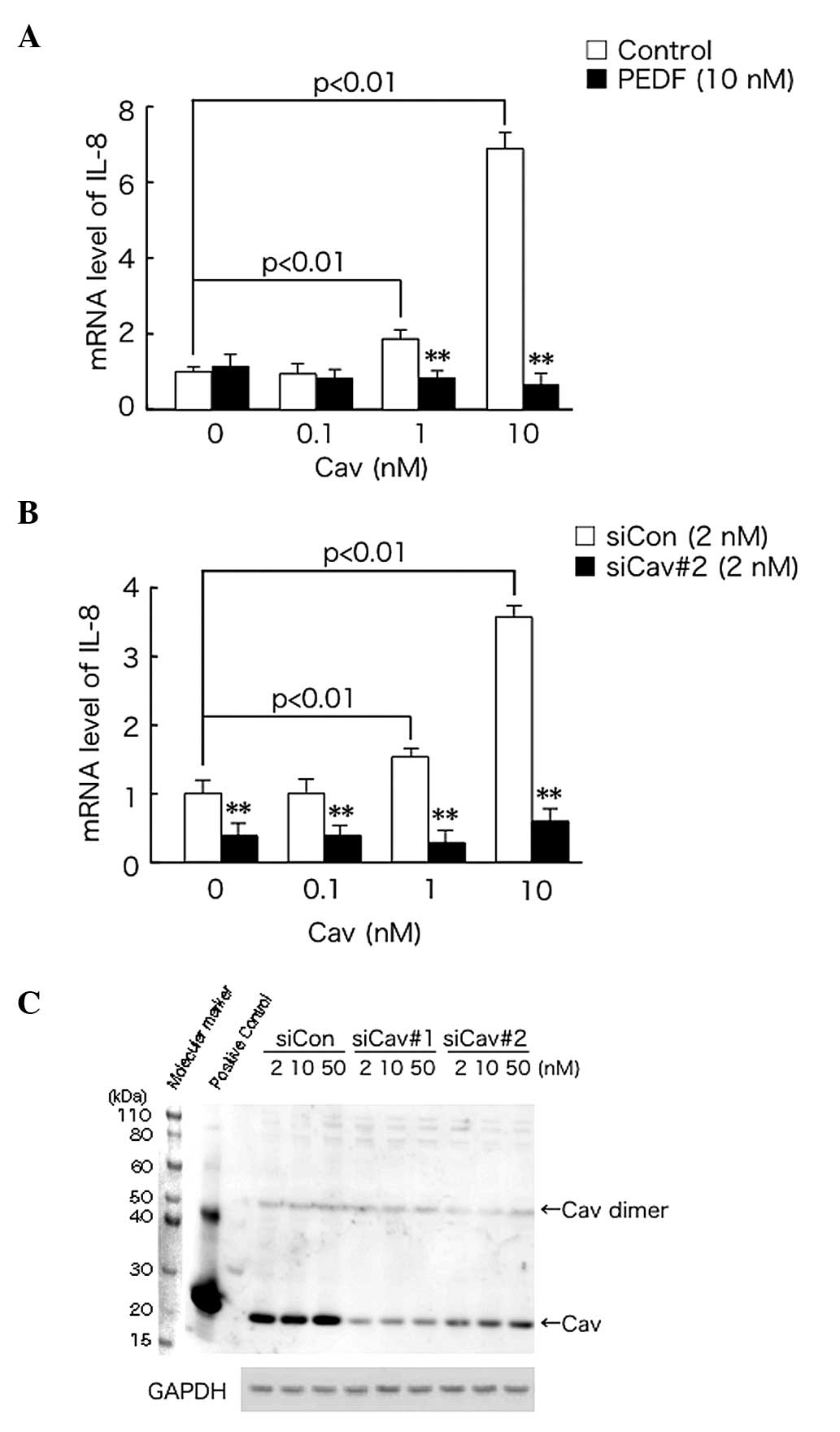
Cav leads to upregulation of IL-8
expression in PC-3 cells and this effect is abrogated by PEDF
The effect of PEDF on IL-8 gene expression in
Cav-exposed PC-3 cells was investigated. As shown in Fig. 1A, exogenously administered Cav
upregulated the mRNA expression of IL-8 in PC-3 cells, in a
dose-dependent manner. This effect was abrogated by treatment with
1 or 10 nM PEDF.
Cav increased IL-8 mRNA expression in
siCon-transfected cells, in a dose-dependent manner and this effect
was suppressed by siCav#2 transfection (Fig. 1B). Furthermore, transfection with
siCav#1 or siCav#2, reduced Cav expression in PC-3 cells to
1/5–1/10 of that of the siCon-transfected control cells (Fig. 1C).
PEDF leads to decreased DNA synthesis
in PC-3 cells
The effect of PEDF on DNA synthesis in PC-3 cells
was also examined. As shown in Fig.
2A, 10 nM Cav significantly increased DNA synthesis in PC-3
cells. This effect was inhibited by treatment with 10 nM PEDF. PEDF
(10 nM) also significantly decreased DNA synthesis in PC-3 cells
when administered alone.
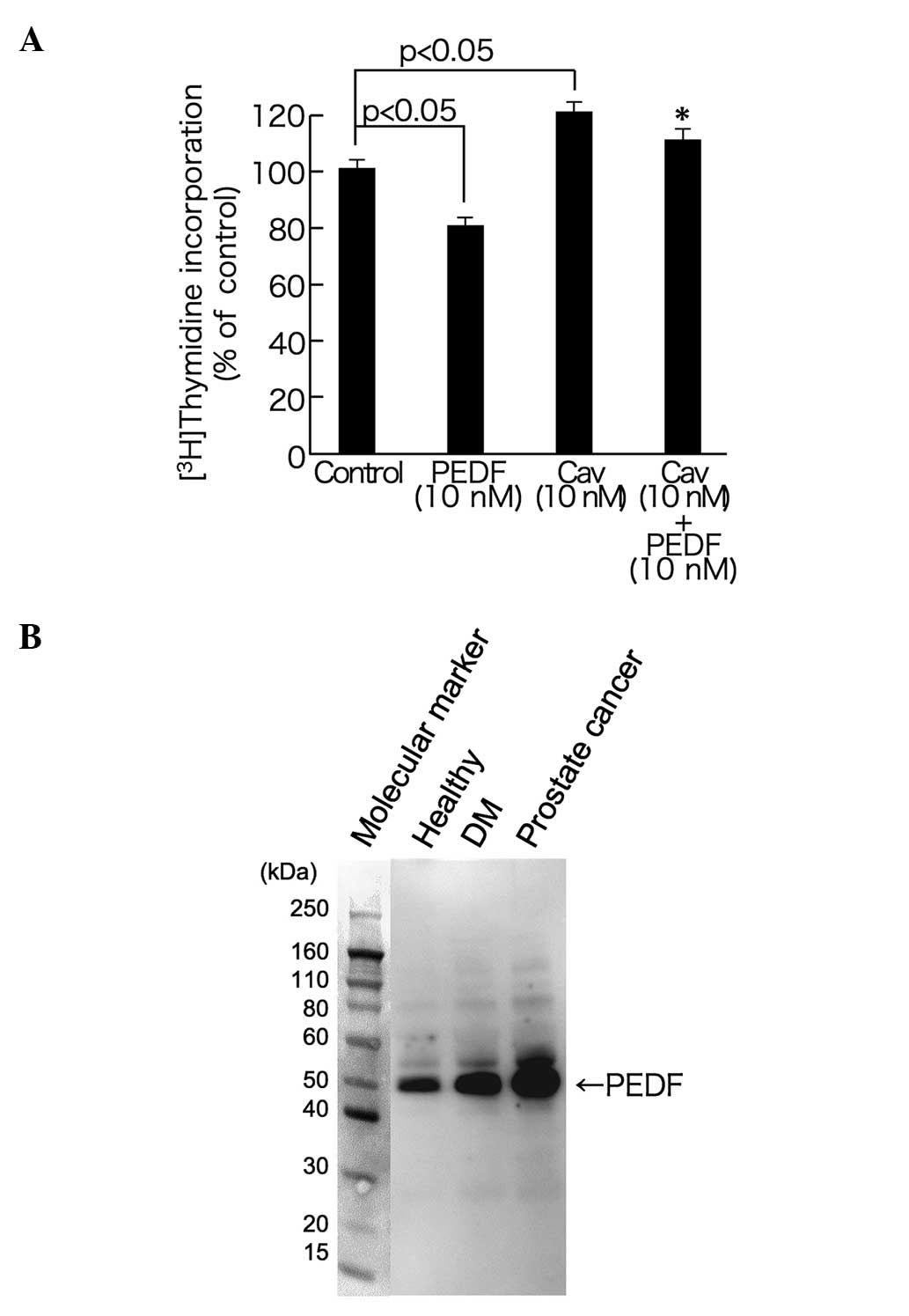 | Figure 2.Effect of PEDF on DNA synthesis in
PC-3 cells (A) and circulating PEDF levels in three subjects (B).
(A) PC-3 cells were treated with or without 10 nM Cav, in the
presence or absence of 10 nM PEDF for 20 h. Subsequently,
[3H]thymidine was added, to produce a final
concentration of 1 µCi/ml, and cells were further incubated for 4
h. Following incubation, PC-3 cells were fixed, and
[3H]thymidine incorporation into PC-3 cells was
measured. *P<0.05, compared with the 10 nM Cav alone group. n=4.
(B) Serum was separated using SDS-PAGE and transferred to
nitrocellulose membranes. Membranes were then probed with
antibodies against PEDF. PEDF, pigment epithelium-derived factor;
Cav, caveolin-1; DM, diabetes mellitus. |
Finally, the circulating levels of PEDF in the serum
of the patient with prostate cancer were higher than those of the
healthy control or the diabetic subjects (Fig. 2B).
Discussion
There is accumulating evidence that overexpression
of Cav promotes tumor growth and metastasis of prostate cancer via
diverse pathways (3–6), while PEDF may protect against the
progression of prostate cancer, via anti-angiogenic,
anti-inflammatory and proapoptotic effects (17–19). To
the best of our knowledge, the present study demonstrated for the
first time that treatment with 1 or 10 nM PEDF significantly
inhibited the Cav-induced increase in IL-8 mRNA expression in PC-3
cells. Transfection with siCav#2 reduced endogenous Cav expression
in PC-3 cells to ~1/5 of that of siCon-transfected cells, and
decreased IL-8 mRNA expression in Cav-exposed PC-3 cells. Our group
has recently demonstrated that PEDF directly binds to Cav at a
KD value of 7.36×10−7 M; that
exogenously administered Cav is taken up into endothelial cells,
resulting in increases in the membrane levels of Cav and promoting
inflammatory reactions; and that siCav#1 treatment reduces Cav
expression and suppresses inflammatory gene expression in
endothelial cells (25). These
observations indicate that PEDF may reduce Cav-induced IL-8 gene
expression by binding to exogenously administered Cav and
modulating its membrane levels in PC-3 cells, a type of
hormone-refractory prostate cancer (HRPC) cell. IL-8 may promote
the transition of prostate cancer to HRPC by upregulating androgen
receptor expression and activation, and may stimulate the
proliferation and migration of this cell type (17). Circulating Cav levels have been shown
to be increased in patients with HRPC, while they were not
increased in those with hormone-sensitive cancer (9). These findings further support the
hypothesis that PEDF may inhibit the growth and metastasis of HRPC
cells, such as PC-3 cells, by blocking Cav-evoked IL-8
overexpression.
The present study also demonstrated that PEDF
inhibited the Cav-induced increase in DNA synthesis in PC-3 cells.
Treatment with PEDF alone also decreased [3H] thymidine
incorporation into PC-3 cells. Therefore, PEDF may directly inhibit
the proliferation of PC-3 cells via modulation of the
growth-promoting activity of Cav.
To the best of our knowledge, the current study also
demonstrated for the first time that circulating PEDF levels were
higher in a patient with prostate cancer, compared with levels in
the healthy control or diabetic subjects. As reported previously
(31), PEDF levels in diabetic
subjects were higher than those in healthy controls. The antitumor
effects of PEDF that were observed in the present study, suggest
that circulating PEDF levels may be elevated as a counter-system
against Cav overexpression in prostate cancer. Treatment with Cav
antisera reduced the development and growth of primary tumors and
metastases in mouse models of prostate cancer (32). Since it has been demonstrated that the
laminin receptor mediates the anti-inflammatory and
antithrombogenic effects of PEDF in malignant myeloma cells
(33), blockade of the effects of Cav
using a PEDF-laminin receptor system may be a novel therapeutic
approach for prostate cancer.
It has previously been reported that the human blood
concentration of PEDF is 100–200 nM (31,34).
However, the majority of circulating PEDF in serum may exist as a
protein-bound and inactive from, and the free active form of PEDF
comprises only a small portion of the total level do of this
protein (31). In addition, a 4
nM-increase in serum PEDF levels has been shown to exert
anti-inflammatory effects in animal models (35,36).
Therefore, it is unlikely that PEDF at 10 nM would produce toxic
effects on PC-3 cells. Recently, serum levels of Cav were
demonstrated to be increased in patients with prostate cancer
compared with healthy controls. Serum levels of Cav were highest in
those in the advanced stages of prostate cancer (8). Furthermore, epithelial cell hyperplasia
in the prostate, in association with increased angiogenesis, was
observed in PEDF-deficient mice (16). The present results support the
involvement of an interaction between PEDF and Cav in prostate
cancer progression in vivo.
Acknowledgements
This study was supported by Grants-in-Aid for
Scientific Research (B) from the Ministry of Education, Culture,
Sports, Science and Technology, Japan (grant no. 25293127).
References
|
1
|
Sowa G: Caveolae, caveolins, cavins and
endothelial cell function: new insights. Front Physiol.
2:1202102.
|
|
2
|
Guan TH, Chen G, Gao B, Janssen MR,
Uttarwar L, Ingram AJ and Krepinsky JC: Caveolin-1 deficiency
protects against mesangial matrix expansion in a nouse model of
type 1 diabetic nephropathy. Diabetologia. 56:2068–2077. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thompson TC, Tahir SA, Li L, Watanabe M,
Naruishi K, Yang G, Kadmon D, Logothetis CJ, Troncoso P, Ren C, et
al: The role of caveolin-1 in prostate cancer: clinical
implications. Prostate Cancer Prostatic Dis. 13:6–11. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Freeman MR, Yang W and Di Vizio D:
Caveolin-1 and prostate cancer progression. Adv Exp Med Biol.
729:95–110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li L, Ren C, Yang G, Goltsov AA, Tabata K
and Thompson TC: Caveolin-1 promotes autoregulatory, Akt-mediated
induction of cancer-promoting growth factors in prostate cancer
cells. Mol Cancer Res. 7:1781–1791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tahir SA, Yang G, Goltsov AA, Watanabe M,
Tabata K, Addai J, Fattah el MA, Kadmon D and Thompson TC: Tumor
cell-secreted caveolin-1 has proangiogenic activities in prostate
cancer. Cancer Res. 68:731–739. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tahir SA, Frolov A, Hayes TG, Mims MP,
Miles BJ, Lerner SP, Wheeler TM, Ayala G, Thompson TC and Kadmon D:
Preoperative serum caveolin-1 as a prognostic marker for recurrence
in a radical prostatectomy cohort. Clin Cancer Res. 12:4872–4875.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gumulec J, Sochor J, Hlavna M,
Sztalmachova M, Krizkova S, Babula P, Hrabec R, Rovny A, Adam V,
Eckschlager T, et al: Caveolin-1 as a potential high-risk prostate
cancer biomarker. Oncol Rep. 27:831–841. 2012.PubMed/NCBI
|
|
9
|
Sugie S, Mukai S, Tsukino H, Toda Y,
Yamauchi T, Nishikata I, Kuroda Y, Morishita K and Kamoto T:
Increased plasma caveolin-1 levels are associated with progression
of prostate cancer among Japanese men. Anticancer Res.
33:1893–1897. 2013.PubMed/NCBI
|
|
10
|
TombranTink J, Chader CG and Johnson LV:
PEDF: A pigment epithelium-derived factor with potent neuronal
differentiative activity. Exp Eye Res. 53:411–414. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dawson DW, Volpert OV, Gillis P, Crawford
SE, Xu H, Benedict W and Bouck NP: Pigment epithelium-derived
factor: a potent inhibitor of angiogenesis. Science. 285:245–248.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Duh EJ, Yang HS, Suzuma I, Miyagi M,
Youngman E, Mori K, Katai M, Yan L, Suzuma K, West K, et al:
Pigment epithelium-derived factor suppresses ischemia-induced
retinal neovascularization and VEGF-induced migration and growth.
Invest Ophthalmol Vis Sci. 43:821–829. 2002.PubMed/NCBI
|
|
13
|
Yamagishi S, Amano S, Inagaki Y, Okamoto
T, Takeuchi M and Inoue H: Pigment epithelium-derived factor
inhibits leptin-induced angiogenesis by suppressing vascular
endothelial growth factor gene expression through anti-oxidative
properties. Microvasc Res. 65:186–190. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamagishi S, Nakamura K, Matsui T, Inagaki
Y, Takenaka K, Jinnouchi Y, Yoshida Y, Matsuura T, Narama I,
Motomiya Y, et al: Pigment epithelium-derived factor inhibits
advanced glycation end product-induced retinal vascular
hyperpermeability by blocking reactive oxygen species-mediated
vascular endothelial growth factor expression. J Biol Chem.
281:20213–20220. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsui T, Nishino Y, Maeda S and Yamagishi
S: PEDF-derived peptide inhibits corneal angiogenesis by
suppressing VEGF expression. Microvasc Res. 84:105–108. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Doll JA, Stellmach VM, Bouck NP, Bergh AR,
Lee C, Abramson LP, Cornwell ML, Pins MR, Borensztajn J and
Crawford SE: Pigment epithelium-derived factor regulates the
vasculature and mass of the prostate and pancreas. Nat Med.
9:774–780. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hirsch J, Johnson CL, Nelius T, Kennedy R,
Riese Wd and Filleur S: PEDF inhibits IL8 production in prostate
cancer cells through PEDF receptor/phospholipase A2 and regulation
of NFκB and PPARγ. Cytokine. 55:202–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mirochnik Y, Aurora A, SchulzeHoepfner FT,
Deabes A, Shifrin V, Beckmann R, Polsky C and Volpert OV: Short
pigment epithelial-derived factor-derived peptide inhibits
angiogenesis and tumor growth. Clin Cancer Res. 15:1655–1663. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gong Q, Qiu S, Li S, Ma Y, Chen M, Yao Y,
Che D, Feng J, Cai W, Ma J, et al: Proapoptotic PEDF functional
peptides inhibit prostate tumor growth-A mechanistic study. Biochem
Pharmacol. 92:425–437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Abe R, Shimizu T, Yamagishi S, Shibaki A,
Amano S, Inagaki Y, Watanabe H, Sugawara H, Nakamura H, Takeuchi M,
et al: Overexpression of pigment epithelium-derived factor
decreases angiogenesis and inhibits the growth of human malignant
melanoma cells in vivo. Am J Pathol. 164:1225–1232. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takenaka K, Yamagishi S, Jinnouchi Y,
Nakamura K, Matsui T and Imaizumi T: Pigment epithelium-derived
factor (PEDF)-induced apoptosis and inhibition of vascular
endothelial growth factor (VEGF) expression in MG63 human
osteosarcoma cells. Life Sci. 77:3231–3241. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Seki R, Yamagishi S, Matsui T, Yoshida T,
Torimura T, Ueno T, Sata M and Okamura T: Pigment
epithelium-derived factor (PEDF) inhibits survival and
proliferation of VEGF-exposed multiple myeloma cells through its
anti-oxidative properties. Biochem Biophys Res Commun. 431:693–697.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hoshina D, Abe R, Yamagishi SI and Shimizu
H: The role of PEDF in tumor growth and metastasis. Curr Mol Med.
10:292–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamagishi S and Matsui T: Pigment
epithelium-derived factor (PEDF) and cardiometabolic disorders.
Curr Pharm Des. 20:2377–2386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matsui T, Higashimoto Y, Taira J and
Yamagishi S: Pigment epithelium-derived factor (PEDF) binds to
caveolin-1 and inhibits the pro-inflammatory effects of caveolin-1
in endothelial cells. Biochem Biophys Res Commun. 441:405–410.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Taira J and Higashimoto Y: Caveolin-1
interacts with protein phosphatase 5 and modulates its activity in
prostate cancer cells. Biochem Biophys Res Commun. 431:724–728.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamagishi S, Inagaki Y, Amano S, Okamoto
T, Takeuchi M and Makita Z: Pigment epithelium-derived factor
protects cultured retinal pericytes from advanced glycation end
product-induced injury through its antioxidative properties.
Biochem Biophys Res Commun. 296:877–882. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ojima A, Ishibashi Y, Matsui T, Maeda S,
Nishino Y, Takeuchi M, Fukami K and Yamagishi S: Glucagon-like
peptide-1 receptor agonist inhibits asymmetric dimethylarginine
generation in the kidney of streptozotocin-induced diabetic rats by
blocking advanced glycation end product-induced protein arginine
methyltranferase-1 expression. Am J Pathol. 182:132–141. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Higashimoto Y, Matsui T, Nishino Y, Taira
J, Inoue H, Takeuchi M and Yamagishi S: Blockade by
phosphorothioate aptamers of advanced glycation end
products-induced damage in cultured pericytes and endothelial
cells. Microvasc Res. 90:64–70. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamagishi S, Adachi H, Abe A, Yashiro T,
Enomoto M, Furuki K, Hino A, Jinnouchi Y, Takenaka K, Matsui T, et
al: Elevated serum levels of pigment epithelium-derived factor in
the metabolic syndrome. J Clin Endocrinol Metab. 91:2447–2450.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tahir SA, Yang G, Ebara S, Timme TL, Satoh
T, Li L, Goltsov A, Ittmann M, Morrisett JD and Thompson TC:
Secreted caveolin-1 stimulates cell survival/clonal growth and
contributes to metastasis in androgen-insensitive prostate cancer.
Cancer Res. 61:3882–3885. 2001.PubMed/NCBI
|
|
33
|
Matsui T, Higashimoto Y and Yamagishi S:
Laminin receptor mediates anti-inflammatory and anti-thrombogenic
effects of pigment epithelium-derived factor in myeloma cells.
Biochem Biophys Res Commun. 443:847–851. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Petersen SV, Valnickova Z and Enghild JJ:
Pigment epithelium-derived factor (PEDF) occurs at a
physiologically concentration in human blood: purification and
characterization. Biochemical J. 374:199–206. 2003. View Article : Google Scholar
|
|
35
|
Fujimura T, Yamagishi S, Ueda S, Fukami K,
Shibata R, Matsumoto Y, Kaida Y, Hayashida A, Koike K, Matsui T, et
al: Administration of pigment epithelium-derived factor (PEDF)
reduces proteinuria by suppressing decreased nephrin and increased
VEGF expression in the glomeruli of adriamycin-injected rats.
Nephrol Dial Transplant. 24:1397–1406. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matsui T, Nishino Y, Ojima A, Maeda S,
Tahara N and Yamagishi S: Pigment epithelium-derived factor
improves metabolic derangements and ameliorates dysregulation of
adipocytokines in obese type 2 diabetic rats. Am J Pathol.
184:1094–1103. 2014. View Article : Google Scholar : PubMed/NCBI
|