Introduction
Colorectal cancer is a leading cause of malignant
tumor-related mortality in developed countries (1), and is the third most common cancer type
in China (2). Every year, ~300,000
mortalities from colorectal cancer occur globally (3,4).
Chemotherapy with 5-fluorouracil (5-FU) and associated adjuvant
agents remains the standard drug treatment regimen against
early-stage and advanced colorectal cancers. Despite progress in
the elucidation of the carcinogenic processes of colon cancer, more
effective treatment regimens based on novel and innovative
approaches are required, since the remission rate induced by
treatment with 5-FU and associated adjuvant agents is low (5,6). In
addition, standard chemotherapy produces numerous systemic adverse
reactions, including a decrease in leukocyte and platelet counts
(7,8).
Hence, more effective and safer therapeutic strategies are urgently
required.
Certain non-steroidal anti-inflammatory drugs
(NSAIDs) exhibit inhibitory effects on COX-2, and are used
extensively for analgesic and antipyretic treatments (9). Previous studies have demonstrated that
NSAIDs reduce the risk and mortality of several cancer types
(10,11). While their anti-tumorigenic mechanisms
are not completely understood, cyclooxygenase (COX)-dependent and
COX-independent pathways may play significant roles (12,13). In
2004, a study reported increased cardiovascular toxicity occurring
in patients who received the drug Vioxx for longer than 18 months
(14). Thus, drugs from
herbal/natural sources with COX-2 inhibitory properties that
produce fewer side effects are of interest. Sinomenium
acutum has a long history of medicinal use in traditional
Chinese medicine, and is now commonly used as a COX-2 inhibitor and
as an anti-inflammatory agent in mixed herbal decoctions for the
treatment of neuralgia and rheumatic diseases (15,16). It is
capable of potently releasing histamine in association with the
degranulation of mast cells in mammalian tissues. The release of
histamine is responsible for the dominant pharmacological actions
of sinomenine (SIN) (17), including
vasodilatation, increased vascular permeability, acceleration of
thoracic and peripheral lymph flow, contraction of the plain
muscles, increased peristalsis of the intestines, and stimulation
of gastric acid secretion (18). The
active pharmacological constituents of Sinomenium acutum
include alkaloids, sterols, phospholipids and several other
components. Extensive pharmacological and clinical research on SIN
has primarily focused on the immune, cardiovascular and nervous
system (17).
SIN possesses antitumor activity in certain cancer
types and is already prescribed to patients with cardiac diseases
(19). SIN exhibits a significant
apoptotic effect on NCI-H460 cells through the
mitochondria-mediated apoptosis pathway. SIN-induced apoptosis is
accompanied by the collapse of the mitochondrial membrane
potential, the release of cytochrome c and the activation of
caspase-9 and caspase-3. SIN also increases the levels of Bax
protein and decreases the levels of Bcl-2 protein in NCI-H460 cells
(20). It also induces apoptosis in
NCI-H226 and NCI-H522 cells through the activation of pAkt and Perk
(21). However, the anti-tumorigenic
action of SIN in colon carcinogenesis has not been clearly
determined.
The present study examined the anti-tumorigenic
effect of SIN from Sinomenium acutum by focusing on the
anti-tumorigenic effects and molecular mechanisms of SIN in SW1116
human colon cancer cells. The growth-inhibitory effects of SIN were
examined in vitro and in vivo using a nude mouse
xenograft model. We hypothesized that the anti-carcinogenic action
of SIN might be due to the inhibition of COX-2 expression in the
cancer cells and/or effects on cell cycle regulation.
Materials and methods
Materials
Sinomenine hydrochloride was obtained from Hunan
Zhengqing Pharmaceutical Co. Ltd. (Huaihua, Hunan, China). Primary
antibodies against COX-2, cyclin D1, cyclin E, Cip1/p21 and
Kip1/p27 were purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA). The antibody against GAPDH was purchased from
Sigma-Aldrich (St. Louis, MO, USA). The bicinchoninic acid (BCA)
protein assay kit was purchased from the Beyotime Institute of
Biotechnology (Haimen, China). An enhanced chemiluminescence (ECL)
western blotting kit was purchased from Millipore (Billerica, MA,
USA). A PrimeScript™ RT reagent kit was obtained from Takara
Biotechnology Co., Ltd. (Dalian, China). Universal SYBR-Green I was
purchased from Bioteke Corporation (Beijing, China). TRIzol reagent
was purchased from Invitrogen Life Technologies (Carlsbad, CA,
USA). The RNeasy kit was purchased from Qiagen (Hilden, Germany).
Diethylpyrocarbonate was purchased from Sigma (Poole, Dorset, UK).
All other reagents were widely available commercially. All
quantitative polymerase chain reaction (qPCR) experiments were
performed on an Applied Biosystems 7900HT Fast Real-Time PCR system
(Life Technologies, Grand Island, NY, USA).
Cell culture and synchronization
The human colon adenocarcinoma cell line SW1116 was
purchased from the Type Culture Collection of the Chinese Academy
of Sciences, Shanghai, China. Cells were maintained in L-15 medium
supplemented with 10% fetal bovine serum (FBS) in a humidified
atmosphere of 100% air at 37°C. A subculture of cells was processed
by enzymatic digestion (trypsin/ethylenediaminetetraacetic acid
solution: 0.25/0.02%). Sinomenine hydrochloride was dissolved in
phosphate-buffered saline (PBS) as a 100 mM stock solution and then
diluted with the L-15 medium. All experiments were performed using
media containing 1% serum following 24 h of serum starvation. This
procedure has been effective for the synchronization of cells in
the G0 phase in cell cycle studies (22,23).
Cell viability assay
Cell viability was detected using CCK-8. When 70–80%
confluence was reached, SW1116 cells (2×104) were
cultured in 96-well plates and exposed to various concentrations of
SIN (1, 2, 4, 8 and 16 mM) for 24, 48 or 72 h. L-15 medium (0 mM)
was added to the control wells at the various treatment times. The
CCK-8 solution (10 µl) diluted 1:10 in FBS-free L-15 (100 µl) was
added to each well and incubated for 3 h at 37°C. The absorbance
was measured at 450 nm using a Bio-Rad 680 microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The mean optical
density (OD) of five wells in the indicated groups was used to
calculate the percentage of cell viability as follows: Cell
viability (%) =(ODtreatment group - ODblank
group) / (ODcontrol group - ODblank
group) × 100%. The experiment was performed in
triplicate.
Ultrastructure observation
SW1116 cells (4×106) were planted in
100-mm2 culture dishes and treated with 8 mM SIN for 48
h. The scratched cells were harvested and washed twice with
ice-cold PBS, centrifuged (200 × g, 4°C, 10 min) and fixed with
ice-cold 2.5% glutaraldehyde for 24 h at 4°C, washed twice in PBS,
postfixed in 1% osmium tetroxide in 0.1 M phosphate buffer for 2–3
h at 4°C, then double-washed in distilled water. The samples were
blocked in warm agar (50°C), dehydrated in ethanol series (50, 75,
95 and double 100%) and embedded in 618 epoxy resin. Finally, glass
knives were used to obtain ultrathin sections of 90 nm that were
stained with uranyl acetate and lead citrate for observation under
transmission electron microscopy (CM120, Philips, Amsterdam, The
Netherlands).
Flow cytometry analysis for cell cycle
distribution
To synchronize the cell cycle at the G0 boundary,
SW1116 cells (4×106) were planted in 100-mm2
culture dishes and treated with various concentrations of SIN (0,
4, 8, 10 and 16 mM) for 24, 48 or 72 h. Cells were harvested by
brief trypsinization and centrifugation. The cells were washed
twice with ice-cold PBS, centrifuged (200 × g, 4°C, 10 min) and
fixed with ice-cold 70% ethanol for 4 h. After staining with 50
µg/ml propidium iodide and 500 µg/ml RNase at room temperature for
30 min, the cells were subjected to fluorescence-activated cell
sorting with FACScan (Becton Dickinson, Franklin Lakes, NJ, USA) to
analyze the cell cycle.
Analysis of COX-2 protein expression
by western blot analysis
SW1116 cells (4×106) plated in
100-mm2 culture dishes were treated with various
concentrations of SIN (0, 4, 8 and 10 mM) for 24, 48 or 72 h.
Proteins were obtained by cell lysis in ice-cold cell
radioimmunoprecipitation assay buffer. Total proteins in the
supernatant were measured using the BCA protein assay kit
(Beyotime). Thirty micrograms of total proteins from each sample
were separated by 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The proteins in the gel were transferred to
nitrocellulose membranes and probed with the COX-2 (D5H5) XP®
rabbit monoclonal antibody. The immunoblots were developed and
visualized using an ECL detection system (Bioshine, Shanghai,
China). Each blot was stripped and reprobed with GAPDH antibody as
an internal control. The signal was visualized using an ECL
detection system. Western blot analysis of COX-2 protein levels in
tumor tissues from nude mice was performed as described for the
western blot analysis of SW1116 cells.
Analysis of COX-2 mRNA expression by
qPCR
SW1116 cells (4×106) plated in
100-mm2 culture dishes were treated with various
concentrations of SIN (0, 4, 8, 10 mM) for 24, 48 or 72 h. RNA
isolation was followed by qPCR to detect COX-2 mRNA. Total RNA was
extracted from all cell lines using the RNeasy kit (Qiagen).
Complementary DNA was synthesized from total RNA using the
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd.). qPCR
measurement of individual cDNA was performed using SYBR-Green dye
to measure the formation of duplex DNA, and the result was
normalized to the expression of GAPDH. Amplification was performed
by denaturation at 95°C for 30 sec, followed by 40 cycles of 5 sec
at 95°C (melting) and 30–34 sec at 60°C (annealing and elongation)
on an Applied Biosystems 7900HT sequence detection system (Applied
Biosystems, Foster City, CA, USA). The primers used were as
follows: COX-2 upstream primer, 5′-GGAAACTGTGGCGTGATGGCCG-3′; COX-2
downstream primer, 5′-GTTGGCAGTGGGGACACGGAAG-3′ (COX-2 product
size, 199 bp); GAPDH upstream primer, 5′-GCTGGTGGTAGGAATGTTCC-3′;
GAPDH downstream primer, 5′-CAGCATCGATGTCACCATAG-3′ (GAPDH product
size, 140 bp). The qPCR analysis of COX-2 mRNA in tumor tissues
from nude mice was performed as described for SW1116 cells.
Tumor xenografts in nude mice
The experimental procedures were approved by the
Animal Care and Use Committee of Shanghai University of Traditional
Chinese Medicine and conformed to the international standards on
the ethical treatment of animals. Balb/c nu/nu mice (male, body
mass 16–20 g) were purchased from Shanghai Super-B&K Laboratory
Animal Co. Ltd. (Shanghai, China). The mice were maintained under
sterile and pathogen-free conditions in isolated pathogen-free
ventilation chambers at an ambient temperature of 20–22°C with
45–50% relative humidity. The animal rearing facility was
maintained on a 12-h light-dark cycle. All animals were given free
access to sterilized food and water and were habituated for 7 days
before experimentation.
Mice were randomly assigned to the control and
treatment groups. The treatment regimens were as follows (n=8):
control group, intraperitoneal (IP) injection with normal saline
(0.2 ml) once daily (days 0–30); SIN treatment group, IP injection
with various doses of SIN (25, 50 and 100 mg/kg) once daily (days
0–30); 5-FU group, IP injection with 5-FU (20 mg/kg) once daily
(days 0–14). The cell suspension was injected subcutaneously into
the right thigh of each animal (at a cell density of
5×106 in 200 µl PBS). The day of tumor implantation was
designated day 0. Tumors became palpable 10 days after the
xenograft procedure. Tumor volume was measured using a digital
caliper every 3 days and calculated as (length × width2)
/ 2 (24). Mice were monitored for 40
days following tumor inoculation. The body weight of all animals
was recorded throughout the entire experimental to assess drug
toxicity. Any mortality during the course of the study was also
recorded.
Immunohistochemical analysis of the
expression of cyclin D1, cyclin E, Cip1/p21 and Kip1/p27 in nude
mice xenografts
All mice were sacrificed by IP injection with an
overdose (35 mg/kg) of pentobarbital followed by cervical
dislocation. Tumor samples were fixed in 10% neutral buffered
formalin for 12 h and processed according to standard procedures.
Tumor sections were incubated overnight with monoclonal antibodies
against cyclin D1, cyclin E, Cip1/p21 or Kip1/p27 at a 1:400
dilution, followed by incubation with a biotinylated anti-mouse
secondary antibody. Then sections were exposed to
streptavidin-conjugated horseradish peroxidase and
3,3′-diaminobenzidine. Positive controls for each antibody were
included to confirm the adequacy of staining for each experiment.
The immunohistochemical method and a computer-assisted image
analysis system were used to detect the expression of cyclin D1,
cyclin E, Cip1/p21 and Kip1/p27 in the tumor tissues of nude
mice.
Statistical analysis
Statistical analysis of the data was performed using
SPSS version 19.0 (IBM SPSS, Armonk, NY, USA). Student's t-test was
used to compare between the mean values of two groups. Data from
each group were compared by one-way analysis of variance, followed
by the least significant difference test. Final values are
expressed as the means ± standard error of the mean (SEM).
P<0.05 was considered to indicate a statistically significant
difference.
Results
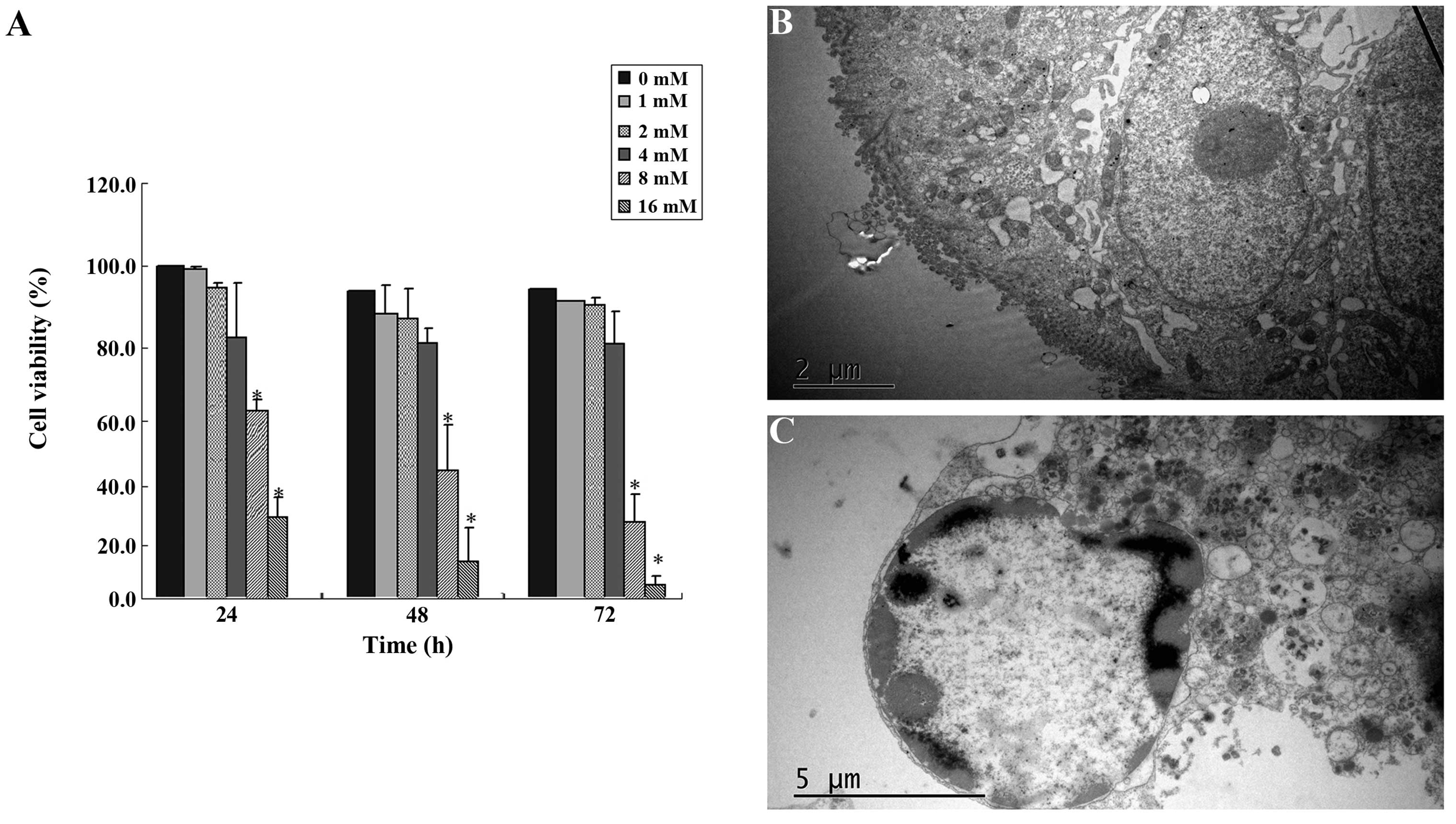
Cell viability assays and transmission
electron microscopy analysis of SW1116 cells following SIN
treatment
SIN inhibited SW1116 cell viability in a
dose-dependent manner (Fig. 1). The
estimated median effective concentrations (IC50) of SIN after 24,
48 and 72 h of incubation were 11.75, 9.85 and 7.91 mM,
respectively. SIN significantly reduced the viability of SW1116
cells at concentrations of 8 and 16 mM (Fig. 1A). We investigated the morphology of
SIN-treated cells using transmission electron microscopy. Cells
treated with 8 mM SIN for 48 h exhibited a morphology typical of
nuclear chromatin condensation (Fig.
1C) compared with the control cells without drug treatment
(Fig. 1B).
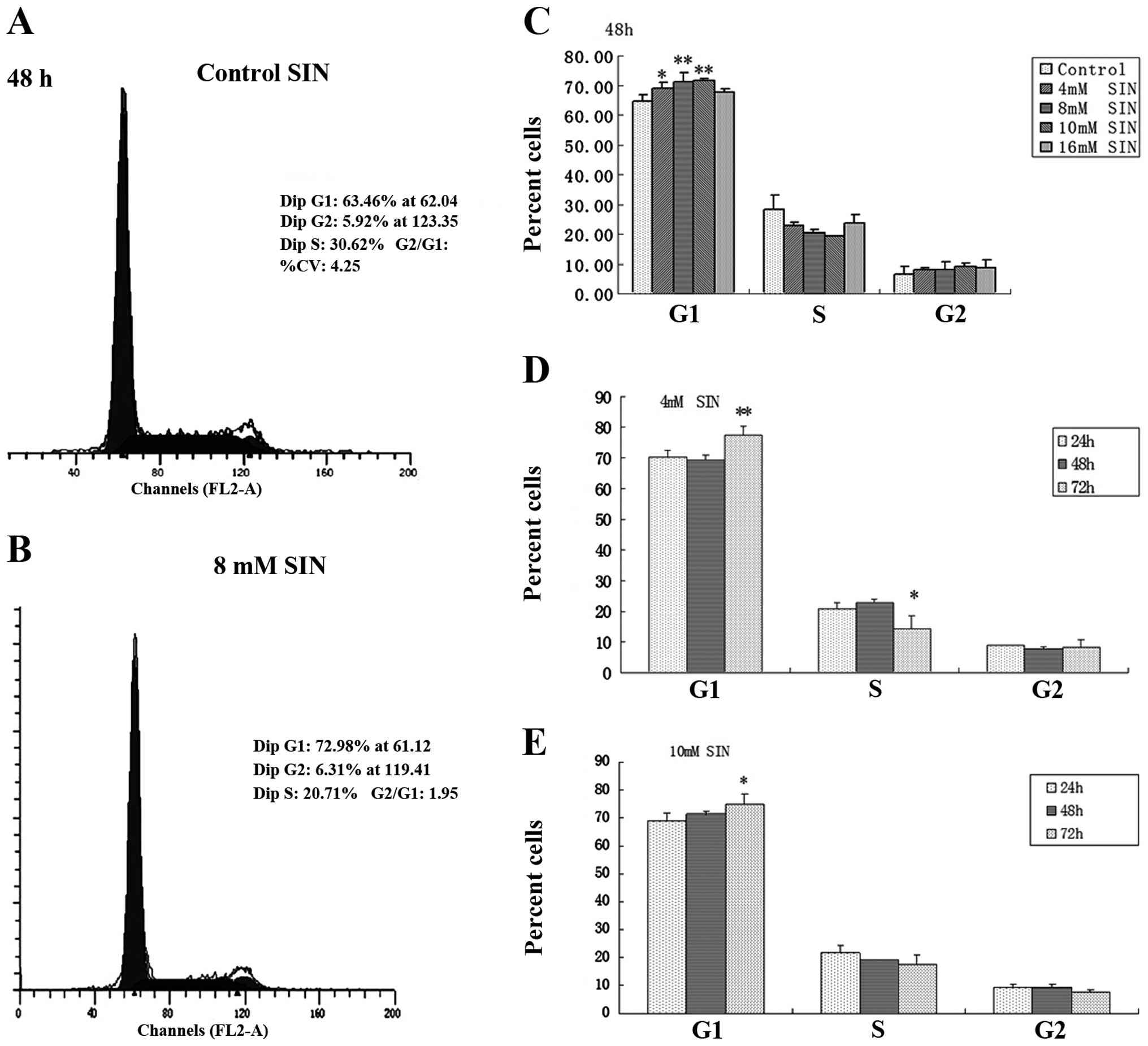
SIN causes cell cycle arrest in SW1116
cells
Based on its cell growth inhibitory effect, the
effect of SIN on cell cycle progression in SW1116 cells was
examined by cell cycle distribution analysis (Fig. 2). Following SIN treatment for 48 h, a
significant and dose-dependent accumulation of cells was noted in
the G1 phase, with a concomitant decrease in the percentage of
cells in the S phase compared with the control (0 mM SIN; Fig. 2C). Representative flow cytometry scans
of the control and SW1116 cells treated with 8 mM SIN for 48 h are
shown in Fig. 2A and B. These clearly
indicate an increase in the number of cells in the G1 phase. The
effect of SIN treatment for various incubation times (24, 48 and 72
h) on the cell cycle response in SW1116 cells was investigated at
concentrations of 4 and 10 mM. At a longer treatment time (72 h),
G1 arrest was observed compared with shorter treatment times (24 h;
Fig. 2D and E). These results suggest
that cell cycle arrest (predominantly in G1 phase) induced by SIN
underlies its cell growth inhibitory effect in colorectal cancer
cells.
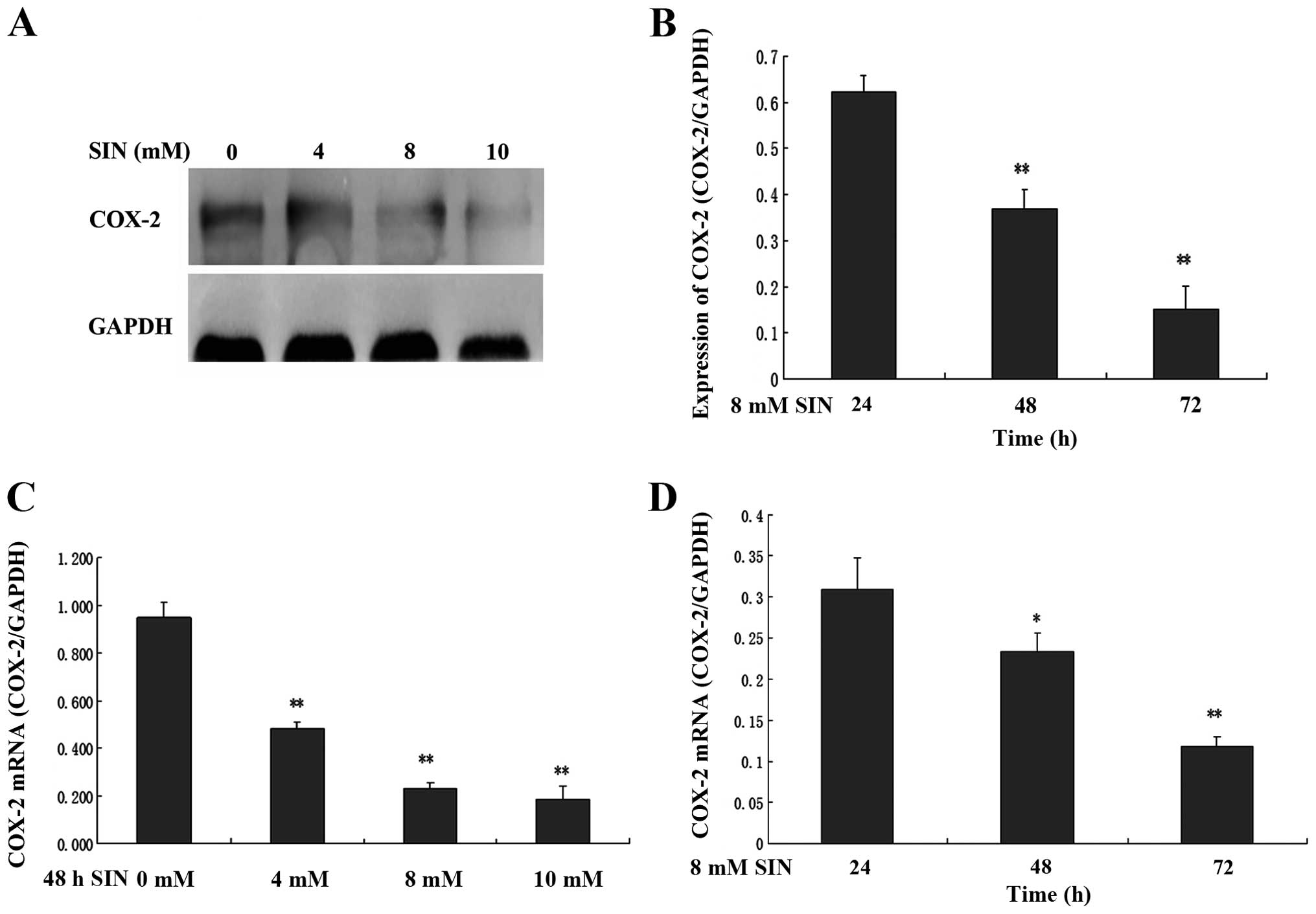
SIN modulates the expression of COX-2
in SW1116 cells
To determine whether the cell growth inhibitory
effect of SIN was correlated with the expression of COX-2, the
expression of COX-2 mRNA and protein in SW1116 cells was analyzed.
SW1116 cells were exposed to the indicated concentrations of SIN
(0, 4, 8 and 10 mM) for 24, 48 and 72 h. As shown in Fig. 3A and B, COX-2 protein expression
significantly decreased in a time- and dose-dependent manner in
cells treated with SIN. Consistent with the western blot analysis
results, the treatment of SW1116 cells with SIN led to a
significant decrease in cytoplasmic COX-2 mRNA expression in a
time- and dose-dependent manner (Fig. 3C
and D).
Anti-tumorigenic effects of SIN in
nude mice xenograft
The anti-tumorigenic effect of SIN on colon cancer
was further examined in vivo in nude mice xenografts. The
tumor growth inhibition efficacy of SIN was compared with that of
the standard chemotherapeutic drug 5-FU. As shown in Tables I and II, treatment with SIN (25, 50 and 100
mg/kg) or 5-FU (10 mg/kg) significantly increased tumor growth
inhibition from day 15 onwards. On the day of sacrifice (day 18),
the group that received a high dose of SIN (100 mg/kg) demonstrated
a tumor suppression rate of 50.8%, approaching that induced by 5-FU
alone (Table II). Along with the
extension of the intervention time, a trend towards an increase in
tumor suppression was observed from days 21–24 onwards.
 | Table I.Tumor volume (in mm3) of
nude mice xenografts (mean ± SEM). |
Table I.
Tumor volume (in mm3) of
nude mice xenografts (mean ± SEM).
| Treatment | Day 15 | Day 18 | Day 21 | Day 24 | Day 27 | Day 30 |
|---|
| Control |
156.0±29.4 |
222.1±66.8 |
280.1±107.2 |
311.8±119.8 |
383.3±145.1 |
456.9±141.7 |
| SIN (25 mg/kg) |
97.8±46.5b |
136.2±64.6b |
204.6±99.9a |
239.4±111.4a |
303.0±172.6 |
396.6±227.0 |
| SIN (50 mg/kg) |
104.8±48.1a |
140.3±57.7b |
191.6±65.9a |
228.8±80.2a |
307.7±149.2 |
393.4±158.0 |
| SIN (100
mg/kg) |
80.6±22.2b |
109.2±44.5b |
159.8±47.2b |
178.3±55.5b |
259.7±98.5 |
294.4±129.3 |
| 5-FU |
56.5±21.4b |
63.4±23.6b |
72.7±28.1b |
104.3±45.8a |
132.7±51.4b |
149.0±69.1b |
| Table II.Changes in tumor inhibitory rate in
the four treatment groups. |
Table II.
Changes in tumor inhibitory rate in
the four treatment groups.
|
| Tumor inhibitory
rate (%) |
|---|
|
|
|
|---|
| Treatment | Day 15 | Day 18 | Day 21 | Day 24 | Day 27 | Day 30 |
|---|
| SIN (25 mg/kg) | 37.3 | 38.7 | 27.0 | 23.2 | 20.9 | 13.2 |
| SIN (50 mg/kg) | 32.8 | 36.8 | 31.6 | 26.6 | 19.7 | 13.9 |
| SIN (100
mg/kg) | 48.3 | 50.8 | 42.9 | 42.8 | 32.2 | 35.6 |
| 5-fluorouracil | 63.8 | 71.5 | 74.0 | 66.5 | 66.4 | 67.4 |
The body weights of the animals in the control and
drug treatment groups were monitored throughout the experimental
period to asses drug-induced toxicity. SIN was not observed to
cause any significant decrease in the body weight of the mice from
days 0–18; however, a significant reduction in body weight was
observed on day 24 in the animals treated with a high dose of SIN
(Table III).
| Table III.Changes in body weight and mortality
rate of nude mice with SW1116 xenografts (n=8 per group). |
Table III.
Changes in body weight and mortality
rate of nude mice with SW1116 xenografts (n=8 per group).
|
| Change in body
weight |
|
|---|
|
|
|
|
|---|
| Treatment | Day 0 | Day 6 | Day 12 | Day 18 | Day 24 | Day 30 | Mortality |
|---|
| Control |
18.6±0.8 |
19.0±0.7 |
20.3±1.0 |
20.9±1.5 |
21.5±1.4 |
21.9±1.4 | 1 |
| SIN (25 mg/kg) |
19.2±0.8 |
19.1±1.1 |
19.9±0.9 |
20.3±1.0 |
21.5±1.2 |
22.1±1.3 | 0 |
| SIN (50 mg/kg) |
19.6±1.0 |
19.8±1.2 |
19.4±2.9 |
21.5±1.3 |
21.8±1.4 |
20.9±2.6 | 1 |
| SIN (100
mg/kg) |
17.9±1.1 |
18.0±1.2 |
18.7±1.2 |
18.8±1.7 |
19.5±2.2a |
18.2±4.0b | 2 |
| 5-fluorouracil |
17.7±1.0 |
18.1±1.1 |
18.4±1.7a |
19.1±3.0 |
21.4±1.5 |
22.2±1.3 | 0 |
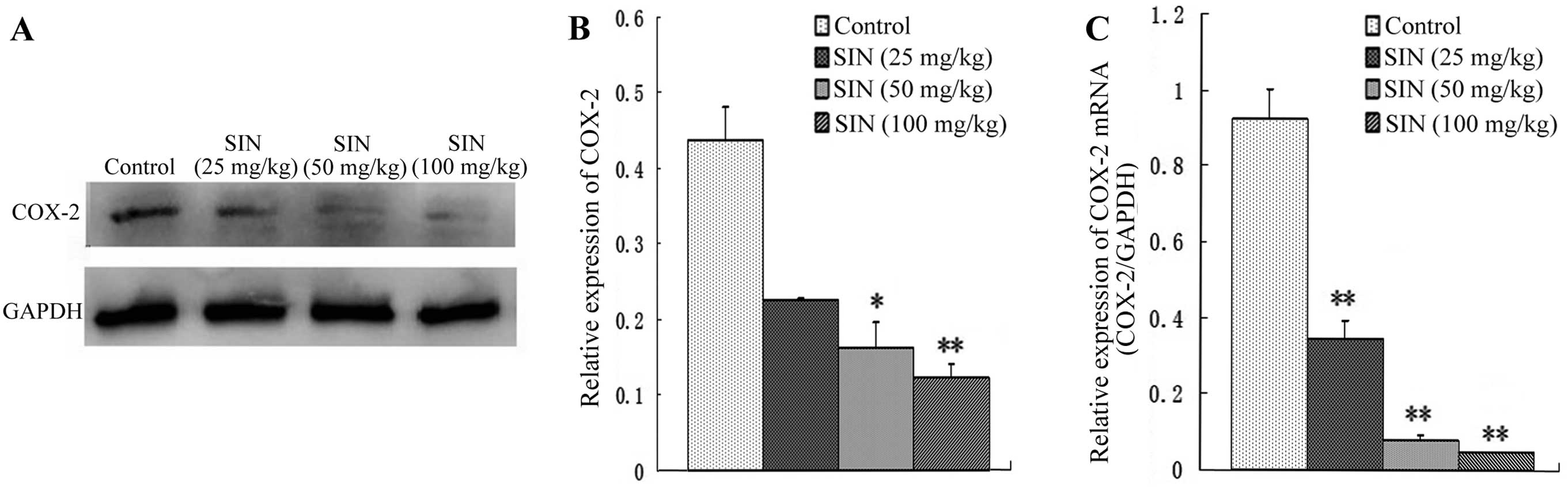
SIN modulates the expression of COX-2
in nude mice xenografts
To determine whether the tumor growth inhibitory
effect of SIN was correlated with COX-2 expression, the effect of
SIN treatment on COX-2 protein and mRNA levels was evaluated by
western blot analysis and qPCR. As shown in Fig. 4A and B, daily IP injections of SIN at
doses of 50 and 100 mg/kg/d administered to nude mice resulted in
dose-dependent decreases in COX-2 protein expression compared with
the control group. Similar results were observed for COX-2 mRNA
expression in the treatment group compared with the control group
(Fig. 4C).
SIN modulates the expression of cyclin
D1, cyclin E, Cip1/p21 and Kip1/p27 in nude mice xenografts
As SIN induced notable cell cycle arrest in the G1
phase in SW1116 cells, we next evaluated the in vivo effect
of SIN treatment on cell cycle regulators of the G1 phase. Cyclin
D1, Cyclin E, Cip1/p21 and Kip1/p27 expression was
immunohistochemically analyzed in tumor samples. Qualitative
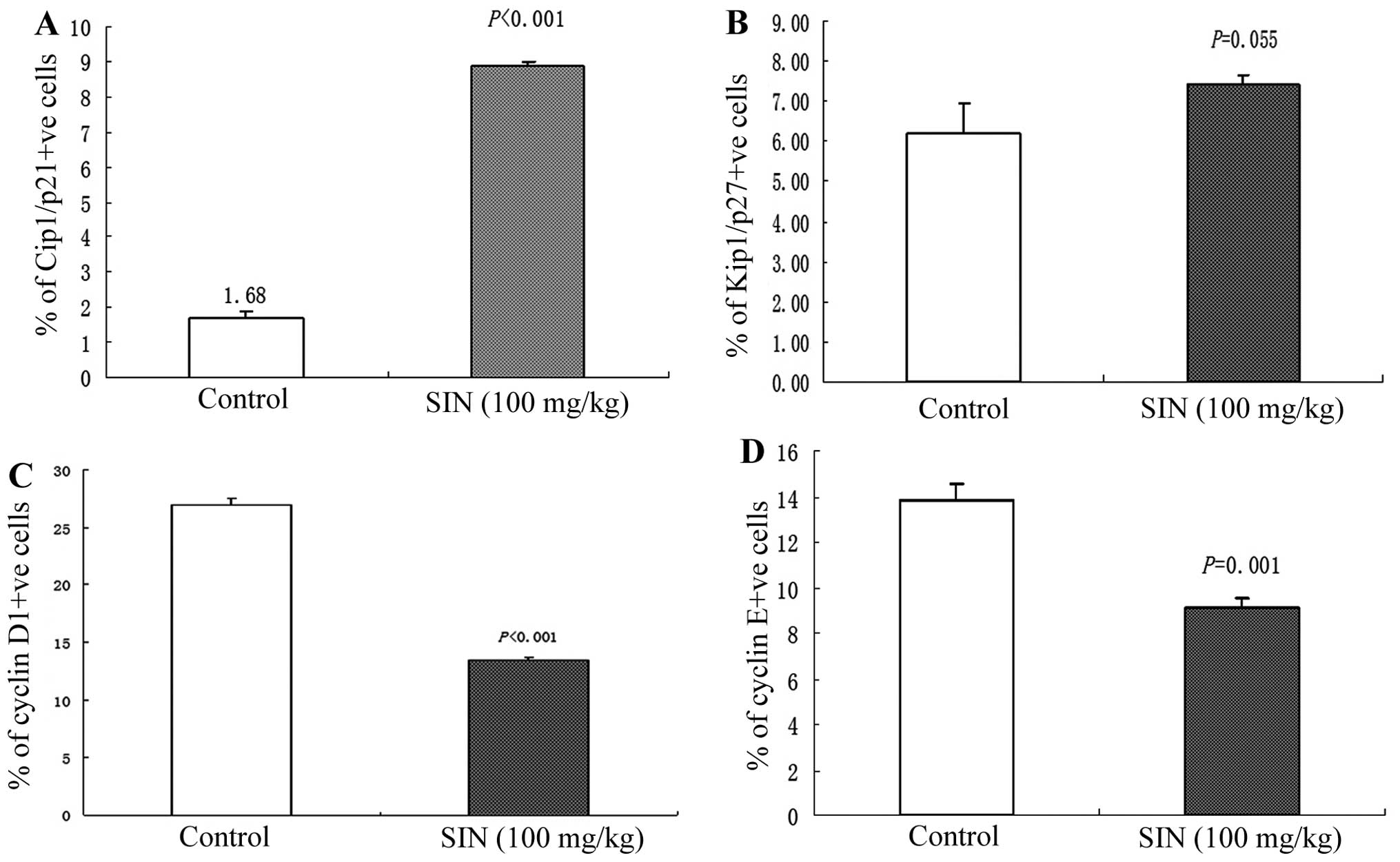
microscopic examination of Cip1/p21-stained tumor sections revealed
an increase in Cip1/p21-positive cells in tumors from SIN-treated
(100 mg/kg) mice compared with tumors from the control group
(Fig. 5A). Immunohistochemical
analysis of cyclin D1 and cyclin E protein expression in tumor
sections revealed a significant decrease (P<0.001) in cyclin D1
and cyclin E immunoreactivity in tumors from SIN-treated mice
(Fig. 5C and D). There was no
significant difference in the intensity of Kip1/p27-positive
staining between tumors from the SIN treatment (100 mg/kg) and
control group (Fig. 5B).
Discussion
The primary aim of the present study was to evaluate
the anticancer effects and associated mechanisms of SIN in human
colorectal cancer-derived SW1116 cells in vitro and to
translate the in vitro findings in an in vivo
preclinical model. SW1116 cell culture studies revealed that SIN
strongly inhibited SW1116 cell growth in a time- and dose-dependent
manner. The growth inhibitory effect was particularly notable in
cells treated with a mid-dose and high dose of SIN (8 and 16 mM).
However, the accumulation of cells in the G1 phase decreased as the
treatment dose of SIN increased to 16 mM. It was inferred that cell
cycle arrest might not be the only mechanism, and that other factor
may also be involved.
Based on the promising in vitro anticancer
effects of SIN against SW1116 cells, its efficacy was examined in a
preclinical animal model by ectopic implantation of SW1116
xenografts in athymic nude mice, which revealed an anti-tumorigenic
effect of SIN. The tumor growth inhibition efficacy of SIN was
compared with that of the standard chemotherapeutic drug 5-FU.
Treatment with SIN (25, 50 and 100 mg/kg) significantly increased
the tumor inhibition rate from day 15 onwards. On day 18, treatment
with high doses of SIN (100 mg/kg) resulted in tumor suppression of
50.8%, approaching that induced by 5-FU alone. However, with
increasing treatment time, the tumor inhibition rate tended to
decrease. We also monitored the body weight of animals throughout
the experimental period to assess drug-induced toxicity, which
revealed a significant reduction on day 24 in the body weight of
animals treated with a high dose of SIN. Based on these results, we
inferred that daily IP injection of SIN inhibits the growth of
SW1116 tumor xenografts, but that long-term treatment with
high-dose SIN (100 mg/kg) should be carefully investigated and
considered.
Previous studies have implicated COX-2 in
oncogenesis in a number of cancers (25–27), and
have demonstrated that COX-2 inhibitors are effective in the
prevention of the development of tumors, including colon cancer.
Increased expression of COX-2 is most likely associated with a poor
prognosis (28). We demonstrated that
SIN inhibits COX-2 expression in SW1116 cells and nude mice
xenografts. We thus inferred that COX-2 inhibition may be a
significant antitumor mechanism and that COX-2 may be a key target
of the drug activity of SIN.
Studies in SW1116 cell culture revealed that SIN may
inhibit cell growth via cell cycle arrest, specifically in G1
phase. We next evaluated the effect of SIN treatment on G1 phase
cell cycle regulators, including cyclin D1, cyclin E, Cip1/p21 and
Kip1/p27. Cell cycle regulation requires the periodic formation,
activation and inactivation of unique protein kinase complexes
composed of cyclin (regulatory) and cyclin-dependent kinase (CDK;
catalytic) subunits (29). The
association of cyclin D1 and CDK4, cyclin E and cdk2 result in
rubidium phosphorylation in the G0/G1 and G1/S phase transitions of
the cell cycle (30–32). The cdk inhibitors Cip1/p21 and
Kip1/p27 have been proposed to exert redundant functions in cell
cycle progression (33–36). Cyclin levels are strictly controlled
by precise synthesis and degradation at the appropriate time points
during cell cycle progression (37).
Immunohistochemical analysis revealed an increase in Cip1/p21
expression and a decrease in cyclin D1 and cyclin E expression,
suggesting an effect of SIN on cell cycle arrest in vivo.
Abnormal accumulation of cyclin E, cyclin D1 and Cip1/p21 could in
turn prevent SW1116 cells from entering the mitotic phase, which
might cause a delay in subsequent cell cycle progression. Taken
together, the present data demonstrate that SIN has a significant
antitumor effect in vivo, and that these molecular
alterations by SIN may lead to cell cycle arrest.
In summary, we have demonstrated that SIN inhibits
human colon cancer cell growth in vitro and in vivo
via COX-2 inhibition and cell cycle arrest in G1 phase. In contrast
to standard chemotherapy with cytotoxic drugs, mid/low doses (25
and 50 mg/kg) of this herbal molecule induce antitumor effects with
reduced toxicity. These results suggest the potential of SIN for
development as an alternative treatment option or adjuvant
chemotherapeutic agent in colon cancer therapy.
Acknowledgements
The authors would like to thank Yanchun Ma for
technical assistance and Dr Peihao Yin for valuable advice. This
study was funded by the Shanghai Education Committee (2012JW66) and
by the Putuo district Science and Technology Committee
(2012PTKW008).
References
|
1
|
Ekberg M, Callender M, Hamer H and Rogers
S: Exploring the decision to participate in the national health
service bowel cancer screening programme. Eur J Cancer Prev.
23:391–397. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hou L, Jiang J, Liu B, Nasca PC, Wu Y, Zou
X, Han W, Chen Y, Zhang B, Xue F, et al: Association between
smoking and deaths due to colorectal malignant carcinoma: a
national population-based case-control study in China. Br J Cancer.
110:1351–1358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Estimating the world cancer burden: Globocan 2000. Int J Cancer.
94:153–156. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chatenoud L, Bertuccio P, Bosetti C,
Malvezzi M, Levi F, Negri E and La Vecchia C: Trends in mortality
from major cancers in the Americas: 1980–2010. Ann Oncol.
25:1843–1853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zaniboni A and Labianca R: Gruppo Italiano
per lo Studio e la Cura dei Tumori del Digerente: Adjuvant therapy
for stage II colon cancer: an elephant in the living room? Ann
Oncol. 15:1310–1318. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rao S and Cunningham D: Adjuvant therapy
for colon cancer in the new millenium. Scand J Surg. 92:57–64.
2003.PubMed/NCBI
|
|
7
|
Sinha VR and Honey C: Critical aspects in
rationale design of fluorouracil-based adjuvant therapies for the
management of colon cancer. Crit Rev Ther Drug Carrier Syst.
29:89–148. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lotfi-Jam K, Carey M, Jefford M, Schofield
P, Charleson C and Aranda S: Nonpharmacologic strategies for
managing common chemotherapy adverse effects: a systematic review.
J Clin Oncol. 26:5618–5629. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Latimer N, Lord J, Grant RL, O'Mahony R,
Dickson J and Conaghan PG: Value of information in the
osteoarthritis setting: Cost effectiveness of COX-2 selective
inhibitors, traditional NSAIDs and proton pump inhibitors.
Pharmacoeconomics. 29:225–237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jacobo-Herrera NJ, Pérez-Plasencia C,
Camacho-Zavala E, González GF, Urrutia EL, García-Castillo V and
Zentella-Dehesa A: Clinical evidence of the relationship between
aspirin and breast cancer risk (review). Oncol Rep.
32:4512014.PubMed/NCBI
|
|
11
|
Brasky TM, Liu J, White E, et al:
Non-steroidal anti-inflammatory drugs and cancer risk in women:
results from the Women's Health Initiative. Int J Cancer.
135:1869–1883. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu X, Cai M, Ji F and Lou LM: The impact
of COX-2 on invasion of osteosarcoma cell and its mechanism of
regulation. Cancer Cell Int. 14:272014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liggett JL, Zhang X, Eling TE and Baek SJ:
Anti-tumor activity of non-steroidal anti-inflammatory drugs:
cyclooxygenase-independent targets. Cancer Lett. 346:217–224. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Davies NM and Jamali F: COX-2 selective
inhibitors cardiac toxicity: getting to the heart of the matter. J
Pharm Pharm Sci. 7:332–336. 2004.PubMed/NCBI
|
|
15
|
Zhang MF, Zhao Y, Jiang KY, et al:
Comparative pharmacokinetics study of sinomenine in rats after oral
dministration of sinomenine monomer and sinomenium acutum extract.
Molecules. 19:12065–12077. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao T, Hao J, Wiesenfeld-Hallin Z, et al:
Analgesic effect of sinomenine in rodents after inflammation and
nerve injury. Eur J Pharmacol. 721:5–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mo ZX, An SL and Zhou JY: Effects of
caulis sinomenii and sinomenine on morphine-induced place
preference and brain histamine level in mice. Nan Fang Yi Ke Da Xue
Xue Bao. 26:1709–1713. 2006.(In Chinese). PubMed/NCBI
|
|
18
|
Yamasaki H: Pharmacology of sinomenine, an
anti-rheumatic alkaloid from sinomenium acutum. Acta Med Okayama.
30:1–20. 1976.PubMed/NCBI
|
|
19
|
Candinas D, Mark W, Kaever V, et al:
Immunomodulatory effects of the alkaloid sinomenine in the high
responder ACI-to-Lewis cardiac allograft model. Transplantation.
62:1855–1860. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang T, Zhou L, Zhang W, Qu D, Xu X, Yang
Y and Li S: Effects of sinomenine on proliferation and apoptosis in
human lung cancer cell line NCI-H460 in vitro. Mol Med Rep.
3:51–56. 2010.PubMed/NCBI
|
|
21
|
Zhou L, Luan H, Liu Q, Jiang T, Liang H,
Dong X and Shang H: Activation of PI3K/Akt and ERK signaling
pathways antagonized sinomenine-induced lung cancer cell apoptosis.
Mol Med Rep. 5:1256–1260. 2012.PubMed/NCBI
|
|
22
|
Kues WA, Anger M, Carnwath JW, et al: Cell
cycle synchronization of porcine fetal fibroblasts: effects of
serum deprivation and reversible cell cycle inhibitors. Biol
Reprod. 62:412–419. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen M, Huang J, Yang X, et al: Serum
starvation induced cell cycle synchronization facilitates human
somatic cells reprogramming. PLoS One. 7:e282032012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tan Y, Sun X, Xu M, et al: Efficacy of
recombinant methioninase in combination with cisplatin on human
colon tumors in nude mice. Clin Cancer Res. 5:2157–2163.
1999.PubMed/NCBI
|
|
25
|
Tjandrawinata RR, Dahiya R and
Hughes-Fulford M: Induction of cyclo-oxygenase-2 mRNA by
prostaglandin E2 in human prostatic carcinoma cells. Br J Cancer.
75:1111–1118. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maekawa M, Sugano K, Sano H, et al:
Increased expression of cyclooxygenase-2 to −1 in human colorectal
cancers and adenomas, but not in hyperplastic polyps. Jpn J Clin
Oncol. 28:421–426. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hao X, Bishop AE, Wallace M, et al: Early
expression of cyclo-oxygenase-2 during sporadic colorectal
carcinogenesis. J Pathol. 187:295–301. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sheehan KM, Sheahan K, O'Donoghue DP,
MacSweeney F, Conroy RM, Fitzgerald DJ and Murray FE: The
relationship between cyclooxygenase-2 expression and colorectal
cancer. JAMA. 282:1254–1257. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kõivomägi M and Skotheim JM: Docking
interactions: cell-cycle regulation and beyond. Curr Biol.
24:R647–R649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dorée M and Hunt T: From Cdc2 to Cdk1:
when did the cell cycle kinase join its cyclin partner? J Cell Sci.
115:2461–2464. 2002.PubMed/NCBI
|
|
31
|
Sherr CJ and Roberts JM: Living with or
without cyclins and cyclin-dependent kinases. Genes Dev.
18:2699–2711. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fisher D, Krasinska L, Coudreuse D and
Novák B: Phosphorylation network dynamics in the control of cell
cycle transitions. J Cell Sci. 125:4703–4711. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yoon MK, Mitrea DM, Ou L and Kriwacki RW:
Cell cycle regulation by the intrinsically disordered proteins p21
and p27. Biochem Soc Trans. 40:981–988. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weinberg WC and Denning MF: P21Waf1
control of epithelial cell cycle and cell fate. Crit Rev Oral Biol
Med. 13:453–464. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shankland SJ and Wolf G: Cell cycle
regulatory proteins in renal disease: Role in hypertrophy,
proliferation and apoptosis. Am J Physiol Renal Physiol.
278:F515–F529. 2000.PubMed/NCBI
|
|
36
|
Todd R, Hinds PW, Munger K, Rustgi AK,
Opitz OG, Suliman Y and Wong DT: Cell cycle dysregulation in oral
cancer. Crit Rev Oral Bio Med. 13:51–61. 2002. View Article : Google Scholar
|
|
37
|
Schafer KA: The cell cycle: a review. Vet
Pathol. 35:461–478. 1998. View Article : Google Scholar : PubMed/NCBI
|