Introduction
Ovarian cancer only accounts for ~4% of cancer cases
in women, but it is the eighth most common cause of
cancer-associated mortality resulting from gynecological tumors
worldwide (1,2). Epithelial ovarian cancer (EOC), which is
categorized into serous, mucinous, endometrioid and non-clear cell
types, accounts for 90% of all ovarian cancer (3,4). The
earlier cancer is identified, the better the prognosis; however,
ovarian cancer is difficult to detect early, as overt symptoms do
not occur until later stages of the disease. Despite of advanced
diagnostic technologies such as the combination of computed
tomography/positron emission tomography scan and application of
novel targeted drugs such as Bevacizumab and Olaparib, the 5-year
survival rate ranges between ~30–50% (1,5).
Furthermore, EOC is characterized by accelerated and aggressive
growth, which may result in high recurrence rates. These features
comprise the major challenges for the early diagnosis and effective
treatment of patients with EOC (6).
For a number of years, specific diagnostic biomarkers have been
reported in various studies of EOC (7,8); Chong
et al (9) conducted a
microarray based microRNA expression study in primary and recurrent
EOC patient tissues samples and identified 8 specific miRNAs whose
expression was most significantly changed. In addition, Eitan et
al (10) identified miR-200a,
miR-34a, and miR-449b as the most downregulated miRNAs in advanced
(stage III) ovarian tumors. Fan et al (11) reported the association of high levels
of miR-196a expression and worse overall survival in ovarian cancer
patients, especially in advanced stage tumors. Single nucleotide
polymorphisms (SNPs) have also been examined as potential EOC risk
indictors (12–14). However, the mechanism and genetic
factors involved in the progression of this cancer remain to be
elucidated.
MicroRNAs (miRNAs/miRs) are non-coding RNAs with
complex biological functions that regulate the expression of
numerous proteins through direct binding to the 3′-untranslated
region of target genes (15,16). It has been well-documented that miRNAs
are involved in a wide range of ovarian cancer tumorigenic
processes, including malignant transformation, differentiation,
proliferation and apoptosis (17–19).
Furthermore, SNPs located in miRNAs (miRSNPs) have
begun to draw attention, due to their critical regulatory role in
cancer progression. miRSNPs influence the transcription of the
primary target gene, disturbing pri-/pre-miRNA processing, or
affecting miRNA-mRNA interactions (20).
A previous study located the common genetic variant
rs11614913 in the 3p mature miRNA region of Homo sapiens
(hsa)-miR-196a2, and revealed that it led to a variation from G:T
to G:C in the stem region of the miR-196a-2 precursor (21). Previous studies have suggested that
this SNP may affect the processing of pre-miRNA and is associated
with risk in various types of cancer, including breast and
colorectal cancer (22–24). However, the genetic factors associated
with EOC susceptibility have not previously been identified. It was
hypothesized that the miR-196a-2 genotype may be associated with
susceptibility to EOC. The present study aimed to investigate this
hypothesis in a population-based case-control study and determine
the function of this variant in the OVCAR3 cell line in
vivo.
Materials and methods
Study population
The study protocol was approved by the Institutional
Review Board of Zhengzhou University (Zengzhou, China) and Henan
Academy of Medical Science (Zengzhou, China). The present study
included 479 women with EOC that underwent tumor resections in the
First Affiliated Hospital of Zhengzhou University and Woman and
Infants Hospital of Zhengzhou between 2006 and 2012. In addition,
431 healthy women living in the same area, without any history of
hereditary or malignant disease were selected as a control group.
All patients and controls were of the one ethnic group (Han
Chinese). Healthy control individuals were recruited from a large
pool of individuals seeing a physician for routine health checkups
in the First Affiliated Hospital of Zhengzhou University and Woman
and Infants Hospital of Zhengzhou. Both cases and controls were
prospectively recruited and matched by age and ethnicity.
Demographic and epidemiology information, including height and
weight were collected for all subjects. Histopathological diagnoses
and clinical stages were classified based on the criteria of the
International Federation of Gynecology and Obstetrics (25). Genomic DNA was immediately extracted
from 5 ml peripheral blood samples of all patients using a Wizard
Genomic DNA Purification kit (Promega Corp., Madison, WI, USA) and
stored at −80°C. Written informed consent was obtained from all
participants or their families if direct consent could not be
obtained.
Genotyping
Genotyping was performed using a polymerase chain
reaction (PCR)-based restriction fragment length polymorphism
assay. The following primers were used to amplify the rs11614913
T>C polymorphism site in the miR-196a-2 precursor: F
5′-CCCCTTCCCTTCTCCTCCAGATA-3′ and R 5′-CGAAAACCGACTGATGTAACTCCG-3′.
All primers was synthesized by Shengong Company (Shanghai, China).
PCR was performed with a total volume of 25 µl with 100 ng DNA
template, 2.5 µl of 10X PCR buffer, 1 U of Taq DNA polymerase, 0.2
mM dNTPs (Invitrogen, Carslbad, CA, USA) and 0.5 µmol/l of each
primer. The PCR conditions were 94°C for 5 min followed by 35
cycles of 30 sec at 94°C, 30 sec at 63°C, and 1 min at 72°C, and
final elongation step at 72°C for 10 min. A total of 10 µl PCR
product was then digested using 2 µl (10 U/µl) Fermentas
MspI restriction enzyme (Thermo Fisher Scientific, Inc.,
Pittsburgh, PA, USA) for 16 h at 37°C. The resulting fragments were
separated by electrophoresis on a 3.0% agarose gel (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) and visualized in three
distinct patterns of restriction fragments. The CC genotype
produced two fragments (125 and 24 bp), the TT homozygote produced
one 149-bp fragment and the TC heterozygote produced three
fragments (125, 149 and 24 bp). The experiment was performed in
triplicate; 10% of the PCR products were randomly selected from
each of the miRNA polymorphism patterns for repeat assay and
validated by direct DNA sequencing using an ABI3730xl DNA Analyzer
(Applied Biosystems, Foster City, CA, USA).
Tissue samples
To detect the expression levels of miRNA-196a,
ovarian cancer tissue samples were collected from 83 patients who
were histopathologically diagnosed with EOC and had undergone
resection for EOC between June and October 2010 at Henan Cancer
Hospital (Zhengzhou, China). The recruitment process was the same
as outlined above. No patients had received radiotherapy or
chemotherapy prior to undergoing surgery.
Reverse transcription-quantitative PCR
(RT-qPCR)
RNA was isolated from the ovarian cancer tissues
using the Applied Biosystems mirVana miRNA Isolation kit (Thermo
Fisher Scientific, Inc.). The quality of miRNA extracted was
determined using a 2100 Bioanalyzer (Agilent Technologies, Inc.,
Santa Clara, CA, USA). Samples with a high RNA Integrate Number
(RIN) of miRNA (>7.0) were included in subsequent
investigations. RIN was calculated using Agilent 2100 Expert
software (Agilent Biotechnology) (26). RNA was extracted from 83 tissue
samples and nearly 40 were selected for RIN calculation in the
present study.
An RT-qPCR assay (Applied Biosystems TaqMan
microRNA; Thermo Fisher Scientific, Inc.) was used to determine
mature miRNA expression levels. The primers were as follows:
Forward, 5′-GCTCTGGCTCCGTGTCTTCACTCCC-3′ and reverse,
5′-TGCCCCAGCACAGCCCCCGTCCCCC-3′. The following primers were used to
amplify the β-actin gene, which was used as the internal control:
Forward, 5′-AGAAAATCTGGCACCACACC-3′ and reverse,
5′-GGGGTGTTGAAGGTCTCAAA-3′. Small nuclear RNA U6 (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used for
normalization. The PCR was performed in a 20 µl reaction (2 µl of
template cDNA, 1 µl of 10 µmol/l primers, 10 µl of 2X SYBR Green
Master Mix, 4 µl of 25 mmol/l Mg2+ and 2 µl of
ddH2O) was performed with the following cycling
parameters: 3 min at 95°C followed by 40 cycles at 95°C for 10 sec
and 60°C for 30 sec. All quantitative reactions were performed in
an Applied Biosystems ABI 7000 Real-Time PCR System (Thermo Fisher
Scientific, Inc.) in triplicate. Quantification cycle values were
obtained from Applied Biosystems ABI PRISM 7000 SDS software,
version 1.2.3 and used to calculate relative expression: Transcript
levels were calculated according to the comparative cycle threshold
(Cq). The 2−(∆∆Cq) method was applied to present final
results, where ∆Cq for each sample was determined by subtracting
the value for RNA U6 gene. Only triplicates with Cq values with a
standard deviation <0.20 were acceptable.
Transient transfection and miRNA
expression in an ovarian cancer cell line
To further explore the influence of this
polymorphism on the production of mature miRNA, the commercial
expression plasmid pCMV-MIR-196a-2 rs11614913-T/C (OriGene
Technologies, Inc., Beijing, China) was transfected into OVCAR3
cells (American Type Culture Collection, Manassas, VA, USA), which
exhibits wild-type and low expression of miR-196a. Briefly,
5×104 OVCAR3 cells/well were seeded on 24-well plates 24
h prior to transfection. A total of 1 µg of DNA vector diluted in
100 µl Opti-MEM I (ThermoFisher Scientific); 3 µl transfection
agent MegaTran 1.0 (OriGene) was added to diluted DNA and incubate
for 10 min at room temperature; 100 µl Megatran/DNA mixture was
then added into each well and incubate 37°C for 48 h. An empty
vector (pCMV-MIR) transfection was performed in parallel as a
negative control. Transfection efficiency was confirmed by
monitoring the GFP signal contained in the vector. The total RNA of
cells that carried pCMV miR-196a-2 rs11614913 T or C was extracted
and qPCR was used to detect miR-196a expression following 72 h of
culture.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
cell proliferation assay
Cell proliferation was evaluated using an MTT cell
viability assay. Cells (3×103/ml) were initially seeded
in a 96-well plate and collected once growth in the log phase was
achieved. A total of 10 µl MTT (5 mg/ml; Sigma-Aldrich) was added
at 24 and 36 h following plasmid transfection, and cells were
cultured for a further 4 h. Subsequently, cells were lysed with
dimethyl sulfoxide (Sigma-Aldrich, St. Louis, MO, USA). The ratio
of optical density (OD) was measured at an absorbance of 490 nm
using a Bio-Rad Laboratories 680 microplate reader, and the assay
was repeated a minimum of three times.
Cell invasion and migration assay
Trypsinized cells of the various groups were
collected from culture flasks and resuspended at a density of
2×105/ml in Dulbecco's modified Eagle's medium
(Sigma-Aldrich). This cell suspension (200 µl) was added to the
upper chamber of an 8-µm pore-size Transwell® insert (Corning Life
Sciences, Carlsbad, CA, USA) in 6 wells. For the invasion assay,
the Transwell was coated with 100 µl Matrigel (Corning Life
Sciences). Subsequently, 600 µl RPMI-1640 culture solution
(Sigma-Aldrich) containing 10% fetal bovine serum was added to the
lower chamber of each well. Incubation at 37°C occurred for 24 h
for migration and 48 h for invasion detection. Non-migratory cells
on the upper surface of the membrane were removed, stained with
0.1% crystal violet and counted: the number of invading and
migrated cells under the microscope at approximately 40X total
magnification. Count cells in five randomly selected fields in
triplicate.
Statistical analysis
The SPSS software package (version 11.5; SPSS, Inc.,
Chicago, IL, USA) was used for all statistical analyses.
Hardy-Weinberg equilibrium analysis was initially performed and
then the χ2 test, was used to compare the distribution
of SNPs between EOC patients and healthy controls. The odds ratio
(OR) and 95% confidence interval (CI) were calculated using an
unconditional logistic regression model. P<0.05 was considered
to indicate a statistically significant difference.
Results
Patient characteristics
The current results showed that 164 patients with
EOC were stage I/II, while 315 were stage III/IV. The predominant
tumor pathology type observed was serous-papillary (163 samples),
followed by 121 endometrioid, 95 undifferentiated, 60 mucinous and
40 no clear cell-type samples (Table
I). The 431 healthy control women exhibited a similar age
distribution to that of the patients with EOC included in the
current study.
 | Table I.Baseline characteristics of the EOC
cases and controls investigated. |
Table I.
Baseline characteristics of the EOC
cases and controls investigated.
| Characteristic | EOC patients,
n(n=479) | Controls,
n(n=431) |
|---|
| Age, years |
|
|
|
<41 | 75 | 67 |
|
41–50 | 118 | 112 |
|
51–60 | 168 | 156 |
|
>60 | 118 | 106 |
| Tumor
histology |
|
|
|
Serous | 163 |
|
|
Mucinous | 60 |
|
|
Endometrioid | 121 |
|
|
Undifferentiated | 95 |
|
| No cell
type | 40 |
|
| Tumor stage |
|
|
|
I/II | 164 |
|
|
III/IV | 315 |
|
miR-196a-2 polymorphism CC genotype
increases EOC risk
The observed genotype distribution of the miR-196a-2
polymorphism in the EOC and control groups conformed to the
Hardy-Weinberg equilibrium (P=0.600 and P=0.243, respectively). The
current results revealed a significantly higher frequency of CT and
CC genotypes in the patients compared with that of the control
group (51.6 vs. 47.1%, P=0.063 and 25.2 vs. 20.0%, P=0.018,
respectively; Table II). Subsequent
grouping of the TT and CT genotypes in the recessive genetic model
revealed a significantly increased risk of ovarian cancer in CC
genotype carriers when compared with that of the wild-type
homozygous TT and heterozygous CT genotype carriers (OR, 1.36; 95%
CI, 1.04–2.17; P=0.023).
| Table II.Association between the
microRNA-196a-2 single nucleotide polymorphism and the risk of
EOC. |
Table II.
Association between the
microRNA-196a-2 single nucleotide polymorphism and the risk of
EOC.
| Genotype | EOC cases, n (%;
n=479) | Control, n (%;
n=431) | ORa (95% CI) | P-value |
|---|
| TT | 111 (23.2) | 142 (32.9) |
1
(Reference) |
|
| CT | 247 (51.6) | 203 (47.1) | 1.53
(0.77–2.12) | 0.063 |
| CC | 121 (25.2) | 86
(20.0) | 1.78
(1.03–2.05) | 0.018 |
| TT/CTb | 358 (74.8) | 345 (80.0) |
1
(Reference) |
|
rs11614913-C enhances mature miR-196a
expression
The effect of miR-196a rs11614913 on mature miR-196a
expression was analyzed in 83 ovarian cancer tissue samples.
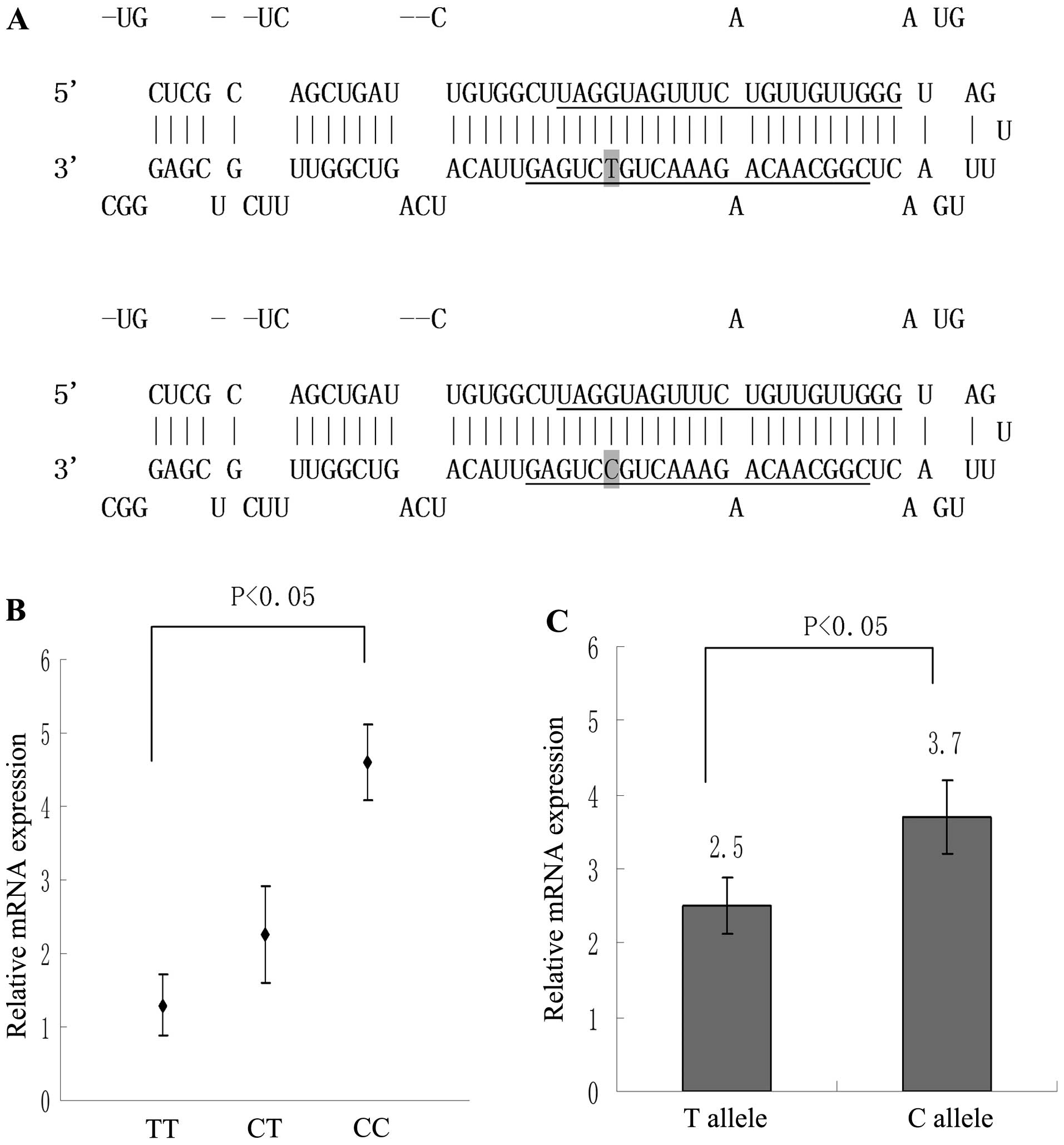
Fig. 1A indicates the site-specific
mutagenesis at rs11614913 from T to C in the plasmid used in the
current study. Significantly upregulated expression of miR-196a was
observed in CC genotype patients compared with that of the TT
genotype (P<0.05; Fig. 1B).
Furthermore, the CT genotype demonstrated increased expression of
miR-196a compared with that of the TT genotype, however, no
significant difference was observed (P>0.05; Fig. 1B). Following 72 h of culture,
pCMV-MIR-miR-196a-2 rs11614913-C-transfected OVCAR3 cells expressed
significantly increased expression levels of miR-196a when compared
with those of the wild-type T allele group (P<0.01; Fig. 1C).
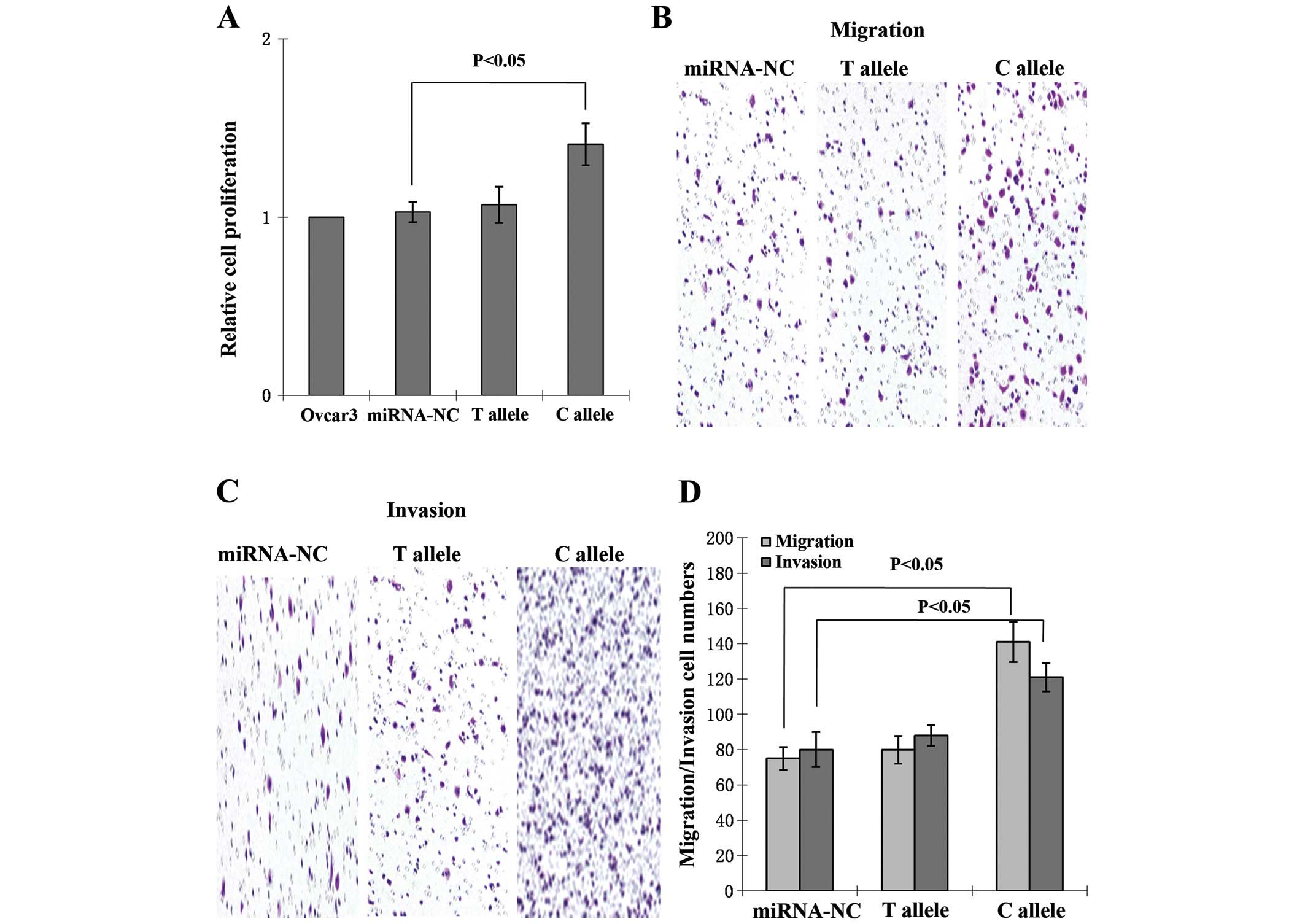
miR-196a SNP promotes cell
proliferation, migration and invasion
Following transfection, no significant differences
in cell viability were observed between the control and T allele
groups. Proliferation in the C allele group was significantly
increased compared with that of the control group (P<0.05;
Fig. 2A). The migration capacity of C
allele-transfected cells was significantly enhanced compared with
that of the non-transfected and T allele-transfected groups
(Fig. 2B). The number of cells
migrating to Matrigel was counted, and compared with
non-transfected and T allele-transfected cells, invasion was
increased in C allele-transfected cells (Fig. 2C). The increases in migration and
invasion rates were significant (P<0.05; Fig. 2D).
Discussion
It is well established that miRNAs participate in
the initial and developmental stages of numerous types of cancer,
as oncogenes or tumor suppressors (27–29).
However, the correlation between miRNA coding region variants and
cancer risk, as well as prognosis, has not been fully elucidated
(30,31). The present study revealed the
distribution frequencies of miRNA miR-196a-2 rs11614913 in a Han
Chinese population with EOC. An association between the CC genotype
of miR-196a-2 rs11614913 and a 1.34-fold increase in ovarian cancer
risk was observed. In addition, it was demonstrated that the
miR-196a-2 rs11614913 C allele induced mature miR-196a expression
in vivo in tissue and in vitro in cells. Finally,
mechanism data suggested that the alternation of mature miR-196a-2
expression may result in abnormal cell viability and
migration/invasion capacity, which may eventually be responsible
for increasing susceptibility to ovarian cancer. To the best of our
knowledge, the present study is the first combined clinical and
cell functional study of the association between rs11614913 SNP and
susceptibility to ovarian cancer.
It is widely accepted that gene variants located in
pri- and pre-miRNA regions may influence the biological functions
of miRNAs and eventually lead to alterations in disease incidence
(32). Considering this, various
studies have investigated the application of pri- and pre-miRNA
polymorphisms as predictors of cancer risk (29,31,33).
Notably, miRNAs are frequently located in cancer-associated genomic
regions (34) and may regulate almost
all cancer-associated genes (35).
The SNPs in miRNAs may perturb multiple miRNA-mediated gene
regulations and make it a novel cancer diagnostic and prognostic
biomarker. One example is the G to C variant (rs2910164) of the
miR-146a precursor, which induces a change from a G:U pair to a C:U
mismatch in the stem region. Evidence indicates that this variant
may facilitate early diagnosis in patients with ovarian cancer
(36). Another study demonstrated
that carriers of the variant homozygote CC of miR-196a-2 were more
likely to develop gastric cancer compared with wild-type homozygote
TT and heterozygote CT carriers (adjusted OR, 1.57; 95% CI,
1.03–2.39; P=0.038) (37). In the
same study, it was also indicated this C allele was significantly
associated with lymph node metastasis of gastric cancer (adjusted
OR, 2.25; 95% CI, 1.21–4.18; P=0.011) (37).
Hu et al (30)
previously reported significantly higher expression of miR-196a in
non-small cell lung tumor samples with CC genotypes compared with
that of CT and TT individuals. However, no observed differential
expression was found in pri- and pre-miRNA sequences based on
genotype in subsequent studies (21,38).
Hoffman et al (33)
demonstrated upregulation of mature miR-196a expression in breast
cancer cells transfected with pre-miR-196a-C compared with
pre-miR-196a-T-transfected empty vector control; however, marked
differential expression of the precursor miRNA was not observed in
this study. Similarly, the present study revealed upregulation of
miR-196a in patients with ovarian cancer who carried the C allele
and in pre-miR-196a-C-transfected cells. Similarly, the present
study identified that CC genotype carriers were more susceptible to
ovarian cancer compared with TT and CT genotype carriers. Together,
these results suggest that the rs11614913 polymorphism may affect
the processing of the pre-miRNA to its mature form.
To clarify the impact of miR-196a on tumor cells, a
series of functional experiments were performed in the present
study. It was observed that elevated miR-196a expression from C
allele-transfected cells promoted cell proliferation, migration and
invasion capacity in vitro. A previous study, conducted in
colorectal cancer, demonstrated a positive correlation between
enhanced migration, invasion ability and increased expression
levels of miR-196a (39). Results
derived from studies of other types of cancer, including lung
(38), colorectal (40) and early breast (22) cancer, also demonstrated similar
results, suggesting that the miR-196a-2 genotype may result in
altered processing of the pre-miRNA. However, other studies have
reported the absence of an association between miR-196a-2
rs11614913 and the risk for various types of cancer or disease
(41–43). These contradictory results may be a
result of population sample selection bias or differences in human
genotype distribution. A large population-based gene variant
correlation study should be conducted to overcome this
limitation.
Despite the merits of the current study, including
it being the first report of miRNA SNPs and EOC risk, as well as
having a relatively large study population and a high statistical
power, certain limitations should be taken into consideration. All
the patients were clinically followed up for 12–24 months. The
short follow-up period for certain patients, particularly those of
stage I, mean that analysis of the association between this SNP and
EOC prognosis could not be performed. In addition, the direct
target genes of hsa-miR-196a-2 miRNA were not identified in the
current study; this will require further exploration in future
in vitro and in vivo investigations.
In conclusion, the present study suggested a
potential role for miR-196a-2 rs11614913 in predicting EOC risk,
and indicated the critical use of miRNAs as diagnostic and
prognostic biomarkers in cancer.
Acknowledgements
The present study was supported by a grant from the
National Natural and Science Foundation of China (grant no.
81202070).
References
|
1
|
Ovarian Cancer 2014 Report. World Cancer
Research Fund/American Institute for Cancer Research, Food,
Nutrition, Physical Activity, and the Prevention of Cancer.
https://www.wcrf.org/sites/default/files/Ovarian-Cancer-2014-Report.pdfAccessed.
July 21–2014
|
|
2
|
Ferlay J, Soerjomataram I, Ervik M,
Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D and
Bray F: GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality
Worldwide: IARC CancerBase No. 11 [Internet]. (Lyon, France).
International Agency for Research on Cancer. simpleglobocan.iarc.frAccessed. July 21–2014
|
|
3
|
Sung PL, Chang YH, Chao KC and Chuang CM:
Task Force on Systematic Review and Meta-analysis of Ovarian
Cancer: Global distribution pattern of histological subtypes of
epithelial ovarian cancer: A database analysis and systematic
review. Gynecol Oncol. 133:147–154. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang C, Guo Z, Wu C, Li Y and Kang S: A
polymorphism at the miR-502 binding site in the 3′ untranslated
region of the SET8 gene is associated with the risk of epithelial
ovarian cancer. Cancer Genet. 205:373–376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suh DH, Lee KH, Kim K, Kang S and Kim JW:
Major clinical research advances in gynecologic cancer in 2014.
Gynecol Oncol. 26:156–167. 2015. View Article : Google Scholar
|
|
6
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Munksgaard PS and Blaakaer J: The
association between endometriosis and gynecological cancers and
breast cancer: A review of epidemiological data. Gynecol Oncol.
123:157–163. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gram IT, Lukanova A and Brill I: Cigarette
smoking and risk of histological subtypes of epithelial ovarian
cancer in the EPIC cohort study. Int J Cancer. 130:2204e22102012.
View Article : Google Scholar
|
|
9
|
Chong GO, Jeon HS, Han HS, Son JW, Lee YH,
Hong DG, Lee YS and Cho YL: Differential MicroRNA Expression
Profiles in Primary and Recurrent Epithelial Ovarian Cancer.
Anticancer Res. 35:2611–2617. 2015.PubMed/NCBI
|
|
10
|
Eitan R, Kushnir M, Lithwick-Yanai G,
David MB, Hoshen M, Glezerman M, Hod M, Sabah G, Rosenwald S and
Levavi H: Tumor microRNA expression patterns associated with
resistance to platinum based chemotherapy and survival in ovarian
cancer patients. Gynecol Oncol. 114:253–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan Y, Fan J, Huang L, Ye M, Huang Z, Wang
Y, Li Q and Huang J: Increased expression of microRNA-196a predicts
poor prognosis in human ovarian carcinoma. Int J Clin Exp Pathol.
8:4132–4137. 2015.PubMed/NCBI
|
|
12
|
Yuan C, Liu X, Yan S, Wang C and Kong B:
Analyzing association of the XRCC3 gene polymorphism with ovarian
cancer risk. BioMed Res Int. 2014:6481372014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Diaz-Padilla I, Amir E, Marsh S, Liu G and
Mackay H: Genetic polymorphisms as predictive and prognostic
biomarkers in gynecological cancers: A systematic review. Gynecol
Oncol. 124:354–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vella N, Aiello M, Russo AE, Scalisi A,
Spandidos DA, Toffoli G, Sorio R, Libra M and Stivala F: ‘Genetic
profiling’ and ovarian cancer therapy (Review). Mol Med Rep.
4:771–777. 2011.PubMed/NCBI
|
|
15
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wienholds E and Plasterk RH: MicroRNA
function in animal development. FEBS Lett. 579:5911–5922. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hatfield SD, Shcherbata HR, Fischer KA,
Nakahara K, Carthew RW and Ruohola-Baker H: Stem cell division is
regulated by the microRNA pathway. Nature. 435:974–978. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu Y, Yu CY, Wang JL, Guan J, Chen HY and
Fang JY: MicroRNA sequence polymorphisms and the risk of different
types of cancer. Sci Rep. 4:36482014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Christensen BC, Avissar-Whiting M, Ouellet
LG, Butler RA, Nelson HH, McClean MD, Marsit CJ and Kelsey KT:
Mature microRNA sequence polymorphism in MIR196A2 is associated
with risk and prognosis of head and neck cancer. Clin Cancer Res.
16:3713–3720. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee SJ, Seo JW, Chae YS, Kim JG, Kang BW,
Kim WW, Jung JH, Park HY, Jeong JY and Park JY: Genetic
polymorphism of miR-196a as a prognostic biomarker for early breast
cancer. Anticancer Res. 34:2943–2949. 2014.PubMed/NCBI
|
|
23
|
Hoffman AE, Zheng T, Yi C, Leaderer D,
Weidhaas J, Slack F, Zhang Y, Paranjape T and Zhu Y: microRNA
miR-196a-2 and breast cancer: A genetic and epigenetic association
study and functional analysis. Cancer Res. 69:5970–5977. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang N, Li Y, Zhu LJ, Zhou RM, Jin W, Guo
XQ, Wang CM, Chen ZF and Liu W: A functional polymorphism
rs11614913 in microRNA-196a2 is associated with an increased risk
of colorectal cancer although not with tumor stage and grade.
Biomed Rep. 1:737–742. 2013.PubMed/NCBI
|
|
25
|
Prat J: FIGO Committee on Gynecologic
Oncology: Staging classification for cancer of the ovary, fallopian
tube, and peritoneum. Int J Gynaecol Obstet. 124:1–5. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schroeder A, Mueller O, Stocker S,
Salowsky R, Leiber M, Gassmann M, Lightfoot S, Menzel W, Granzow M
and Ragg T: The RIN: an RNA integrity number for assigning
integrity values to RNA measurements. BMC Mol Biol. 7:32006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen PS, Su JL and Hung MC: Dysregulation
of microRNAs in cancer. J Biomed Sci. 19:902012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fabbri M, Calore F, Paone A, Galli R and
Calin GA: Epigenetic regulation of miRNAs in cancer. Adv Exp Med
Biol. 754:137–148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Salzman DW and Weidhaas JB: SNPing cancer
in the bud: microRNA and microRNA-target site polymorphisms as
diagnostic and prognostic biomarkers in cancer. Pharmacol Ther.
137:55–63. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang D, Meyer L, Chang DW, Lin J, Pu X,
Ye Y, Gu J, Wu X and Lu K: Genetic variants in MicroRNA
biosynthesis pathways and binding sites modify ovarian cancer risk,
survival, and treatment response. Cancer Res. 70:9765–9776. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu Q, Dong Q, He C, Liu W, Sun L, Liu J,
Xing C, Li X, Wang B and Yuan Y: A new polymorphism biomarker
rs629367 associated with increased risk and poor survival of
gastric cancer in Chinese by up-regulated miRNA-let-7a expression.
PLoS One. 9:e952492014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Slaby O, Bienertova-Vasku J, Svoboda M and
Vyzula R: Genetic polymorphisms and microRNAs: New direction in
molecular epidemiology of solid cancer. J Cell Mol Med. 16:8–21.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Srivastava K and Srivastava A:
Comprehensive review of genetic association studies and
meta-analyses on miRNA polymorphisms and cancer risk. PLoS One.
7:e509662012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nicoloso MS, Sun H, Spizzo R, Kim H,
Wickramasinghe P, Shimizu M, Wojcik SE, Ferdin J, Kunej T, Xiao L,
et al: Single-nucleotide polymorphisms inside microRNA target sites
influence tumor susceptibility. Cancer Res. 70:2789–2798. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen J, Ambrosone CB, DiCioccio RA, Odunsi
K, Lele SB and Zhao H: A functional polymorphism in the miR-146a
gene and age of familial breast/ovarian cancer diagnosis.
Carcinogenesis. 29:1963–1966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peng S, Kuang Z, Sheng C, Zhang Y, Xu H
and Cheng Q: Association of microRNA-196a-2 gene polymorphism with
gastric cancer risk in a Chinese population. Dig Dis Sci.
55:2288–2293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L,
Zeng Y, Miao R, Jin G, Ma H, et al: Genetic variants of miRNA
sequences and non-small cell lung cancer survival. J Clin Invest.
118:2600–2608. 2008.PubMed/NCBI
|
|
39
|
Schimanski CC, Frerichs K, Rahman F,
Berger M, Lang H, Galle PR, Moehler M and Gockel I: High miR196a
levels promote the oncogenic phenotype of colorectal cancer cells.
World J Gastroenterol. 15:2089–2096. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhan JF, Chen LH, Chen ZX, Yuan YW, Xie
GZ, Sun AM and Liu Y: A functional variant in microRNA196a2 is
associated with susceptibility of colorectal cancer in a Chinese
population. Arch Med Res. 42:1441482011. View Article : Google Scholar
|
|
41
|
Chen H, Sun LY, Chen LL, Zheng HQ and
Zhang QF: A variant in microRNA196a2 is not associated with
susceptibility to and progression of colorectal cancer in Chinese.
Intern Med J. 42:e115–e119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pu JY, Dong W, Zhang L, Liang WB, Yang Y
and Lv ML: No association between single nucleotide polymorphisms
in pre-mirnas and the risk of gastric cancer in Chinese population.
Iran J Basic Med Sci. 17:128–133. 2014.PubMed/NCBI
|
|
43
|
Zhu R, Liu X, He Z and Li Q: miR-146a and
miR-196a2 polymorphisms in patients with ischemic stroke in the
northern Chinese Han population. Neurochem Res. 39:1709–1716. 2014.
View Article : Google Scholar : PubMed/NCBI
|