Introduction
Gastric cancer is one of the most common types of
cancer in the world (1). The
incidence and mortality of gastric cancer vary geographically, with
the highest rates reported in Eastern Asian countries (2). Although the incidence of gastric cancer
has declined in recent years, it remains a remarkable burden for
public health in China (3). Due to
the lengthy time required to establish a definite diagnosis, a
large number of patients with gastric cancer fail to be diagnosed
prior to the optimal time for surgery, or tend to develop novel
tumors or metastasis during the diagnostic phase (4,5).
Post-surgical chemical therapy for gastric cancer causes resistance
and significant side effects, leading to poor prognosis (6). In recent years, a large number of
studies have focused on biological therapy for gastric cancer,
among which, the interferon (IFN) family has exhibited anti-tumor
abilities (7,8). IFN has been applied clinically as
biological therapy for the treatment of hairy cell leukemia,
chronic myelogenous leukemia, renal carcinoma and melanoma
(9). Human (h)IFN-λ1, also known as
interleukin (IL)-29, is a member of the IFN family, and has
demonstrated anti-tumor effects in the treatment of lung cancer and
colon carcinoma in previous studies (10,11). In
order to examine the potential use of hIFN-λ1 for the treatment of
stomach carcinoma, the present study investigated the anti-tumor
mechanism of hIFN-λ1, and evaluated its effects on the stomach
carcinoma cell line SGC-7901.
Materials and methods
Materials
The stomach carcinoma cell line SGC-7901 and 293A
cells were provided by the Shanghai Institute of Biochemistry and
Cell Biology (Shanghai, China). The recombinant plasmid adenovirus
(pAd)-hIFN-λ1 was generated with primers obtained from Sangon
Biotech Co., Ltd. (Shanghai, China). Cell culture reagents were
acquired from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA), while polymerase chain reaction (PCR) reagents and TRIzol
were purchased from Toyobo Co., Ltd. (Osaka, Japan) and Invitrogen
(Thermo Fisher Scientific, Inc.), respectively. The kits used for
Annexin V (cat no. KGA105) and terminal deoxynucleotidyl
transferase deoxyuridine triphosphate nick end-labeling (TUNEL)
assays (cat no. KGA702) were obtained from KeyGen Biotech Co., Ltd.
(Nanjing, China), and methyl thiazolyl tetrazolium (MTT; cat no.
0793) was obtained from Amresco LLC (Solon, OH, USA). Cyanine
(Cy)3-labeled goat anti-rabbit immunoglobulin (Ig)G (cat no.
KGAB019) was obtained from KeyGen Biotech Co., Ltd., rabbit
anti-human polyclonal hIFN-λ1 antibody (cat no. ab38569) was
obtained from Abcam (Cambridge, UK) and horseradish-peroxidase
(HRP)-conjugated monoclonal mouse anti-GAPDH (cat no. kc5G5) was
obtained from Kangcheng Biology Engineering Co., Ltd. (Shanghai,
China). Furthermore, ECL (cat no. WBLUC0100) was obtained from EMD
Millipore (Billerica, MA, USA) and the BCA kit (cat no. CW0013) was
purchased from CWbio Co., Ltd. (Beijing, China).
Preparation of recombinant
adenovirus
To produce the recombinant virus pAd-hIFN-λ1, a
monolayer of 293A cells cultured in a 6-well plate was transfected
with PacI-linearized plasmid pAd-hIFN-λ1 (4 µg/well; cat no.
FD2204; Thermo Fisher Scientific, Inc.) using Lipofectamine 2000
(Invitrogen, Thermo Fisher Scientific, Inc.). The viruses were
propagated in 293A cells and purified by viral plaque three times.
At 7–14 days post-transfection, when 90% of the cells displayed
cytopathogenic alterations, the cell supernatants were collected by
centrifugation at 10,000 × g for 10 min at room temperature (Jouan
BR4i; Thermo Fisher Scientific, Inc.), and stored at −70°C for
future use. The viral titer (T) was calculated according to the
tissue culture infectious dose (TCID)50. SGC-7901 cells
were infected with recombinant pAd-hIFN-λ1 viruses, with a
multiplicity of infection (MOI) of 12.5, 25, 50, 100, 200, 400 and
800 pfu/cell, respectively. The optimal MOI was determined based on
the morphology presented by the cells at 48 h post-infection.
Indirect immunofluorescence assay
SGC-7901 cells were cultured in a 24-well plate and
infected with pAd-hIFN-λ1. The cells were collected at 48 h
post-infection, fixed with cold 4% paraformaldehyde for 1 h at
−20°C, and washed three times with phosphate-buffered saline (PBS).
The fixed cells were incubated with rabbit anti-human hIFN-λ1
antibody for 1 h at 37°C in a humidified chamber, and then washed
three times with PBS, followed by 1-h staining with Cy3-labeled
goat anti-rabbit IgG at 37°C in a humidified chamber, and three
washes with PBS. All antibodies were diluted to 1:300 and each
washing step was performed for 5 min at room temperature.
Subsequently, the cells were stained with Hoechst 33342.
Photographs of the cells were then captured with a fluorescence
microscope (Leica DFC300 FX, Leica Microsystems (UK) Ltd., Milton
Keynes, UK).
Protein extraction and western blot
analysis
At 48 h post-infection with pAd-hIFN-λ1, pAd-LacZ
and PBS, respectively, 1×106 cells/group were lysed with
100 µl radioimmunoprecipitation assay buffer [50 mM Tris pH 7.4,
150 mM NaCl, 1% TritonX-100, 1% sodium deoxycholate, 0.1% sodium
dodecyl sulfate (SDS), 1 mM dithiothreitol, 1 mM
phenylmethanesulfonyl fluoride, 1 mM protease inhibitors and 1 mM
sodium orthovanadate; CWbio Co., Ltd.]. The cell lysates were then
centrifuged at 12,000 × g for 15 min at 4°C (Jouan BR4i; Thermo
Fisher Scientific, Inc.), and the protein concentration in the
supernatant was determined with a bicinchoninic acid protein assay
kit (Bio-Rad Laboratories, Inc., Berkeley, CA, USA). Subsequently,
the supernatant was mixed with loading buffer at a ratio of 1:1
(v:v), and heated at 100°C for 5 min. Next, 30 µg protein was
loaded onto SDS-polyacrylamide gels and subjected to
electrophoresis using a Mini-PROTEAN Tetra Cell system (Bio-Rad
Laboratories, Inc.). A 10% gel and buffers for electrophoresis were
prepared according to the manufacturer's protocol (Bio-Rad
Laboratories, Inc.). Electrophoresis was performed at a constant
voltage of 120 V for 90 min at room temperature to isolate the
proteins. A Protein Molecular Weight Marker (cat no. 3452; Takara
Bio, Inc., Otsu, Japan). Subsequently, the proteins were
electrotransferred to polyvinylidene difluoride Immobilon membranes
(EMD Millipore) with Mini Trans-Blot Cell (Bio-Rad Laboratories,
Inc.) at a constant current of 350 mA for 90 min at 4°C. The
membranes were then probed with rabbit anti-human hIFN-λ1 antibody,
and washed with Tris-buffered saline pH 7.4 containing 1% Tween-20,
followed by incubation with HRP-labeled goat anti-rabbit IgG (cat.
no. CW0159, Beijing CoWin Biotech Co., Ltd., Beijing, China), and
washed as indicated above. All antibodies were diluted to 1:300.
Immunoblots were detected with enhanced chemiluminescence. Typhoon
9400 was used for scanning western blots (GE Healthcare Life
Sciences, Little Chalfont, UK).
Reverse transcription (RT)-PCR
analysis
SGC-7901 cells were infected with pAd-hIFN-λ1,
pAd-LacZ and PBS, respectively, and total RNA was extracted 24 h
later with TRIzol, following the manufacturers protocol. The RNA
was converted into complementary (c)DNA via RT, using SuperScript
III First-Strand Synthesis System (Invitrogen, Thermo Fisher
Scientific, Inc.). RT was conducted with oligo(dT) primers in 25 µl
reactions, which were incubated at 50°C for 50 min, and terminated
at 85°C for 5 min. The cDNA obtained was stored at −20°C until
further use.
Amplification of hIFN-λ1 cDNA was performed using a
MasterCycler Gradient Thermal Cycler (Eppendorf, Hamburg, Germany)
with primers 5-TATCCAGCCTCAGCCCACAGCA-3 (sense) and
5-ACAGGTTCCCATCGGCCACATA-3 (anti-sense), under the following PCR
conditions: 4 min at 95°C for pre-denaturation, 40 sec at 94°C for
denaturation, 40 sec at 61°C for primer annealing, and 35 sec at
72°C for elongation during 35 cycles of amplification, followed by
10-min incubation at 72°C for final extension. To identify the PCR
products, 5 µl of the reaction was loaded onto 1% agarose gels, and
subjected to electrophoretic analysis. DL2000 DNA Marker (Clontech
Laboratories, Inc., Mountainview, CA, USA) was used as DNA
molecular size marker.
MTT proliferation assay
SGC-7901 cells at logarithmic growth phase were
seeded in 96-well plates at a density of 5×104 cells/ml
in the presence of 200 µl culture medium supplemented with 10%
fetal bovine serum (Gibco, Thermo Fisher Scientific, Inc.), and
incubated for 24 h. Next, the cells were placed in quintuplicate
wells in the presence of 200 µl pAd-hIFN-λ1 (6.3×106
pfu/ml), pAd-LacZ and PBS, respectively. MTT solution (5 mg/ml) was
prepared by dissolving MTT in PBS, and subsequently
filter-sterilizing the solution using a filter paper (EMD
Millipore) of 0.2-µm pores. MTT (20 µl) was then added to each
well, and the plate was incubated for 4 h at 37°C, prior to reading
the absorbance at 490 nm using an iMARK microplate absorbance
reader (Bio-Rad Laboratories, Inc.). Quintuplicate samples/group
were evaluated.
TUNEL assay
A TUNEL assay kit was used to assess the presence of
apoptotic cells in SGC-7901 cells infected with pAd-hIFN-λ1,
pAd-LacZ and PBS, respectively. SGC-7901 cells were fixed in 4%
formaldehyde (pH 7.4) for 48 h at room temperature, rinsed in PBS,
and permeabilized in PBS with 0.1% Triton X-100 and 0.1% sodium
citrate at 4°C for 2 min. Next, the cells were stained following
the protocol provided by the manufacturer, observed under a light
microsocope, and photographed. The number of positive cells/100
cells was counted, and the average ratio of positive cells was
defined as the apoptotic index.
Analysis of apoptosis by flow
cytometry
Apoptosis in SGC-7901 cells infected with
pAd-hIFN-λ1, pAd-LacZ and PBS, respectively, was determined with an
Annexin V-fluorescein isothiocyanate (FITC) staining kit, according
to the manufacturers protocol. Propidium iodide (PI) was used to
differentiate apoptotic cells with membrane integrity (Annexin
V+/PI−) from necrotic cells that had lost
their membrane integrity (Annexin V+/PI+).
The percentage of apoptotic cells was calculated using CellQuest
Pro Software (BD Biosciences, Franklin Lakes, NJ, USA), and the
data were analyzed with WinMDI version 2.9 software (http://en.bio-soft.net/other/WinMDI.html).
Statistical analysis
Data are expressed as the mean ± standard deviation,
unless otherwise stated. Statistically significant differences
between the cells in the pAd-hIFN-λ1 group and the cells in the
control groups were determined by paired t-test or one-way analysis
of variance. SPSS software (version 14.0; SPSS, Inc., Chicago, IL,
USA) was used for all statistical analysis (ANOVA with Fisher's
Least Significant Difference post hoc) and P<0.05 was considered
to indicate a statistical significant difference.
Results
Viral T of adenovirus
The viral T of pAd-hIFN-λ1 was calculated as
follows:
T=101+(1+1+1+1+1+1+1+1–0.5)=108.5/100 µl,
which equates to 109.5 TCID50/ml or
6.3×108 pfu/ml. Thus, there is an exponential
association between T and the MOI. For pAd-LacZ, T=4×108
pfu/ml was observed.
MOI of recombinant virus for infection
of SGC-7901 cells
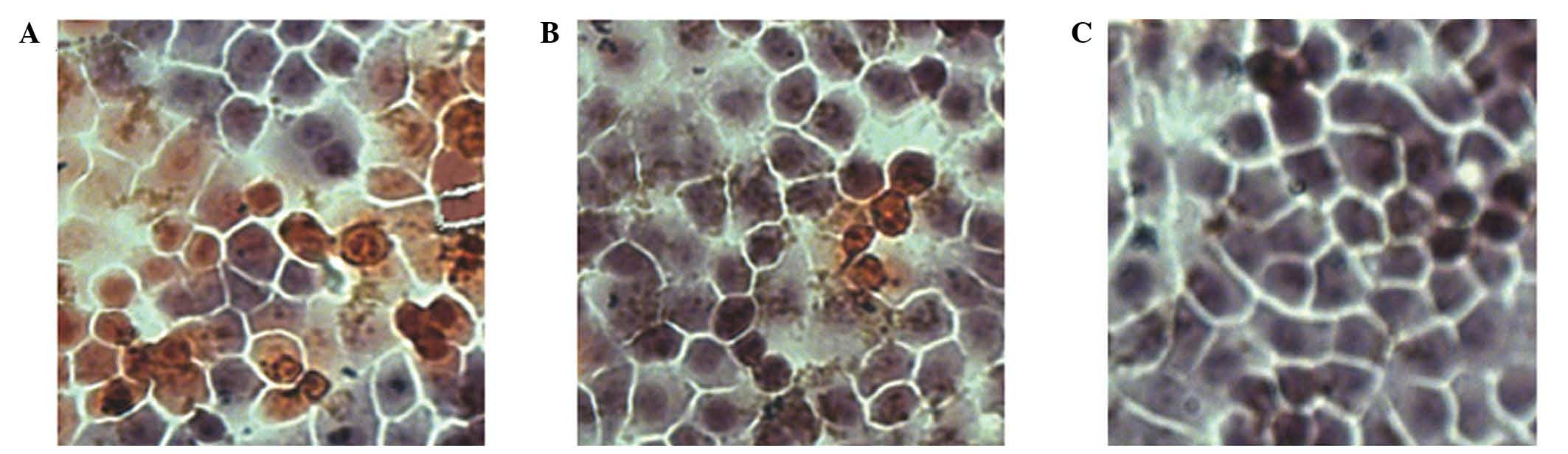
SGC-7901 cells were seeded on 24-well plates at a
density of 5×104 cells/well, and incubated overnight at
37°C with 5% CO2, prior to be infected with pAd-hIFN-λ1
or pAd-LacZ viruses with variable MOI (12.5, 25, 50, 100, 200, 400
and 800 pfu/cell, respectively). At 48 h post-infection, the cells
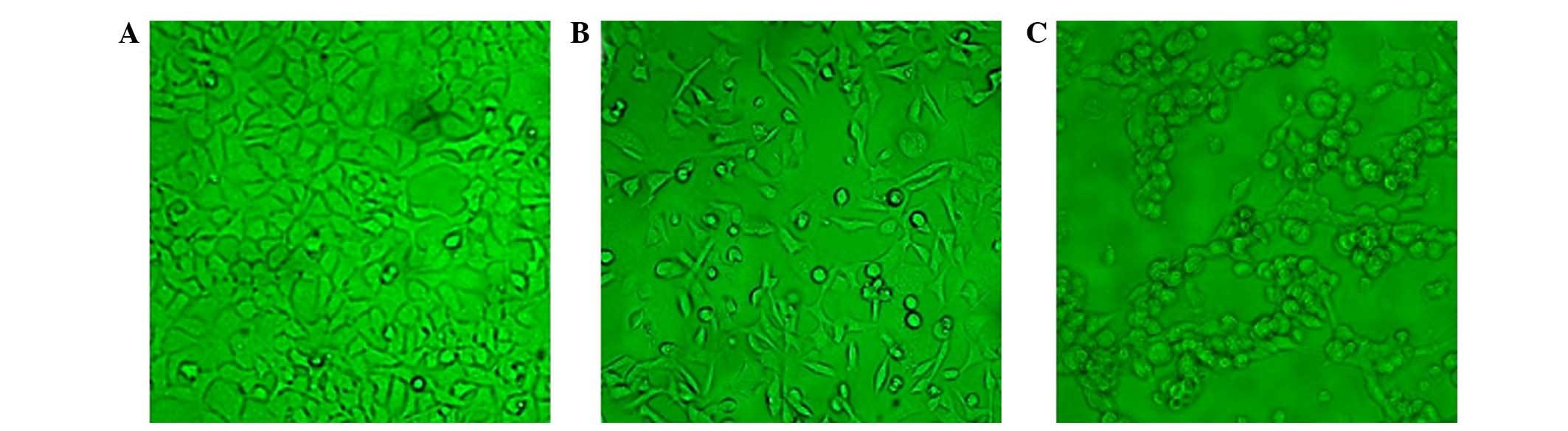
were monitored under the microscope. As presented in Fig. 1, the number of cells infected with
pAd-hIFN-λ1 increased with increasing MOI, until MOI = 200, while
at higher MOI values (MOI = 400 and 800), the cells became necrotic
and their size reduced, indicating that the optimal MOI was
200.
Messenger (m)RNA and protein
expression levels of hIFN-λ1 in SGC-7901 cells
IFN-λs have been recently identified as cytokines
whose target cells remain to be elucidated (12,13).
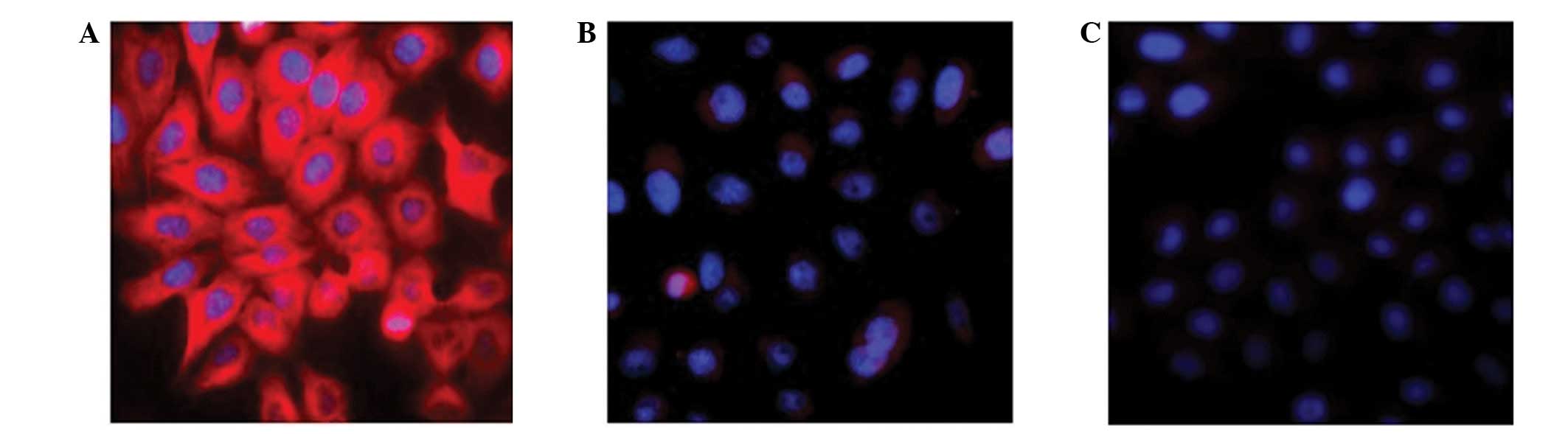
Immunofluorescence was used to detect the expression of hIFN-λ1 in
SGC-7901 cells infected with pAd-hIFN-λ1, pAd-LacZ and PBS,
respectively. The results are presented in Fig. 2. The pAd-LacZ and PBS control groups
did not exhibit any red fluorescence, while the pAd-hIFN-λ1 group
markedly displayed red fluorescence, indicating expression of
hIFN-λ1 protein in these cells.
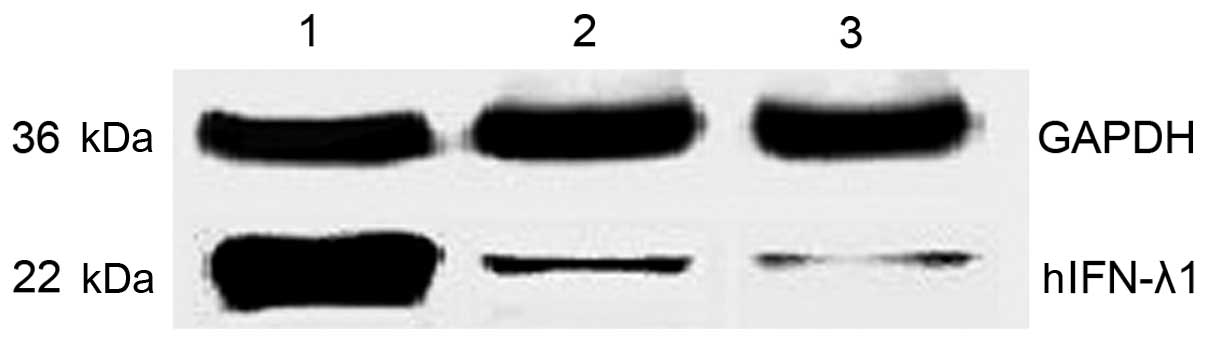
To quantify the protein expression levels of hIFN-λ1
in the pAd-hIFN-λ1, pAd-LacZ and PBS groups, western blot assay was
performed. The results are expressed as relative to the protein
levels of glyceraldehyde 3-phosphate dehydrogenase measured in
these cells. As presented in Fig 3,
the protein expression levels of hIFN-λ1 were higher in the
pAd-hIFN-λ1 group, compared with the control groups, suggesting
that hIFN-λ1 was overexpressed in these cells.
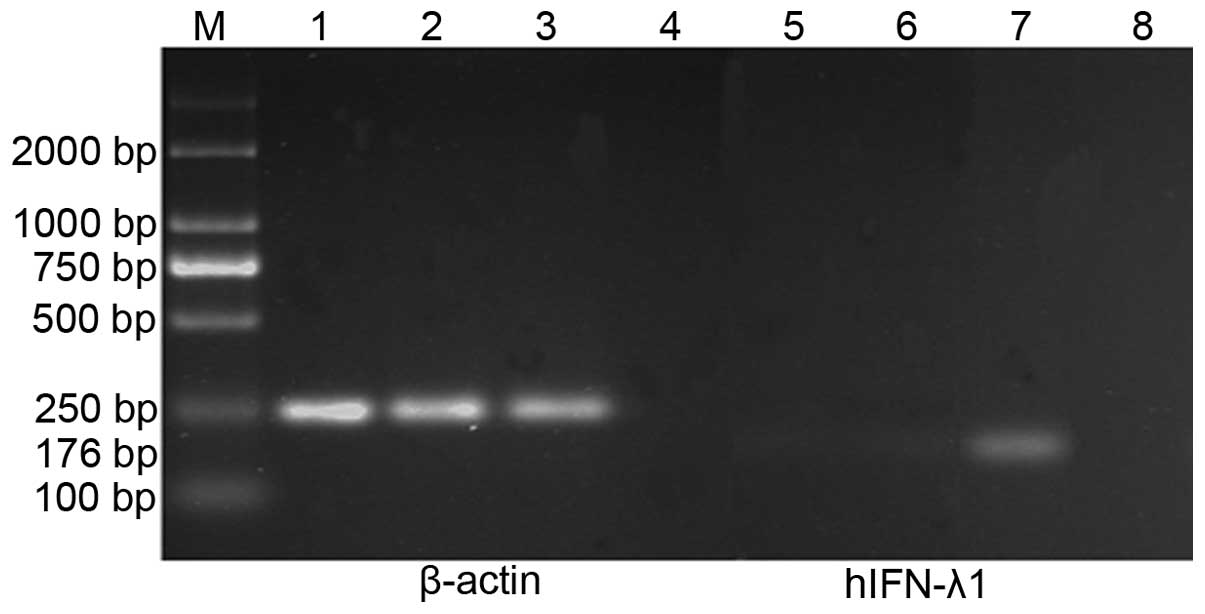
In addition, RT-PCR was used to evaluate the mRNA
expression levels of hIFN-λ1 in each group. mRNA was isolated from
SGC-7901 cells infected with pAd-hIFN-λ1, pAd-LacZ and PBS,
respectively, and the levels of hIFN-λ1 transcript present in these
mRNA samples were determined by RT-PCR. The results demonstrated
the presence of high mRNA expression levels of hIFN-λ1 in the
pAd-hIFN-λ1 group, but not in the controls (Fig. 4).
Apoptosis induced by infection with
pAd-hIFN-λ1
To assess whether the hIFN-λ1-induced
anti-proliferative effects observed in SGC-7901 cells infected with
pAd-hIFN-λ1 may be due to the induction of apoptosis, MTT assay was
performed. The results obtained confirmed the initial hypothesis.
Thus, pAd-hIFN-λ1 significantly inhibited cell proliferation in
SGC-7901 cells (43.28±1.65% inhibition vs. 3.12±2.91% inhibition
for the pAd-LacZ control) (data not shown).
These results were in agreement with the findings of
TUNEL assay. As presented in Fig. 5,
the pAd-hIFN-λ1 group exhibited a higher number of apoptotic cells
that the pAd-LacZ control group.
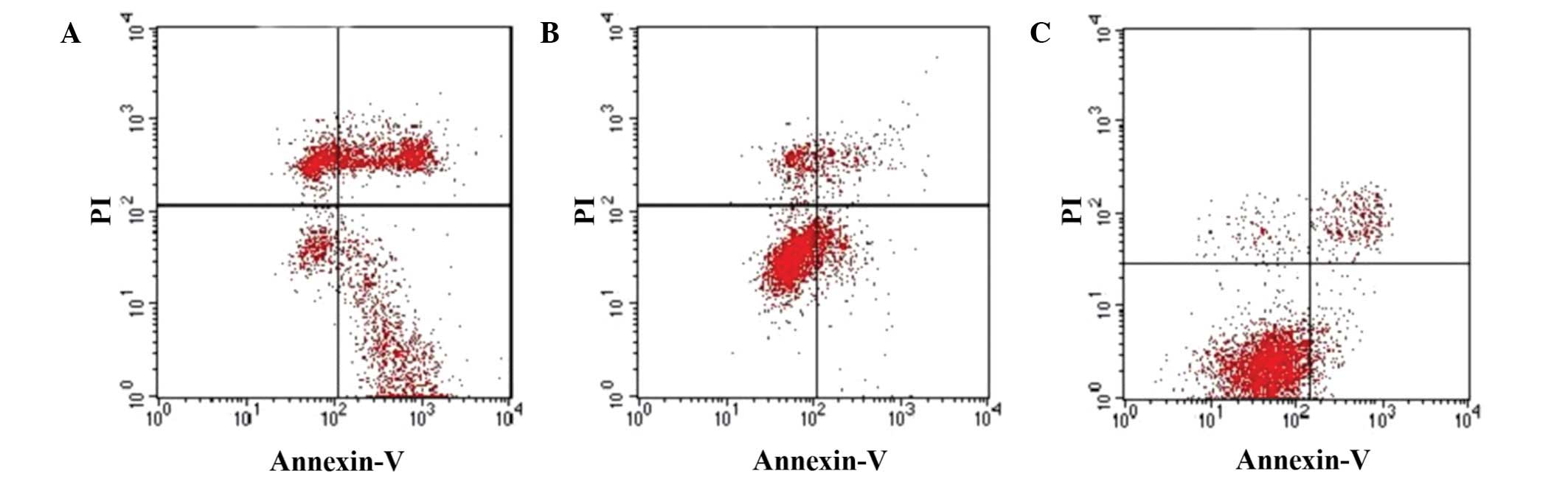
These observations were further confirmed by flow
cytometry using Annexin V and PI staining. As depicted in Fig. 6, the apoptotic index significantly
increased in SGC-7901 cells infected with pAd-hIFN-λ1, whereas a
slight increase in the apoptotic index was observed in
pAd-hIFN-λ1-infected cells, compared with the PBS control.
Discussion
Members of the type III IFN-λ family include IFN-λ1,
2 and 3, also termed IL-29, −28A and −28B, respectively (14). IFN-λs exhibit similar effects to type
I IFNs, which are involved in signal transduction pathways,
anti-virus, anti-proliferation and immunological regulation
(15–17). Gene therapy is a highly effective and
specific type of therapy that enables the translation of a gene of
interest in the corresponding target cells via a vector (18). The success of gene therapy mainly
depends on the selection of an adequate vector (19). Since viruses are able to translate
independently their genetic material inside the host cell, and
their DNA does not integrate into the host cell chromosome,
virus-derived vectors such as adenoviruses are considered safer
than other type of vectors that require integration of their
genetic material in the DNA of the host (20). Therefore, virus-derived vectors, and
in particular adenoviruses, have become the most widely used type
of vector in gene therapy (21).
In the present study, pAd-hIFN-λ1 was infected into
the human gastric cell line SGC-7901, and the genetic material
carried by the vector was efficiently transcribed inside these
cells, as demonstrated by the high mRNA expression levels of
hIFN-λ1 detected by RT-PCR. In addition, immunofluorescence and
western blot analyses revealed high protein expression levels of
hIFN-λ1 in pAd-hIFN-λ1-infected SGC-7901 cells, indicating that
hIFN-λ1 was also efficiently translated inside these tumor cells.
Previous studies have demonstrated that hIFN-λ1, a recently
identified member of the IFN-λs family, exhibits anti-tumor effects
in lung carcinoma (11), colon cancer
(10) and esophageal cancer (22). The results of the MTT assay conducted
in the present study confirmed that pAd-hIFN-λ1 is effective in
inhibiting the proliferation of SGC-7901 cells, which is in
agreement with the anti-tumor effects of hIFN-λ1 reported in
previous studies. Therefore, hIFN-λ1 may be a potential target for
biological therapy of stomach carcinoma. hIFN-λ1 may exert its
anti-tumor effects by inhibiting tumor replication (23), inducing cell apoptosis (24,25),
inhibiting cell proliferation (16,26),
regulating the immune system in order to kill tumor cells (27), targeting tumor matrix cells, and
promoting anti-angiogenesis (28).
In the present study, the results of TUNEL assay and
Annexin V-FITC/PI flow cytometry revealed that the apoptosis rate
was higher in the pAd-hIFN-λ1 group than in the pAd-LacZ and PBS
control groups. Previous studies using IFN-λs to regulate the
apoptosis of colon carcinoma cells have demonstrated that IFN-λs
are able to arrest cells in the G1/G0 phase
of the cell cycle, leading to the extracellular translocation of
phosphatidylserine from the inner side of the cell membrane, DNA
damage and activation of caspase-3, −8 and −9, which eventually
results in apoptosis (28,29). Therefore, one of the possible
mechanisms of inhibition of cell proliferation in gastric carcinoma
cells is the induction of apoptosis via phosphatidylserine
(30).
In conclusion, the results of the present study
confirm that pAd-hIFN-λ1 is capable of inhibiting proliferation and
inducing apoptosis in stomach cancer cells, which provides evidence
for future studies on the use of pAd-hIFN-λ1 as biological therapy
for the treatment of stomach cancer. Compared with type I IFNs and
cytotoxic drugs, pAd-hIFN-λ1-based therapy does not appear to
produce any side effects. However, whether the combination of
pAd-hIFN-λ1 with a reduced dosage of cytotoxic drugs may enhance
its anti-tumor effects or reduce the side effects caused by the
cytotoxic drugs requires further investigation.
Acknowledgements
This study was supported by the Natural Science
Foundation of Jiangsu Province (grant no. BK20151333).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pourhoseingholi MA, Vahedi M and
Baghestani AR: Burdenof gastrointestinal cancer in Asia; an
overview. Gastroenterol Hepatol Bed Bench. 8:19–27. 2015.PubMed/NCBI
|
|
3
|
Tänzer M, Liebl M and Quante M: Molecular
biomarkers in esophageal, gastric, and colorectal adenocarcinoma.
Pharmacol Ther. 140:133–147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liang H and Kim YH: Identifying molecular
drivers of gastric cancer through next-generation sequencing.
Cancer Lett. 340:241–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
González CA and Agudo A: Carcinogenesis,
prevention and early detection of gastric cancer: Where we are and
where we should go. Int J Cancer. 130:745–753. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yared JA and Tkaczuk KH: Update on taxane
development: New analogs and new formulations. Drug Des Devel Ther.
6:371–384. 2012.PubMed/NCBI
|
|
7
|
Bu XF, Zhang J, Jia LJ, Liang B, Zhang J,
Liu Y and Yan YL: Effect of human interferon-λ1 recombinant
adenovirus on a gastric cancer orthotopic transplantation model.
Exp Ther Med. 8:1115–1122. 2014.PubMed/NCBI
|
|
8
|
Gao Z, Zhu M, Wu Y, Gao P, Qin Z and Wang
H: Interferon-λ1 induces G1 phase cell cycle arrest and apoptosis
in gastric carcinoma cells in vitro. Oncol Rep. 32:199–204.
2014.PubMed/NCBI
|
|
9
|
Ferrantini M, Capone I and Belardelli F:
Interferon-α and cancer: Mechanisms of action and new perspectives
of clinical use. Biochimie. 89:884–893. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hui X, Chen H, Zhang S, Ma X, Wang X and
Huang B: Antitumor activities of recombinant human interferon
(IFN)-λ1 in vitro and in xenograft models in vivo for
colon cancer. Cancer Lett. 311:141–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan Y, Zhang J, Liu Y, Zhu T, Yuan L, Ge
Y, Ding H and Bu X: Inhibition of lung adenocarcinoma transfected
with interleukin 28A recombinant adenovirus (Ad-mIFN-λ2) in
vivo. Cancer Biother Radiopharm. 28:124–130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lopušná K, Režuchová I, Betáková T, et al:
Interferons lambda, new cytokines with antiviral activity. Acta
Virol. 57:171–179. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Steen HC and Gamero AM: Interferon-lambda
as a potential therapeutic agent in cancer treatment. J Interferon
Cytokine Res. 30:597–602. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Donnelly RP and Kotenko SV:
Interferon-lambda: A new addition to an old family. J Interferon
Cytokine Res. 30:555–564. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meager A, Visvalingam K, Dilger P, Bryan D
and Wadhwa M: Biological activity of interleukins −28 and −29:
Comparison with type I interferons. Cytokine. 31:109–118. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dumoutier L, Tounsi A, Michiels T,
Sommereyns C, Kotenko SV and Renauld JC: Role of the interleukin
(IL)-28 receptor tyrosine residues for antiviral and
antiproliferative activity of IL-29/interferon-λ1: Similarities
with type I interferon signaling. J Biol Chem. 279:32269–32274.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lasfar A, Lewis-Antes A, Smirnov SV,
Anantha S, Abushahba W, Tian B, Reuhl K, Dickensheets H, Sheikh F,
Donnelly RP, et al: Characterization of the mouse IFN-lambdas
ligand-receptor system: IFN-λs exhibit antitumor activity against
B16 melanoma. Cancer Res. 66:4468–4477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Khalighinejad N, Hariri H, Behnamfar O,
Yousefi A and Momeni A: Adenoviral gene therapy in gastric cancer:
A review. World J Gastroenterol. 14:180–184. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rincon MY, VandenDriessche T and Chuah MK:
Gene therapy for cardiovascular disease: Advances in vector
development, targeting, and delivery for clinical translation.
Cardiovasc Res. 108:4–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang I and Huang I: Adenovirus technology
for gene manipulation and functional studies. Drug Discov Today.
5:10–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Büning H, Huber A, Zhang L, Meumann N and
Hacker U: Engineering the AAV capsid to optimize
vector-host-interactions. Curr Opin Pharmacol. 24:94–104. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Q, Kawamura K, Okamoto S, Fujie H,
Numasaki M, Namba M, Nagata M, Shimada H, Kobayashi H and Tagawa M:
Adenoviruses-mediated transduction of human oesophageal carcinoma
cells with the interferon-λ genes produced anti-tumour effects. Br
J Cancer. 105:1302–1312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kelly C, Klenerman P and Barnes E:
Interferon-lambdas: The next cytokine storm. Gut. 60:1284–1293.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tagawa M, Kawamura K, Li Q, Tada Y,
Hiroshima K and Shimada H: A possible anticancer agent, type III
interferon, activates cell death pathways and produces antitumor
effects. Clin Dev Immunol. 2011:4790132011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sato A, Ohtsuki M, Hata M, Kobayashi E and
Murakami T: Antitumor activity of IFN-λ in murine tumor models. J
Immunol. 176:7686–7694. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maher SG, Sheikh F, Scarzello AJ,
Romero-Weaver AL, Baker DP, Donnelly RP and Gamero AM: IFNalpha and
IFNlambda differ in their antiproliferative effects and duration of
JAK/STAT signaling activity. Cancer Biol Ther. 7:1109–1115. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li M, Liu X, Zhou Y and Su SB:
Interferon-lambdas: The modulators of antivirus, antitumor, and
immune responses. J Leukoc Biol. 86:23–32. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li W, Lewis-Antes A, Huang J, Balan M and
Kotenko SV: Regulation of apoptosis by type III interferons. Cell
Prolif. 41:960–979. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Q, Kawamura K, Tada Y, Shimada H,
Hiroshima K and Tagawa M: Novel type III interferons produce
anti-tumor effects through multiple functions. Front Biosci
(Landmark Ed). 18:909–918. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woehlecke H, Pohl A, Alder-Baerens N, Lage
H and Herrmann A: Enhanced exposure of phosphatidylserine in human
gastric carcinoma cells overexpressing the half-size ABC
transporter BCRP (ABCG2). Biochem J. 376:489–495. 2003. View Article : Google Scholar : PubMed/NCBI
|