Introduction
Molecules in the breast tumor microenvironment may
impact the progression of breast tumor cells throughout all stages
of the metastatic process (1).
Adiponectin (AdipoQ) is a 244-amino acid protein, and the most
abundant adipokine among the host of autocrine, endocrine and
paracrine-acting adipokines secreted by adipocytes (2). Adipocytes are key constituents of the
mammary stroma (3). Therefore, it is
not surprising that the presence of AdipoQ has been previously
reported in breast cancer and adjacent normal breast epithelial
tissue (4–6), in addition to its well-characterized
presence in circulating plasma, where its levels range from 2 to 30
µg/ml (2). Thus, we hypothesize that
AdipoQ may act directly on tumor cells and potentially contribute
to the microenvironmental regulation of the early steps of
metastasis.
AdipoQ is synthesized as a ~30 kDa monomer,
consisting of an amino-terminal sequence, a variable domain, a
collagen-like domain and a carboxyl-terminal globular domain
(2). This full-length monomer (fAd)
assembles into higher-order structures such as trimers, hexamers
and multimers of low, intermediate and high molecular weight
(7,8).
AdipoQ additionally exists as a cleaved isoform named globular
adiponectin (gAd), whereby fAd is truncated into a fragment
primarily containing its globular domain (9,10). This
cleavage event is initiated by leukocyte elastase (10), a serine protease secreted by activated
monocytes and neutrophils alongside breast cancer cells (11), which has been associated with poor
prognosis in patients with breast cancer (11–13).
A substantial body of clinical and experimental
evidence suggests that AdipoQ possesses an inhibitory function in
the development (14–32) and progression (22,24,32–35)
of breast tumors. Notably, the majority of clinical evidence
supporting antitumor roles for AdipoQ in breast cancer is based on
circulating levels of serum AdipoQ, which often are not
representative of tissue concentrations (36,37).
Furthermore, previous tissue-based studies have suggested an
association between high expression levels of AdipoQ and cancer
(4,5).
In addition, despite numerous observations suggesting that fAd and
gAd diverge in biological function (9,38–40), the majority of studies on the role of
AdipoQ in breast cancer have focused on fAd or total AdipoQ,
without considering gAd (35). In
previous studies, gAd has been observed to elicit pro-metastatic
responses, including reduced hypoxia-associated apoptosis (41) and increased cell proliferation,
migration, angiogenesis, activation of matrix metalloproteinases,
secretion of proinflammatory cytokines and production of reactive
oxygen species (ROS) (38–40).
Therefore, the present study aimed to assess the
influence of gAd versus fAd on the metastatic potential of breast
cancer cells. The results revealed a more invasive cell phenotype
and an increase in indicators of autophagic induction upon addition
of gAd, but not fAd. In addition, the gAd-induced increase in
invasion was observed to be partly dependent on autophagic
function. An enhanced understanding of novel isoform-specific
functions for AdipoQ in the breast tumor microenvironment may aid
the development of novel therapeutic approaches for the treatment
of metastatic breast cancer.
Materials and methods
Cells and cell culture
The metastatic human breast carcinoma cell line
MDA-MB-231, previously described by Hurst et al (42), was acquired from the American Type
Culture Collection (Manassas, VA, USA). The cell line was cultured
in a 1:1 (v/v) mixture of Dulbecco's modified Eagle's medium and
Ham's F-12 Nutrient Mixture (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 5% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc.), 2 mM L-glutamine (Thermo Fisher
Scientific, Inc.) and 0.02 mM non-essential amino acids (Thermo
Fisher Scientific, Inc.). Cells were maintained at 37°C with 5%
CO2 in a humidified atmosphere and were regularly tested
for Mycoplasma spp. contamination with PlasmoTest™ Reagent
Kit (InvivoGen, San Diego, CA, USA), according to the
manufacturer's protocol, and identified to be negative.
Transwell migration and invasion
assays
Cell migration assays were performed with a
Transwell chamber comprising 24-well inserts with membranes of 8-µm
diameter pores (Corning Life Sciences, Manassas, VA, USA). Cells
were seeded (1×105 cells/insert) in serum-free medium
(SFM) and treated with 0.5 µg/ml human recombinant fAd
(SouthernBiotech, Birmingham, AL, USA), 0.5 µg/ml human recombinant
gAd (R&D Systems, Inc., Minneapolis, MN, USA) or 100 nM
rapamycin (Sigma-Aldrich, St. Louis, MO, USA). The concentration of
gAd used to treat the cells was selected based on a previous study
reporting the relatively low levels of gAd compared with total
AdipoQ (9). To ensure equal
comparisons, the same concentration of fAd was used in the present
study. Complete culture medium was added to the lower wells and
employed as chemoattractant. Each treatment condition was evaluated
in triplicate. Following incubation for 19 h, non-migrated cells
remaining on the upper surface of the insert membranes were removed
using sterile cotton swabs (Thermo Fisher Scientific, Inc.), and
the membranes were next washed with deionized water, fixed in 100%
methanol (Thermo Fisher Scientific, Inc.) for 20 min, washed and
stained with crystal violet (Thermo Fisher Scientific, Inc.) for 18
min. The inserts were air-dried overnight. Images of migrated cells
(8 images/insert) were captured with ECLIPSE TE2000-U microscope
(Nikon Corporation, Tokyo, Japan), and the average number of
migrated cells per field was compared. Similar procedures were used
for invasion assays with the following exceptions: Membranes were
coated with a layer of Matrigel™ Basement Membrane Matrix (BD
Biosciences, San Jose, CA, USA), the seeding density was
5×105 cells/insert and the incubation period was 22
h.
Three-dimensional (3D) cell morphology
studies
3D cell culture assays were performed in 24-well
plates (Thermo Fisher Scientific, Inc.) with 400 µl/well of
Matrigel™ Matrix Growth Factor Reduced (BD Biosciences). Cells were
suspended in complete medium supplemented with 2% Matrigel™, plated
at a density of 4×103 cells/well and incubated at 37°C
for 9 days. A fresh layer of complete medium supplemented with
Matrigel™ was added following 3 days of incubation, and 5 days
later, the medium was replaced with SFM containing the following
treatments: i) 0.5 µg/ml human recombinant fAd; ii) 0.5 µg/ml human
recombinant gAd; or iii) 100 nM rapamycin, individually or with 50
µM chloroquine (Sigma-Aldrich). Each condition was evaluated in
triplicate. Images were captured at 9 days post-incubation using
ECLIPSE TE2000-U microscope.
Proliferation assay
Proliferation assays were performed as previously
described (43). Briefly, cells were
cultured in SFM, seeded onto a 96-well tissue culture plate
(1×103 cells/well; Thermo Fisher Scientific, Inc.) and
treated with 0.5 µg/ml human recombinant fAd, 0.5 µg/ml human
recombinant gAd or 100 nM rapamycin. Cell viability was measured at
1, 3, 5 and 7 days post-incubation with alamarBlue® (Thermo Fisher
Scientific, Inc.). Fluorescence intensity at 570/580 nm
excitation/emission was determined via F-7000 Fluorescence
Spectrophotometer (Hitachi, Ltd., Tokyo, Japan).
Immunoblot analysis
Cells grown in two-dimensional (2D) cultures were
treated for 3 and 6 h under identical conditions to those used in
3D cell morphology studies. Whole-cell lysates were collected with
1X radioimmunoprecipitation assay lysis buffer (EMD Millipore,
Billerica, MA, USA) containing 1X Halt™ Protease and Phosphatase
Inhibitor Single-Use Cocktail (Thermo Fisher Scientific, Inc.).
Proteins were separated by sodium dodecyl sulfate-polyacrylamide
gel electrophoresis (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at 100 V for 2 h and transferred to nitrocellulose membranes
(Bio-Rad Laboratories, Inc.). Membranes were next blocked for 1 h
at room temperature using 5% skimmed milk (Bio-Rad Laboratories,
Inc.) dissolved in Tris-buffered saline (TBS) supplemented with
0.05% Tween 20 (TBST; Bio-Rad Laboratories, Inc.), and incubated
overnight at 4°C with the corresponding primary antibody. Membranes
were then washed with TBST four times, incubated with the
corresponding monoclonal donkey anti-rabbit (cat. no. NA934V) or
sheep anti-mouse (cat. no. NA931V) IgG secondary antibody
(1:10,000; GE Healthcare Life Sciences, Chalfont, UK) for 1 h at
room temperature and washed with TBST four times. Blots were
developed using ECL Western Blotting Substrate (Thermo Fisher
Scientific, Inc.) and Supersignal West Dura Extended Duration
Substrate (Thermo Fisher Scientific, Inc.), and the results were
quantified with Image Studio™ Software version 4.0 (LI-COR
Biotechnology, Lincoln, NE, USA). The following primary antibodies
were employed: Rabbit anti-human polyclonal
anti-microtubule-associated protein 1 light chain 3 beta (LC3B)
(1:3,000; cat. no. ab51520; Abcam, Cambridge, MA, USA), rabbit
anti-human monoclonal autophagy related protein 7 (ATG7) (1:5,000;
cat. no. 04–1055; EMD Millipore); and mouse anti-human monoclonal
β-actin (1:10,000; cat. no. A5441; Sigma-Aldrich).
Immunofluorescence studies
Glass coverslips pretreated with 0.01% poly-L-lysine
(Thermo Fisher Scientific, Inc.) were placed in 6-well culture
plates, whereby cells were seeded and grown to 80–90% confluence.
Next, cells were incubated with SFM containing 0.5 µg/ml human
recombinant gAd or 100 nM rapamycin, individually or with the
addition of 50 µM chloroquine. Each treatment condition was
evaluated in triplicate. At 3 and 6 h post-incubation, cells were
washed 4 times for 3 min in phosphate-buffered saline (PBS; (Thermo
Fisher Scientific, Inc.), fixed in 3% formaldehyde (Thermo Fisher
Scientific, Inc.) dissolved in PBS for 45 min at room temperature,
permeabilized with 0.5% Triton X-100 (Sigma-Aldrich) for 3 min at
room temperature, blocked for 1 h with PBS containing 1% bovine
serum albumin (BSA; Thermo Fisher Scientific, Inc.) and incubated
overnight at 4°C with rabbit anti-human polyclonal LC3B antibody
(1:2,000; cat. no. ab51520; Abcam) dissolved in PBS containing 1%
BSA. Cells were subsequently incubated with goat anti-rabbit IgG
fluorescein isothiocyanate-labelled secondary antibody (1:2,000;
cat. no. ab6717; Abcam) for 1 h at room temperature. The coverslips
were then mounted onto glass slides (Thermo Fisher Scientific,
Inc.) using VECTASHIELD® Antifade Mounting Medium with DAPI
(catalogue no. H-1200; Vector Laboratories, Inc., Burlingame, CA,
USA). Immunofluorescent images were captured with ECLIPSE TE2000-U
fluorescence microscope.
RNA interference
Gene silencing assays were performed using small
interfering (si)RNA targeting three non-overlapping sequences of
ATG7 (MISSION® siRNAs catalogue nos. SASI_Hs01_00077648,
SASI_Hs01_00077650 and SASI_Hs01_00077652, which were termed ATG7
siRNA1, 2 and 3, respectively; Sigma-Aldrich). MISSION® siRNA
Universal Negative Control #1 (Sigma-Aldrich) was selected as the
non-targeting siRNA control, and referred to as scramble siRNA. To
achieve transfection, Lipofectamine® 3000 (Thermo Fisher
Scientific, Inc.) was utilized according to the manufacturer's
protocol.
Statistical analysis
Between-group differences were assessed by unpaired
Student's t-test, and P<0.05 was considered to indicate a
statistically significant difference. Statistical analysis was
performed with SigmaStat® software version 3.5 (Systat Software,
Inc., Chicago, IL, USA).
Results
gAd increases the metastatic potential
of breast cancer cells
The ability to migrate and invade through breast
tissue stroma is an essential property of metastatic tumor cells
(44). In order to investigate how
different AdipoQ isoforms modulate this capacity, the effect of gAd
and fAd on the 3D growth in Matrigel™ of the human metastatic
breast carcinoma cell line 231 was evaluated. In the absence of
serum, parental 231 cells grew with a relatively non-invasive
grape-like morphology (Fig. 1A). By
contrast, 231 cells treated with gAd developed spicules typical of
a more invasive cell phenotype, whereas fAd-treated cells
maintained a non-invasive grape-like structure similar to that of
the untreated controls (Fig. 1A).
Since none of the AdipoQ isoforms tested altered the proliferation
ability of 231 cells in 2D culture (Fig.
1B), Transwell assays with and without Matrigel™ were
subsequently performed to quantify the invasion and migration
abilities of 231 cells. Treatment with gAd promoted migration by
221% (Fig. 1C and D; P<0.001) and
invasion by 91% (Fig. 1E and F;
P<0.001), compared with untreated cells. No significant
alterations were noted following treatment with fAd. These results
suggested that the different AdipoQ isoforms performed different
functions. Thus, gAd appeared to promote the metastatic potential
of 231 cells, contrarily to fAd.
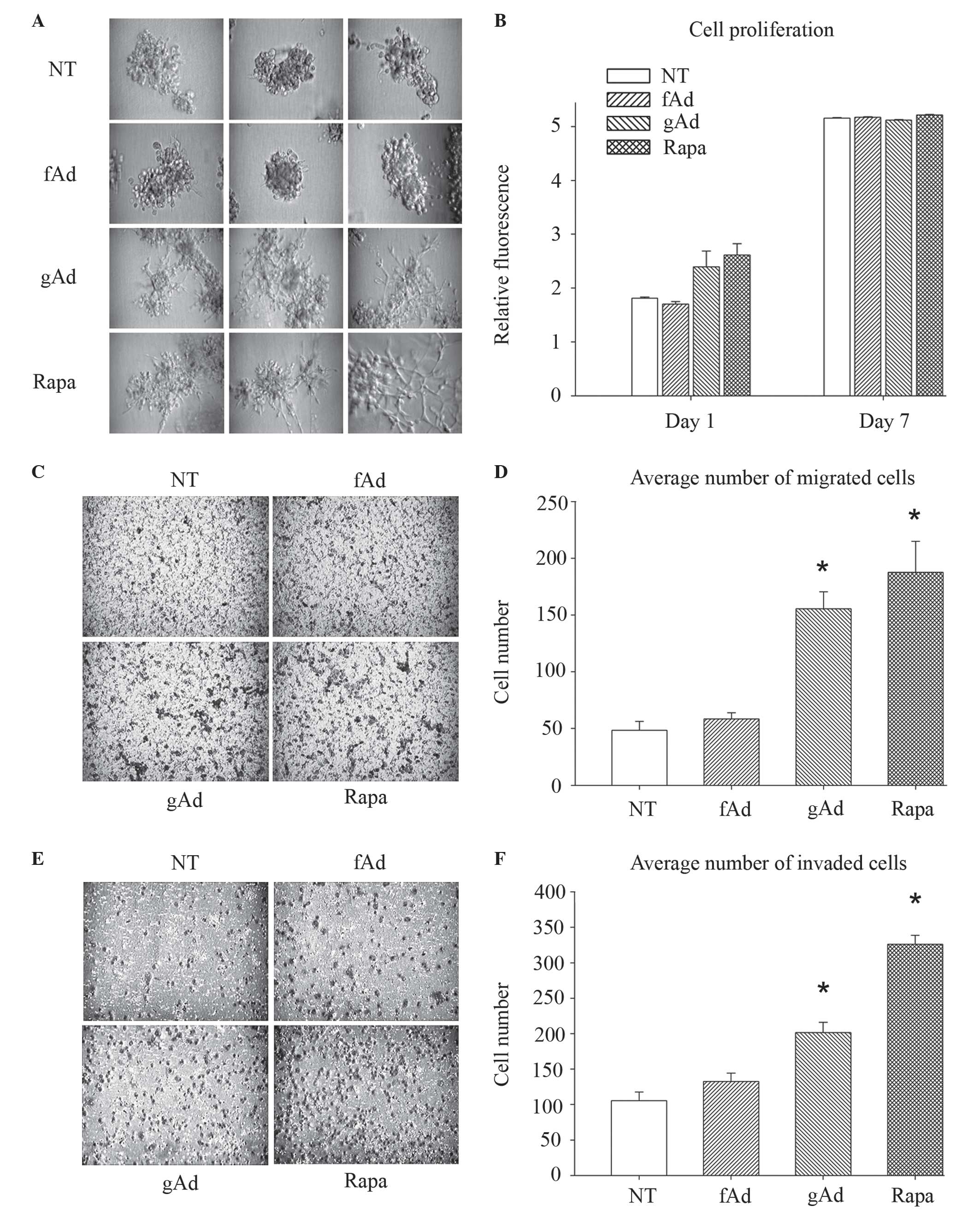 | Figure 1.gAd increases the metastatic
potential of breast cancer cells. (A) MDA-MB-231 cells were
cultured in SFM, seeded onto Matrigel™-coated cell culture plates
and treated with 0.5 µg/ml human recombinant fAd, 0.5 µg/ml human
recombinant gAd or 100 nM rapa. Untreated cells were used as
negative controls. All conditions were evaluated in triplicate.
Representative images of each replicate were obtained 9 days
subsequent to seeding. (B) Cell proliferation was assessed with
alamarBlue®, which revealed that neither fAd, gAd nor rapa
significantly altered cell growth over a 7-day period, compared
with NT control cells. Each bar represents the mean fluorescence
intensity obtained for each of the conditions, which were evaluated
in quadruplicate. (C-F) Cells were cultured in SFM, seeded onto the
top chamber of Transwell (C and D) migration and (E and F) invasion
plates, and treated with fAd (0.5 µg/ml), gAd (0.5 µg/ml) or rapa
(100 nM). Migration and invasion towards complete serum-containing
medium were measured at 19 and 22 h post-incubation, respectively.
gAd and rapa significantly increased cell migration and invasion,
compared with NT controls. Data are represented as the
mean±standard error from assays performed in triplicate. *P≤0.001.
SFM, serum-free medium; fAd, full-length adiponectin; gAd, globular
adiponectin; rapa, rapamycin; NT, no treatment. |
Signaling pathways support the association between
gAd and the induction of autophagy, a cellular stress response
broadly considered to suppress tumorigenesis but promote the
progression of established tumor cells (45). The authors of the present study have
previously proposed a model in which autophagy is a plausible
mechanism for gAd-stimulated breast cancer metastasis (35). In agreement with that model, the
present study observed that gAd altered the metastatic potential of
231 cells in a manner akin to rapamycin, an established inducer of
autophagy (46). Similar to gAd,
rapamycin significantly increased cell migration (286%; P=0.001;
Fig. 1C and D) and invasion (210%;
P<0.001; Fig. 1E and F), and
promoted the development of extended spikes in 3D cultures of 231
cells (Fig. 1A).
gAd promotes autophagic induction in
breast cancer cells
Since gAd and rapamycin elicited similar invasive
phenotypes in breast cancer cells, the association between
different AdipoQ isoforms and autophagic induction was further
investigated using rapamycin as positive control. Pharmacological
inhibition of autophagy with chloroquine markedly reduced the gAd
and rapamycin-induced increase in invasive morphology, whereas no
clear alteration was observed in fAd-treated cells (Fig. 2A). Subsequent biochemical assays of
autophagic induction corroborated these findings. LC3B is a widely
used marker of autophagic induction, and the conversion of LC3B-I
to LC3B-II, along with an increase in LC3B puncta, are considered
to be indicative of autophagosome formation (46). To ensure an accurate comparison
between treatments, any cellular alterations that occurred in the
presence of chloroquine were analyzed (46). Immunofluorescence analysis revealed
that gAd and rapamycin similarly increased the number of LC3B
puncta (Fig. 2B) Additionally,
treatment with gAd and rapamycin increased the conversion of LC3B-I
to LC3B-II in 231 cells by 110 and 220%, respectively, compared
with no treatment (Fig. 2C and D). In
contrast, the levels of LC3B-II remained almost constant upon
treatment with fAd. Therefore, the results of the present study
suggested that gAd and fAd exerted disparate effects on autophagic
induction.
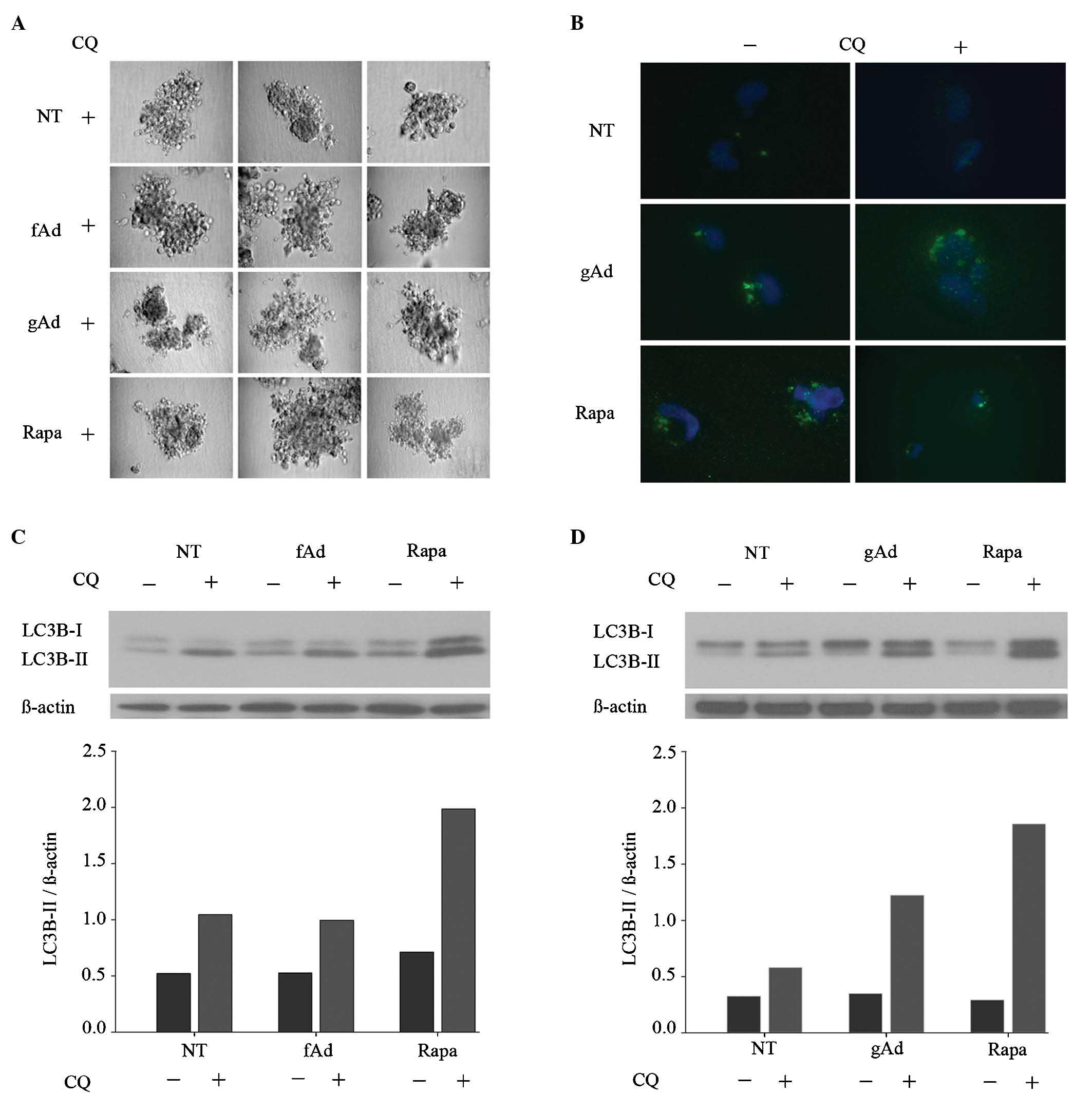 | Figure 2.gAd promotes autophagic induction in
breast cancer cells. (A) MDA-MB-231 cells cultured in serum-free
medium were seeded onto Matrigel™-coated cell culture plates and
treated with human recombinant fAd (0.5 µg/ml), human recombinant
gAd (0.5 µg/ml) or rapa (100 nM). Untreated cells were used as
controls. CQ (50 µM) was added to all the treatment conditions,
which were evaluated in triplicate. Representative images of each
replicate were obtained 9 days subsequent to seeding. (B)
Immunofluorescence analysis revealed that gAd (0.5 µg/ml) and rapa
(100 nM) similarly increased the number of LC3B puncta. Each
condition was evaluated in triplicate, and representative images
are shown. (C and D) Immunoblotting results revealed that fAd (0.5
µg/ml) reduced, while gAd (0.5 µg/ml) and rapa (100 nM) increased
the expression levels of LC3B-II, compared with NT controls. CQ was
added as an autophagy inhibitor to standardize the comparisons
between the different conditions tested. gAd, globular adiponectin;
fAd, full-length adiponectin; rapa, rapamycin; CQ, chloroquine;
LC3B, microtubule-associated protein 1 light chain 3 beta; NT, no
treatment. |
Autophagic induction contributes to
gAd-enhanced invasion
To assess whether autophagic induction may mediate
the gAd-induced increase in migration and invasion observed in 231
cells, RNA interference was used to silence ATG7 in these cells.
ATG7 is responsible for the conversion of LC3B-I to LC3B-II and for
the conjugation of ATG5 to ATG12, which is required for
autophagosome formation during the initial stages of autophagy
(46). Cells were transfected with
three distinct siRNAs that targeted non-overlapping sequences of
ATG7, in addition to a non-targeting negative control siRNA. Upon
verifying the silencing efficiency of the different siRNAs by
immunoblotting (Fig. 3A), an
effective sequence, ATG7 siRNA1, was selected for the subsequent
experimental assays. Silencing ATG7 attenuated the enhancement in
cell migration caused by gAd and rapamycin (Fig. 3B), although knocking down ATG7 also
significantly reduced migration in untreated cells (P<0.001).
While gAd increased invasion by 32% (P=0.09) in control cells
(Fig. 3C), its effect was almost
completely abolished (<0.1%; P=0.97) in cells with impaired
autophagic function. In addition, the rapamycin-induced increase in
invasion was attenuated in ATG7 knockdown (28%; P=0.14), compared
with control (99%; P<0.001) cells. Overall, the results of the
present study are consistent with autophagic induction contributing
to gAd-enhanced cell invasion.
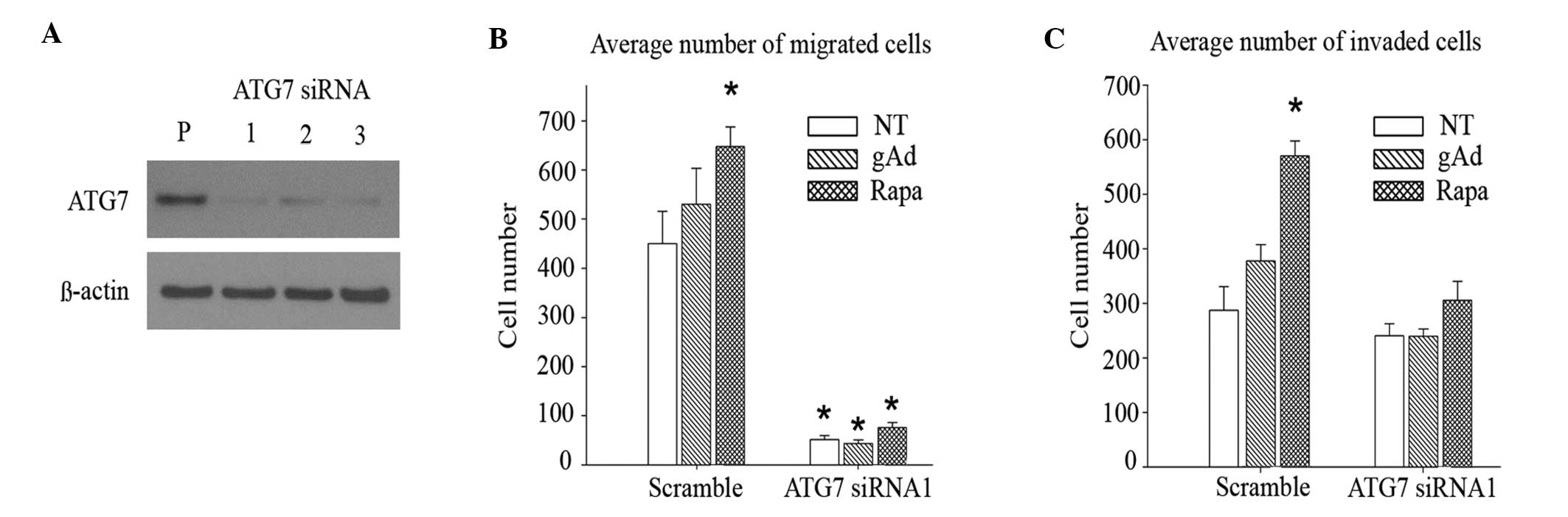 | Figure 3.Autophagic induction contributes to
gAd-enhanced invasion. (A) siRNA silencing of ATG7 was confirmed by
immunoblot analysis. In cells where ATG7 had been knocked down, the
enhanced (B) migration and (C) invasion abilities induced by gAd
(0.5 µg/ml) and rapa (100 nM), respectively, were reduced, compared
with cells transfected with scramble siRNA, which was used as
non-targeting siRNA control. Data are represented as the
mean±standard error of triplicate wells. *P<0.001. gAd, globular
adiponectin; siRNA, small interfering RNA; ATG7, autophagy related
protein 7; rapa, rapamycin; P, parental; NT, no treatment. |
Discussion
In the present study, the effects of different
AdipoQ isoforms on the metastatic potential of breast cancer cells
were compared. In the triple-negative human metastatic cell line
MDA-MB-231, it was identified that gAd, but not fAd, increased
migration, invasion and invasive cell morphology without altering
proliferation. There is currently limited data on the roles of gAd
in breast cancer (20,31). Grossmann et al (20) reported that gAd reduced proliferation
in 231 cells transfected with the estrogen receptor-α (ERα) gene,
but not in parental ERα− 231 cells (20). Mauro et al (31) observed that gAd inhibited
proliferation in ERα− 231 and SK-BR-3 cells, but
increased proliferation in ERα+ MCF7 and T47D cells.
This group additionally reported that gAd downregulated the
expression of cyclin D1 in 231 cells (47) and limited their anchorage-independent
growth, while in MCF7 cells, gAd upregulated the expression of
cyclin D1 (47) and increased their
anchorage-independent growth, cell-cell adhesion and 3D growth.
Furthermore, Jia et al (48)
observed that an unspecified AdipoQ isoform increased the migration
ability of MDA-MB-436 and MFM-223 cells. Although additional
studies are required to fully elucidate the influence of gAd on
breast cancer metastasis, the results of the present study suggest
that gAd possesses unique functions compared with fAd regarding the
promotion of cellular processes that enable the invasion and
dissemination of metastatic breast tumor cells.
A number of studies on other types of cancer support
the hypothesis that gAd and fAd act differentially in terms of
eliciting metastasis-promoting cellular phenotypes (38–41,49,50).
Thus, gAd, but not fAd, increased migration and angiogenesis in
human microvascular endothelial HMEC-1 cells (38). In the human colorectal adenocarcinoma
cell line HT-29, gAd, but not fAd, increased the messenger RNA
(mRNA) expression levels and the secretion of proinflammatory
interleukin 8 (IL-8), granulocyte-macrophage colony-stimulating
factor and monocyte chemoattractant protein-1, and promoted the
nuclear translocation of nuclear factor-kappa B (NF-κB) (40). In human esophageal adenocarcinoma OE19
cells, gAd increased, while fAd reduced the activation of NF-κB,
the production of intracellular ROS and the mRNA levels of tumor
necrosis factor-α, IL-8 and IL-6 (49,50). In
addition, gAd enhanced the production of ROS in monocytes and
neutrophils treated with N-formyl-methionyl-leucyl-phenyl-alanine
(39), and reduced the apoptosis
induced by hypoxia-reoxygenation treatment (41). These observations, together with the
results of the present study, highlight the importance of assessing
the functions of gAd and fAd.
Based on potential associations between the 5′
adenosine monophosphate-activated protein kinase (AMPK)-mediated
signaling pathways and the observation in the present study that
gAd and rapamycin similarly increased metastatic potential in
breast cancer cells, it was hypothesized that gAd may promote
invasiveness of breast cancer cells partly by inducing autophagy.
Cells exposed to a dual treatment of gAd or rapamycin plus
chloroquine, a positive control for autophagy induction, exhibited
a markedly less invasive 3D morphology than those treated with gAd
and rapamycin alone. However, these findings must be interpreted
with caution, since the autophagy inhibitor chloroquine is capable
of exerting a wide range of effects that are not specific to
autophagy (46). Nevertheless, the
results of the present study were supported by the observation that
gAd and rapamycin similarly upregulated LC3B-II, a key marker of
autophagosome formation in the early stages of autophagy (46).
The association between gAd and autophagic induction
in breast cancer cells revealed in the present study is consistent
with the results of previous studies on other types of cancer
(51–53). However, to the best of our knowledge,
the aforementioned association has not been studied in breast
cancer to date. In the present study, fAd did not alter the levels
of LC3B-II, in contrast to a previous study that identified
increased autophagosome and autophagolysosome formation in 231
cells following fAd treatment (54).
In that study, the authors treated cells cultured in
FBS-supplemented medium, as opposed to SFM, and used substantially
higher concentrations of fAd than the ones used in the present
study (54). Therefore, differences
in the experimental conditions may have contributed to the
inconsistencies observed across the two studies, although they were
not evaluated in detail, since they are beyond the scope of the
present study.
Knockdown of ATG7 reduced the gAd and
rapamycin-induced promotion of cell invasion, providing evidence
that gAd may act, at least in part, through autophagic pathways to
affect metastatic behavior. However, ATG7 knockdown also reduced
the migration ability of untreated cells, suggesting that gAd may
promote migration through alternative pathways. By contrast, there
was no significant alteration of invasion between the untreated
control and ATG7 knockdown cells. Although the reasons for the
differences in migration and invasion observed with ATG7 knockdown
remain to be elucidated, the results of the present study are
consistent with autophagic induction as a potential mechanism
contributing to the gAd-enhanced invasiveness of breast cancer
cells.
AdipoQ has been previously detected in breast cancer
and adjacent tissue (4–6), although the distribution of gAd versus
fAd has not been investigated in these previous studies. The
authors of the present study recently proposed a model in which gAd
is locally elevated in tumor tissue, where it enhances the
potential of tumor cells to metastasize by upregulating the
autophagic response through AdipoQ receptor 1 (ADIPOR1)-mediated
activation of AMPK (35). A previous
report in colorectal cancer identified significantly increased
protein expression levels of gAd in tumor tissue of patients with
colorectal cancer, compared with adjacent normal mucosa tissue. By
contrast, fAd displayed the opposite trend (55). Furthermore, increased expression
levels of gAd have been previously correlated with increased mRNA
expression levels of AMPK and ADIPOR1 (2).
In conclusion, although the results of the present
study are limited to a single cell line, they nonetheless provide
initial evidence that the different AdipoQ isoforms present in the
breast tumor microenvironment may exert different effects. The
present study has demonstrated that gAd is able to differentially
act on breast cancer cells to promote processes that facilitate
metastatic progression, and that autophagic induction may mediate
this effect. Furthermore, the present study is timely, considering
the current interest in the preclinical development of AdipoQ and
AdipoQ receptor-based therapies for the treatment of breast cancer
(56–58) and the clinical use of autophagy
modulators (59,60). Additional studies are required to
understand the specific association between different AdipoQ
isoforms and metastasis, in order to optimize the effects of
emerging therapies.
Acknowledgements
The present study was partly funded by the American
Cancer Society (Atlanta, GA, USA) (grant no. RSG-11-259-01-CSM),
METAvivor Research and Support, Inc. (Annapolis, MD, USA) (grant
no. awarded to DRH) and the Cancer Prevention and Control Training
Program of the University of Alabama at Birmingham (Birmingham, AL,
USA) (grant no. R25 CA047888 awarded to WDW and EFL).
Glossary
Abbreviations
Abbreviations:
|
AdipoQ
|
adiponectin
|
|
fAd
|
full-length adiponectin
|
|
gAd
|
globular adiponectin
|
|
rapa
|
rapamycin
|
|
CQ
|
chloroquine
|
|
SFM
|
serum-free medium
|
|
NT
|
no treatment
|
References
|
1
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dalamaga M, Diakopoulos KN and Mantzoros
CS: The role of adiponectin in cancer: A review of current
evidence. Endocr Rev. 33:547–594. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tan J, Buache E, Chenard MP, Dali-Youcef N
and Rio MC: Adipocyte is a non-trivial, dynamic partner of breast
cancer cells. Int J Dev Biol. 55:851–859. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jeong YJ, Bong JG, Park SH, Choi JH and Oh
HK: Expression of leptin, leptin receptor, adiponectin, and
adiponectin receptor in ductal carcinoma in situ and
invasive breast cancer. J Breast Cancer. 14:96–103. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karaduman M, Bilici A, Ozet A, Sengul A,
Musabak U and Alomeroglu M: Tissue levels of adiponectin in breast
cancer patients. Med Oncol. 24:361–366. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morad V, Abrahamsson A and Dabrosin C:
Estradiol affects extracellular leptin:adiponectin ratio in human
breast tissue in vivo. J Clin Endocrinol Metab.
99:3460–3467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Waki H, Yamauchi T, Kamon J, Ito Y, Uchida
S, Kita S, Hara K, Hada Y, Vasseur F, Froguel P, et al: Impaired
multimerization of human adiponectin mutants associated with
diabetes. Molecular structure and multimer formation of
adiponectin. J Biol Chem. 278:40352–40363. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pajvani UB, Du X, Combs TP, Berg AH,
Rajala MW, Schulthess T, Engel J, Brownlee M and Scherer PE:
Structure-function studies of the adipocyte-secreted hormone
Acrp30/adiponectin. Implications for metabolic regulation and
bioactivity. J Biol Chem. 278:9073–9085. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fruebis J, Tsao TS, Javorschi S,
Ebbets-Reed D, Erickson MRS, Yen FT, Bihain BE and Lodish HF:
Proteolytic cleavage product of 30-kDa adipocyte complement-related
protein increases fatty acid oxidation in muscle and causes weight
loss in mice. Proc Natl Acad Sci USA. 98:2005–2010. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Waki H, Yamauchi T, Kamon J, Kita S, Ito
Y, Hada Y, Uchida S, Tsuchida A, Takekawa S and Kadowaki T:
Generation of globular fragment of adiponectin by leukocyte
elastase secreted by monocytic cell line THP-1. Endocrinology.
146:790–796. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamashita JI, Ogawa M, Ikei S, Omachi H,
Yamashita SI, Saishoji T, Nomura K and Sato H: Production of
immunoreactive polymorphonuclear leucocyte elastase in human breast
cancer cells: Possible role of polymorphonuclear leucocyte elastase
in the progression of human breast cancer. Br J Cancer. 69:72–76.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Foekens JA, Ries C, Look MP,
Gippner-Steppert C, Klijn JG and Jochum M: The prognostic value of
polymorphonuclear leukocyte elastase in patients with primary
breast cancer. Cancer Res. 63:337–341. 2003.PubMed/NCBI
|
|
13
|
Akizuki M, Fukutomi T, Takasugi M,
Takahashi S, Sato T, Harao M, Mizumoto T and Yamashita J:
Prognostic significance of immunoreactive neutrophil elastase in
human breast cancer: Long-term follow-up results in 313 patients.
Neoplasia. 9:260–264. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu LY, Wang M, Ma ZB, Yu LX, Zhang Q, Gao
DZ, Wang F and Yu ZG: The role of adiponectin in breast cancer: A
meta-analysis. PLoS One. 8:e731832013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Körner A, Pazaitou-Panayiotou K, Kelesidis
T, Kelesidis I, Williams CJ, Kaprara A, Bullen J, Neuwirth A,
Tseleni S, Mitsiades N, et al: Total and high-molecular-weight
adiponectin in breast cancer: In vitro and in vivo
studies. J Clin Endocrinol Metab. 92:1041–1048. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen DC, Chung YF, Yeh YT, Chaung HC, Kuo
FC, Fu OY, Chen HY, Hou MF and Yuan SS: Serum adiponectin and
leptin levels in Taiwanese breast cancer patients. Cancer Lett.
237:109–114. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mantzoros C, Petridou E, Dessypris N,
Chavelas C, Dalamaga M, Alexe DM, Papadiamantis Y, Markopoulos C,
Spanos E, Chrousos G and Trichopoulos D: Adiponectin and breast
cancer risk. J Clin Endocrinol Metab. 89:1102–1107. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Miyoshi Y, Funahashi T, Kihara S, Taguchi
T, Tamaki Y, Matsuzawa Y and Noguchi S: Association of serum
adiponectin levels with breast cancer risk. Clin Cancer Res.
9:5699–5704. 2003.PubMed/NCBI
|
|
19
|
Tworoger SS, Eliassen AH, Kelesidis T,
Colditz GA, Willett WC, Mantzoros CS and Hankinson SE: Plasma
adiponectin concentrations and risk of incident breast cancer. J
Clin Endocrinol Metab. 92:1510–1516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Grossmann ME, Nkhata KJ, Mizuno NK, Ray A
and Cleary MP: Effects of adiponectin on breast cancer cell growth
and signaling. Br J Cancer. 98:370–379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dos Santos E, Benaitreau D, Dieudonne MN,
Leneveu MC, Serazin V, Giudicelli Y and Pecquery R: Adiponectin
mediates an antiproliferative response in human MDA-MB 231 breast
cancer cells. Oncol Rep. 20:971–977. 2008.PubMed/NCBI
|
|
22
|
Duggan C, Irwin ML, Xiao L, Henderson KD,
Smith AW, Baumgartner RN, Baumgartner KB, Bernstein L,
Ballard-Barbash R and McTiernan A: Associations of insulin
resistance and adiponectin with mortality in women with breast
cancer. J Clin Oncol. 29:32–39. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Arditi JD, Venihaki M, Karalis KP and
Chrousos GP: Antiproliferative effect of adiponectin on MCF7 breast
cancer cells: A potential hormonal link between obesity and cancer.
Horm Metab Res. 39:9–13. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Taliaferro-Smith L, Nagalingam A, Zhong D,
Zhou W, Saxena NK and Sharma D: LKB1 is required for
adiponectin-mediated modulation of AMPK-S6K axis and inhibition of
migration and invasion of breast cancer cells. Oncogene.
28:2621–2633. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taliaferro-Smith L, Nagalingam A, Knight
BB, Oberlick E, Saxena NK and Sharma D: Integral role of PTP1B in
adiponectin-mediated inhibition of oncogenic actions of leptin in
breast carcinogenesis. Neoplasia. 15:23–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Treeck O, Lattrich C, Juhasz-Boess I,
Buchholz S, Pfeiler G and Ortmann O: Adiponectin differentially
affects gene expression in human mammary epithelial and breast
cancer cells. Br J Cancer. 99:1246–1250. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Lam JB, Lam KS, Liu J, Lam MC, Hoo
RL, Wu D, Cooper GJ and Xu A: Adiponectin modulates the glycogen
synthase kinase-3beta/beta-catenin signaling pathway and attenuates
mammary tumorigenesis of MDA-MB-231 cells in nude mice. Cancer Res.
66:11462–11470. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dieudonne MN, Bussiere M, Dos Santos E,
Leneveu MC, Giudicelli Y and Pecquery R: Adiponectin mediates
antiproliferative and apoptotic responses in human MCF7 breast
cancer cells. Biochem Biophys Res Commun. 345:271–279. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakayama S, Miyoshi Y, Ishihara H and
Noguchi S: Growth-inhibitory effect of adiponectin via adiponectin
receptor 1 on human breast cancer cells through inhibition of
S-phase entry without inducing apoptosis. Breast Cancer Res Treat.
112:405–410. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pfeiler GH, Buechler C, Neumeier M,
Schäffler A, Schmitz G, Ortmann O and Treeck O: Adiponectin effects
on human breast cancer cells are dependent on 17-beta estradiol.
Oncol Rep. 19:787–793. 2008.PubMed/NCBI
|
|
31
|
Mauro L, Pellegrino M, De Amicis F,
Ricchio E, Giordano F, Rizza P, Catalano S, Bonofiglio D, Sisci D,
Panno ML and Andò S: Evidences that estrogen receptor α interferes
with adiponectin effects on breast cancer cell growth. Cell Cycle.
13:553–564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim KY, Baek A, Hwang JE, Choi YA, Jeong
J, Lee MS, Cho DH, Lim JS, Kim KI and Yang Y: Adiponectin-activated
AMPK stimulates dephosphorylation of AKT through protein
phosphatase 2A activation. Cancer Res. 69:4018–4026. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang JH, Yu BY and Youn DS: Relationship
of serum adiponectin and resistin levels with breast cancer risk. J
Korean Med Sci. 22:117–121. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Macis D, Gandini S, Guerrieri-Gonzaga A,
Johansson H, Magni P, Ruscica M, Lazzeroni M, Serrano D, Cazzaniga
M, Mora S, et al: Prognostic effect of circulating adiponectin in a
randomized 2 × 2 trial of low-dose tamoxifen and fenretinide in
premenopausal women at risk for breast cancer. J Clin Oncol.
30:151–157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Libby EF, Frost AR, Demark-Wahnefried W
and Hurst DR: Linking adiponectin and autophagy in the regulation
of breast cancer metastasis. J Mol Med Berl. 92:1015–1023. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Llanos AA, Dumitrescu RG, Marian C,
Makambi KH, Spear SL, Kallakury BV, Perry DJ, Convit RJ, Platek ME,
Millen AE, et al: Adipokines in plasma and breast tissues:
Associations with breast cancer risk factors. Cancer Epidemiol
Biomarkers Prev. 21:1745–1755. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sonmez B, Seker M, Bilici A, Yavuz Erkal
F, Oven Ustaalioglu BB, Gumus M, Ozturk Guler D, Karaduman M, Gezen
C, Eser M, et al: Is there any correlation among adiponectin levels
in serum, tumor tissue and normal tissue of the same patients with
breast cancer? J BUON. 16:227–232. 2011.PubMed/NCBI
|
|
38
|
Adya R, Tan BK, Chen J and Randeva HS:
Protective actions of globular and full-length adiponectin on human
endothelial cells: Novel insights into adiponectin-induced
angiogenesis. J Vasc Res. 49:534–543. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chedid P, Hurtado-Nedelec M, Marion-Gaber
B, Bournier O, Hayem G, Gougerot-Pocidalo MA, Frystyk J, Flyvbjerg
A, El Benna J and Marie JC: Adiponectin and its globular fragment
differentially modulate the oxidative burst of primary human
phagocytes. Am J Pathol. 180:682–692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ogunwobi OO and Beales IL: Adiponectin
stimulates proliferation and cytokine secretion in colonic
epithelial cells. Regul Pept. 134:105–113. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Park M, Youn B, Zheng XL, Wu D, Xu A and
Sweeney G: Globular adiponectin, acting via AdipoR1/APPL1, protects
H9c2 cells from hypoxia/reoxygenation-induced apoptosis. PLoS One.
6:e191432011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hurst DR, Xie Y, Vaidya KS, Mehta A, Moore
BP, Accavitti-Loper MA, Samant RS, Saxena R, Silveira AC and Welch
DR: Alterations of BRMS1-ARID4A interaction modify gene expression
but still suppress metastasis in human breast cancer cells. J Biol
Chem. 283:7438–7444. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cody JJ, Markert JM and Hurst DR: Histone
deacetylase inhibitors improve the replication of oncolytic herpes
simplex virus in breast cancer cells. PLoS One. 9:e929192014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Talmadge JE and Fidler IJ: AACR centennial
series: The biology of cancer metastasis: Historical perspective.
Cancer Res. 70:5649–5669. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
White E: Deconvoluting the
context-dependent role for autophagy in cancer. Nat Rev Cancer.
12:401–410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Klionsky DJ, Abdalla FC, Abeliovich H,
Abraham RT, Acevedo-Arozena A, Adeli K, Agholme L, Agnello M,
Agostinis P, Aguirre-Ghiso JA, et al: Guidelines for the use and
interpretation of assays for monitoring autophagy. Autophagy.
8:445–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mauro L, Pellegrino M, Giordano F, Ricchio
E, Rizza P, De Amicis F, Catalano S, Bonofiglio D, Panno ML and
Andò S: Estrogen receptor-α drives adiponectin effects on cyclin D1
expression in breast cancer cells. FASEB J. 29:2150–2160. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jia Z, Liu Y and Cui S: Adiponectin
induces breast cancer cell migration and growth factor expression.
Cell Biochem Biophys. 70:1239–1245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang R, Wu J, Liu D, Shan H and Zhang J:
Anti-inflammatory effect of full-length adiponectin and
proinflammatory effect of globular adiponectin in esophageal
adenocarcinoma cells. Oncol Res. 21:15–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang R, Yin X, Shi H, Wu J, Shakya P, Liu
D and Zhang J: Adiponectin modulates DCA-induced inflammation via
the ROS/NF-κ B signaling pathway in esophageal adenocarcinoma
cells. Dig Dis Sci. 59:89–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Habeeb BS, Kitayama J and Nagawa H:
Adiponectin supports cell survival in glucose deprivation through
enhancement of autophagic response in colorectal cancer cells.
Cancer Sci. 102:999–1006. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nepal S, Kim MJ, Lee ES, Kim JA, Choi DY,
Sohn DH, Lee SH, Song K, Kim SH, Jeong GS, et al: Modulation of
Atg5 expression by globular adiponectin contributes to autophagy
flux and suppression of ethanol-induced cell death in liver cells.
Food Chem Toxicol. 68:11–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nepal S and Park PH: Activation of
autophagy by globular adiponectin attenuates ethanol-induced
apoptosis in HepG2 cells: Involvement of AMPK/FoxO3A axis. Biochim
Biophys Acta. 1833:2111–2125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu J, Xu A, Lam KS, Wong NS, Chen J,
Shepherd PR and Wang Y: Cholesterol-induced mammary tumorigenesis
is enhanced by adiponectin deficiency: Role of LDL receptor
upregulation. Oncotarget. 4:1804–1818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Vetvik KK, Sonerud T, Lindeberg M, Lüders
T, Størkson RH, Jonsdottir K, Frengen E, Pietiläinen KH and Bukholm
I: Globular adiponectin and its downstream target genes are
up-regulated locally in human colorectal tumors: Ex vivo and
in vitro studies. Metabolism. 63:672–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Delort L, Jardé T, Dubois V, Vasson MP and
Caldefie-Chézet F: New insights into anticarcinogenic properties of
adiponectin: A potential therapeutic approach in breast cancer?
Vitam Horm. 90:397–417. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Khan S, Shukla S, Sinha S and Meeran SM:
Role of adipokines and cytokines in obesity-associated breast
cancer: Therapeutic targets. Cytokine Growth Factor Rev.
24:503–513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Otvos L Jr, Haspinger E, La Russa F,
Maspero F, Graziano P, Kovalszky I, Lovas S, Nama K, Hoffmann R,
Knappe D, et al: Design and development of a peptide-based
adiponectin receptor agonist for cancer treatment. BMC Biotechnol.
11:902011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Gewirtz DA: The four faces of autophagy:
Implications for cancer therapy. Cancer Res. 74:647–651. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Vinayak S and Carlson RW: mTOR inhibitors
in the treatment of breast cancer. Oncology (Williston Park).
27:38–48. 2013.PubMed/NCBI
|














