Introduction
Scorpion stings are a major health hazard worldwide,
and in the Saudi Arabian population in particular (1). High rates of incidence of scorpion
stings have been reported in Northwestern regions of Saudi Arabia
(2,3).
Approximately 1,500 species of scorpion have been reported so far,
including one of the oldest species on earth (4). As reported, the incidence of scorpion
stings between the years of 1993–1997 was 72,168 individuals. Of
these, ~25 species of scorpion exhibiting high toxicity have been
identified in Saudi Arabia (5). The
climatic, socioeconomic and cultural circumstances of Saudi Arabia
further promote the occurrence of scorpion stings in the local
inhabitants. There are an estimated 14,500 cases/year of scorpion
stings in the different regions of Saudi Arabia (6). The venom from the Androctonus
genus, which is commonly observed in Saudi Arabia, is considered to
be the most toxic type of scorpion venom, compared with the venom
of other species (7).
The sting and envenomation of the scorpion generates
discomfort and pain in the victims, and may develop into a number
of medical complications, including hypotension, cardiac arrhythmia
and respiratory distress (8). The
mechanism underlying scorpion stings and their associated health
complications is due to the venom acting as a potent neurotoxin,
which causes the inhibition of the
Na+/K+-ATPase pump, thus paralyzing the
sympathetic and parasympathetic nervous systems, with
severe-to-fatal consequences (9).
Biochemically, scorpion venom contains several basic
proteins that are responsible for its neurotoxicity (10). Amino acid sequencing of venom obtained
from different species of scorpion revealed that the above
neurotoxins consist of 60–70 amino acid residues, which are
cross-linked by four disulfide bridges (10). Furthermore, due to the close
similarities in their amino acid composition, all scorpion toxins
possess similar three-dimensional structures (10). In spite of the various negative
effects of scorpion envenomation, scorpion venom contains numerous
beneficial components, which are currently being utilized for drug
design in the pharmaceutical industry (11,12). A
number of antibacterial, antifungal (yeast) and antiviral
substances have been derived from peptides isolated from scorpion
venom (13,14). Additionally, scorpion venom has
demonstrated anticancer potential in pancreatic cancer (15) and leukemia (16). Scorpion venom is constituted by a
complex mixture of salts, nucleotides, biogenic amines, peptides,
enzymes, mucoproteins and other proteins such as neurotoxins
(17). It has been recently observed
that animal venoms and toxins possess anticancer potential.
Previous studies have revealed that scorpion venoms and toxins are
able to reduce cancer growth, induce apoptosis and inhibit
progression and metastasis in vivo and in vitro
(17). A number of active molecules
with anticancer properties, including induction of cell cycle
arrest and apoptosis, inhibition of proliferation and reduction of
cell migration and invasion, have been isolated from scorpion venom
(17). These observations have
provided insight into the application of scorpion venom and toxins
as possible novel anticancer therapeutics (17).
Cancer or malignancy may be defined as the
uncontrolled growth of cells leading to the development of disease.
The most remarkable feature of cancerous cells is their capacity to
invade and metastasize to different regions of the body and form
secondary tumors in these organs. The current study analyzed the
effects of scorpion venom on cancer cells. In particular, of
phenotypic alterations such as variations in cell motility and
clonogenic survival were assessed using an anchorage-dependent
colony formation assay. A total of three distinct cancer cell lines
derived from colorectal (HCT-8 and HCT-116) and breast (MDA-MB-231)
cancer were selected for the present study. The rationale behind
selecting the above cell lines was the ease of availability of
these cells at the Research Center of Prince Sultan Military
Medical City Hospital (Riyadh, Saudi Arabia) and the prevalence of
these two malignancies among the Saudi Arabian population.
Colorectal cancer is cancer of the colon or rectum,
which is part of the large intestine or gastrointestinal system.
Colon cancer is the third most fatal type of cancer worldwide among
men and women. In 2014, it was estimated that ~138,000 patients
were diagnosed with colorectal cancer in USA, of which, ~50%
succumbed to disease (18).
Similarly, breast cancer is the second cause of cancer-associated
mortality worldwide among women (19). As reported by the National Saudi
Cancer Registry (Riyadh, Saudi Arabia), a case study conducted
between 2001 and 2006 identified a significant increase in the
occurrence of colorectal cancer among the Saudi Arabian population
(20). A similar study conducted by
Alghamdi et al (21) between
2001 and 2008 reported a significant increase in female breast
cancer, particularly among young women, in Saudi Arabia. The above
study also specified the geographical locations of the prevailing
disease, and identified that the Eastern region of Saudi Arabia
presented a linear upward trend in the occurrence of breast cancer,
compared with Southern areas of the country, including Jazan, Baha
and Najran (21).
Cell invasion, migration and metastasis to different
parts of the body are the most important characteristic features of
cancer cells. The migration of cancerous cells depends on the tumor
microenvironment, from which the cells receive nourishment and
support for the formation of novel vasculature (angiogenesis),
which enables them to spread (22).
The degradation of the extracellular matrix (ECM) facilitates the
movement of cancer cells in the body (23). Cancerous tumors are comprised of
aberrant cells in addition to macromolecules from the ECM, which
constitute a substantial part of their volume. These macromolecules
are predominantly polysaccharides and proteins of various types
that are secreted locally, which are arranged in a mesh-like
structure and are closely associated with the malignant tumor.
Previous studies have revealed the role of matrix
metalloproteinases (MMPs) in the distortion of the ECM meshwork,
which results in the generation of a free passage for cancer cells
to migrate and invade distal parts of the body, thus leading to the
progression of metastatic disease (24,25). MMPs
are zinc-dependent endopeptidases that belong to the metzincin
enzyme superfamily and may adversely affect the architecture of the
ECM network (26–28).
Recent studies have demonstrated that a 36-amino
acid peptide sequence, which contains four disulfide bridges and
derives from scorpion venom, is effective in inhibiting the action
of various MMPs in glioblastoma (29–31),
pancreatic (15) and breast cancer
(32). However, there are limited
studies on the effect of scorpion venom as an anticancer agent on
colorectal cancer. In those previous studies, MMPs were observed to
be constitutively overexpressed (33). Therefore, the aim of the present study
was to investigate the anticancer potential of the scorpion venom
obtained at the Research Center of Prince Sultan Military Medical
City Hospital, against colorectal and breast cancer cell lines.
The findings of the present study indicated that
reduced cell motility and colony formation correlated with the
inhibitory role of scorpion venom on colorectal and breast cancer
cell lines. Furthermore, these results suggested that venom therapy
may constitute an important step towards the development of a more
specific treatment for aggressive types of cancer. To the best of
our knowledge, the present study is the first to demonstrate the
phenotypic changes that occur in colorectal and breast cancer cell
lines following treatment with scorpion venom.
Materials and methods
Scorpions and venom collection
Medically important species of scorpion, including
Androctonus crassicauda, Androctonus bicolor and Leiurus
quinquestriatus, were collected from various regions of Saudi
Arabia and housed at Prince Sultan Military Medical City Hospital
(Riyadh, Saudi Arabia). A male and a female scorpion were housed in
a plastic box and fed with mealworms and water ad libitum.
Temperature of the room kept at 25°C with 12 h light and dark
cycles. The Research Ethics Committee of Prince Sultan Military
Medical City Hospital approved the present study (project no.
1a/2013). Scorpion venom was milked by electrical stimulation using
6012 Dual Pulse Stimulator (Harvard Apparatus, Holliston, MA, USA).
The ejected venom was collected in glass beakers and immediately
stored at −20°C. The recovery of venom was achieved by mixing the
sample with distilled water, followed by centrifugation at 199 × g
for 10 min. The supernatant was then lyophilized and stored at
−80°C until required. A stock solution of venom at a concentration
of 10 mg/ml was prepared in phosphate-buffered saline (PBS), which
was sterilized by passage through a 0.22-µm membrane filter (Cat
no. SLGN 033RS, Thomas Scientific, Swedesboro, NJ, USA) prior to
use. Additional dilution, as per the experimental requirements, was
performed using an identical buffer system. For simplicity, venoms
were designated as venom-1, −2 and −3, which were obtained from the
scorpion species Androctonus crassicauda, Androctonus bicolor and
Leiurus quinquestriatus, respectively.
Cell culture
Cancer cell lines, namely HCT-8 [derived from
ileocecal adenocarcinoma of a 67-year-old male patient (34)], HCT-116 [derived from colorectal
carcinoma of a male patient (35)]
and MDA-MB-231 [obtained from breast carcinoma (36)], were supplied by the Cancer Research
Facility of King Fahad National Guard Hospital (Riyadh, Saudi
Arabia). The cell lines HCT-8 and HCT-116 were cultured in
RPMI-1640 medium, while MDA-MB-231 cells were cultured in
Dulbecco's Modified Eagle's medium (Life Technologies, Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10%
heat-inactivated fetal bovine serum (PAA Laboratories, GE
Healthcare Life Sciences, Chalfont, UK), supplemented with 2 mM
L-glutamine (cat no. 25030-081), 50 µg/ml penicillin G and 50 µg/ml
streptomycin sulfate (cat no. 10378-016; all Life Technologies,
Thermo Fisher Scientific, Inc.). This combination was designated as
complete medium. The cultures were maintained by replacing the
medium on alternate days until cells reached the desired
confluence. In order to initiate subcultures, 80–90% confluent
cells were washed using Ca2+ and Mg2+-free
warm PBS, prior to be treated with ~3 ml
trypsin-ethylenediaminetetraacetic acid (EDTA) solution (0.05%
trypsin and 0.53 mM EDTA; cat no. 25200-056; Life technologies,
Thermo Fisher Scientific, Inc.) for 5 min at 37°C. Subsequently, 5
ml complete medium was added to the cells to quench the effect of
trypsin, and the cell suspension was next centrifuged at 1,000 rpm
for 5 min. The pelleted cells were resuspended in complete medium
and seeded into 25−cm2 culture flasks at a
density of 1×104 cells/cm2. The cultures were
maintained in a humidified atmosphere at 37°C with 5%
CO2 and 95% O2.
Cell motility assay
Cell motility assays were performed in 6-well cell
culture plates. HCT-8 and MDA-MB-231 cells at ~80% confluence were
washed using PBS, and a fine scratch in the form of a groove was
created in the cell monolayer using a sterile pipette tip, and
immediately photographed. This time point was designated as 0 h.
Subsequently, cells were supplemented with complete medium and
allowed to grow in the presence of various concentrations (20, 40,
60, 80 and 100 µg/ml) of scorpion venom −1, which was obtained from
Androctonus crassicauda. Next HCT-116 cells were treated with 50
and 100 µg/ml scorpion venom −1. Additionally, cell motility and
colony formation in MDA-MB-231 cells were examined following
treatment with 50 and 100 µg/ml of venoms −2 and −3, obtained from
the species of Androctonus bicolor and Leiurus
quinquestriatus, respectively. Control cells were grown in the
absence of venom, and treated with equivalent volumes of PBS
instead.
The migration of cells from the edge of the scratch
towards the center was monitored microscopically at magnification
×10 upon 24 h of incubation, in order to assess the extent of the
scratched area that had been covered by the migrated cells. The
width of the scratch was measured at 0 and 24 h to calculate the
percentage of the gap covered by the cells in a 24-h time
period.
Clonogenic survival assay
The clonogenic survival assay was performed
according to the protocol previously described by Franken et
al (37). Briefly, 500 cells were
cultured in 6-well plates for 2 weeks, in the absence or presence
of various concentrations (20, 40, 60, 80 and 100 µg/ml) of
scorpion venom −1. HCT-116 cells were treated with 50 and 100 µg/ml
scorpion venom −1. Additionally, MDA-MB-231 cells were examined
following treatment with 50 and 100 µg/ml of venoms −2 and −3,
respectively. Control cells were grown in the absence of venom, and
treated with equivalent volumes of PBS instead. Upon termination of
the assay, the cells were washed with PBS, and the colonies formed
were subsequently stained for 45 min with 0.5% (w/v) crystal violet
(cat no. C 3886; Sigma-Aldrich, St. Louis, MO, USA) prepared in
0.6% (v/v) glutaraldehyde solution (cat no. C 3886; Sigma-Aldrich),
rinsed with water and air-dried.
Statistical analysis
Statistical analyses were performed using the
Student's t-test. Results are presented as the mean ± standard
deviation. The data were analyzed in triplicate, using Graph Pad
Prism software, version 4.0 (Graph Pad Software, Inc., San Diego,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Treatment with scorpion venom
decreases motility and colony formation in HCT-8 cells
The present study focused on the in vitro
effects of scorpion venom in three distinct well-established cancer
cell lines, namely HCT-8, HCT-116 and MDA-MB-231. These cell lines
have been well-characterized, and are widely used as models to
investigate various aspects of cancer metastasis. The main aim of
the present study was to evaluate the anticancerous potential of
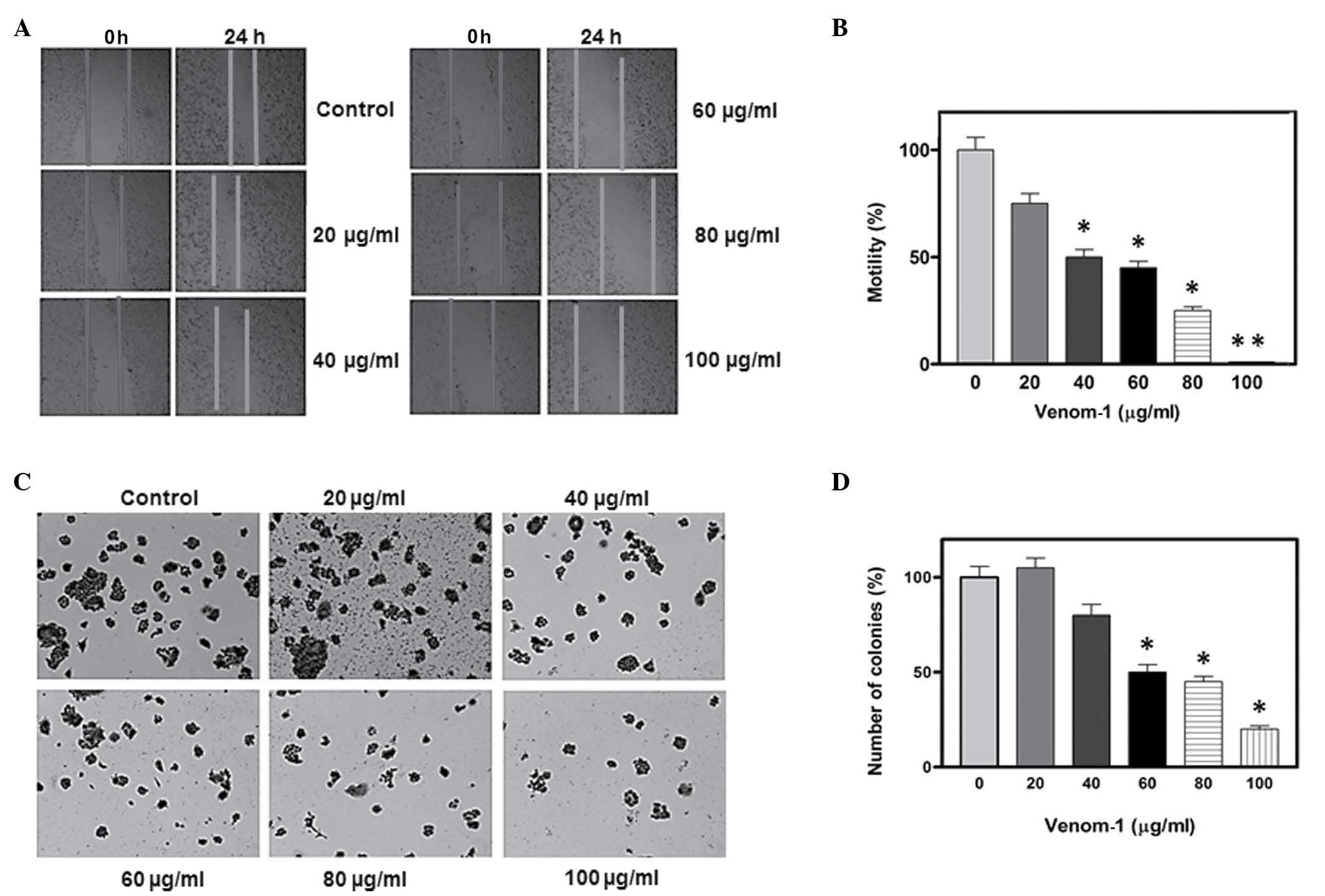
venom obtained from scorpions. As demonstrated in Fig. 1A, the cell motility of the HCT-8 cell
line was observed to decrease in a dose-dependent manner. Notably,
at a concentration of 100 µg/ml of venom-1, a complete halt in cell
motility was observed following 24 h of incubation. Fig. 1B reveals the gradual percentage of
scratch covered in the HCT-8 cell line 24 h subsequent to wounding.
In addition, anchorage-dependent colony formation, a hallmark of
cancer cell survival, was examined. A marked decrease in colony
formation was observed following venom-1 treatment in the HCT-8
cell line, compared with the control (Fig. 1C). Fig.
1D reveals a ~70–90% reduction in colony formation at a
concentration of 80–100 µg/ml of venom-1.
Treatment with scorpion venom
decreases motility and colony formation in HCT-116 cells
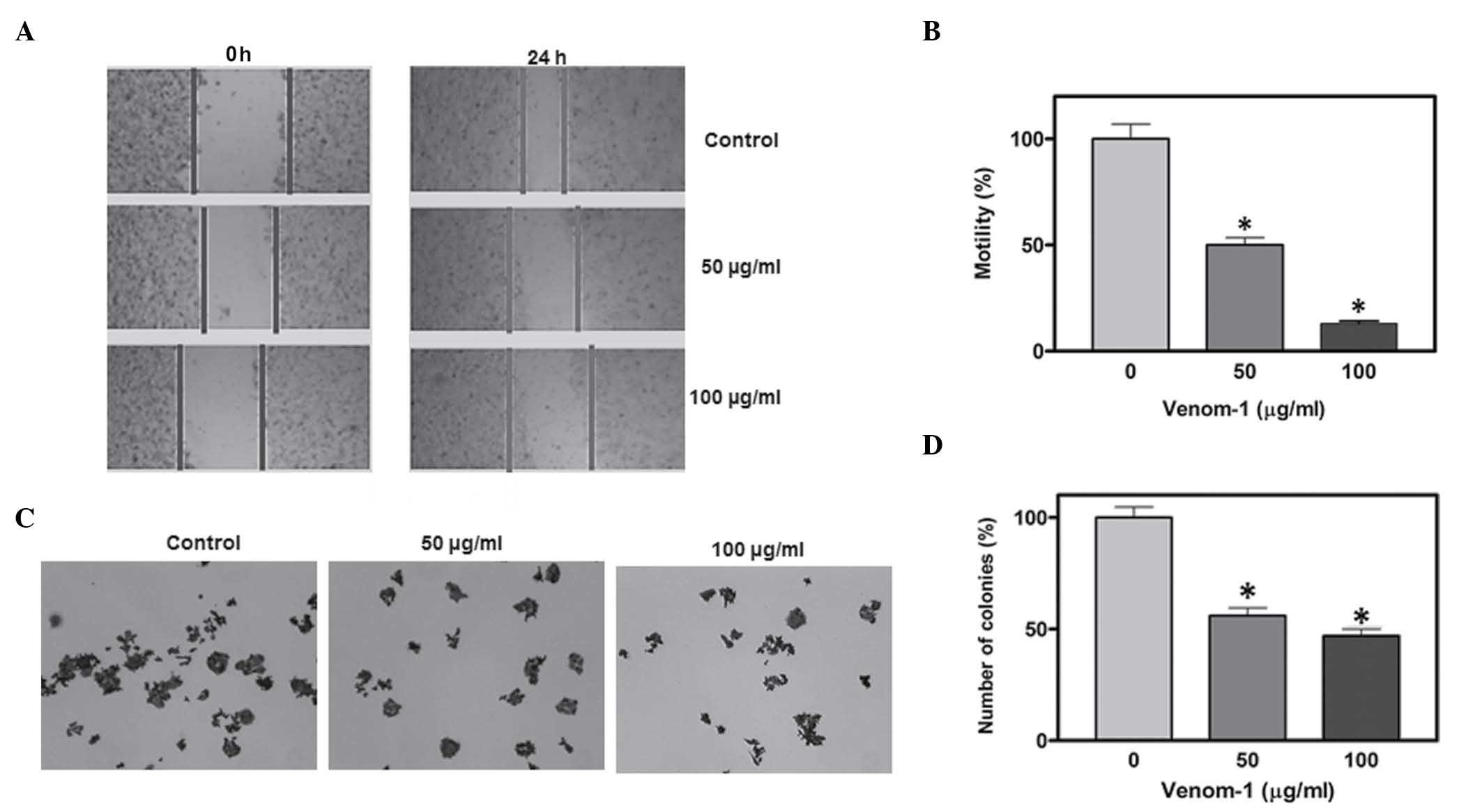
In HCT-116 cells, a significant reduction in cell
motility and colony formation was observed following treatment with
50 and 100 µg/ml venom-1 (Fig. 2A and
C). Statistical analysis of cell motility and colony formation
is depicted in the form of bar graphs (Fig. 2B and D). Asterisks indicate the
significance of cell motility and colony formation in the treatment
group, compared with the control (P<0.05).
Treatment with scorpion venom
decreases motility and colony formation in MDA-MB-231 cells
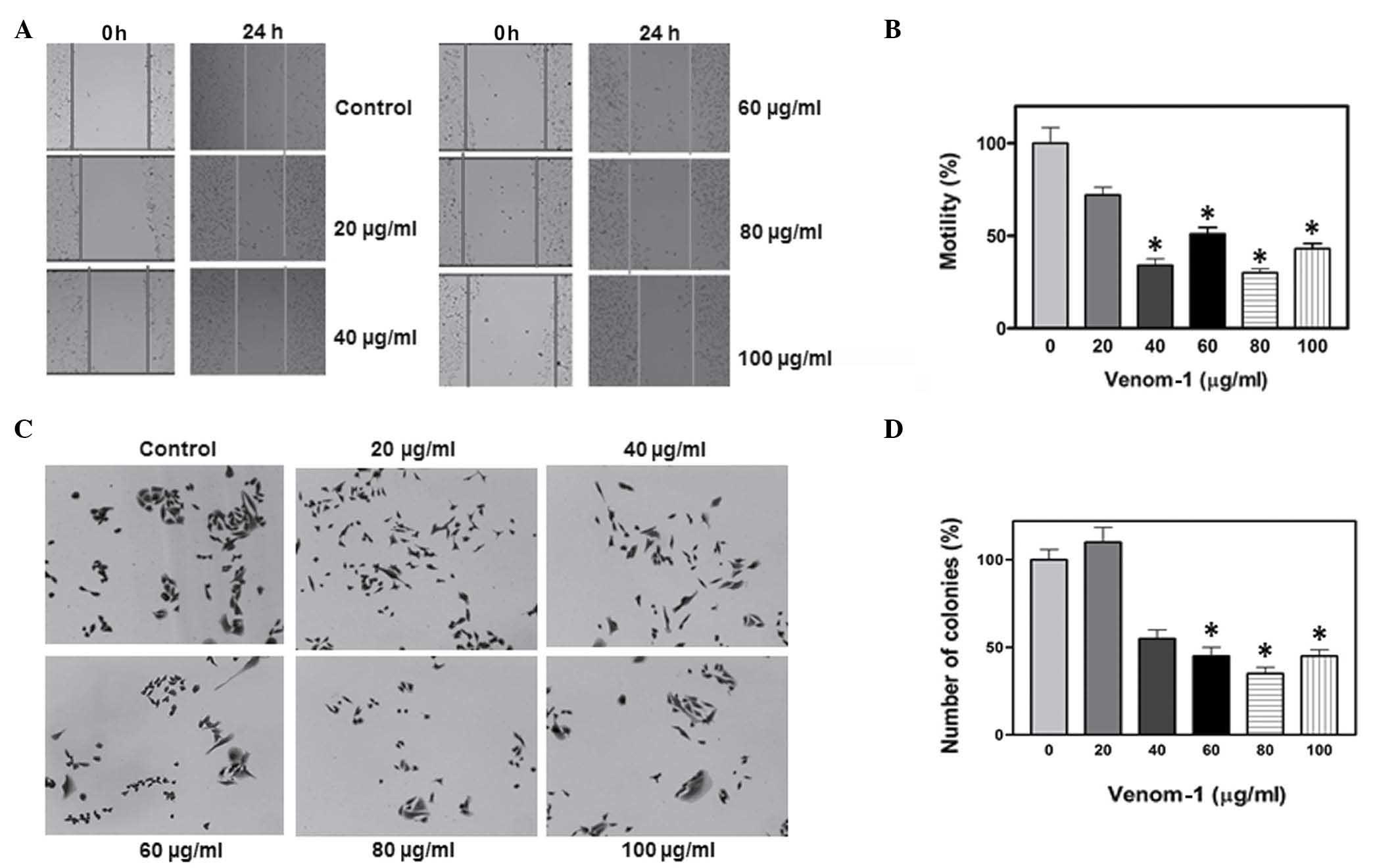
In order to investigate the efficacy of venom-1 as
an anticancer agent in different types of cancer other than
colorectal, a breast cancer cell line, namely MDA-MB-231, was also
included in the present study. A similar pattern of inhibition in
cell motility and colony formation was observed (Fig. 3), confirming that the action of
venom-1 was not restricted to one type of cell line, but may
display a wide range of anticancer properties, as evidenced by the
phenotypic changes observed in the cell lines analyzed in the
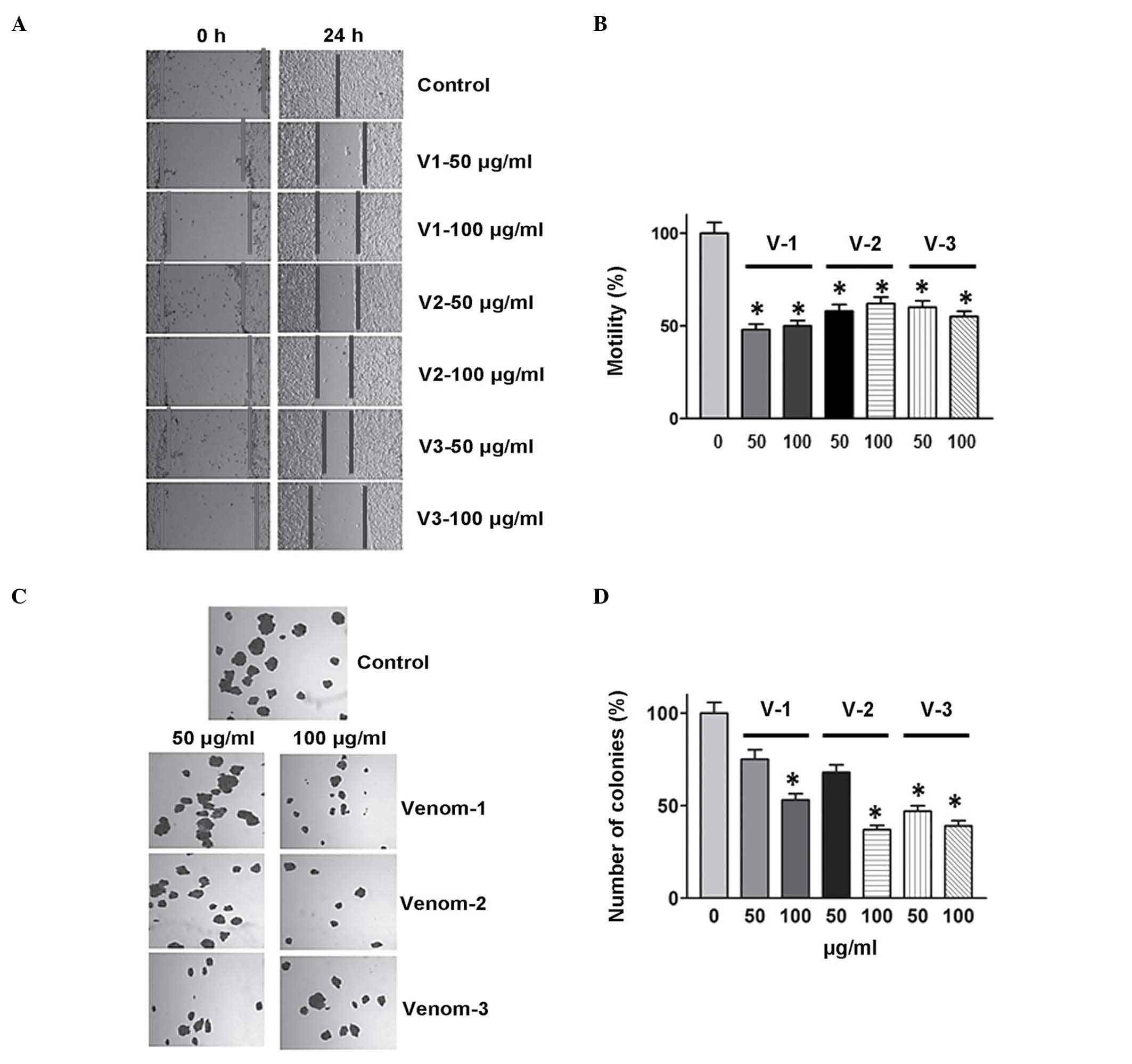
present study. Furthermore, venom obtained from Androctonus bicolor
and Leiurus quinquestriatus was used against the MDA-MB-231 cell
line, and a similar inhibitory effect was observed on colony
formation and cell motility (Fig. 4).
These findings suggest a potential novel area of research aimed to
elucidate the molecular mechanism responsible for the anticancer
activities of scorpion venom.
Discussion
Scorpion venom is a chemically enriched substance,
containing various proteins and peptides, which may act as an
anticancer agent. Scorpion venom exhibits anticancer potential via
the inhibition of cell proliferation and growth in vitro by
interfering with the activity of various signaling molecules such
as phosphatase and tensin homolog, which are involved in a number
of signal transduction cascades (38–40). The
present study evaluated the effects of scorpion venom on cell
motility and colony formation in two well-established and
characterized colorectal cancer cell lines, HCT-8 and HCT-116, in
addition to the breast cancer cell line MDA-MB-231. A marked
reduction in cell motility and colony formation was observed in a
dose-dependent manner in the above colorectal cancer cell lines.
Due to the limited availability of venom, the median concentration
of venom-1 that was used to treat HCT-8 cells was selected to
evaluate the effects of venom-1 in the HCT-116 cell line, and the
same inhibitory effect was observed in cell motility and
anchorage-dependent colony formation assays. In order to
investigate whether scorpion venom exhibited a broad range of
anticancer activity rather than being specific only for colorectal
cancer cells, a breast cancer cell line, namely MDA-MB-231, was
additionally studied. A perceptive decrease in cell motility and
colony formation was also observed in these cell line, although
within minor experimental variation, indicating that venom-1 also
exhibited an inhibitory effect on breast cancer cells. Minor
fluctuations in the outcome of the treatments may be attributed to
the differential uptake of venom-1 in the different experimental
setups. Nevertheless, the results demonstrated that venom-1 acted
as an inhibitor of cancer cell motility. Furthermore, a similar
phenotypic inhibitory effect was observed following treatment with
venoms obtained from Androctonus bicolor (venom-2) and
Leiurus quinquestriatus (venom-3). Cell motility and colony
formation are two important factors that are widely used in cancer
research to evaluate the effectiveness of anticancer drugs or
micro-RNAs on cancer progression (41,42). Focal
adhesion kinase (FAK) is an important molecule for focal assembly
and contraction of cells, which facilitates movement and invasion
into the ECM. The activation of FAK is necessary to achieve
adhesion, followed by movement of cells in various directions and
enhancement of cell motility (43).
Reduced cell motility in the presence of scorpion venom, as
demonstrated in the present study, may be a manifestation of loss
of active FAK due to the action of the venom, which may instigate a
signaling cascade leading to phenotypic changes via the retardation
of normal cell activities. Similarly, clonogenic assay is a
representation of cell proliferation, and assesses the survival of
a single cell, which may form colonies to ensure its growth and
survival (37). As presented in the
current study, a marked decrease in colony formation following
treatment with venom was an additional indication of the inhibition
of cancer cell proliferation in the presence of the three scorpion
venoms analyzed.
The findings of the present study suggest the
identification of a novel and productive area of future research
aimed to elucidate the underlying molecular mechanism that enables
scorpion venom to inhibit cell motility and colony formation in
cancer cell lines. Two possible mechanisms may be proposed to
explain this reduction in cell motility and colony formation: i) A
decrease in the expression of MMPs, and ii) a reduction in the
phosphorylation levels of FAK, which is involved in cell migration
and invasion. However, these hypothetical mechanisms will require
future investigation.
In conclusion, the present study demonstrated that
scorpion venom is able to act as an anticancer agent by decreasing
cell motility and colony formation in colorectal and breast cancer
cell lines. The results of the present study add to the increasing
body of evidence supporting the potential of scorpion venom to be
used as an anticancer agent, and indicate the requirement for
molecular analysis of the action of the underlying signaling
mechanisms. Therefore, scorpion venom may represent a valuable
therapeutic tool to be used as part of the therapeutic regimen for
the treatment of patients with colorectal and breast cancer.
Acknowledgments
The authors would like to thank the Research Center
of Prince Sultan Military Medical City Hospital and King Abdulaziz
City for Science and Technology (KACST) for providing the necessary
facilities and financial support. The authors would also like to
thank Dr Bandar Al-Knawy, Director of the Research Center of King
Saud Bin Abdulaziz Medical City Hospital (Riyadh, Saudi Arabia) for
providing the colorectal and breast cancer cell lines used in the
present study.
References
|
1
|
Al-Asmari AK and Al-Saif AA: Scorpion
sting syndrome in a general hospital in Saudi Arabia. Saudi Med J.
25:64–70. 2004.PubMed/NCBI
|
|
2
|
Mahaba HM: ElS ayed SA: Scorpion sting: Is
it a health problem in Saudi Arabia? Evaluation of management of
820 cases. Saudi Med J. 17:15–21. 1996.
|
|
3
|
Mahaba HM: Scorpion sting syndrome:
Epidemiology clinical presentations and management of 2240 cases.
East Mediterr Health J. 3:82–89. 1997.
|
|
4
|
Simard MJ and Watt DD: Venoms and toxins.
The Biology of Scorpions. Polis GA: X:(Yth). (Redwood City, CA).
Stanford University Press. 414–444. 1990.
|
|
5
|
Al Asmari AK, Khan HA and Manthiri RA: AlY
ahya KM and Al Otaibi KE: Effects of Echis pyramidum snake
venom on hepatic and renal antioxidant enzymes and lipid
peroxidation in rats. J Biochem Mol Toxicol. 28:407–412. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Al Asmari AK, Al Zahrani AG, Al Jowhary S
and Arshaduddin M: Clinical aspects and frequency of scorpion
stings in the Riyadh region of Saudi Arabia. Saudi Med J.
33:852–858. 2012.PubMed/NCBI
|
|
7
|
Ozkan O and Filazi A: The determination of
acute lethal dose-50 (LD50) levels of venom in mice obtained by
different methods from scorpions, Androctonus crassicauda
(Oliver 1807). Turkiye Parazitol Derg. 28:50–53. 2004.
|
|
8
|
Bawaskar HS and Bawaskar PH: Scorpion
sting: Update. J Assoc Physicians India. 60:46–55. 2012.PubMed/NCBI
|
|
9
|
Petricevich VL: Effect of Tityus
serrulatus venom on cytokine production and the activity of
murine macrophages. Mediators Inflamm. 11:23–31. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rochat H, Bernard P and Couraud F:
Scorpion toxins: Chemistry and mode of action. Advances in
Cytopharmacology, Neurotoxins Tools in Neurobiology. Ceccarelli B
and Clementi F: 3:(Yth). (New York, NY). Raven. 325–334. 1979.
|
|
11
|
Andreotti N, Jouirou B, Mouhat S, Mouhat L
and Sabatier JM: Therapeutic value of peptides from animal Venoms.
Amino Acids, Peptides and Proteins, Comprehensive Natural Products
II Chemistry and Biology. Mander L and Lui HD: X:(Yth). (Oxford).
Elsevier. 287–303. 2010.
|
|
12
|
Ahn MY, Ryu KS, Lee YW and Kim YS:
Cytotoxicity and L-amino acid oxidase activity of crude insect
drugs. Arch Pharm Res. 23:477–481. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Z, Hong W, Zeng Z, Wu Y, Hu K, Tian
X, Li W and Cao Z: Mucroporin-M1 inhibits hepatitis B virus
replication by activating the mitogen-activated protein kinase
(MAPK) pathway and down-regulating HNF4-α in vitro and in
vivo. J Biol Chem. 287:30181–30190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen Y, Cao L, Zhong M, Zhang Y, Han C, Li
Q, Yang J, Zhou D, Shi W, He B, et al: Anti-HIV-1 activity of a new
scorpion venom peptide derivative Kn2-7. PLoS One. 7:e349472012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
El-Ghlban S, Kasai T, Shigehiro T, Yin HX,
Sekhar S, Ida M, Sanchez A, Mizutani A, Kudoh T, Murakami H and
Seno M: Chlorotoxin-Fc fusion inhibits release of MMP-2 from
pancreatic cancer cells. Biomed Res Int. 2014:1526592014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gupta SD and Gomes A, Debnath A, Saha A
and Gomes A: Apoptosis induction in human leukemic cells by a novel
protein Bengalin, isolated from Indian black scorpion venom,
Through mitochondrial pathway and inhibition of heat shock
proteins. Chem Biol Interact. 183:293–303. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ding J, Chua PJ, Bay BH and
Gopalakrishnakone P: Scorpion venoms as a potential source of novel
cancer therapeutic compounds. Exp Biol Med (Maywood). 239:387–393.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Colorectal Cancer Facts & Figures
2014-2016. Atlanta: American Cancer Society Inc. 2014.
|
|
19
|
Breast Cancer Facts & Figures
2013-2014. Atlanta: American Cancer Society, Inc. 2013.
|
|
20
|
Mosli MH and Al-Ahwal MS: Colorectal
cancer in the Kingdom of Saudi Arabia: Need for screening. Asian
Pac J Cancer Prev. 13:3809–3813. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alghamdi IG, Hussain II, Alghamdi MS and
El-Sheemy MA: The incidence rate of female breast cancer in Saudi
Arabia. An observational descriptive epidemiological analysis of
data from Saudi Cancer Registry 2001-2008. Breast Cancer (Dove Med
Press). 5:103–109. 2013.PubMed/NCBI
|
|
22
|
Folkman J: Angiogenesis: I nitiation and
control. Ann N Y Acad Sci. 401:212–227. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis. An imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sato H, Takino T, Okada Y, Cao J,
Shinagawa A, Yamamoto E and Seiki M: A matrix metalloproteinase
expressed on the surface of invasive tumour cells. Nature.
370:61–65. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Itoh Y and Seiki M: MT1-MMP: A potent
modifier of pericellular microenvironment. J Cell Physiol. 206:1–8.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gross J and Lapiere CM: Collagenolytic
activity in amphibian tissues: A tissue culture assay. Proc Natl
Acad Sci USA. 48:1014–1022. 1962. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gross J and Nagai Y: Specific degradation
of the collagen molecule by tadpole collagenolytic enzyme. Proc
Natl Acad Sci USA. 54:1197–1204. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Van Lint P and Libert C: Chemokine and
cytokine processing by matrix metalloproteinases and its effect on
leukocyte migration and inflammation. J Leukoc Biol. 82:1375–1381.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Deshane J, Garner CC and Sontheimer H:
Chlorotoxin inhibits glioma cell invasion via matrix
metalloproteinase-2. J Biol Chem. 278:4135–4144. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Soroceanu L, Gillespie Y, Khazaeli MB and
Sontheimer H: Use of chlorotoxin for targeting of primary brain
tumors. Cancer Res. 58:4871–4879. 1998.PubMed/NCBI
|
|
31
|
Lyons SA, O'Neal J and Sontheimer H:
Chlorotoxin, a scorpion-derived peptide, specifically binds to
gliomas and tumors of neuroectodermal origin. Glia. 39:162–173.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qin C, He B, Dai W, Zhang H, Wang X, Wang
J, Zhang X, Wang G, Yin L and Zhang Q: Inhibition of metastatic
tumor growth and metastasis via targeting metastatic breast cancer
by chlorotoxin-modified liposomes. Mol Pharm. 11:3233–3241. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zucker S and Vacirca J: Role of matrix
metalloproteinases (MMPs) in colorectal cancer. Cancer Metastasis
Rev. 23:101–117. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vermeulen SJ, Chen TR, Speleman F and
Nollet F: VanR oy FM and Mareel MM: Did the four human cancer cell
lines DLD-1, HCT-15, HCT-8 and HRT-18 originate from one and the
same patient? Cancer Genet Cytogenet. 107:76–79. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Crowley-Weber CL, Payne CM, Gleason-Guzman
M, Watts GS, Futscher B, Waltmire CN, Crowley C, Dvorakova K,
Bernstein C, Craven M, et al: Development and molecular
characterization of HCT-116 cell lines resistant to the tumor
promoter and multiple stress-inducer, deoxycholate. Carcinogenesis.
23:2063–2080. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cailleau R, Young R, Olivé M and Reeves WJ
Jr: Breast tumor cell lines from pleural effusions. J Natl Cancer
Inst. 53:661–674. 1974.PubMed/NCBI
|
|
37
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat
Protoc. 1:2315–2319. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Omran MAA: In vitro anticancer
effect of scorpion Leiurus quinquestriatus and Egyptian
cobra venom on human breast and prostate cancer cell lines. Res J
Med Sci. 3:66–86. 2003.
|
|
39
|
Fu YJ, Yin LT, Liang AH, Zhang CF, Wang W,
Chai BF, Yang JY and Fan XJ: Therapeutic potential of
chlorotoxin-like neurotoxin from the Chinese scorpion for human
gliomas. Neurosci Lett. 412:62–67. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gao F, Li H, Chen YD, Yu XN, Wang R and
Chen XL: Upregulation of PTEN involved in scorpion venom-induced
apoptosis in a lymphoma cell line. Leuk Lymphoma. 50:633–641. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Islam M, Datta J, Lang JC and Teknos TN:
Down regulation of RhoC by microRNA-138 results in de-activation of
FAK, Src and Erk1/2 signaling pathway in head and neck squamous
cell carcinoma. Oral Oncol. 50:448–456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Islam M, Sharma S, Kumar B and Teknos TN:
Atorvastatin inhibits RhoC function and limits head and neck cancer
metastasis. Oral Oncol. 49:778–786. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Schlaepfer DD and Mitra SK: Multiple
connections link FAK to cell motility and invasion. Curr Opin Genet
Dev. 14:92–101. 2004. View Article : Google Scholar : PubMed/NCBI
|