Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common type of cancer worldwide, and the incidence of hepatitis B
virus (HBV)-related primary HCC is 55 cases/100,000 individuals for
each gender (1). HCC can be diagnosed
by non-invasive imaging, including computed tomography or magnetic
resonance imaging. At present, there are multiple therapeutic
methods available for HCC, which depends on the stage of HCC,
however, surgical treatment is the most common method. Despite
this, the prognosis for HCC is poor (2). HBV is an important pathogenic factor for
HCC (3). Hippo is a highly-conserved
signaling pathway that has been identified in both drosophila and
mammals (4). Yes-associated protein 1
(YAP) is a transcription factor in the Hippo signaling pathway
(5), which regulates a number of
transcription factors, including erb-b2 receptor tyrosine kinase 4,
runt related transcription factor 2, p73, and TEA domain
transcription factor 1 (6–9). Furthermore, YAP promotes the
proliferation of liver cancer in mice by regulating the
transcription of certain target genes, including Ki-67, c-myc,
SRY-Box 4 (SOX4), H19 and α-fetoprotein (AFP) (10). In addition, transgenic mice
overexpressing YAP exhibited an increased liver size and eventually
developed liver cancer (11). It has
also been demonstrated that YAP overexpression induces
epithelium-mesothelium conversion and anchorage-independent growth
in mammary epithelial MCF10A cells (12). A recent study in HCC revealed that YAP
overexpression overcame cell contact inhibition and promoted cell
growth (13). In addition, the
expression and nuclear accumulation of YAP was elevated in
prostate, large intestine, breast, esophageal, ovarian and liver
cancer (14–16). Clinical studies in HCC have
demonstrated that YAP is an independent predictive factor of poor
prognosis and overall survival (16).
YAP is involved in the development of HCC, indicating that YAP
exhibits an extremely important function in HBV-associated HCC
(17). However, the mechanism of
HBV-induced YAP upregulation remains unclear. In four HBV encoded
proteins, HBV X protein (HBx) acts as a multifunctional regulatory
protein, which exhibits an important function in HBV-induced HCC
(18). Although HBx does not combine
directly with DNA, HBx interacts with nuclear transcription
factors, including activator protein-1, nuclear factor-κB,
specificity protein-1 and cyclic adenosine monophosphate response
element-binding protein to regulate transcriptional activity,
subsequently affecting the regulation of intracellular signal
transduction pathways (19,20). Therefore, it is hypothesized that HBx
may be closely associated with YAP upregulation.
Subjects and methods
Patient samples
A total of 20 HCC tissues and corresponding adjacent
non-tumor liver tissues were obtained immediately after surgical
resection, performed in Nanyang City Central Hospital (Nanyang,
China) between March 2013 and March 2015. Patient's clinical and
pathological information was obtained from case records. All
patients were known to be HBV-positive. All liver tumor tissue
samples were confirmed as primary HCC by at least two pathologists,
according to the Barcelona Clinic Liver Cancer staging system
(21). No patients had received
radiotherapy or chemotherapy prior to surgery. A total of 5 normal
liver tissues were obtained from trauma patients during resection.
This study was conducted in accordance with the Declaration of
Helsinki. This study was conducted with approval from the Ethics
Committee of Nanyang City Central Hospital and written informed
consent was obtained from all patients.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total cellular RNA was extracted using Trizol
reagent (Takara, Dalian, China) according to the manufacturer's
instructions. RevertAid First Strand cDNA Synthesis Kit (MBI
Fermentas, St. Leon-Rot, Germany) was used for cDNA synthesis and
PCR amplification. DyNAmo™ Flash SYBR® Green qPCR kit
(Finnzymes Oy, Espoo, Finland) was used for qPCR. The primer
sequences used for PCR were as follows: Yap forward,
5′-GGGTGTTCATCCATTCTC-3′ and reverse, 5′-CCCAGCATCTTGTGTTTC-3′; HBx
forward, 5′-GTGGGATGATGACGACG-3′ and reverse,
5′-TACGACCAGAGGCATACAGG-3′; and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) forward, 5′-GCGGGAAATCGTGCGTGAC-3′ and
reverse, CGTCATACTCCTGCTTGCTG-3′. The reaction conditions were
pre-degeneration at 94°C for 30 sec, degeneration at 94°C for 5
sec, annealing at 94°C for 30 sec and extension at 72°C for 40 sec,
for 35 cycles, followed by terminal extension for 5 min at 72°C.
PCR was performed according to a previously described protocol
(22). The expression levels were
calculated and normalized to GAPDH.
Western blot analysis
The liver tissue was homogenized with liquid
nitrogen. Cells were lysed using cell lysis solution [0.3% NP40, 1
mM EDTA, 50 mM Tris-Cl (pH 7.4), 2 mM EDTA, 1% Triton X-100, 150 mM
NaCl, 25 mM NaF, 1 mM Na3VO3, 10 µg PMSF].
The experiment was conducted as described previously (23). The membranes were incubated with
monoclonal mouse YAP (1:100 dilution; catalog no. sc-15407; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and rabbit
polyclonal HBx (1:200 dilution; catalog no. ab39716; Abcam,
Cambridge, UK) primary antibodies, and β-actin mouse monoclonal
antibody (1:200 dilution; catalog no., sc-47778; Santa Cruz
Biotechnology, Inc.) overnight at 4°C. The membranes were then
washed with phosphate-buffered saline, followed by incubation with
goat anti-mouse immunoglobulin G secondary antibody (1:2,000
dilution; catalog no. BA1031; Boster Inc., Wuhan, China) at room
temperature for 1 h, and further washing 3 times. Next, the
membranes were incubated with enhanced chemiluminescence colored
liquid (Boster Inc.). The expression of YAP, HBx and β-actin was
quantified using Image J software (version 1.38; National
Institutes of Health, Bethesda, MD, USA). The experiment was
performed in triplicate.
Immunohistochemical analysis
For routine immunohistochemical analysis, liver
tissue was resected, embedded in paraffin and cut into 5-µm thick
sections. After the sections were dewaxed and hydrated, the
endogenous catalase was removed with 3% hydrogen peroxide. The
primary and secondary antibodies used were those applied in the
western blotting, with identical incubation times and temperatures.
The sections were incubated with DAB, stained with hematoxylin and
observed under an optical microscope (Olympus AX80; Olympus
Corporation, Tokyo, Japan). YAP immunoreactivity was classified
according to the following staining scores: Negative, 0; and
positive, 1–3; as described previously (16)
Statistical analysis
All data were analyzed using SPSS 17.0 statistical
software (SPSS Inc., Chicago, IL, USA). Student's t-test was
conducted and the data presented as the mean ± standard deviation.
Pearson's correlation coefficient was used to analyze the
association between YAP and HBx mRNA expression. The Wilcoxon
signed rank test was used to compare YAP expression in tumor tissue
and matched adjacent non-tumor tissue. P<0.05 was considered to
indicate a statistically significant difference.
Results
YAP and HBx mRNA expression are
positively correlated in HBV-positive HCC tumor tissues
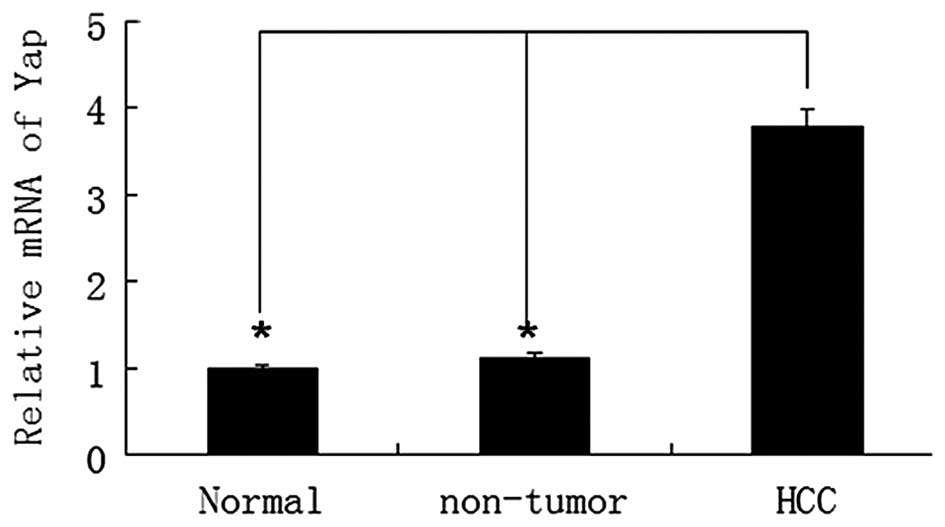
YAP mRNA expression was analyzed in 20 paired HCC
and adjacent non-tumor liver tissues. The results revealed that YAP
mRNA levels in HBV-associated HCC tissues were significantly higher
than that of non-tumor liver tissues (P<0.05) (Fig. 1). According to a previous study
(24), RT-qPCR identified HBx mRNA in
HCC tissue. The present study demonstrated that HBx mRNA expression
was identified in all HBV-infected clinical samples by RT-qPCR.
Notably, upregulation of YAP was significantly correlated with HBx
expression (P<0.05; r=0.719) (Fig.
2). Western blot analysis of YAP expression in HBx-infected
liver tissue revealed that HBx expression was positively correlated
with YAP upregulation.
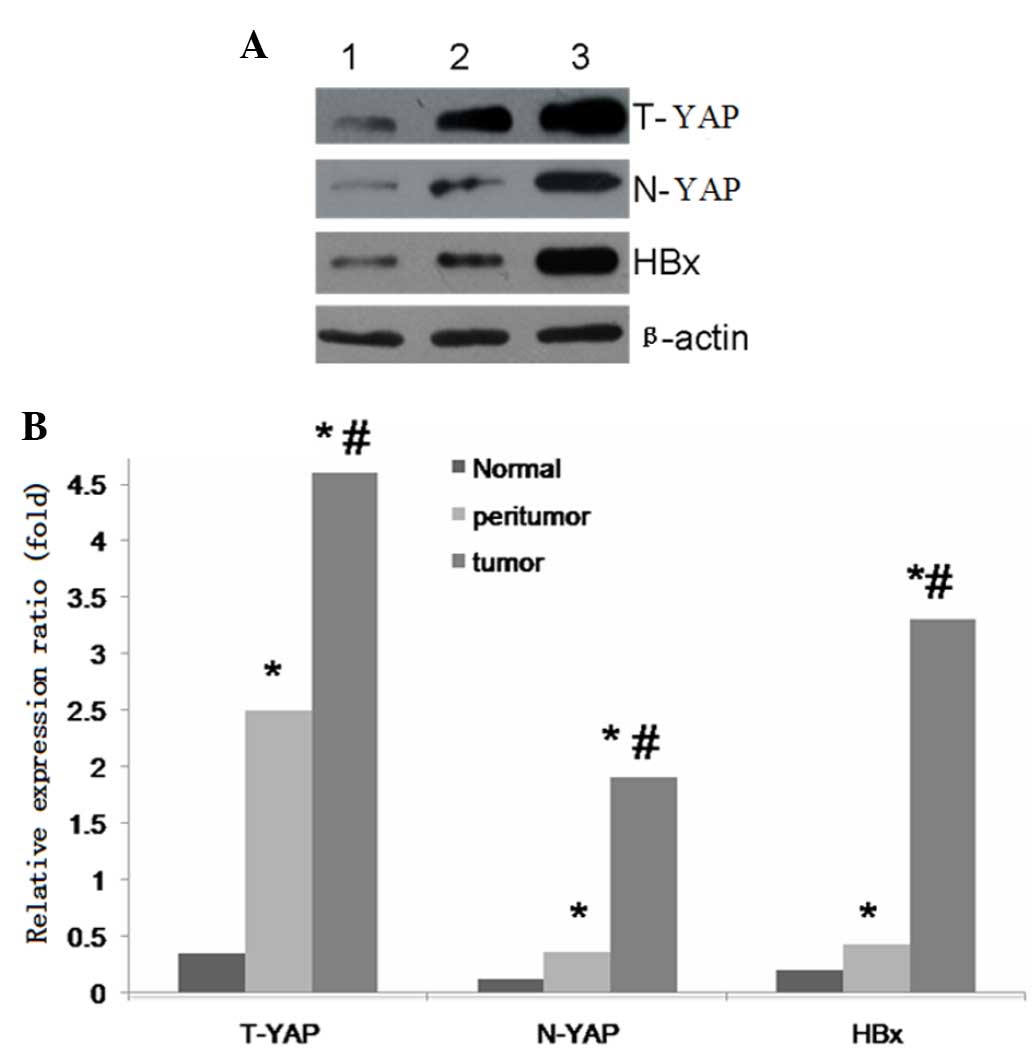 | Figure 2.(A) Western blot analysis of T-YAP,
N-YAP, HBx and β-actin expression. Lanes 1, 2 and 3 indicate
normal, peritumor and HCC tissue, respectively. (B) Relative
protein expression of total Yap, nuclear Yap and HBx. *P<0.01
for tumor versus peritumoral or normal tissue;
#P<0.01 for tumor versus peritumoral tissue for
T-YAP, N-YAP and HBx protein. T-YAP, total yes-associated protein
1; N-YAP, nuclear YAP; HBx, hepatitis B virus × protein. |
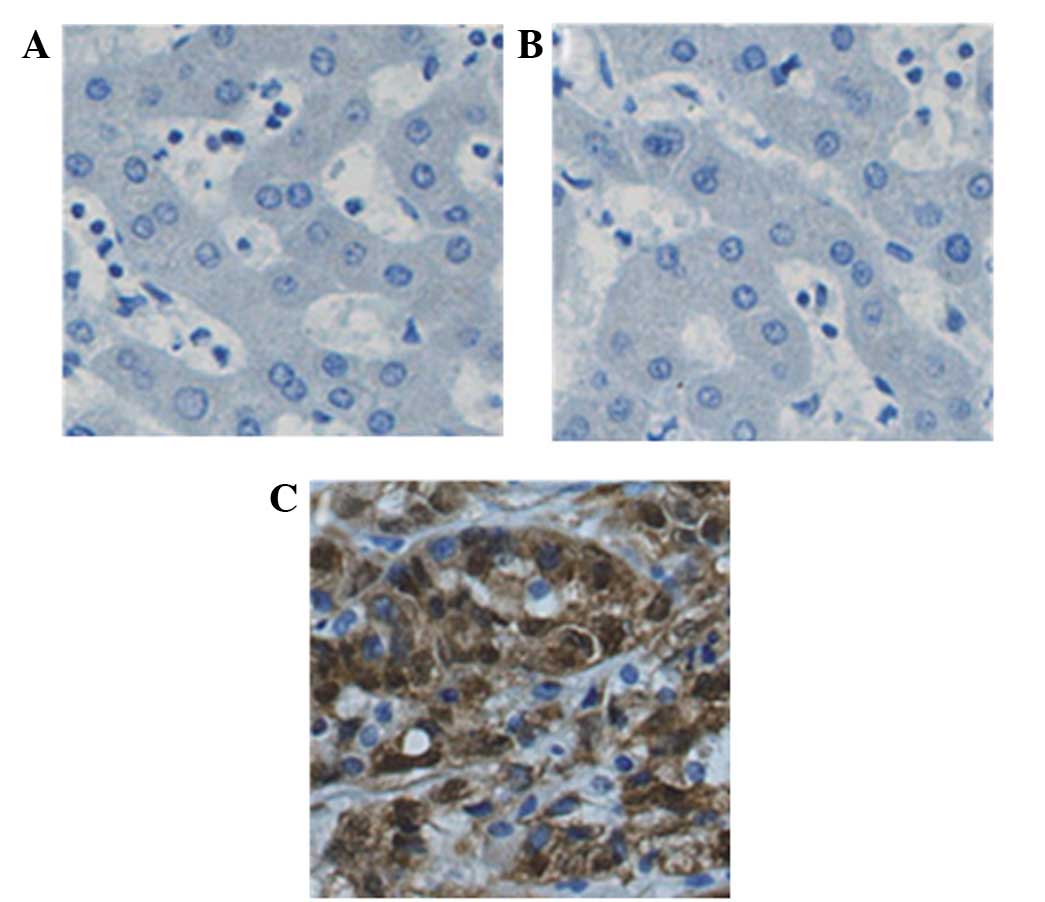
HCC tissues exhibit strong nuclear YAP
expression
YAP immunoreactivity was classified according to the
following staining scores: Negative, 0; and positive, 1–3; as
described previously (16). The
results confirmed that the expression of YAP was increased in 70%
of HCC tissues in comparison to adjacent HCC tissues. A total of
40.0% (8/20), 15% (3/20) and 10% (2/20) of HCC tissues exhibited
strong, moderate and low nuclear staining, respectively. The rest
of the HCC samples exhibited negative YAP nuclear staining. In HCC
tissues, cytoplasmic localization of YAP with strong and moderate
immunoreactivity was observed in 35 (7/20) and 25% (5/20) of
tissues, respectively. The rest HCC sample showed weak and negative
YAP intracellular immunoreactivities. Positive cytoplasmic YAP
expression was identified in adjacent non-tumor and normal liver
tissues (Fig. 3). Strong YAP staining
was observed in HCC cell nuclei, but not in non-tumor cell
nuclei.
Discussion
HBx exhibits an important function in HBV-induced
hepatocellular carcinoma (18). It
has been reported that HBx regulates various genes (25). HBx involves numerous functions,
including transcription regulation, signal transduction, cell cycle
progression, protein degradation pathways, apoptosis and the
interaction between different transcription factors and signal
transduction pathway components (19,20,24,26).
YAP is an important transcription factor that promotes the
proliferation of liver cancer in mice by regulating the
transcription of certain target genes, including Ki-67, c-myc,
SOX4, H19 and AFP (18). In the
present study, YAP expression in 20 primary HCC, 20 adjacent
non-tumor liver tissues and 5 normal HBV-negative liver tissues
were analyzed by RT-qPCR and western blot analysis. YAP expression
was increased in the cell nucleus of primary HCC liver tissue when
compared with non-tumor tissues and was positively correlated with
HBx expression. Immunohistochemical staining also revealed that YAP
was strongly expressed in HBx-induced HCC liver tissue, however,
YAP expression was negative or weak in adjacent non-tumor tissue
and normal liver tissue, indicating that YAP and HBx expression are
positively correlated. We hypothesized that although HBx does not
directly combine with DNA, it induces the transcription factor,
YAP, in the Hippo pathway to shuttle and translocate to the
nucleus. Additionally, YAP forms complexes with other factors to
regulate the expression of target genes, such as the AFP protein
(18,27). Zhao et al (13) reported that 54% of 115 HCC patients
exhibited YAP overexpression in the United States and Xu et
al (16) reported that 62% of 117
HCC patients exhibited YAP overexpression in Hong Kong.
Furthermore, YAP was predominantly located in the cell nuclei of
liver cancer cells and YAP staining was weak in non-tumor tissues
(13,16), which was consistent with the results
of the present study. Notably, nuclear YAP expression has also been
identified in other tumors, including prostate, colorectal, breast,
ovarian and esophageal cancer (13,16).
Kaposi's sarcoma-associated herpes virus is an oncogenic virus. The
genome-encoded viral G protein-coupled receptor inhibits kinase
Last1/2 in the Hippo signaling pathway via Gq/11 and G12/13, which
promotes YAP activation leading to cell proliferation and
transformation (28). This indicates
that certain tumor viruses lead to cell transformation via the
Hippo signaling pathway. Therefore, YAP inhibitors may present can
potential antiviral or anticancer drug candidates.
In conclusion, YAP is a critical oncogenic protein
that is involved in human cancer. The present study revealed that
HBx expression upregulates YAP in HCC cells. Therefore, HBx
upregulates YAP in liver cancer cells and may contribute to the
development of HBV infection-associated HCC, indicating that YAP is
a key driver gene of HBx-induced liver cancer. Thus, YAP may
present a novel target for the treatment of HBV-associated HCC.
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Han YF, Zhao J, Ma LY, Yin JH, Chang WJ,
Zhang HW and Cao GW: Factors predicting occurrence and prognosis of
hepatitis-B-virus-related hepatocellular carcinoma. World J
Gastroenterol. 17:4258–4270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Engelke M, Mills K, Seitz S, Simon P,
Gripon P, Schnölzer M and Urban S: Characterization of a hepatitis
B and hepatitis delta virus receptor binding site. Hepatology.
43:750–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu L, Li Y, Kim SM, Bossuyt W, Liu P, Qiu
Q, Wang Y, Halder G, Finegold MJ, Lee JS and Johnson RL: Hippo
signaling is a potent in vivo growth and tumor suppressor pathway
in the mammalian liver. Proc Natl Acad Sci USA. 107:1437–1442.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu AM, Xu MZ, Chen J, Poon RT and Luk JM:
Targeting YAP and Hippo signaling pathway in liver cancer. Expert
Opin Ther Targets. 14:855–868. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Komuro A, Nagai M, Navin NE and Sudol M:
WW domain-containing protein YAP associates with ErbB-4 and acts as
a co-transcriptional activator for the carboxyl-terminal fragment
of ErbB-4 that translocates to the nucleus. J Biol Chem.
278:33334–33341. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zaidi SK, Sullivan AJ, Medina R, Ito Y,
van Wijnen AJ, Stein JL, Lian JB and Stein GS: Tyrosine
phosphorylation controls Runx2-mediated subnuclear targeting of YAP
to repress transcription. EMBO J. 23:790–799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lapi E, Di Agostino S, Donzelli S, Gal H,
Domany E, Rechavi G, Pandolfi PP, Givol D, Strano S, Lu X and
Blandino G: PML, YAP and p73 are components of a proapoptotic
autoregulatory feedback loop. Mol Cell. 32:803–814. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao B, Ye X, Yu J, Li L, Li W, Li S, Yu
J, Lin JD, Wang CY, Chinnaiyan AM, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong J, Feldmann G, Huang J, Wu S, Zhang
N, Comerford SA, Gayyed MF, Anders RA, Maitra A and Pan D:
Elucidation of a universal size-control mechanism in Drosophila and
mammals. Cell. 130:1120–1133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kowalik MA, Saliba C, Pibiri M, Perra A,
Ledda-Columbano GM, Sarotto I, Ghiso E, Giordano S and Columbano A:
Yes-associated protein regulation of adaptive liver enlargement and
hepatocellular carcinoma development in mice. Hepatology.
53:2086–2096. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Overholtzer M, Zhang J, Smolen GA, Muir B,
Li W, Sgroi DC, Deng CX, Brugge JS and Haber DA: Transforming
properties of YAP, a candidate oncogene on the chromosome 11q22
amplicon. Proc Natl Acad Sci USA. 103:12405–12410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Steinhardt AA, Gayyed MF, Klein AP, Dong
J, Maitra A, Pan D, Montgomery EA and Anders RA: Expression of
Yes-associated protein in common solid tumors. Hum Pathol.
39:1582–1589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Muramatsu T, Imoto I, Matsui T, Kozaki K,
Haruki S, Sudol M, Shimada Y, Tsuda H, Kawano T and Inazawa J: YAP
is a candidate oncogene for esophageal squamous cell carcinoma.
Carcinogenesis. 32:389–398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang T, Zhang J, You X, Liu Q, Du Y, Gao
Y, Shan C, Kong G, Wang Y, Yang X, Ye L and Zhang X: Hepatitis B
virus X protein modulates oncogene Yes-associated protein by CREB
to promote growth of hepatoma cells. Hepatology. 56:2051–2059.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang H, Oishi N, Kaneko S and Murakami S:
Molecular functions and biological roles of hepatitis B virus ×
protein. Cancer Sci. 97:977–983. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ng SA and Lee C: Hepatitis B virus X gene
and hepatocarcinogenesis. J Gastroenterol. 46:974–990. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Zhang H and Ye L: Effects of
hepatitis B virus X protein on the development of liver cancer. J
Lab Clin Med. 147:58–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Llovet JM, Di Bisceglie AM, Bruix J,
Kramer BS, Lencioni R, Zhu AX, Sherman M, Schwartz M, Lotze M,
Talwalkar J and Gores GJ: Panel of Experts in HCC-Design Clinical
Trials: Design and endpoints of clinical trials in hepatocellular
carcinoma. J Natl Cancer Inst. 100:698–711. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ji JF and Ma XH: Effect of baculovirus P35
protein on apoptosis in brain tissue of rats with acute cerebral
infarction. Genet Mol Res. 14:9353–9360. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang JH, Li XJ, Chai SJ and Wang XY:
Neuroprotective effect and its mechanism of lentivirus mediated
VEGF on rat model with cerebral ischemic injury. Int J Clin Exp
Med. 8:4094–4100. 2015.PubMed/NCBI
|
|
24
|
Wang Y, Lu Y, Toh ST, Sung WK, Tan P, Chow
P, Chung AY, Jooi LL and Lee CG: Lethal-7 is down-regulated by the
hepatitis B virus × protein and targets signal transducer and
activator of transcription 3. J Hepatol. 53:57–66. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen JY, Chen YJ, Yen CJ, Chen WS and
Huang WC: HBx sensitizes hepatocellular carcinoma cells to
lapatinib by up-regulating ErbB3. Oncotarget. 7:473–489.
2016.PubMed/NCBI
|
|
26
|
Sung WK, Lu Y, Lee CW, Zhang D, Ronaghi M
and Lee CG: Deregulated direct targets of the hepatitis B virus
(HBV) protein, HBx, identified through chromatin
immunoprecipitation and expression microarray profiling. J Biol
Chem. 284:21941–21954. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou D, Conrad C, Xia F, Park JS, Payer B,
Yin Y, Lauwers GY, Thasler W, Lee JT, Avruch J and Bardeesy N: Mst1
and Mst2 maintain hepatocyte quiescence and suppress hepatocellular
carcinoma development through inactivation of the Yap1 oncogene.
Cancer Cell. 16:425–438. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu G, Yu FX, Kim YC, Meng Z, Naipauer J,
Looney DJ, Liu X, Gutkind JS, Mesri EA and Guan KL: Kaposi
sarcoma-associated herpesvirus promotes tumorigenesis by modulating
the Hippo pathway. Oncogene. 34:3536–3546. 2015. View Article : Google Scholar : PubMed/NCBI
|