Introduction
Chordoma is a rare malignant bone tumor with an
incidence of ~0.1/100,000 individuals (1). It is thought to originate from
notochordal remnants and is usually localized to the skull base,
vertebral column and sacrum. Despite improvements in therapeutic
modalities, the treatment of chordoma remains challenging because
of its location and local recurrence (2). Therefore, understanding the molecular
mechanisms underlying the development and progression of chordoma
is crucial to identify novel therapeutic targets.
Brachyury, which is a transcription factor encoded
by the T gene, is specifically expressed in chordomas and in the
notochord (3). T gene duplication is
common in familial chordoma, while gain of the T locus has also
been found in sporadic cases (4,5). Silencing
of brachyury using short hairpin RNA has been shown to result in
decreased cell proliferation and induction of a senescent phenotype
(5,6).
It is well-documented that brachyury orchestrates several
downstream pathways in chordoma; however, the mechanisms underlying
brachyury regulation remain unclear (7).
MicroRNAs (miRNAs or miRs) are a class of small,
non-coding, single-stranded RNAs that can regulate the process of
translation by binding to the 3′-untranslated region (UTR) of
mature mRNAs. Importantly, miRNAs have been shown to be involved in
tumorigenesis, angiogenesis, invasion and metastasis (8–10).
Furthermore, miRNAs may served as oncogenes or tumor suppressors,
and have therefore been reported to be dysregulated in various
human cancers, including gastric cancer (11), colorectal cancer (12), thyroid cancer (13) and squamous cell carcinoma (14). However, a role for miRNAs in the
regulation of brachyury expression in chordoma has yet to be
elucidated.
In the present study, it was first demonstrated that
miR-219-5p was downregulated in chordoma tissue samples and the
U-CH2 cell line. Therefore, whether brachyury is a direct target of
miR-219-5p was evaluated. In vitro analyses showed that
miR-219-5p inhibited chordoma cell proliferation and clonogenicity
by downregulating brachyury. The associations between miR-219-5p
expression and various patient clinicopathological factors were
further analyzed and it was shown that miR-219-5p expression was
correlated with the tumor extent and recurrence.
Materials and methods
Patients and tissue samples
Tissue samples were obtained from 40 patients who
had undergone surgery at Xuanwu Hospital, Capital Medical
University (Beijing, China) between February 2008 and February
2013. Freshly received tissues from the operating room were
immediately frozen with liquid nitrogen. All patients had confirmed
pathological diagnosis according to the World Health Organization
classification (15). A total of 40
skull base chordoma samples s and 10 non-neoplastic adjacent tissue
samples (normal skeletal muscles) were used in accordance with the
policies of the hospital's institutional review board. Written
informed consent was obtained prior to initiation of the study from
all patients. Medical records were reviewed to obtain clinical
information for each chordoma case, including age, gender, tumor
site, surgical approach and recurrence. The clinical findings of
the 40 patients are listed in Table
I.
 | Table I.Clinical features of chordoma patients
and relative expression levels of miR-219-5p. |
Table I.
Clinical features of chordoma patients
and relative expression levels of miR-219-5p.
| N | Gender | Age (years) | Tumor site | Surgical
approach |
Initial/recurrent | miR |
|---|
| 1 | F | 25 | Clivus and CV | TA | Recurrent | 0.451 |
| 2 | M | 25 | Clivus | EEA | Recurrent | 0.656 |
| 3 | F | 41 | Clivus | EEA | Recurrent | 0.600 |
| 4 | F | 13 | Clivus and CV | TA and OCF | Recurrent | 0.196 |
| 5 | M | 32 | Clivus and SR | TA | Initial | 0.960 |
| 6 | M | 62 | Clivus | EEA | Initial | 2.646 |
| 7 | F | 39 | Clivus | TA | Recurrent | 0.390 |
| 8 | F | 55 | Clivus | EEA | Initial | 0.210 |
| 9 | M | 34 | Clivus | EEA | Initial | 0.918 |
| 10 | M | 44 | Clivus | EEA | Initial | 6.881 |
| 11 | F | 52 | Clivus | EEA | Initial | 1.879 |
| 12 | M | 19 | Clivus | EEA | Initial | 0.554 |
| 13 | F | 15 | Clivus | TA | Initial | 0.230 |
| 14 | M | 50 | Petroclival
region | TA | Recurrent | 0.342 |
| 15 | M | 62 | Clivus | EEA | Initial | 1.332 |
| 16 | M | 43 | Clivus and ASB | EEA | Recurrent | 0.292 |
| 17 | M | 23 | Clivus | EEA | Initial | 1.609 |
| 18 | F | 31 | Clivus | TA | Recurrent | 2.154 |
| 19 | M | 42 | Petroclival
region | EEA | Recurrent | 0.662 |
| 20 | M | 33 | Clivus | EEA | Recurrent | 0.380 |
| 21 | F | 49 | Clivus | EEA | Recurrent | 0.898 |
| 22 | F | 32 | Clivus | EOA | Initial | 1.822 |
| 23 | M | 56 | Clivus | EEA | Initial | 1.589 |
| 24 | M | 51 | Clivus and SR | EEA | Recurrent | 0.820 |
| 25 | F | 59 | Clivus | EEA | Initial | 1.398 |
| 26 | F | 46 | Clivus and CV | EOA | Initial | 0.853 |
| 27 | M | 69 | Clivus and CV | EOA | Initial | 1.400 |
| 28 | F | 42 | Clivus and ASB | EEA | Recurrent | 0.143 |
| 29 | F | 16 | Clivus | EEA | Initial | 1.664 |
| 30 | F | 72 | Clivus and CV | EOA | Initial | 0.882 |
| 31 | F | 60 | Clivus | EEA | Recurrent | 1.526 |
| 32 | F | 66 | Clivus | EEA | Initial | 0.151 |
| 33 | M | 43 | Clivus | EEA | Recurrent | 1.058 |
| 34 | M | 22 | Clivus | EEA | Initial | 4.216 |
| 35 | M | 32 | Clivus | EEA | Initial | 1.211 |
| 36 | M | 42 | Clivus and CV | EOA and OCF | Initial | 0.189 |
| 37 | M | 47 | Clivus and ASB | EEA | Recurrent | 0.350 |
| 38 | F | 21 | Clivus | EEA | Initial | 2.359 |
| 39 | F | 28 | Clivus | EEA | Recurrent | 0.883 |
| 40 | M | 50 | Clivus | EEA | Initial | 0.418 |
Cell lines and reagents
The human chordoma cell line U-CH2 (ATCC, Manassas,
VA, USA) was cultivated in Iscove's Modified Dulbecco's modified
Eagle's medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
and RPMI-1640 medium (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA) at a ratio of 4:1 (vol/vol), supplemented with 10% fetal
bovine serum (FBS; Seromed Biochrom, Berlin, Germany). Culture
flasks were coated with rat tail type I collagen (BD Biosciences,
San Diego, CA, USA) prior to use. The human embryonic kidney 293T,
nasopharyngeal carcinoma CNE-2 and laryngeal carcinoma Hep-2 cell
lines were obtained from the Institute of Basic Medical Sciences of
the Chinese Academy of Medical Sciences (Beijing, China). 293T
cells were cultured in RPMI-1640 medium containing 10% FBS. CNE-2
and Hep-2 cells were cultured in Dulbecco's Modified Eagle's Medium
containing 10% FBS. All cell lines were cultured in a humidified
atmosphere at 37°C in 5% CO2.
Bioinformatics analysis
miRNAs targeting the 3′-UTR of brachyury mRNA were
predicted using miRanda (http://www.microrna.org/microrna/home.do) and
TargetScan (http://www.targetscan.org).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from tissue samples and cell
lines using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and was reverse transcribed using the TianScript RT kit
(Tiangen Biotech Co., Ltd., Beijing, China). The reverse
transcription reaction was performed in a total volume of 14.5 µl,
which included 2.5 µl (1 µg) total RNA, 2 µl dNTP mix (2.5 mM), 2
µl specific reverse transcription primer (10 µM) and 8 µl
RNase-free water. The reaction mixture was initially incubated at
70°C for 5 min, followed by incubation on ice for 2 min. The tubes
were centrifuged at 1,000 × g at 4°C for 5 sec, after which,
4 µl 5X First-Strand Buffer, 0.5 µl RNasin (40 U/µl) and 2 µl
reverse transcriptase (200 U/µl) were added to a total reaction
volume of 21 µl. The reaction mixture was then incubated at 42°C
for 50 min, followed by incubation at 95°C for 5 min. qPCR was
performed on an ABI PRISM 7500 Sequence Detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using SYBR FAST qPCR
Master Mix (Kapa Biosystems, Inc., Wilmington, MA, USA), according
to the manufacturer's protocol. The PCR conditions were as follows:
95°C for 3 min, 60°C for 20 sec; 40 cycles of 95°C for 3 sec, 60°C
for 20 sec; and 95°C for 15 sec, 60°C for 15 sec and 95°C for 15
sec. Each sample was run in triplicate. The relative expression of
miRNA compared with U6 was calculated using the 2−ΔΔCq
method (16). The primer sequences,
which were designed by Beijing Microread Genetics, Co., Ltd.
(Beijing, China), were as follows: miR-219-5p stem-loop,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGAAGAATTGC-3′;
miR-219-5p forward, 5′-GGACGGTTGATTGTCCAAAC-3′; common loop
reverse, 5′-TCGTATCCAGTGCAGGGTC-3′; U6 forward,
5′-CTCGCTTCGGCAGCACATATACT-3′; and U6 reverse,
5′-ACGCTTCACGAATTTGCGTGTC-3′.
Dual luciferase report vector
construction and assay
The 3′-UTR fragment of brachyury mRNA containing the
seed sequence of miR-219-5p was amplified by PCR using T gene
forward and T gene reverse primers. The PCR product was then
digested using Xho I and Not I (Takara Bio, Inc.,
Tokyo, Japan) and inserted into the pmiR-RB-Report™ dual-luciferase
vector (Guangzhou RiboBio, Co., Ltd., Guangzhou, China), which
contains the firefly luciferase gene Luc and Renilla
luciferase gene Rluc, to construct the wild-type T-3′UTR
(T-WT) reporter vector. The mutant 3′UTR of the T gene, containing
mutated miRNA binding sites, was amplified using T-mut forward and
T-mut reverse primers prior to splicing by overlap extension PCR.
This was then inserted into the vector to construct the mutant
T-3′UTR (T-Mut) reporter vector. The following primer sequences
were used (Guangzhou RiboBio, Co., Ltd.): T forward,
5′-GATTACTCGAGAGCAGCAAGGCCCAGG-3′ and T reverse:
5′-ATTGCGGCCGCGCATATTGCGTTTATTTTG-3′; and T-mut forward,
5′-GTAGCCAAACTGTTAGTGCAGAAAGCATTTTCTG-3′ and T-mut reverse,
5′-GCTTTCTGCACTAACAGTTTGGCTACTTTGTCAA-3′. Upon reaching 80%
confluence, U-CH2 cells were seeded into rat tail type I
collagen-coated 96-well plates at a density of 3,000 cells per well
(100 µl medium per well) in triplicate. After 24 h of cultivation,
the cells were transiently co-transfected with 100 ng T-WT or T-Mut
reporter vectors and 75 nM miR-219-5p mimics or non-target control
(NC; Guangzhou RiboBio, Co., Ltd.) using Lipofectamine 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). After 72 h, firefly
and Renilla luminescence were measured using a Modulus™
Microplate Fluorometer (Turner BioSystems, Sunnyvale, CA, USA) with
the Dual-Glo® Luciferase Assay system (Promega
Corporation, Madison, WI, USA), according to the manufacturer's
protocol.
Cell counting kit (CCK)-8 and EdU
assays
U-CH2 cells were seeded into rat tail type I
collagen-coated 96-well plates at a density of 3,000 cells per well
(100 µl medium per well) in triplicate. Subsequently, the cells
were transfected with 75 nM miR-219-5p mimics, NC, miR-219-5p
inhibitor or NC inhibitor (Guangzhou RiboBio, Co., Ltd.) using
Lipofectamine 2000. After 24 h of cultivation, viable cells were
evaluated daily for 4 days using CCK-8 assays (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan), according to the
manufacturer's protocol. Briefly, 10 µl CCK-8 was added to each
well and, after 1 h, the optical density (OD) of the samples was
measured at an absorbance of 450 nm using a microplate
spectrophotometer (Epoch 2; BioTek Instruments, Winooski, VT, USA).
Each transfection was performed in triplicate. The impact of
miR-219-5p on U-CH2 cell proliferation was also assessed using the
Cell-Light™ EdU detection kit (Guangzhou RiboBio, Co., Ltd.),
according to the manufacturer's instructions. Briefly, at 72 h
following transfection, cells were incubated with 10 µM EdU for 2 h
prior to fixation with 4% paraformaldehyde for 30 min and
permeabilization using 0.5% Triton X-100 for 10 min. Proliferating
cells were stained with 100 µl 1X Apollo643 for 30 min, after which
the cells were stained with 100 µl 1X Hoechst 33342. Fluorescent
cells were counted using the automated Acumen eX3 Laser Scanning
Cytometer (TTP Labtech, Ltd., London, UK).
Clone formation assay
U-CH2 or 293T cells were inoculated into rat tail
type I collagen-coated 24-well plates at a density of 300 cells per
well. After 18 h, the cells were transfected with 75 nM miR-219-5p
mimics, NC, miR-219-5p inhibitor or NC inhibitor using
Lipofectamine 2000, according to the manufacturer's protocol.
Subsequently, the cells were cultured in a humidified chamber at
37°C containing 5% CO2, with culture medium changed
every 2 days. After 5 days of cultivation, the cells were stained
with 20 µM Calcein-AM (Invitrogen; Thermo Fisher Scientific, Inc.)
for 45 min. Cell clusters >200-µm in size were considered
colonies and were counted using the automated Acumen eX3 Laser
Scanning Cytometer.
Western blotting
At 72 h following transfection, U-CH2 cells were
lysed in ice-cold radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China). The protein
concentration was estimated using a bicinchoninic acid protein
assay (Beyotime Institute of Biotechnology). A total of 50 µg
protein was separated by 10% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). The membranes were blocked with 5% non-fat dry milk in 0.1%
Tween-20 at 4°C for 1 h, then incubated with primary antibodies
against brachyury (1:1,000; ab20680; Abcam, Cambridge, MA, USA) or
β-actin (1:2,000; sc-130656; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) overnight at 4°C. Membranes were washed with TBS and
incubated with alkaline phosphatase-conjugated goat anti-rabbit
secondary antibody (1:3,000; BA1011; Wuhan Boster Biological
Technology, Ltd., Wuhan, China). Respective bands from these blots
were observed using the BCIP/NBT Color Development Substrate
(Promega Corporation). The experiments were repeated three times.
The mean densities of the bands were analyzed using Quantity One
4.6.2 software (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
normalized to that of β-actin.
Statistical analysis
Statistical analyses were performed using SPSS
software, version 21.0 (IBM SPSS, Armonk, NY, USA) and GraphPad
Prism, version 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Differences in miR-219-5p expression were assessed using
non-parametric tests (Mann-Whitney U test) due to the heterogeneity
of variances of the residuals when applying analysis of variance.
The relative luminescence ratios, ratio of cell proliferation and
colony formation, and densities of bands were analyzed using
Student's t-tests. Two-sided Fisher's exact test was used to
identify differences between categorical variables. P<0.05 was
considered statistically significant.
Results
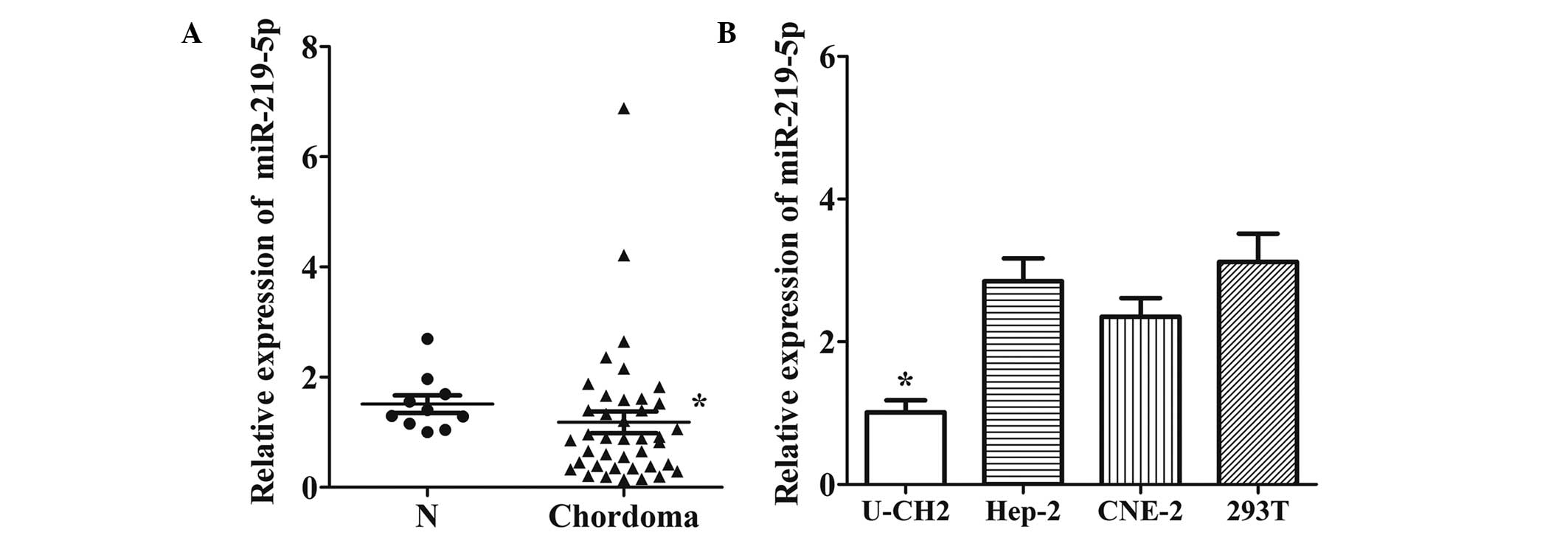
miR-219-5p is downregulated in
chordoma tissues and the U-CH2 cell line
Using miRanda and TargetScan, 16 miRNAs targeting
brachyury mRNA were predicted, including miR-23a, miR-219-5p,
miR-221, miR-222 and miR-448. Of these, RT-qPCR demonstrated that
the expression of miR-219-5p was downregulated in chordoma tissue
compared with normal skeletal muscle (Fig. 1A). Subsequently, the expression levels
of miR-219-5p in U-CH2 cells were compared with CNE-2, Hep-2 and
293T cells (Fig. 1B). These results
suggest that the expression of miR-219-5p is significantly
decreased in chordoma tissues and the U-CH2 cell line.
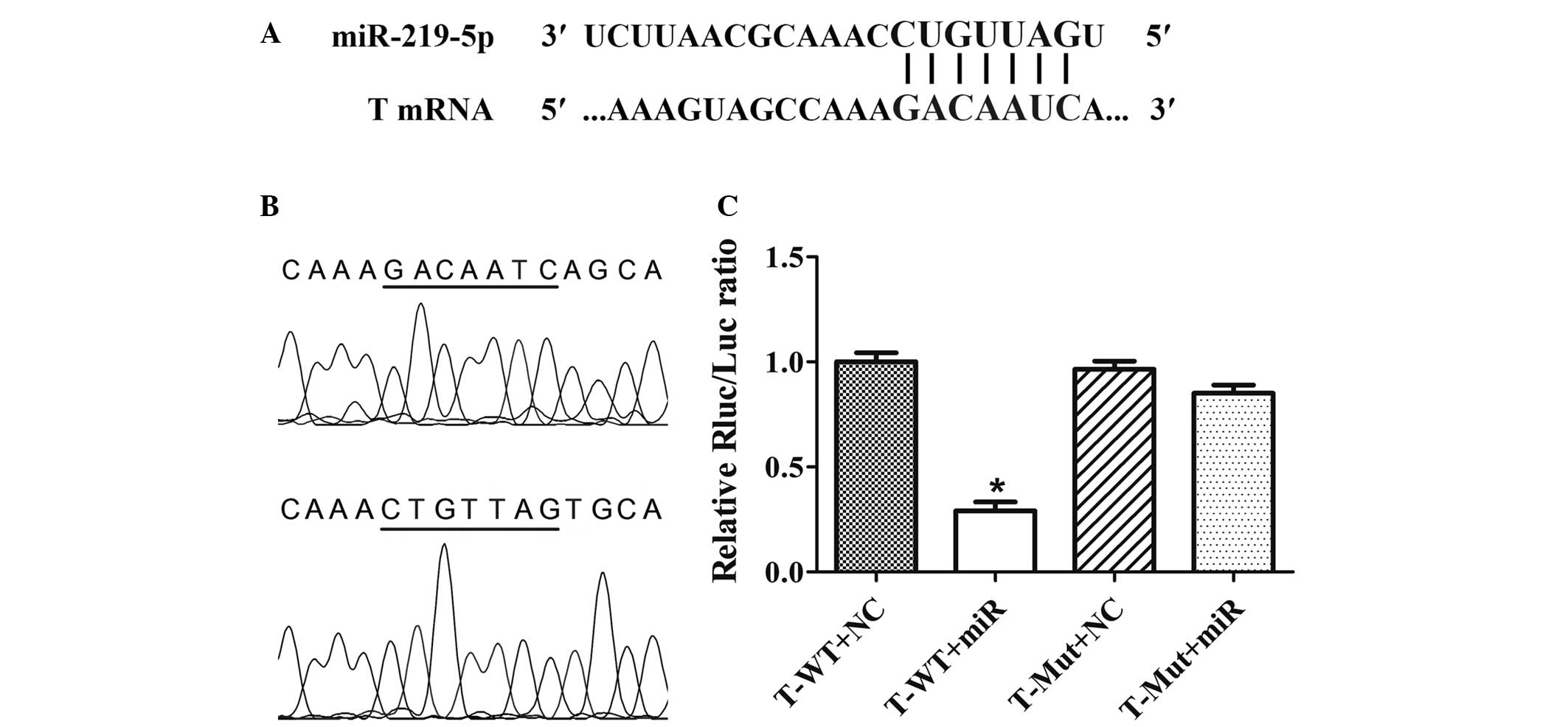
miR-219-5p directly binds to the
3′-UTR of brachyury mRNA
The miR-219-5p seed sequence in the 3′-UTR of
brachyury mRNA was predicted using miRanda (Fig. 2A). The wild-type and mutant T-3′-UTR
fragments, containing the miR-219-5p target site, were inserted
into the reporter vectors and validated by DNA sequencing (Fig. 2B). Dual-luciferase reporter assays
showed that, when miR-219-5p was overexpressed, the luciferase
activity in the wild-type T-3′-UTR reporter vector was
significantly reduced (Fig. 2C).
However, luciferase activity in the reporter vector containing the
mutant T-3′-UTR was not affected by miR-219-5p. These results
suggest that miR-219-5p may negatively regulate brachyury
expression by directly binding to the GACAAUC sequence in the
3′-UTR of T gene mRNA.
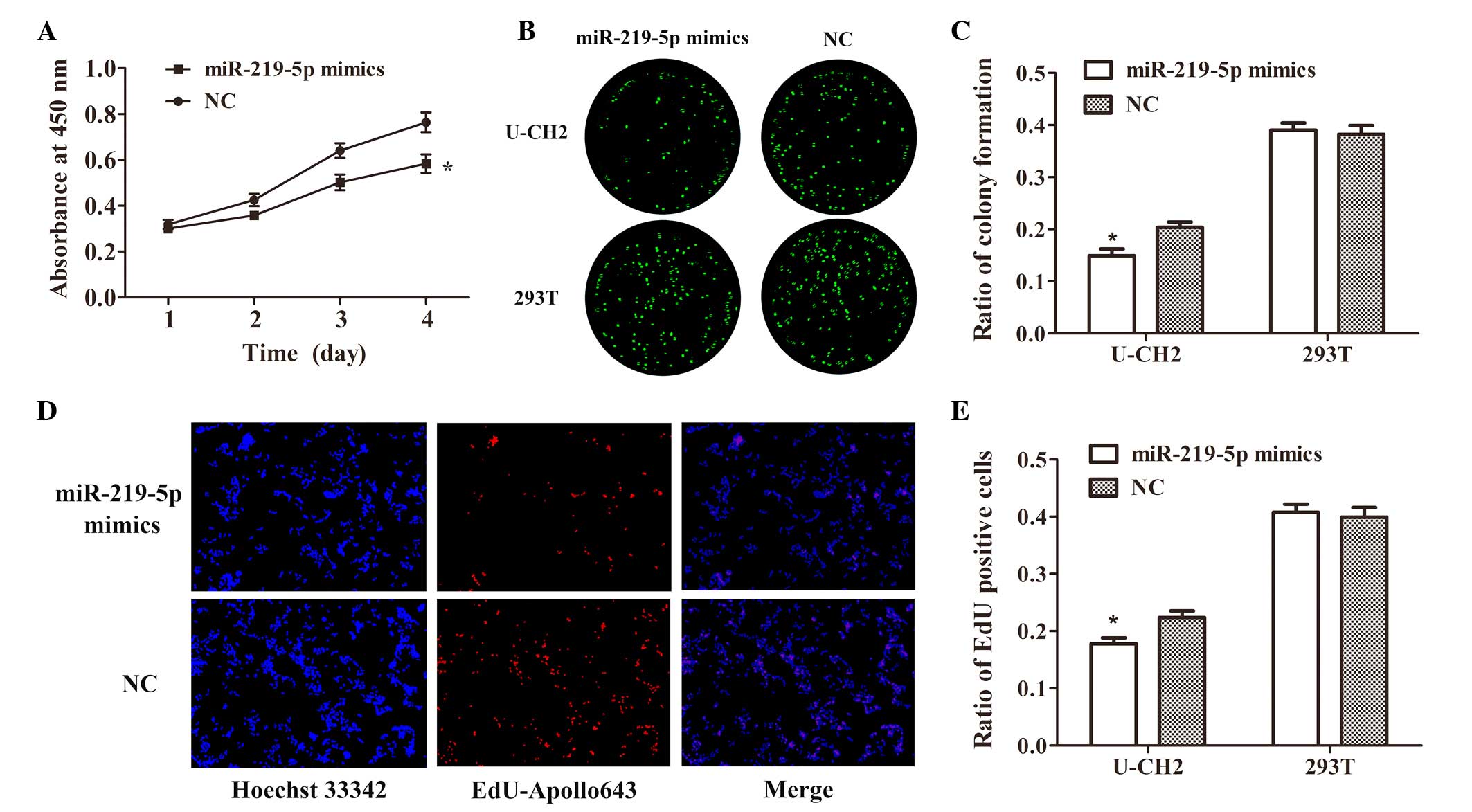
Effect of miR-219-5p overexpression on
cell proliferation and colony formation in chordoma cells
To confirm the biological function of miR-219-5p in
chordoma, CCK-8, EdU and clone formation assays of U-CH2 cells
transfected with miR-219-5p mimics or NC were performed. As a
comparison, the effect of miR-219-5p on proliferation and colony
formation was also measured in 293T cells. Significantly reduced
cell growth activity (P<0.01; Fig.
3A), colony formation (P<0.05; Fig. 3B and C) and proliferation (P<0.05;
Fig. 3D and E) were observed in U-CH2
cells transfected with miRNA-219-5p mimics compared with NC. For
293T cells, there was no significant difference between the cells
transfected with miR-219-5p mimics and NC in terms of colony
formation (Fig. 3B and C) or cell
proliferation (Fig. 3E). These
results suggest that overexpression of miR-219-5p is able to
suppress cell proliferation and colony formation in U-CH2
cells.
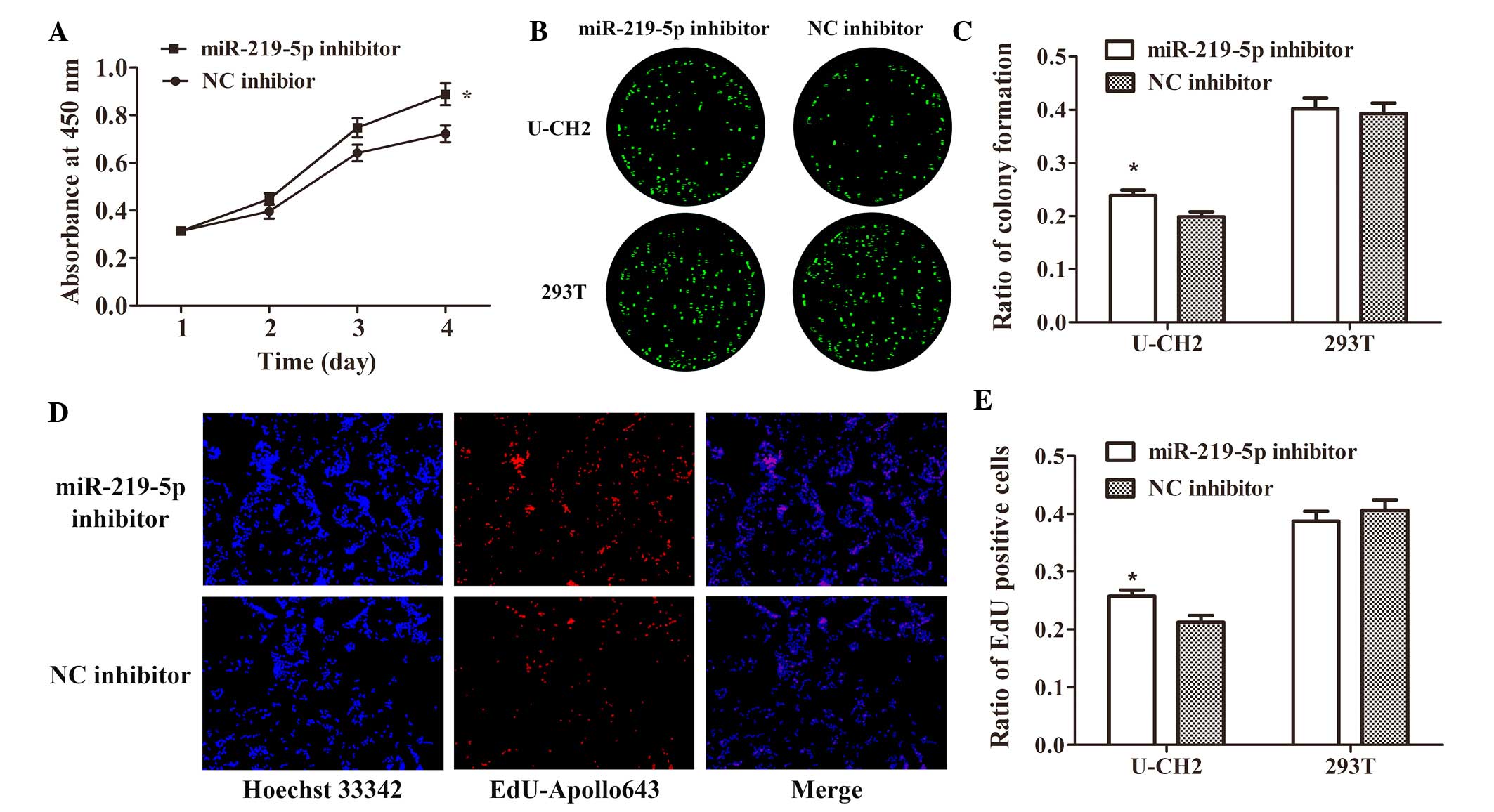
Effect of miR-219-5p knockdown on cell
proliferation and colony formation in UCH-2 cells
The effect of inhibiting miR-219-5p on U-CH2 and
293T cells was examined. The results showed that downregulating the
expression of miR-219-5p by transfection with miR-219-5p inhibitors
significantly increased U-CH2 cell growth activity (P<0.05;
Fig. 4A), colony formation (Fig. 4B and C), and proliferation (Fig. 4D and E), as compared with cells
transfected with NC inhibitor. There was no significant difference
in cell proliferation or colony formation between the 293T cells
transfected with miRNA-219-5p inhibitor and NC inhibitor (Fig. 4). These results indicate that
downregulation of miR-219-5p promotes U-CH2 cell proliferation and
colony formation in vitro.
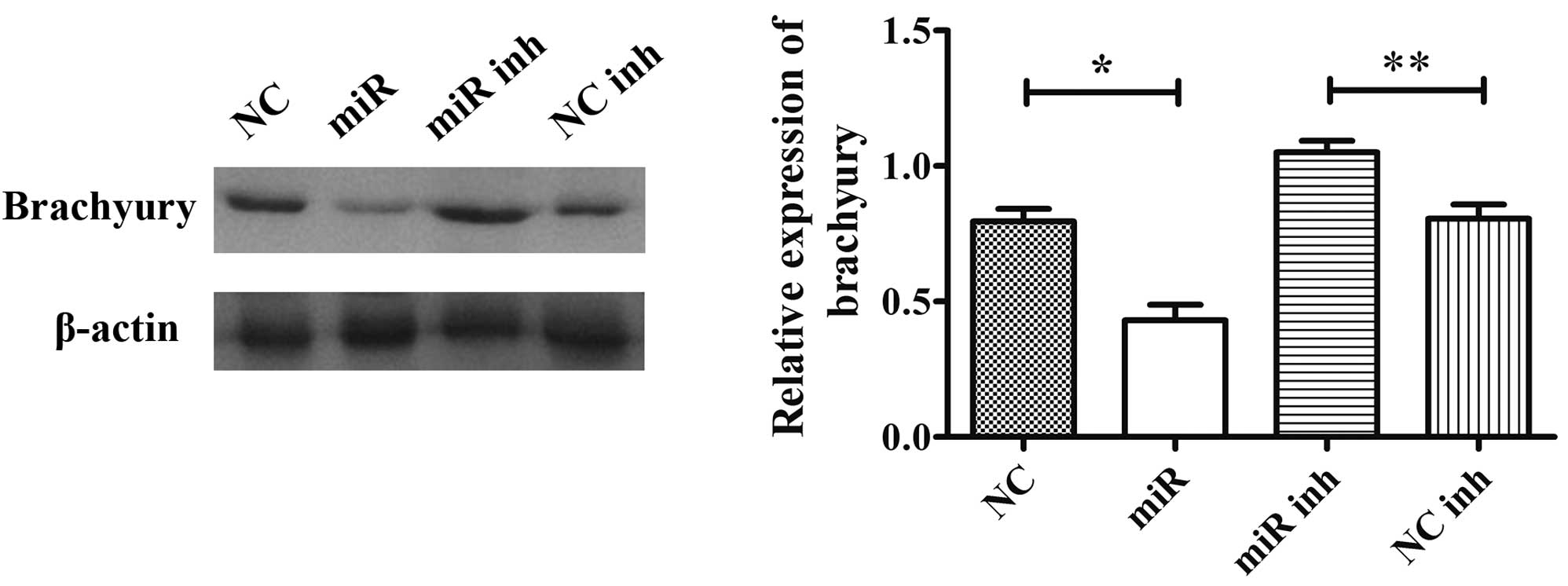
miR-219-5p regulates the expression of
brachyury in U-CH2 cells
U-CH2 cells were transfected with miR-219-5p mimic,
NC, NC inhibitor or miR-219-5p inhibitor, and the effects of
miR-219-5p on the protein expression levels of brachyury were
analyzed using western blotting (Fig.
5). The results demonstrated that the protein expression levels
of brachyury were significantly reduced in miR-219-5p
mimic-transfected U-CH2 cells compared with cells transfected with
NC (P<0.01), but were increased in miR-219-5p
inhibitor-transfected U-CH2 cells compared with cells transfected
with NC inhibitor (P<0.05). These results suggest that
miR-219-5p may regulate the protein expression of brachyury in
U-CH2 cells.
miR-219-5p is correlated with tumor
extent and recurrence
Among the 40 patients with skull base chordoma, 23
patients did not receive treatment prior to surgery. Five patients
experienced tumor recurrence following the primary surgery and had
to be operated on again. Twelve patients underwent the primary
surgery at centers other than Xuanwu hospital. There was no
significant relationship between the relative expression level of
miR-219-5p and patient age (P=1.000) and gender (P=0.755; Table II). Compared with the initial tumors,
the expression levels of miR-219-5p were significantly lower in
recurrent tumors (P=0.022). The expression levels of miR-219-5p
were downregulated in 12 of the 13 chordomas where the tumor had
gone beyond the clivus. No statistically significant relationship
was observed between surgical approaches and the miR-219-5p level
(P=0.210).
| Table II.Correlations between miR-219-5p
expression and various clinicopathological characteristics of skull
base chordoma. |
Table II.
Correlations between miR-219-5p
expression and various clinicopathological characteristics of skull
base chordoma.
|
| miR-219-5p (n,
%) |
|
|
|---|
|
|
|
|
|
|---|
| Features | Downregulation
(<1) | Upregulation
(>1) | Total (n) | P-value |
|---|
| Gender |
|
|
| 0.755 |
|
Male | 12 (57.1) | 9 (42.9) | 21 |
|
|
Female | 12 (63.2) | 7 (36.8) | 19 |
|
| Age (years) |
|
|
| 1.000 |
|
≤40 | 10 (58.8) | 7 (41.2) | 17 |
|
|
>40 | 14 (60.9) | 9 (39.1) | 23 |
|
| Tumor site |
|
|
| 0.005 |
|
Clivus | 12 (44.4) | 15 (55.6) | 27 |
|
| Beyond
clivus | 12 (92.3) | 1 (7.7) | 13 |
|
|
Initial/recurrent |
|
|
| 0.022 |
|
Initial | 10 (43.5) | 13 (56.5) | 23 |
|
|
Recurrent | 14 (82.4) | 3 (17.6) | 17 |
|
| Surgical
approach |
|
|
| 0.210 |
|
Endoscopic | 18 (54.5) | 15 (45.6) | 33 |
|
|
Traditional | 6 (85.7) | 1 (14.3) | 7 |
|
Discussion
Owing to the intricate anatomic location, large
tumor size and extensive bleeding during operation, complete
surgical resection of chordoma is extremely difficult (2). A better understanding of the molecular
mechanisms involved in tumor progression may assist the development
of novel therapeutic strategies for this malignant disease.
Accumulating evidence indicates that miRNAs may have an important
role in the progression of human chordoma (17–21). For
example, reduced expression of miR-1 has been reported in chordoma,
and miR-1 is suggested to suppress the growth of chordoma cells by
inhibiting Met and histone deacetylase 4 (17). Further studies have shown that miR-1
inhibits chordoma cell migration and invasion and correlates with
clinical prognosis (18,19). Another study confirmed that miR-608
and miR-34a were significantly downregulated in chordoma and may
influence chordoma malignancy by targeting epidermal growth factor
receptor (EGFR), B-cell lymphoma-xL and Met (20). Furthermore, some miRNAs are reported
to impact the pathogenesis of chordoma by regulating the
mitogen-activated protein kinase (MAPK) signaling pathway (21). In the present study, the relative
expression level of miR-219-5p in 40 skull base chordoma specimens
and different cell lines were evaluated. Our data suggested that
miR-219-5p was downregulated in chordoma tissues and the U-CH2 cell
line. In addition, the exogenous overexpression of miR-219-5p
inhibited the proliferation and clonogenicity of U-CH2 cells by
targeting brachyury, while suppression of miR-219-5p expression had
the opposite effect.
Brachyury is a master regulator in chordoma that
binds directly to ~100 targets and indirectly influences the
expression of >60 other genes, including genes regulating the
cell cycle, growth factors and cytokines, as well as extracellular
matrix genes (22). Efforts have been
made to clarify the mechanisms underlying the regulation of
brachyury in chordoma. In the genera Ascidians and
Xenopus, as well as zebrafish, brachyury has been shown to
be regulated by fibroblastic growth factor receptors via the
RAS/RAF/MAPK kinase/extracellular signal-regulated kinase-ETS2
signaling pathway, but genetic alterations of these genes have not
been detected in human chordoma (23). The P63 gene can regulate brachyury
expression in mouse embryonic fibroblasts and murine-derived
osteosarcomas, but not in human chordomas (24). In the present study, the inhibitory
effect of miR-219-5p on brachyury, predicted by miRanda and
TargetScan, was validated by the dual-luciferase reporter assay.
In vitro experiments showed that miR-219-5p regulated the
protein expression of brachyury in chordoma cells, and affected
cell proliferation and clonogenicity. The results of the present
study suggest that the loss of regulation of miR-219-5p on
brachyury may be an important mechanism in the initiation and
progression of chordoma.
A review of the literature demonstrated that
miR-219-5p expression is consistently dysregulated in various
tumors (25–29). Downregulation of miR-219-5p has been
reported in glioblastoma (25). The
exogenous overexpression of miR-219-5p in glioma cell lines could
inhibit cell proliferation, growth and migration by targeting EGFR
and inhibiting the receptor tyrosine kinase pathway (26). In addition, low expression of
miR-219-5p has been associated with higher clinical stages and
recurrence rates of meningioma (27).
The level of miR-219-5p was reduced in hepatocellular carcinoma. In
a previous study, miR-219-5p inhibited cell proliferation in
vitro and resulted in the arrest of the cell cycle at the G1/S
transition by inhibiting the expression of glypican-3 (28). miR-219-5p was also downregulated in
papillary thyroid carcinoma and had a critical role in cell growth
by inhibiting estrogen receptor-α (29). These findings, together with the
results of the present study, indicate that miR-219-5p may exert a
tumor suppressor role in the carcinogenesis of various tumors.
It remains controversial as to whether brachyury is
a prognostic indicator for chordoma. Kitamura et al
(30) reported that the expression of
brachyury was an independent prognostic indicator in skull base
chordoma, and was associated with a poor prognosis. Zhang et
al (31) analyzed a chordoma
tissue microarray and determined that brachyury was not a
prognostic indicator for chordoma, although the specimens were
mainly derived from mobile spine or sacrum chordoma. Expression of
miR-219-5p has been correlated with the recurrence rate of
meningioma (27) and overall survival
time of hepatocellular carcinoma patients (28). The present study demonstrated that
miR-219-5p expression was associated with tumor extent and
recurrence, although no definitive conclusion could be drawn owing
to the small sample size of the present study. As a regulator of
brachyury, the prognostic value of miR-219-5p in skull base
chordoma requires further investigation.
In summary, the present study identified that
miR-219-5p was downregulated in chordoma tissues and the U-CH2 cell
line, and was associated with extensive tumor and recurrence.
Furthermore, miR-219-5p exhibited the ability to inhibit cell
proliferation and clonogenicity of chordoma by targeting brachyury.
These findings suggested that miR-219-5p may act as a tumor
suppressor in human chordoma, and it may be a potential target for
therapeutic intervention in chordoma.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81170894), the
Specialized Project on Scientific Research within the Healthcare
Circle by National Health and Family Planning Commission of China
(grant no. 201202005) and the Key Discipline Construction Project
of the Finance Bureau of Beijing (grant no. 3500–114235).
References
|
1
|
Casali PG, Stacchiotti S, Sangalli C, Olmi
P and Gronchi A: Chordoma. Curr Opin Oncol. 19:367–370. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stippler M, Gardner PA, Snyderman CH,
Carrau RL, Prevedello DM and Kassam AB: Endoscopic endonasal
approach for clival chordomas. Neurosurgery. 64:268–277. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vujovic S, Henderson S, Presneau N, Odell
E, Jacques TS, Tirabosco R, Boshoff C and Flanagan AM: Brachyury, a
crucial regulator of notochordal development, is a novel biomarker
for chordomas. J Patho. 209:157–165. 2006. View Article : Google Scholar
|
|
4
|
Yang XR, Ng D, Alcorta DA, Liebsch NJ,
Sheridan E, Li S, Goldstein AM, Parry DM and Kelley MJ: T
(brachyury) gene duplication confers major susceptibility to
familial chordoma. Nat Genet. 41:1176–1178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Presneau N, Shalaby A, Ye H, Pillay N,
Halai D, Idowu B, Tirabosco R, Whitwell D, Jacques TS, Kindblom LG,
et al: Role of the transcription factor T (brachyury) in the
pathogenesis of sporadic chordoma: A genetic and functional-based
study. J Pathol. 223:327–335. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hsu W, Mohyeldin A, Shah SR, ap Rhys CM,
Johnson LF, Sedora-Roman NI, Kosztowski TA, Awad OA, McCarthy EF,
Loeb DM, et al: Generation of chordoma cell line JHC7 and the
identification of Brachyury as a novel molecular target. J
Neurosurg. 115:760–769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Szuhai K and Hogendoorn PC: ‘The chicken
or the egg?’ dilemma strikes back for the controlling mechanism in
chordoma(#). J Pathol. 228:261–265. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: MiRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Le XF, Merchant O, Bast RC and Calin GA:
The roles of microRNAs in the cancer nvasion-metastasis cascade.
Cancer Microenviron. 3:137–147. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li H, Xie S, Liu M, Chen Z, Liu X, Wang L,
Li D and Zhou Y: The clinical significance of downregulation of
mir-124-3p, mir-146a-5p, mir-155-5p and mir-335-5p in gastric
cancer tumorigenesis. Int J Oncol. 45:197–208. 2014.PubMed/NCBI
|
|
12
|
Xu XM, Qian JC, Deng ZL, Cai Z, Tang T,
Wang P, Zhang KH and Cai JP: Expression of miR-21, miR-31, miR-96
and miR-135b is correlated with the clinical parameters of
colorectal cancer. Oncol Lett. 4:339–345. 2012.PubMed/NCBI
|
|
13
|
Jikuzono T, Kawamoto M, Yoshitake H,
Kikuchi K, Akasu H, Ishikawa H, Hirokawa M, Miyauchi A, Tsuchiya S,
Shimizu K and Takizawa T: The miR-221/222 cluster, miR-10b and
miR-92a are highly upregulated in metastatic minimally invasive
follicular thyroid carcinoma. Int J Oncol. 42:1858–1868.
2013.PubMed/NCBI
|
|
14
|
Ries J, Vairaktaris E, Agaimy A, Kintopp
R, Baran C, Neukam FW and Nkenke E: MiR-186, miR-3651 and miR-494:
Potential biomarkers for oral squamous cell carcinoma extracted
from whole blood. Oncol Rep. 31:1429–1436. 2014.PubMed/NCBI
|
|
15
|
Flanagan AM and Yamaguchi T: ChordomaWHO
Classification of Tumours of Soft Tissue and Bone. IARC Press;
Lyon: pp. 328–329. 2013
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Duan Z, Choy E, Nielsen GP, Rosenberg A,
Iafrate J, Yang C, Schwab J, Mankin H, Xavier R and Hornicek FJ:
Differential expression of microRNA (miRNA) in chordoma reveals a
role for miRNA-1 in Met expression. J Orthop Res. 28:746–752.
2010.PubMed/NCBI
|
|
18
|
Osaka E, Yang X, Shen JK, Yang P, Feng Y,
Mankin HJ, Hornicek FJ and Duan Z: MicroRNA-1 (miR-1) inhibits
chordoma cell migration and invasion by targeting slug. J Orthop
Res. 32:1075–1082. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Duan Z, Shen J, Yang X, Yang P, Osaka E,
Choy E, Cote G, Harmon D, Zhang Y, Nielsen GP, et al: Prognostic
significance of miRNA-1 (miR-1) expression in patients with
chordoma. J Orthop Res. 32:695–701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Schiff D, Park D and Abounader R:
MicroRNA-608 and microRNA-34a regulate chordoma malignancy by
targeting EGFR, Bcl-xL and MET. PLoS One. 12:e915462014. View Article : Google Scholar
|
|
21
|
Long C, Jiang L, Wei F, Ma C, Zhou H, Yang
S, Liu X and Liu Z: Integrated miRNA-mRNA analysis revealing the
potential roles of miRNAs in chordomas. PLoS One. 8:e666762013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nelson AC, Pillay N, Henderson S, Presneau
N, Tirabosco R, Halai D, Berisha F, Flicek P, Stemple DL, Stern CD,
et al: An integrated functional genomics approach identifies the
regulatory network directed by brachyury (T) in chordoma. J Pathol.
228:274–285. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shalaby AA, Presneau N, Idowu BD, Thompson
L, Briggs TR, Tirabosco R, Diss TC and Flanagan AM: Analysis of the
fibroblastic growth factor receptor-RAS/RAF/MEK/ERK-ETS2/brachyury
signalling pathway in chordomas. Mod Pathol. 22:996–1005. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pillay N, Amary FM, Berisha F, Tirabosco R
and Flanagan AM: P63 does not regulate brachyury expression in
human chordomas and osteosarcomas. Histopathology. 59:1025–1072.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rao SA, Santosh V and Somasundaram K:
Genome-wide expression profiling identifies deregulated miRNAs in
malignant astrocytoma. Mod Pathol. 23:1404–1417. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rao SA, Arimappamagan A, Pandey P, Santosh
V, Hegde AS, Chandramouli BA and Somasundaram K: MiR-219-5p
inhibits receptor tyrosine kinase pathway by targeting EGFR in
glioblastoma. PLoS One. 8:e631642013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhi F, Zhou G, Wang S, Shi Y, Peng Y, Shao
N, Guan W, Qu H, Zhang Y, Wang Q, et al: A microRNA expression
signature predicts meningioma recurrence. Int J Cancer.
132:128–136. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang N, Lin J, Ruan J, Su N, Qing R, Liu
F, Lv C, Zheng D and Luo R: MiR-219-5p inhibits hepatocellular
carcinoma cell proliferation by targeting glypican-3. FEBS Lett.
23:884–891. 2012. View Article : Google Scholar
|
|
29
|
Huang C, Cai Z, Huang M, Mao C, Zhang Q,
Lin Y, Zhang X, Tang B, Chen Y, Wang X, et al: MiR-219-5p modulates
cell growth of papillary thyroid carcinoma by targeting estrogen
receptor α. J Clin Endocrinol Metab. 100:E204–E213. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kitamura Y, Sasaki H, Kimura T, Miwa T,
Takahashi S, Kawase T and Yoshida K: Molecular and clinical risk
factors for recurrence of skull base chordomas: Gain on chromosome
2p, expression of brachyury and lack of irradiation negatively
correlate with patient prognosis. J Neuropathol Exp Neurol.
72:816–823. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang L, Guo S, Schwab JH, Nielsen GP,
Choy E, Ye S, Zhang Z, Mankin H, Hornicek FJ, Duan Z, et al: Tissue
microarray immunohistochemical detection of brachyury is not a
prognostic indicator in chordoma. PLoS One. 8:e758512013.
View Article : Google Scholar : PubMed/NCBI
|