Introduction
MicroRNAs (miRNAs) are a class of small non-coding
RNAs that negatively regulate gene expression by binding to the
3′untranslated region (3′UTR) of target mRNAs (1,2). There is
emerging evidence that miRNAs function as oncogenes or tumor
suppressors through interaction with a number of the signal
transduction pathways involved in transformation and carcinogenesis
(3–5).
miRNA-221/222 (miR-221/222) are located in tandem on the X
chromosome and have been reported as oncogenic miRNAs in multiple
types of advanced cancer (6–9). A number of studies have observed
overexpression of miR-221/222 in malignancies and the available
data suggest that miR-221/222 may become suitable targets for
cancer treatment (7–9).
The miRNA sponge method was introduced in 2007 by
Ebert et al (10); it is used
to create RNAs containing multiple tandem binding sites
complementary to the miRNAs of interest and leads to continuous
loss of miRNA function in cells and transgenic organisms. The miRNA
sponge has proven to be a valuable tool for miRNA loss of function
experimental systems (11). In a
previous study, a miR-221/222 sponge was successfully constructed
and its inhibitory effects on miR-221/222 in oral squamous cell
carcinoma (OSCC) cells were investigated (12).
Phosphatase and tensin homolog (PTEN) is a tumor
suppressor gene that is frequently mutated in many types of cancer
(13–16). PTEN expression is downregulated in a
wide range of malignancies, including glioblastoma, pancreatic
cancer, colorectal carcinoma, breast cancer and OSCC (13,14,16–20).
The phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) signaling
pathway is involved in multiple biological processes, including
cellular apoptosis, cell cycle regulation, survival and
proliferation (21). Previous studies
have demonstrated that aberrant activation of the PI3K/Akt
signaling pathway has a significant role in tumorigenesis and tumor
metastasis (14). PTEN functions as a
tumor suppressor by negatively regulating the PI3K/Akt signaling
pathway (22). miRNAs, including
miR-17-5p and miR29b, have been reported to regulate the expression
of PTEN (19,23). However, little is known about the
roles of miR-221/222 in the expression of PTEN in OSCC.
In the present study, bioinformatics analysis
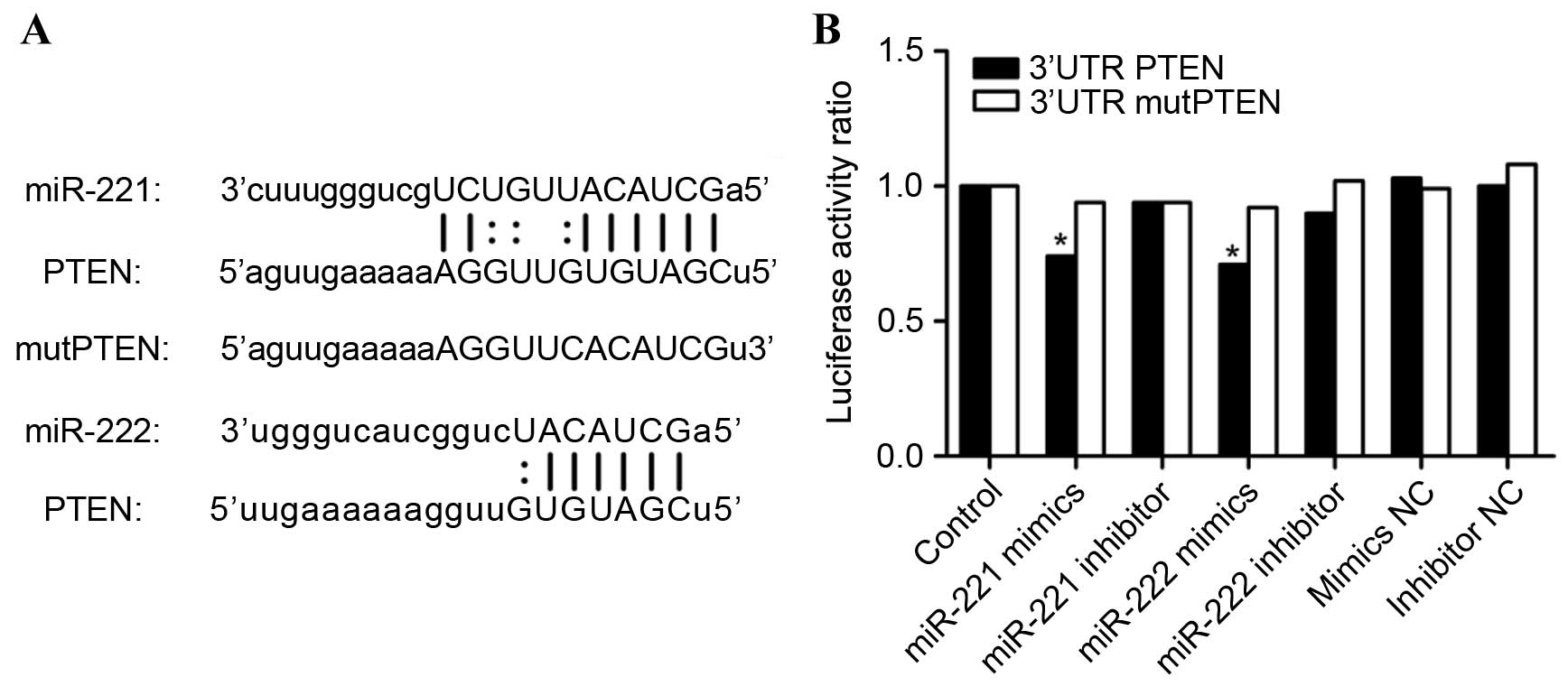
revealed that the PTEN gene may be a direct target of miR-221/222.
Binding sites for miR-221/222 were identified in the 3′UTR of PTEN
using miRanda (http://www.microrna.org/) and the result is shown in
Fig. 1A. miR-221/222 were identified
as potent regulators of PTEN. An miR-221/222 sponge was constructed
in CAL27 and HSC6 OSCC cells to validate the induction of
apoptosis, and reduction of cell proliferation and invasion,
through miR-221/222 inhibition and the upregulation of PTEN
expression.
Materials and methods
Cells and cell culture
The 293T cell line was purchased from Land Unicomed
(Guangzhou, China). The OSCC CAL27 cell line was purchased from
American Type Culture Collection (ATCC; Manassas, VA, USA) and the
OSCC HSC6 cell line was kindly provided by Dr J. Silvio Gutkind
(NIH; Besthesda, MD). All cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.,), 100 IU/ml penicillin
and 100 µg/ml streptomycin, and maintained at 37°C in a humidified
5% CO2 atmosphere.
Dual luciferase reporter assay
The 293T cells were cultured in 24-well plates,
transfected with 0.5 µg of the psi-CHECK2-PTEN (Land Unicomed) or
0.5 µg of the psi-CHECK2-mutPTEN (Land Unicomed), and 20 µM of
miR-221/222 inhibitor (Gene Pharma, Shanghai, China) or 20 µM of
miR-221/222 mimics (Gene Pharma) using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc. USA). Cells were
lysed by Passive Lysis Buffer (Promega Corporation, Madison, WI,
USA) and collected at 48 h post-transfection, and luciferase
activity was detected using the Dual-Luciferase Reporter Assay
system (Promega Corporation) according to the manufacturer's
protocol.
Construction of miRNA sponge and
transfection
An miR-221/222 sponge was constructed by inserting
tandemly arrayed miRNA binding sites into the 3′UTR of a reporter
gene encoding destabilized enhanced green fluorescent protein
driven by the murine cytomegalovirus promoter. Binding sites
contained 3 hsa-miR-221 and hsa-miR-222 antisense sequences
complementary to miR-221/222. The miR-221/222 sponge and empty
vector were transfected using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol, and the transfection medium was replaced
with fresh DMEM containing 10% FBS 6 h later. Following treatment
for 48 h, CAL27 and HSC6 cells were divided into 3 groups,
including the control, scramble and miR-221/222 sponge, and used
for subsequent analysis.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total miRNA and mRNA were extracted using the
miRNeasy mini kit (Qiagen GmbH, Hilden, Germany) and the RNeasy
Micro kit (Qiagen GmbH) according to the manufacturer's protocol.
RT-qPCR was performed using the Primescript RT reagent kit (Takara,
Otsu, Japan) with U6 and 18S rRNA as the loading controls. For the
detection of miR-221/222 and PTEN, the SYBR Green PCR assay
(SYBR® Premix Ex Taq™; Takara) was performed and the
primer sequences used were as follows: miR-221 forward,
ACACTCCAGCTGGGAGCTACATTGTCTGCTGG and reverse, CTCAACTGGTGTCGTGGA;
miR-222 forward, ACACTCCAGCTGGGAGCTACATCTGGCTACTG and reverse,
CTCAACTGGTGTCGTGGA; U6 forward, CTCGCTTCGGCAGCACA and reverse,
AACGCTTCACGAATTTGCGT; PTEN forward, TTGTGGTCTGCCAGCTAAA and
reverse, CGCTCTATACTGCAAATGCT; 18S rRNA forward,
CCTGGATACCGCAGCTAGGA and reverse, GCGGCGCAATACGAATGCCCC. The PCR
was performed in triplicate consisting of 40 cycles of a
denaturation step at 95°C for 5 sec, annealing at 60°C for 30 sec
and extension at 72°C for 30 sec after a cycle of a
pre-denaturation step at 95°C for 30 sec on an ABI
PRISM® 7500 Sequence Detector (Applied Biosystems,
Foster City, CA, USA). Relative expression levels were calculated
using the 2−ΔΔCq method (24).
Western blot analysis
Following 48 h of treatment with empty vector and
miR-221/222 sponge, 3 groups of cells were washed with pre-chilled
phosphate-buffered saline (PBS) 3 times and solubilized in RIPA
buffer with 1% protease inhibitor (Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany) and 10% phosphatase inhibitor (Roche
Diagnostics GmbH, Mannheim, Germany). Cell lysates were centrifuged
for 15 min at 14,000 × g and 4°C, and the protein
concentration was subsequently measured using the Enhanced BCA
protein assay kit (CWBIO, Beijing, China) according to the
manufacturer's protocol. The proteins were diluted with 5X loading
buffer (CWBIO, Beijing, China) and denatured at 99°C for 10 min. A
total of 35 g of protein per group was separated on 10% sodium
dodecyl sulfate-polyacrylamide gel and transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA) by electroblotting. The membranes were blocked with 5% skimmed
milk at room temperature for 1 h, then washed in Tris-buffered
saline and Tween 20 (TBST) and incubated with primary antibodies
against PTEN (catalog no. 9188), phosphorylated Akt (pAkt; catalog
no. 4060), Akt (catalog no. 4691) and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; catalog no. 5174) (dilution, 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) overnight at 4°C.
Subsequently, the blots were washed 3 times with TBST and incubated
with Horseradish Peroxidase-conjugated AffiniPure Goat Anti-Rabbit
IgG (1:3,000; Cell Signaling Technology, Inc.) for 1 h at room
temperature. The membranes were detected by an enhanced
chemiluminescence reagent kit (GE Healthcare Life Sciences,
Chalfont, UK) and visualized using the AlphaView SA system (Protein
Simple, San Jose, CA, USA). The specific protein bands were
quantified by ImageJ 1.48 (National Institutes of Health, Bethesda,
MD, USA) with the density of GAPDH used as the loading control.
Cell Counting Kit-8 (CCK8) assay
The viabilities of the control and miR-221/222
sponge transfected cells were evaluated using the CCK8 assay
(Beyotime Institute of Biotechnology, Haimen, China). Respectively,
CAL27 (4,000 cells/well) and HSC6 (3,000 cells/well) cell lines
were seeded into 96 well plates 1 day prior to transfection.
Following the transfection method previously mentioned, at time
points 12, 24, 36, 48, 60 and 72 h, 10 µl CCK8 reagent per well was
added and incubated for 2 h at 37°C in a humidified environment.
Optical density values were measured at a wavelength of 450 nm. The
data were derived from quadruplicate samples of at least three
independent experiments.
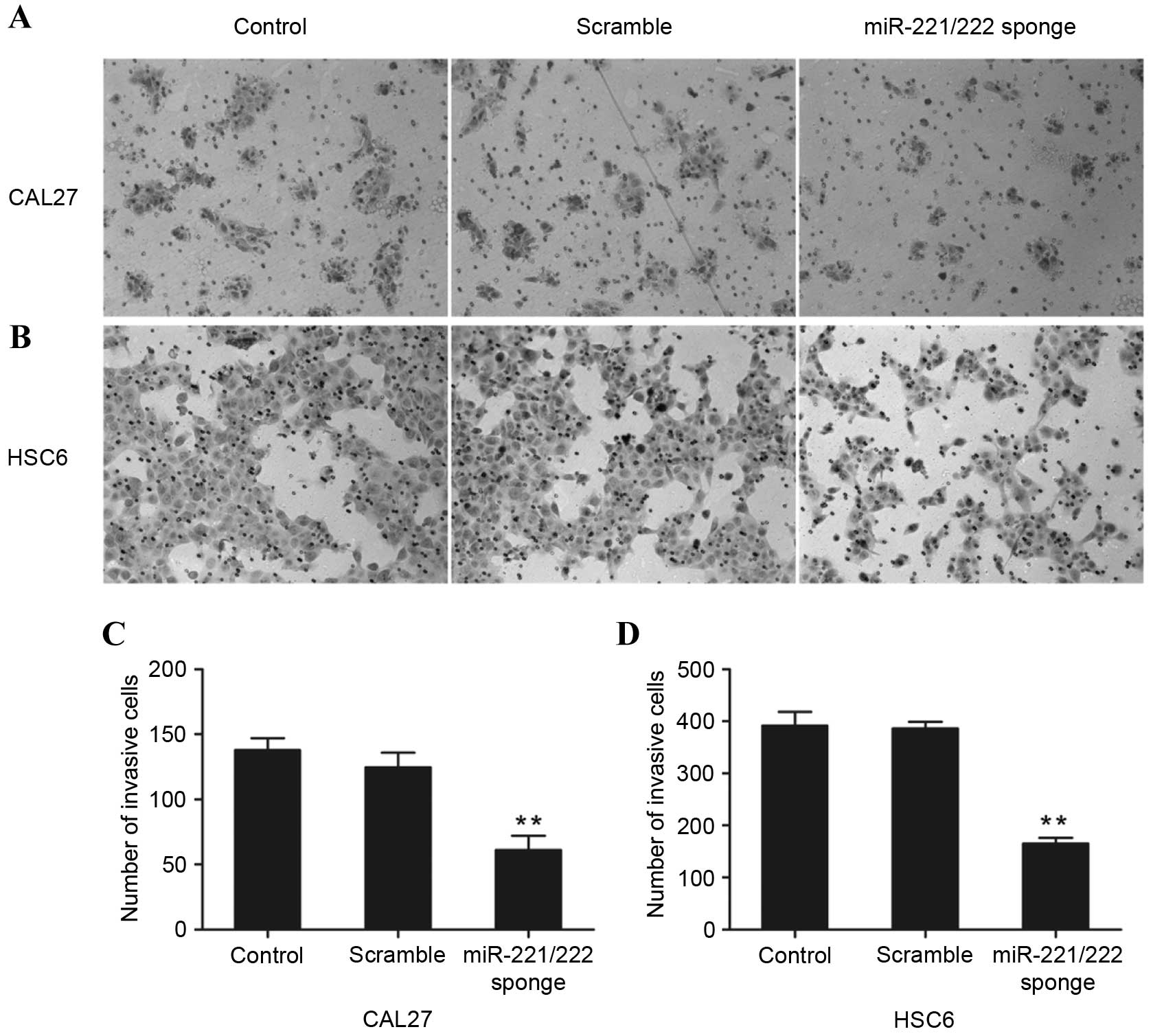
Transwell® assay
The invasive ability of the CAL27 and HSC6 cell
lines was evaluated by the Transwell assay, which measures the
movement of cells across a Matrigel®-coated membrane.
Transwell upper chambers were coated with 50 µl of 20% growth
factor-reduced Matrigel prior to the experiment. The parental and
transfected cells (CAL27, 1×105 cells/chamber; HSC6,
8×104 cells/chamber) were seeded into the upper chambers
in 100 µl serum-free DMEM, while 600 µl DMEM supplemented with 10%
FBS was added to the lower chambers. The cells were incubated at
37°C in a humidified 5% CO2 atmosphere for 24 h, and the
cells and Matrigel in the upper chambers were subsequently
discarded. Cells in the lower chamber were fixed with 4%
paraformaldehyde solution for 20 min and then stained with 0.05%
crystal violet for 15 min, followed by washing with PBS. Images
were captured using an inverted microscope and the number of
invasive cells was counted.
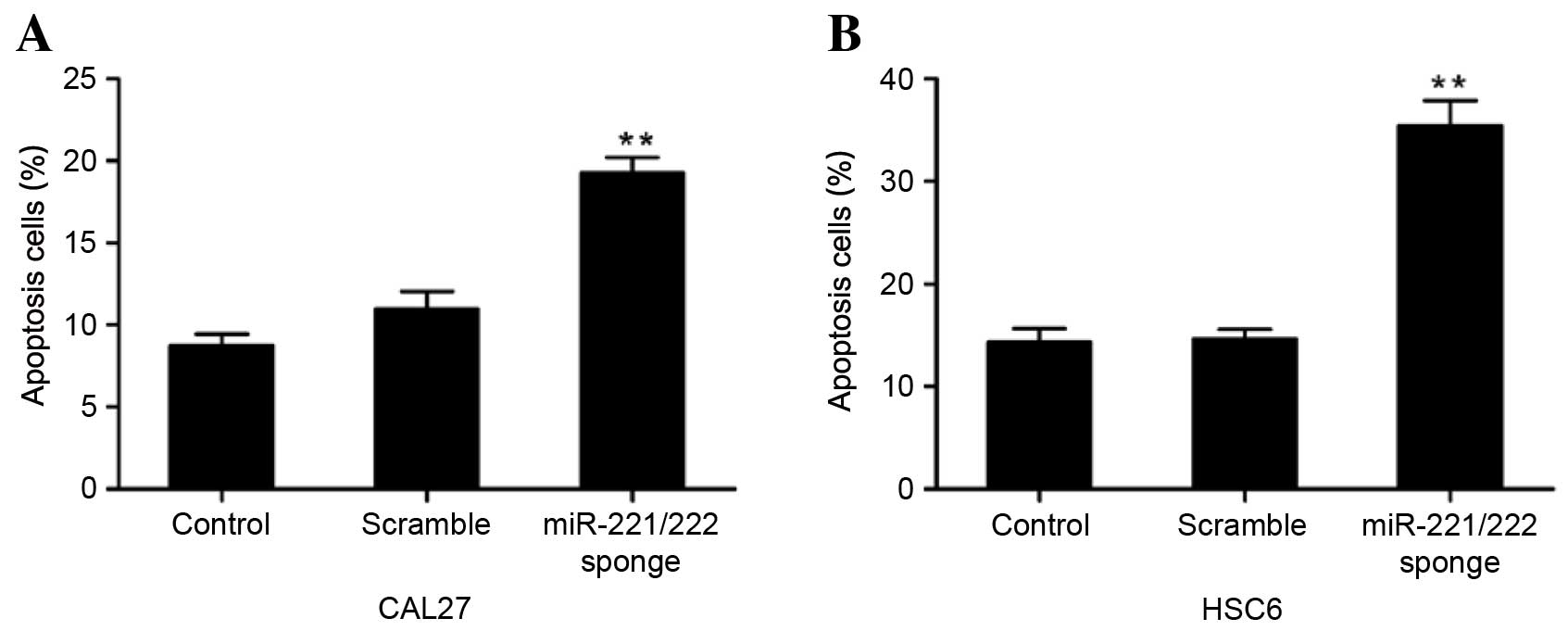
Flow cytometry
After transfection for 48 h, CAL27 and HSC6 cells
were collected in the log phase of growth by centrifugation for 5
min at 1,000 × g and 25°C, and rinsed with PBS 3 times. The
cell apoptosis assay was performed using an Annexin-V-FLUOS
Staining kit (Roche Diagnostics GmbH). Cells were resuspended in
incubation buffer at a density of 106/ml. Subsequently,
5 µl Annexin V-fluorescein isothiocyanate and 5 µl propidium iodide
(PI) were added to the cell suspension according to the
manufacturer's protocol. The cells were incubated for 15 min at
room temperature in the dark. The stained cells were analyzed by an
FC500 flow cytometer (Beckman Coulter, Fullerton, CA, USA).
Statistical analysis
All data are presented as the mean ± SD. The results
were determined by Student's t-test using SPSS version 15.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
miR-221/222 direct targeting of the
PTEN mRNA 3′UTR
To validate whether PTEN is a direct target of
miR-221/222, PTEN 3′UTR and mutPTEN 3′UTR luciferase constructs
were transfected into into 293T cells with miR-221/222 mimics,
mimic negative control (NC), miR-221/222 inhibitor or inhibitor NC.
Luciferase activity was detected using a Dual-Luciferase Reporter
Assay System. As can be observed in Fig.
1B, the luciferase activity of 293T cells transfected with
miR-221/222 and PTEN 3′UTR was reduced when compared with all other
treatment groups.
miR-221/222 and PTEN expression in
CAL27 and HSC6 cells
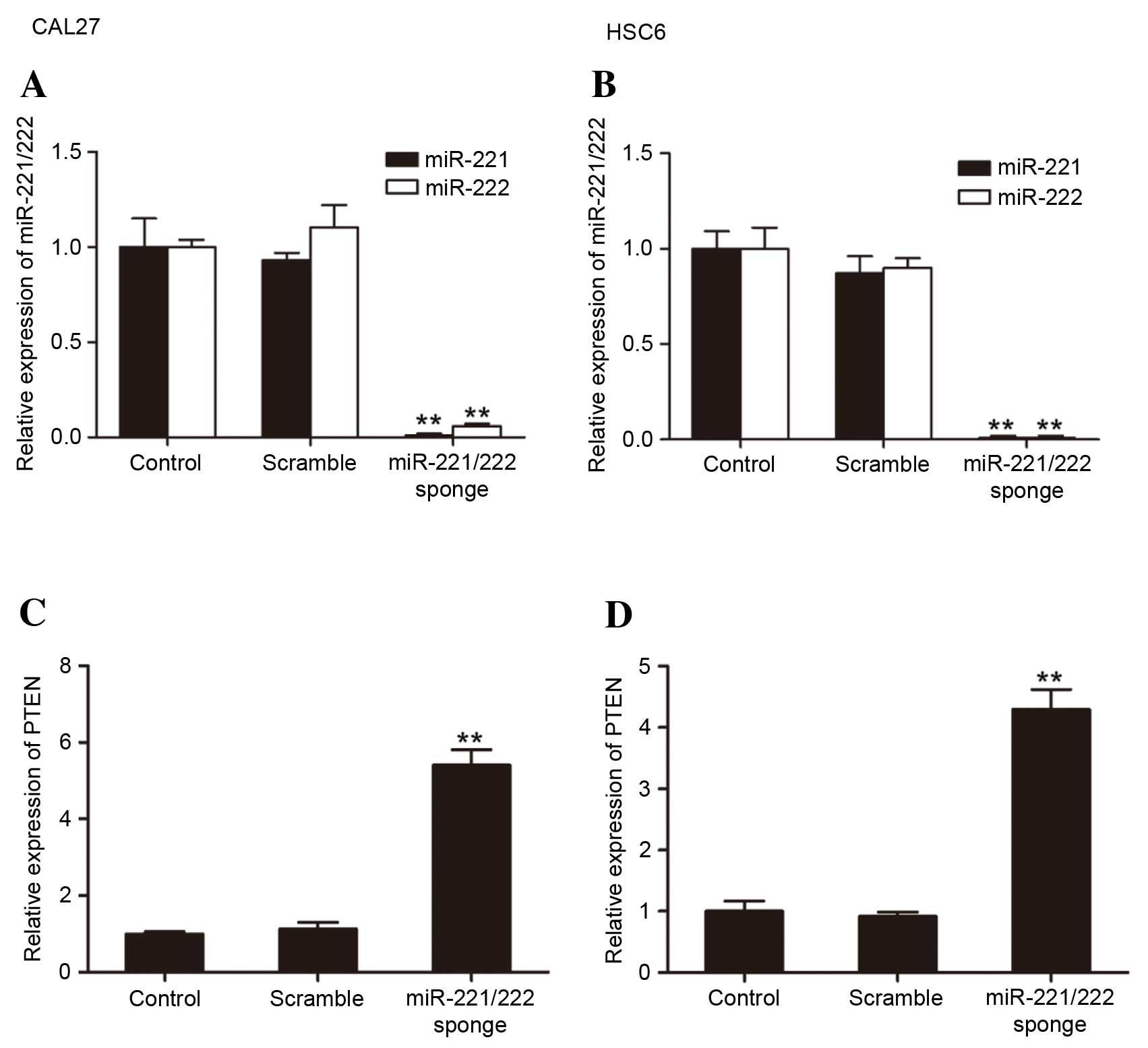
The present study analyzed the expression of
miR-221/222 and PTEN by RT-qPCR. miR-221/222 sponge transfection
significantly reduced the miR-221/222 levels compared with the
control groups (CAL27: miR-221 vs. control, P=0.0063; miR-222 vs.
control, P=0.0090; HSC6: miR-221 vs. control, P=0.00072; miR-222
vs. control, P=0.00058) (Fig. 2A and
B). The bioinformatics analysis predicted that miR-221/222
would regulate the expression of PTEN. The results of the present
study showed that in miR-221/222 sponge groups, a notable
upregulation of PTEN was observed in CAL27 and HSC6 cells (Fig. 2C and D).
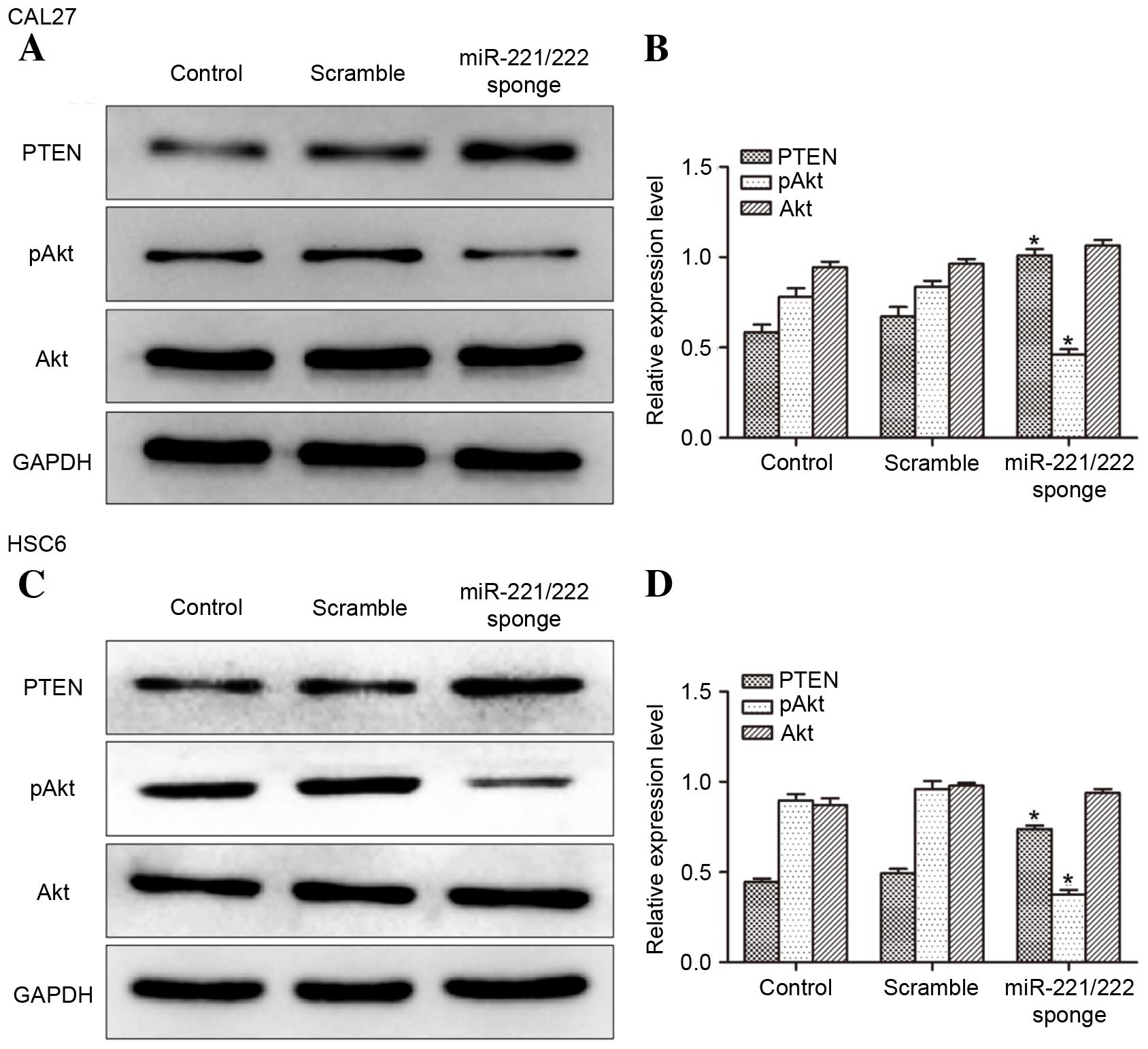
Determination of PTEN and pAkt
expression in CAL27 and HSC6 cells
To investigate the impact of miR-221/222 on protein
expression (Fig. 3) western blot
analysis was performed. It was observed that the level of PTEN
expression was increased and the expression of pAkt was decreased
in miR-221/222 sponge transfected CAL27 and HSC6 cells compared
with controls. The data suggest that the expression of PTEN in
CAL27 and HSC6 cells was negatively regulated by miR-221/222.
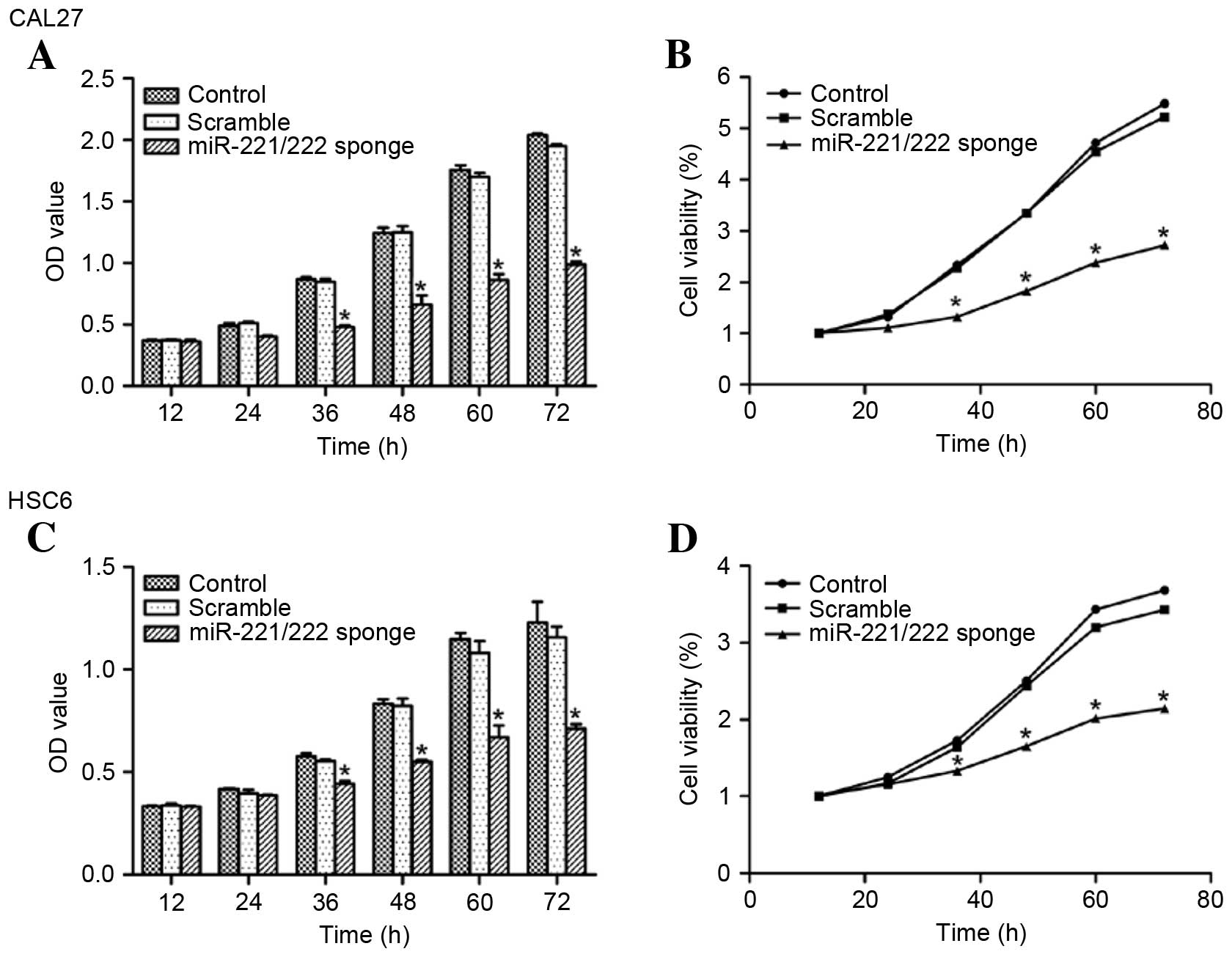
miR-221/222 sponge inhibits CAL27 and
HSC6 cell proliferation
The effect of miR-221/222 on the growth of CAL27 and
HSC6 cells was determined using a CCK8 assay. Compared with the
control group, the miR-221/222 sponge inhibited CAL27 and HSC6 cell
growth (P<0.05 vs. control) (Fig.
4).
miR-221/222 sponge induces CAL27 and
HSC6 cell invasion
The role of miR-221/222 on cell invasion was further
assessed by a Transwell assay. As shown in Fig. 5, the invasive ability of CAL27 and
HSC6 cells that were transfected with the miR-221/222 sponge was
significantly reduced when compared with the control groups
(P<0.01, P=0.0087 and P=0.00063 vs. control, respectively).
miR-221/222 sponge induces CAL27 and
HSC6 cell apoptosis
To examine the effect of the miR-221/222 sponge on
CAL27 and HSC6 cell apoptosis, Annexin V-PI analysis was performed.
The results showed that the number of apoptotic cells was
significantly increased following treatment with the miR-221/222
sponge, compared with the untreated groups, suggesting that
miR-221/222 sponge treatment induced apoptosis (P=0.00024 and
P=0.00003 vs. control, respectively) (Fig. 6).
Discussion
In previous years, increasing evidence has
highlighted the importance of miRNAs in the development and
progression of various types of cancer, including OSCC (25). miRNAs are considered to be novel
molecular targets in the diagnosis and treatment of human carcinoma
(1). Abnormal expression of
miR-221/222 has been observed and implicated in multiple neoplasms,
but to the best of our knowledge the equivalent research into OSCC
has not been performed (7,8,26).
PTEN was demonstrated to be a direct target gene of
miR-221/222, and the binding sites for miR-221/222 in the PTEN
3′UTR were predicted by bioinformatic analysis. Furthermore, the
results of the Dual-Luciferase Reporter Assay confirmed that PTEN
was a target gene of miR-221/222. To investigate further, it was
necessary to remove miR-221/222 function. This could be achieved
through an antisense oligonucleotide inhibitor, genetic knockout or
miRNA sponge (10,11,27). In
the present study, a miR-221/222 sponge was constructed in order to
downregulate the expression of miR-221/222. Results from RT-qPCR
confirmed that transfection with the miR-221/222 sponge
significantly reduced the miR-221/222 levels compared with the
control groups. miRNA sponge technology was initially introduced
and verified by Ebert et al (10) through the creation of RNAs containing
multiple binding sites complementary to the miRNA of interest,
leading to the continuous loss of miRNA function (28,29). The
sponge is usable in a long-term miRNA knockout experiment, while
maintaining an efficiency comparable with knockout by antisense
oligonucleotide inhibitors (10). It
is more convenient to use the sponge rather than perform a gene
knockout to build miRNA silencing cells or animal models.
Additionally, a sponge with common sequences could block a whole
miRNA family, which has the same seed regions (10,12).
Moreover, a sponge can be constructed to work on multiple miRNAs
synchronously by connecting different inhibiting sequences of
relevant miRNAs in series (10,12). It
should, however, be noted that the sponge may exhibit inefficient
inhibition at excessively high concentrations of miRNA, as the
sponge reaches maximum saturation (11). In previous studies, an miR-221/222
sponge was successfully constructed and its effect in OSCC cells
was investigated (12). In addition
to this, the results of the present study demonstrated that the
miR-221/222 sponge satisfactorarily inhibited the expression of
miR-221/222.
In the present study, it was identified that
miR-221/222 regulates OSCC cell proliferation, invasive ability and
apoptosis. The results of the present study show that CAL27 and
HSC6 cell proliferation and invasive ability was inhibited, and
apoptosis was promoted, when miR-221/222 expression was suppressed.
Yang et al (26) studied the
miR-221/222 expression in OSCC and demonstrated that upregulating
the expression of miR-221/222 increased the growth and
tumorigenesis of OSCC cells. Jiang et al (30) demonstrated that miR-222 targets the
p53 upregulated modulator of apoptosis expression, impacting OSCC
cell growth, apoptosis and invasion, which is consistent with the
results of the present study. The RT-qPCR and western blotting data
suggest that the expression of PTEN in CAL27 and HSC6 is negatively
correlated with miR-221/222 levels. The results demonstrated that
PTEN is a crucial functional target of miR-221/222 in OSCC
cells.
Previous studies have observed abnormal expression
of PTEN in OSCC and that its expression levels were correlated with
the degree of carcinoma differentiation (13,14,31). In
the present study, it was observed that knockdown of miR-221/222 in
CAL27 and HSC6 cells resulted in upregulation of PTEN and
downregulation of pAkt. PTEN has been identified as a tumor
suppressor and negative regulator of the PI3K/Akt signaling
pathway. The PI3K/Akt signaling pathway has been identified as an
oncogenic pathway and regulator of numerous cellular processes,
including proliferation, cell cycle, migration, invasion and
apoptosis (32,33). The results of the present study
suggest that OSCC cell proliferation, invasion and apoptosis are
associated with expression of PTEN. Activated PTEN dephosphorylates
phosphatidylinositol 3,4,5-trisphosphate to give
phosphatidylinositol 4,5-bisphosphate, and inhibits the
phosphorylation status of Akt, which results in decreased oncogenic
activity (34). Akt is a key molecule
in the PI3K signaling pathway, and the activity of Akt is tightly
regulated by its phosphorylation status (33). pAkt is activated in various types of
OSCC and has been confirmed to promote cell proliferation,
invasion, migration and reduce apoptosis (23,35).
Research has shown that loss of PTEN function may be a factor
leading to Akt signaling pathway activation and is consistent with
the results of the present study (14,36).
In the present study, it was demonstrated that PTEN
is a crucial functional target of miR-221/222 in OSCC cells. When
miR-221/222 is inhibited by the miR-221/222 sponge, the results
suggest that miR-221/222 has a regulatory function in OSCC cell
proliferation and invasion. Through the targeting of PTEN, the
PI3K/Akt signaling pathway may also be involved. The results of the
present study indicate a potential novel therapy for OSCC.
Acknowledgements
The present study was supported by the National
Natural Sciences Foundation of China (grant nos. 81272554 and
81472526), the Guangdong Natural Sciences Foundation (grant no.
S2011020003247) and the Guangdong Sciences and Technology Project
(grant nos. 2011B050400030 and 2012B031800387).
References
|
1
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ebrahimi F, Gopalan V, Smith RA and Lam
AK: miR-126 in human cancers: Clinical roles and current
perspectives. Exp Mol Pathol. 96:98–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang X, Han L, Zhang A, Wang G, Jia Z,
Yang Y, Yue X, Pu P, Shen C and Kang C: Adenovirus-mediated shRNAs
for co-repression of miR-221 and miR-222 expression and function in
glioblastoma cells. Oncol Rep. 25:97–105. 2011.PubMed/NCBI
|
|
5
|
Mittal SP, Mathai J, Kulkarni AP, Pal JK
and Chattopadhyay S: miR-320a regulates erythroid differentiation
through MAR binding protein SMAR1. Int J Biochem Cell Biol.
45:2519–2529. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miller TE, Ghoshal K, Ramaswamy B, Roy S,
Datta J, Shapiro CL, Jacob S and Majumder S: MicroRNA-221/222
confers tamoxifen resistance in breast cancer by Targeting p27Kip1.
J Biol Chem. 283:29897–29903. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang C, Zhang J, Zhang A, Wang Y, Han L,
You Y, Pu P and Kang C: PUMA is a novel target of miR-221/222 in
human epithelial cancers. Int J Oncol. 37:1621–1626.
2010.PubMed/NCBI
|
|
8
|
Quintavalle C, Garofalo M, Zanca C, Romano
G, Iaboni M, del Basso De Caro M, Martinez-Montero JC, Incoronato
M, Nuovo G, Croce CM and Condorelli G: miR-221/222 overexpession in
human glioblastoma increases invasiveness by targeting the protein
phosphate PTPµ. Oncogene. 31:858–868. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jikuzono T, Kawamoto M, Yoshitake H,
Kikuchi K, Akasu H, Ishikawa H, Hirokawa M, Miyauchi A, Tsuchiya S,
Shimizu K and Takizawa T: The miR-221/222 cluster, miR-10b and
miR-92a are highly upregulated in metastatic minimally invasive
follicular thyroid carcinoma. Int J Oncol. 42:1858–1868.
2013.PubMed/NCBI
|
|
10
|
Ebert MS, Neilson JR and Sharp PA:
MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian
cells. Nat Methods. 4:721–726. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ebert MS and Sharp PA: MicroRNA sponges:
Progress and possibilities. RNA. 16:2043–2050. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou LJ, Zhao W, Jiang FF, Liu ZF, Ou Yang
Y and Yu DS: Construction and validation of microRNA-221/222 sponge
vector. Chinese Journal of Stomatological Research (Electronic
Version). 106–112. 2015.
|
|
13
|
Rahmani A, Alzohairy M, Babiker AY, Rizvi
MA and Elkarimahmad HG: Clinicopathological significance of PTEN
and bcl2 expressions in oral squamous cell carcinoma. Int J Clin
Exp Pathol. 5:965–971. 2012.PubMed/NCBI
|
|
14
|
Wang H, Wu Q, Liu Z, Luo X, Fan Y, Liu Y,
Zhang Y, Hua S, Fu Q, Zhao M, et al: Downregulation of FAP
suppresses cell proliferation and metastasis through PTEN/PI3K/AKT
and Ras-ERK signaling in oral squamous cell carcinoma. Cell Death
Dis. 5:e11552014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dorr C, Janik C, Weg M, Been RA, Bader J,
Kang R, Ng B, Foran L, Landman SR, O'Sullivan MG, et al: Transposon
mutagenesis screen identifies potential lung cancer drivers and
CUL3 as a tumor suppressor. Mol Cancer Res. 13:1238–1247. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin PC, Lin JK, Lin HH, Lan YT, Lin CC,
Yang SH, Chen WS, Liang WY, Jiang JK and Chang SC: A comprehensive
analysis of phosphatase and tensin homolog deleted on chromosome 10
(PTEN) loss in colorectal cancer. World J Surg Oncol. 13:1862015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gwak HS, Kim TH, Jo GH, Kim YJ, Kwak HJ,
Kim JH, Yin J, Yoo H, Lee SH and Park JB: Silencing of MicroRNA-21
confers radio-sensitivity through Inhibition of the PI3K/AKT
pathway and enhancing autophagy in malignant glioma cell lines.
PLoS One. 7:e474492012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sarkar S, Dubaybo H, Ali S, Goncalves P,
Kollepara SL, Sethi S, Philip PA and Li Y: Down-regulation of
miR-221 inhibits proliferation of pancreatic cancer cells through
up-regulation of PTEN, p27(kip1), p57(kip2), and PUMA. Am J Cancer
Res. 3:465–477. 2013.PubMed/NCBI
|
|
19
|
Fang L, Li H, Wang L, Hu J, Jin T, Wang J
and Yang BB: MicroRNA-17-5p promotes chemotherapeutic drug
resistance and tumour metastasis of colorectal cancer by repressing
PTEN expression. Oncotarget. 5:2974–2987. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Beg S, Siraj AK, Prabhakaran S, Jehan Z,
Ajarim D, Al-Dayel F, Tulbah A and Al-Kuraya KS: Loss of PTEN
expression is associated with aggressive behavior and poor
prognosis in Middle Eastern triple-negative breast cancer. Breast
Cancer Res Treat. 151:541–553. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun MM, Zhang MZ, Chen Y, Li SL, Zhang W,
Ya GW and Chen KS: Effect of PTEN antisense oligonucleotide on
oesophageal squamous cell carcinoma cell lines. J Int Med Res.
40:2098–2108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xue Q, Sun K, Deng HJ, Lei ST, Dong JQ and
Li GX: Anti-miRNA-221 sensitizes human colorectal carcinoma cells
to radiation by upregulating PTEN. World J Gastroentero.
19:9307–9317. 2013. View Article : Google Scholar
|
|
23
|
Jia LF, Huang YP, Zheng YF, Lyu MY, Wei
SB, Meng Z and Gan YH: miR-29b suppresses proliferation, migration,
and invasion of tongue squamous cell carcinoma through PTEN-AKT
signaling pathway by targeting Sp1. Oral Oncol. 50:1062–1071. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang D, Ni Z, Xu X and Xiao J: MiR-32
Functions as a tumor suppressor and directly targets EZH2 in human
oral squamous cell carcinoma. Med Sci Monit. 20:2527–2535. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang C, Shen WG, Liu CJ, Chen YW, Lu HH,
Tsai MM and Lin SC: miR-221 and miR-222 expression increased the
growth and tumorigenesis of oral carcinoma cells. J Oral Pathol
Med. 40:560–566. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krützfeldt J, Rajewsky N, Braich R, Rajeev
KG, Tuschl T, Manoharan M and Stoffel M: Silencing of microRNAs in
vivo with ‘antagomirs’. Nature. 438:685–689. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kluiver J, Slezak-Prochazka I,
Smigielska-Czepiel K, Halsema N, Kroesen BJ and van den Berg A:
Generation of miRNA sponge constructs. Methods. 58:113–117. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kluiver J, Gibcus JH, Hettinga C, Adema A,
Richter MK, Halsema N, Slezak-Prochazka I, Ding Y, Kroesen BJ and
van den Berg A: Rapid generation of MicroRNA Sponges for MicroRNA
Inhibition. PLoS One. 7:e292752012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang F, Zhao W, Zhou L, Zhang L, Liu Z
and Yu D: miR-222 regulates the cell biological behavior of oral
squamous cell carcinoma by targeting PUMA. Oncol Rep. 31:1255–1262.
2014.PubMed/NCBI
|
|
31
|
Kurasawa Y, Shiiba M, Nakamura M, Fushimi
K, Ishigami T, Bukawa H, Yokoe H, Uzawa K and Tanzawa H: PTEN
expression and methylation status in oral squamous cell carcinoma.
Oncol Rep. 19:1429–1434. 2008.PubMed/NCBI
|
|
32
|
Cohen Y, Goldenberg-Cohen N, Shalmon B,
Shani T, Oren S, Amariglio N, Dratviman-Storobinsky O,
Shnaiderman-Shapiro A, Yahalom R, Kaplan I and Hirshberg A:
Mutational analysis of PTEN/PIK3CA/AKT pathway in oral squamous
cell carcinoma. Oral Oncol. 47:946–950. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu YZ, Wu K, Huang J, Liu Y, Wang X, Meng
ZJ, Yuan SX, Wang DX, Luo JY, Zuo GW, et al: The PTEN/PI3K/Akt and
Wnt/β-catenin signaling pathways are involved in the inhibitory
effect of resveratrol on human colon cancer cell proliferation. Int
J Oncol. 45:104–112. 2014.PubMed/NCBI
|
|
34
|
Giudice FS and Squarize CH: The
determinants of head and neck cancer: Unmasking the PI3K pathway
mutations. J Carcinog Mutagen. (Suppl 5): pii: 003. 2013.PubMed/NCBI
|
|
35
|
Xie Q, Yan Y, Huang Z, Zhong X and Huang
L: MicroRNA-221 targeting PI3-K/Akt signaling axis induces cell
proliferation and BCNU resistance in human glioblastoma.
Neuropathology. 34:455–464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao F, Huang W, Zhang Y, Tang S, Zheng L,
Ma F, Wang Y, Tang H and Li X: Hes1 promotes cell proliferation and
migration by activating Bmi-1 and PTEN/Akt/GSK3β pathway in human
colon cancer. Oncotarget. 6:38667–38680. 2015.PubMed/NCBI
|