Introduction
Breast cancer is the most common carcinoma in women,
with low survival rates in patients due to metastatic lesions
(1,2).
Triple negative breast cancer (TNBC) is an aggressive breast cancer
subtype in which the tumor cells lack expression of the estrogen,
progesterone and human epidermal growth factor 2 receptors. TNBC
has a high rate of relapse and metastasis, and accounts for
approximately 12–17% of all breast cancer cases (1,3). Due to
poor prognosis and a lack of treatment options, TNBC patients have
a disproportionately high mortality rate: No more than 30% of
patients with metastatic TNBC survive for 5 years (4). Therefore, understanding the mechanism
regulating TNBC progression may assist with the development of
accurate prognosticators and more effective treatments.
Cancer comprises a collection of complicated genetic
and epigenetic alterations that arise via multistep processes
(5–7).
Ubiquitin-specific proteases (USPs) are frequently involved in
cancer regulation, as oncogenic mutations in USP genes are able to
disrupt deubiquitination of proteins that control cell growth and
apoptosis (8–10). To date, 50 USP family members have
been identified in humans (11).
USP21 is able to facilitate initiation of transcriptional activity
via catalyzing the hydrolysis of the ubiquitylation of histone H2A
(12). USP21 is also able to regulate
the stability of proteins through deubiquitination. For example,
USP21 can mediate deubiquitination of GATA3 and maintain GATA3
expression in regulatory T cells (13). Thus, USP proteins may serve as a good
point of intervention for the prevention of cancer and other
mutation-associated diseases (14).
Nucleotide oligomerization domain (NOD)-like
receptor (NLR) signaling pathways have a significant role in
numerous human diseases, including bacterial infections, autoimmune
and inflammatory disorders, and cancer (15). Stimulation of NLRs results in the
activation of nuclear factor (NF)-κB and mitogen-activated protein
kinases (MAPKs), which drive the transcription of numerous genes
involved in both innate and adaptive immune responses (16). Previously, USPs have been shown to
participate in the regulation of the NF-κB signaling pathway
(17,18). For example, USP4 promotes stimulation
of NF-κB mediated by tumor necrosis factor α (TNF-α) through
deubiquitination-dependent downregulation of TGFβ-activated kinase
1 (19). USP11 is able to modulate
TNF-α-induced NF-κB activation through regulation of nuclear factor
of κ light polypeptide gene enhancer in B-cells inhibitor α (IκBα)
stability (20). By contrast, USP21
inhibits TNF-α-induced NF-κB signaling by promoting the
deubiquitination of receptor-interacting protein 1 (RIP1) in HeLa
cells (21). However, the role of
USP21 in breast cancer remains to be elucidated.
In the present study, bioinformatics tools were used
to study data online and characterize the gene alteration status of
USP family members in breast cancer. Subsequently, the expression
of the most altered member, USP21, was validated in vitro.
The expression of USP21 was then knocked down using small
interfering RNA (siRNA) in TNBC cell lines, and cellular
experiments were performed to investigate its biological function,
in the hope that the results may provide useful insights into the
prognosis and treatment of TNBC.
Materials and methods
Cell culture and siRNA
transfection
All breast cancer cells were obtained from American
Type Culture Collection (Manassas, VA, USA) and maintained in
Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1% PenStrep
(100 U/ml penicillin and 100 µg/ml streptomycin; Invitrogen; Thermo
Fisher Scientific, Inc.), in an incubator at 37°C with 5%
CO2. MDA-MB-231 or MDA-MB-157 cells were plated onto
tissue culture plates 24 h prior to transfection. Transient
transfection of siUSP21 (siUSP21-1: 5′-GCUAGAAGAACCUGAGUUA-3′;
siUSP21-2: 5′-GAGCUGUCUUCCAGAAAUA-3′) or siControl
(5′-GUACUUGUACUCCAGCUUUGUG-3′) (Invitrogen; Thermo Fisher
Scientific, Inc.) at a final concentration of 50 nM was
accomplished with Lipofectmine® 2000 reagent according
to the manufacturer's protocol (Invitrogen; Thermo Fisher
Scientific, Inc.).
Cell migration assay
For scratch wound-healing assays, 48 h following
siRNA transfection, 1.5×105 cells were seeded into
six-well plates and serum starved for 24 h. Cells were wounded by
scratching with a pipette tip and cultured in medium containing 0.5
g/ml mitomycin-C (Sigma-Aldrich; EMD Millipore, Billerica, MA, USA)
to block cell division. Cells were photographed at 0, 5 and 10 h
timepoints. The distance of migration was calculated using ImageJ
software (version 1.46; imagej.nih.gov/ij/). Matrigel invasion assay was
performed using a Corning® BioCoat™ Matrigel Invasion
Chamber (Corning Incorportated, Corning, NY, USA). Tumor cells
(1×105) treated with control or USP21 siRNA in 200 µl
serum-free DMEM were placed in the upper chamber. The lower chamber
was filled with 600 µl conditioned medium (DMEM medium containing
1% FBS for 24 h) as chemoattractant. After 24 h of incubation at
37°C, the cells on the upper surface of the filter were removed
with a cotton swab. The cells that had invaded the Matrigel and
reached the lower surface of the filter were fixed in methanol,
stained with hematoxylin and eosin, and counted under
magnification, ×400. A total of five fields were randomly selected
and the number of invasive cells was counted.
Protein isolation, western blot
analysis and co-immunoprecipitation (Co-IP)
Cells were lysed with radioimmunoprecipitation assay
buffer (Sigma-Aldrich; EMD Millipore) as previously described
(22–24). Cells were centrifuged at 4°C for 10
min at 16,000 × g. Protein concentrations were determined by
the Bradford assay (25). Aliquots
containing 20 µg of total protein were separated by 10% sodium
dodecyl-polyacrylamide gel electrophoresis and transferred to
nitrocellulose membranes. Blots were probed with primary antibodies
against USP21 (1:1,000; goat polyclonal; sc-79305; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and β-actin (1:5,000, mouse
monoclonal; A5316; Sigma-Aldrich; EMD Millipore). Appropriate
secondary antibodies (1:3,000; rabbit anti-goat; ab6741; 1:5,000;
rabbit anti-mouse; ab97046; Abcam, Cambridge, MA, USA) conjugated
to horseradish peroxidase and enhanced chemiluminescence (GE
Healthcare Life Sciences, Chalfont, UK) were used to detect the
bound primary antibodies. Co-IP was performed with cell lysate (500
µg) incubated with USP21 (1:1,000; goat polyclonal; sc-79305; Santa
Cruz Biotechnology, Inc.), relA (1:1,000; rabbit polyclonal;
sc-372; Santa Cruz Biotechnology, Inc.) or non-specific-IgG
antibodies (1:1,000; rabbit IgG, monoclonal; ab172730; Abcam) using
µMACS™ Protein A/G MicroBeads and MACS® Separation
Columns according to the manufacturer's protocol (Miltenyi Biotec,
Auburn, USA).
RNA isolation, cDNA preparation and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR)
As previously described (26–28), total
RNA was isolated from cells using the RNeasy Mini kit (Qiagen,
Inc., Valencia, CA, USA). cDNA was prepared from 1 µg of RNA using
the iScript cDNA Synthesis kit (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). RT-qPCR was performed using a GeneAmp Gold RNA
PCR Core kit (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and an iCycler iQ™ Real-Time PCR Detection System (Bio-Rad
Laboratories, Inc.), where each 20 µl reaction included 1% cDNA
preparation, 0.5 µM primers and 10 µl SYBR Green (Bio-Rad
Laboratories, Inc.). Primer sequences are presented in Table I. Expression of
glyceraldehyde-3-phosphate dehydrogenase was used to normalize the
gene expression level. The relative difference in the expression
level was calculated using the ΔΔCq method (29). The data presented are representative
of three independent biological repeats each assayed in triplicate
and show the relative expression levels.
 | Table I.Reverse transcription-qPCR primer
sequences. |
Table I.
Reverse transcription-qPCR primer
sequences.
| Gene name | F/R | Sequences of the
qPCR primer pairs (5′-3′ direction) |
|---|
| GAPDH | F |
GGTGAAGGTCGGAGTCAACGG |
| GAPDH | R |
GAGGTCAATGAAGGGGTCATTG |
| USP21 | F |
ATCTCGGACCAACTTAGCCC |
| USP21 | R |
GTGCCCTCCCAAGGCAATC |
| IL8 | F |
TTTTGCCAAGGAGTGCTAAAGA |
| IL8 | R |
AACCCTCTGCACCCAGTTTTC |
| CARD8 | F |
GAAGCGAAACTGCATATTCTGGT |
| CARD8 | R |
GGGTTGGAAGAGGCATGGC |
| CCL2 | F |
CAGCCAGATGCAATCAATGCC |
| CCL2 | R |
TGGAATCCTGAACCCACTTCT |
| IL6 | F |
ACTCACCTCTTCAGAACGAATTG |
| IL6 | R |
CCATCTTTGGAAGGTTCAGGTTG |
| CXCL1 | F |
GCGGAAAGCTTGCCTCAA |
| CXCL1 | R |
TCAGCATCTTTTCGATGATTTTCTT |
| NLRP3 | F |
GATCTTCGCTGCGATCAACAG |
| NLRP3 | R |
CGTGCATTATCTGAACCCCAC |
| NFKB1A | F |
AACAGAGAGGATTTCGTTTCCG |
| NFKB1A | R |
TTTGACCTGAGGGTAAGACTTCT |
Microarray hybridization data analysis
and The Cancer Genome Atlas (TCGA) analysis
MDA-MB-231 cells were plated onto a 6-well plate at
70% confluence and transfected with siControl or siUSP21. A total
of 48 h subsequent to transfection, the cells were washed with
phosphate-buffered saline, and total RNA was obtained using the
RNeasy Mini kit (Qiagen, Inc.) and treated with 1 unit of DNase I
(Qiagen, Inc.). Expression profiles were generated by hybridizing
10 µg of total RNA to GeneChip® Human Genome U133
Plus2.0 Gene Chips (Affymetrix, Inc., Santa Clara, CA, USA)
according to the Affymetrix Eukaryote One-cycle protocol (30). Briefly, 5–10 µg of total RNA were used
to generate biotinylated cDNA, which was fragmented and hybridized
to a chip for 16 h at 45°C in a GeneChip Hybridization Oven 640
(Affymetrix, Inc.). Arrays were then washed and stained on a
GeneChip Fluidics Station 450 (Affymetrix, Inc.) and subsequently
scanned on a GeneChip Scanner 3000 7G (Affymetrix, Inc.) to obtain
fluorescence intensities. To eliminate data with low reliability,
genes whose expression was regarded as absent in these cell lines
as a result of software analysis were excluded. Following
identification of the differentially expressed genes, DAVID online
software (david.ncifcrf.gov/) was used to
perform Kyoto Encyclopedia of Genes and Genomes pathway significant
enrichment analysis of 1,007 differentially expressed genes
associated with USP21, and the six signaling pathways with the
smallest P-values were selected. For TCGA analysis, data including
1,105 breast invasive carcinoma samples from 1,098 patients were
obtained from the TCGA website using the cBioPortal tool
(www.cbioportal.org/).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analyses were performed using SPSS version 19.0
software (IBM SPSS, Armonk, NY, USA). Statistically significant
differences were determined by Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
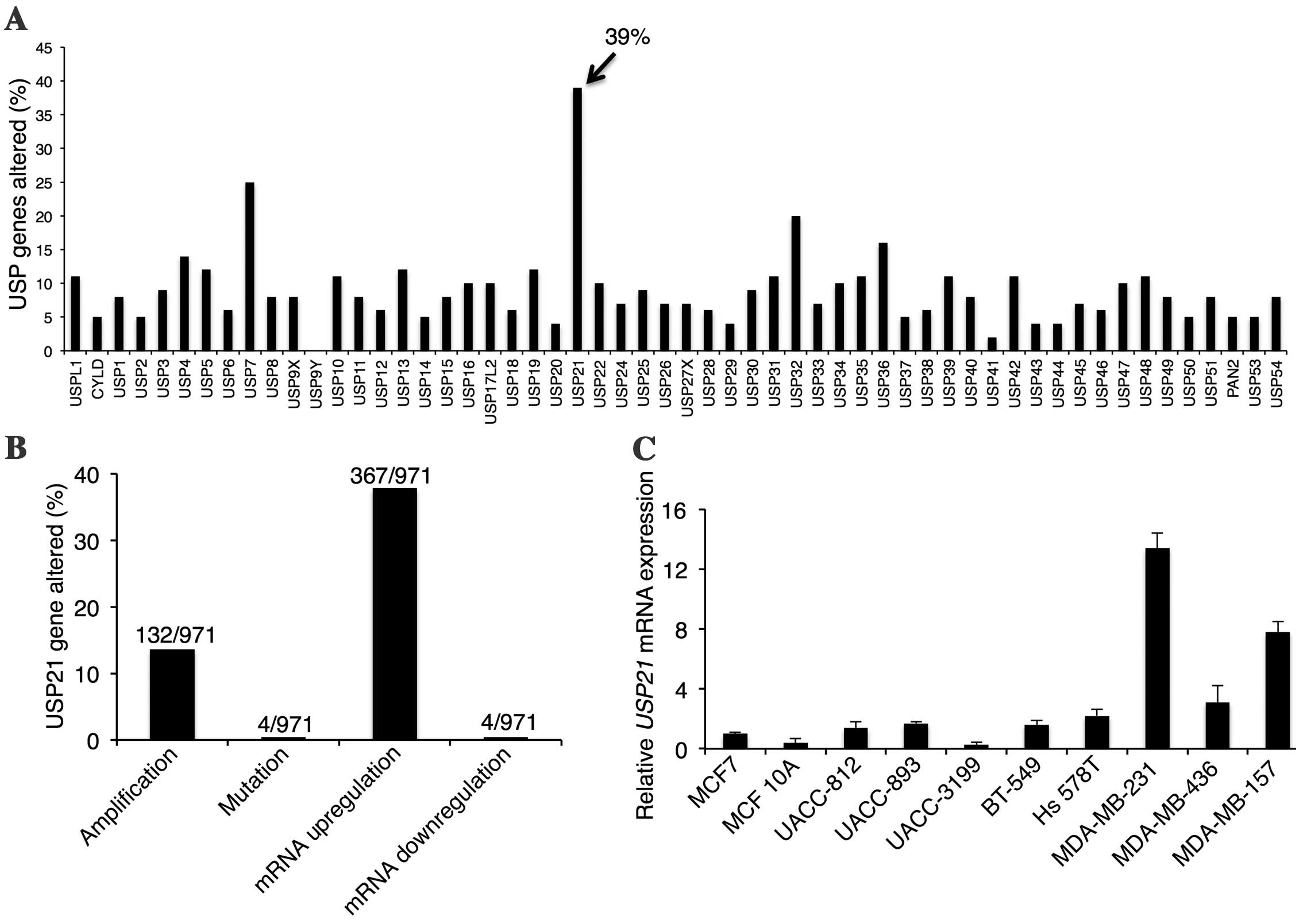
USP21 is the most upregulated USP
family member in breast cancer
USP family proteins have a significant role in
multiple signaling and cell regulatory networks in breast cancer
(9). To characterize the extent of
changes in all 50 USP genes in breast cancer, the present study
generated an alteration-summary for these genes in breast cancer
using the cBioPortal tool for Cancer Genomics (31,32)
(Fig. 1A). The results of the present
study demonstrated that USP21 was altered in 39% (375/971 samples)
of patients with breast invasive carcinomas, indicating it is the
most altered gene among all the USP gene family members. In all the
deregulation situations, including gene copy number amplification
and mRNA expression alteration, of USP21, 13.6% (132/971) of the
patients displayed copy number amplification, while 37.8% (367/971)
of the patients showed mRNA upregulation (Fig. 1B), suggesting a role for USP21 in
breast cancer.
To investigate the expression of USP21 in breast
cancer cell lines in vitro, the present study analyzed a
panel of 10 breast cancer cell lines including luminal, basal and
non-cancerous breast epithelial lines. Notably, it was observed
that the expression of USP21 was enriched in triple negative cell
lines, MDA-MB-231 and MDA-MB-157 (Fig.
1C). Therefore, it appears that USP21 is required for the
cancerous ability of TNBC cells.
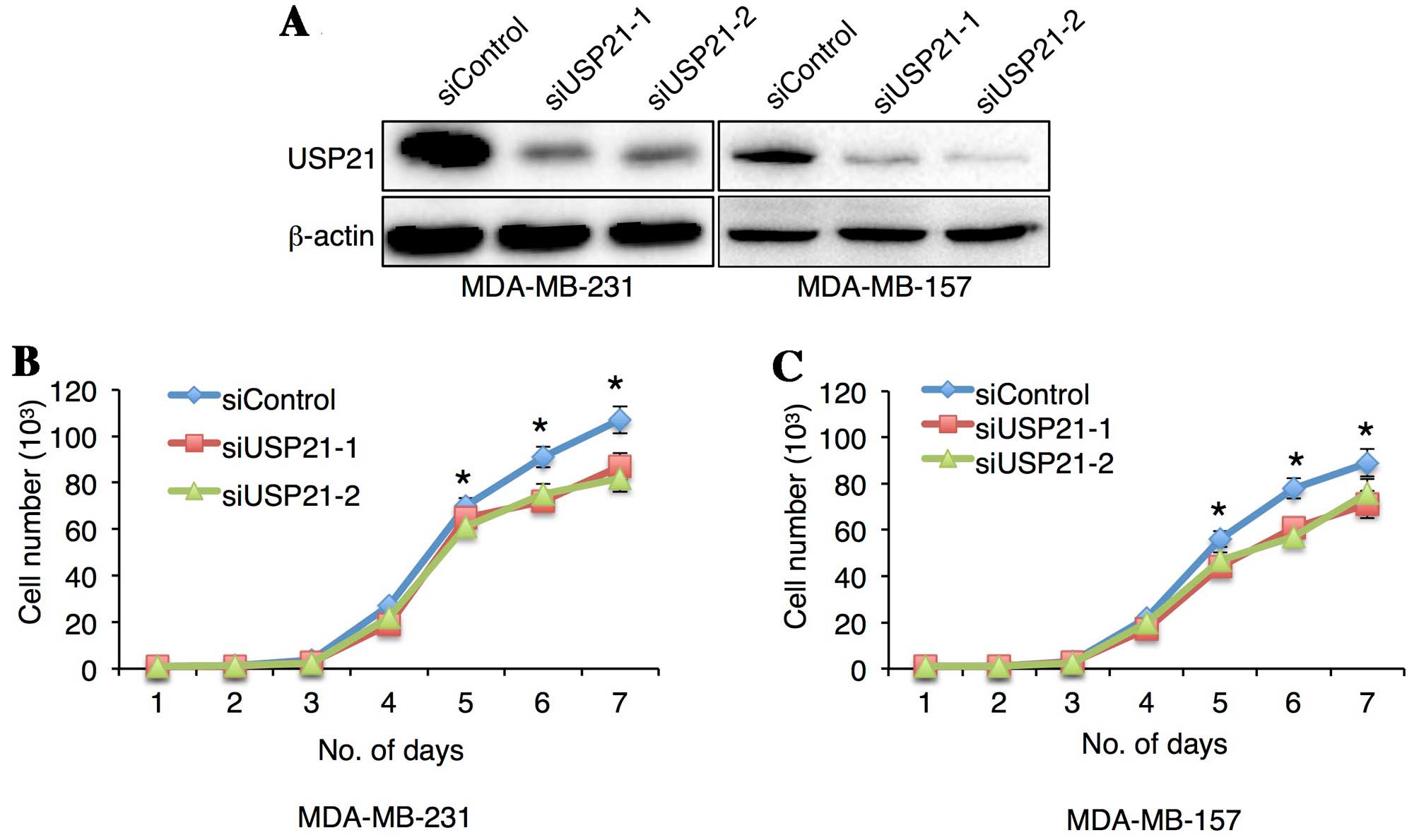
USP21 affects TNBC cell proliferation,
migration and invasion
To directly investigate the contribution of USP21 to
breast tumorigenesis, the present study knocked down USP21 protein
using two specific siRNAs in MDA-MB-231 and MDA-MB-157 cells.
Western blot analysis confirmed the knockdown of USP21 under these
conditions (Fig. 2A). It was
initially investigated whether USP21 is crucial to the
proliferation of these cells. Cells transfected with control or
USP21-siRNAs were cultured for up to 7 days. The difference in cell
proliferation was measured by viable cell counting from day 1 to
day 7. Knockdown of USP21 reduced the proliferation of MDA-MB-157
and MDA-MB-231 cells 5 days after siRNA transfection (Fig. 2B and C).
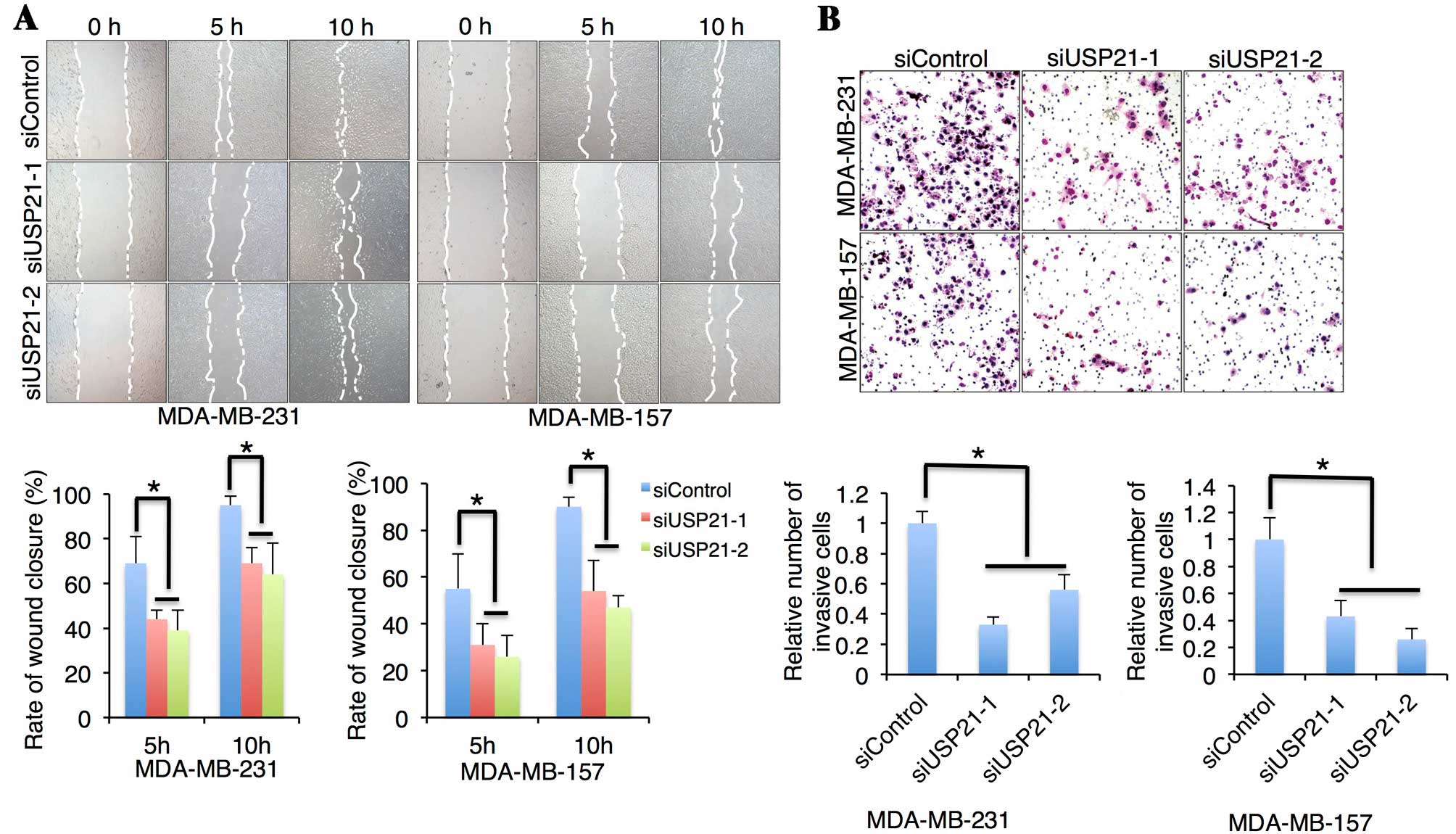
To determine the effects of USP21 on cell migration,
a wound-healing assay was performed following USP21 silencing. The
scratch wounds were almost identical sizes in each experimental
group at 0 h; however, knockdown of USP21 using two different
siRNAs markedly decreased the migration ability of MDA-MB-231 and
MDA-MB-157 cells at 5 and 10 h (Fig.
3A). The present study measured the distance between the
migrating frontlines and calculated the rate of wound closure. It
was observed that the USP21 silenced groups were less effective
than the control group in terms of the healing process (Fig. 3A).
To investigate the role of USP21 in TNBC cell
invasion, the present study measured the invasive ability using
invasion assays. Consistent with the migration assay results,
downregulation of USP21 in MDA-MB-231 and MDA-MB-157 cells by siRNA
knockdown resulted in a marked reduction in the cell invasive
capability compared to the control group (Fig. 3B). These results suggested that USP21
is involved in TNBC cancer growth, migration and invasion.
Microarray data analysis indicates
that USP21 expression is associated with NOD-like receptor
signaling pathway
To further investigate USP21-mediated gene
expression changes, the cDNA from USP21-knockdown and control
MDA-MB-231 cells was subjected to microarray analyses. A total of
1,007 genes in USP21 siRNA treated cells exhibited a differential
expression more than double that of the control sample. A total of
309 genes were upregulated in the USP21 knockdown sample compared
with the control, while 698 genes demonstrated decreased expression
in the USP21 siRNA treated sample. Furthermore, it was observed
that the major signaling pathways of the differentially expressed
genes were associated with the following molecular pathways:
NOD-like receptor signaling, TGF-β signaling pathway, RNA
degradation, small cell lung cancer, pathways in cancer and
extracellular matrix-receptor interaction. It was observed that
USP21 depletion markedly attenuated genes specifically associated
with the NOD-like receptor signaling pathway [interleukin (IL)6,
NLR family, pyrin domain containing (NLRP)3, IκBα and chemokine
(C-X-C motif) ligand (CXCL)8] and stimulated genes associated with
the TGF-β signaling pathway (bone morphogenetic protein (BMP)4, BMP
type IB receptor, ID1, ID2, ID3, ID4 and TGF-β receptor 1)
(Fig. 4A and B). To confirm the
microarray results, 8 genes were selected (USP21, IL8, caspase
recruitment domain family member 8 (CARD8), chemokine (C-C motif)
ligand 2 (CCL2), IL6, CXCL1, NLRP3 and IκBα) and their expression
was measured with RT-qPCR on the same sample used for microarray
studies. All of these genes exhibited moderate to high expression
and demonstrated high concordance between microarray and PCR data
(Fig. 4C).
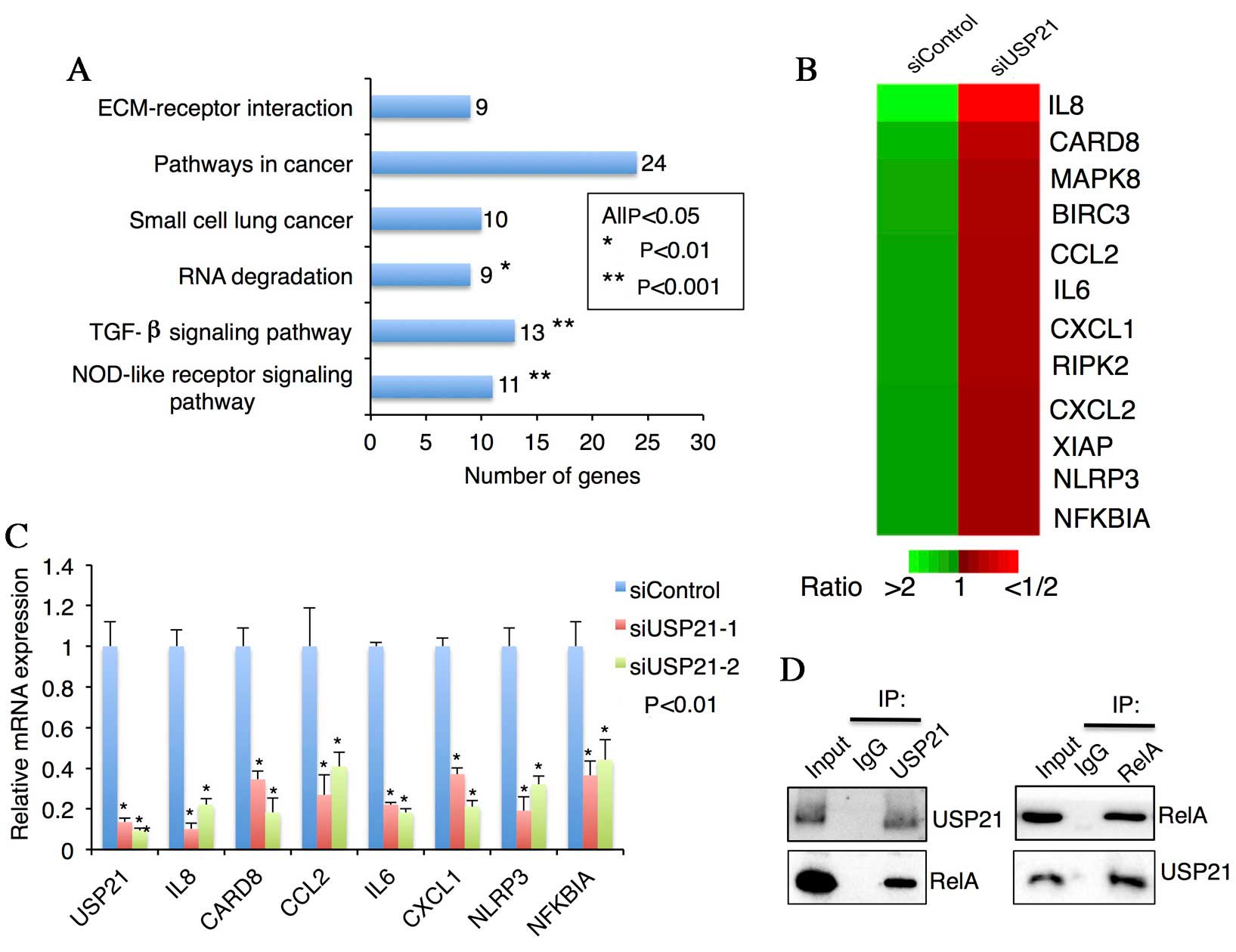 | Figure 4.Global gene profiling of MDA-MB-231
following USP21 knockdown. (A) Summary of the kyoto enclyclopedia
of genes and genomes pathways of genes significantly enriched in
response to USP21 knockdown, using database for annotation,
visualization and integrated discovery software. (B) Changes in
gene expression levels in MDA-MB-231 cells following USP21
knockdown. The heat map depicts relative gene expression changes
(siRNA control/siUSP21-1). (C) Confirmation of downregulated genes
from the microarray dataset by reverse transcription-quantitative
polymerase chain reaction. *P<0.05. (D) Co-immunoprecipitation
of USP21 and relA in MDA-MB-231 cells. USP21, ubiquitin specific
protease 21; ECM, extracellular matrix; TGF, transforming growth
factor; NOD, nucleotide-binding oligomerization domain; siRNA,
small interfering RNA; IL, interleukin; CARD8, caspase recruitment
domain family member 8; MAPK8, mitogen-activated protein kinase 8;
BIRC3, baculoviral IAP repeat containing 3; CCL2, chemokine (C-C
motif) ligand 2; CXCL, chemokine (C-X-C motif) ligand; RIPK2,
receptor interacting serine/threonine kinase 2; XIAP, X-linked
inhibitor of apoptosis protein; NLRP3, NLR family pyrin domain
containing 3; NFKBIA, nuclear factor of κ light polypeptide gene
enhancer in B-cells inhibitor α; IP, immunoprecipitation. |
As USP21 has been reported to be a histone H2A
deubiquitinase that initiates transcriptional activity (12), the present study screened the
association of USP21 with NOD-like receptor associated
transcription factors, including NF-κB, activator protein 1 or
interferon regulatory factors. Notably, it was observed that relA,
an essential subunit of the transcriptionally active NF-κB dimer,
may be ‘pulled down’ with USP21 using a co-IP assay (Fig. 4D). Thus, these results indicate that
USP21 may associate with NF-κB transcription factors.
Discussion
TNBC is an aggressive and deadly subtype of breast
cancer and lacks targeted therapies (33). In the present study, it was observed
that USP21 is the most altered USP member in breast cancer using
online TCGA data sets. Furthermore, the present study demonstrated,
for the first time to the best of our knowledge, that silencing of
USP21 leads to impaired proliferation, migration and invasion
ability of TNBC cells, which indicates that USP21 may be involved
in tumor metastasis. The present study also investigated global
gene profiling upon depletion of USP21 in TNBC cells. The results
of the present study revealed that a subset of genes involved in
NLR signaling pathways were significantly repressed when USP21 was
knocked down in TNBC cells. It was also observed that USP21 was
associated with relA, implying a link between USP21 and NF-κB in
regulating NLR signaling and TNBC progression.
The NLR signaling pathway plays a vital role in
human diseases, including cancer (15). In the current study, it was
demonstrated that silencing of USP21 repressed several NLR
signaling pathway factors, including IL6, IL8, CCL2, CXCL1, NLRP3,
IκBα and CARD8. A number of these genes are involved in TNBC
regulation. For example, inhibition of IL-6 and IL-8 expression in
TNBC led to a decrease in colony formation and cell survival in
vitro and inhibited tumor engraftment and growth in vivo
(34). In addition, RIPK2 can
stimulate triple-negative breast cancer cell migration and invasion
through NF-κB and c-Jun N-terminal kinase signaling pathways
(35). Thus, it appears likely that
the impaired tumorigenic ability in USP21 depleted TNBC cells may
be directly associated with this downregulation of NLR signaling
pathway members.
Several studies have demonstrated a role of USP21 in
inflammation and the NF-κB signaling pathway (21,36,37). It
has been previously shown that USP21 is able to regulate the
expression of IL8 and cancer stem cell properties in human renal
cell carcinoma (28). IL8 has been
demonstrated to be an important cytokine that is required for
growth of TNBC (34), supporting the
results of the present study. Through deubiqutinating RIP1, USP21
is able to repress TNFα-induced NF-κB activation in HeLa cells,
suggesting that USP21 may serve as a negative regulator of the
NF-κB signaling pathway (21). An
additional study has revealed that depletion of USP21 decreases
IL33 protein levels and IL33-mediated NF-κB p65 promoter activity,
indicating USP21 is able to positively regulate the NF-κB signaling
pathway (36). Therefore, although
USP21 is involved in NF-κB signaling transduction activity, the
function of USP21 appears to be context dependent. Based on the
fact that NLR signaling pathway components were repressed upon
USP21 depletion, and USP21 was associated with relA, it is likely
USP21 is a positive regulator of the NF-κB signaling pathway in
TNBC cells. If that is the case, the global regulation of histone
deubiquitination by USP21 requires further investigation. IL6 and
IL8 have critical roles in anchorage-independent growth of TNBC,
and function through the NF-κB signaling pathway (34), meaning that NF-κB may be a potential
therapeutic target in TNBC. A previous report demonstrated that
NF-κB regulates cancer stem cell populations in the basal-like
breast cancer subtype of TNBC (38).
In addition, TGF-β ligands are often enriched in the TNBC tumor
microenvironment and have a role in breast cancer stem cells
(39,40). The results of the present study
suggest that USP21 may serve as a modulator for the TGF-β signaling
pathway in TNBC.
In conclusion, USP21 was observed to be elevated in
breast cancer patient samples. The expression of USP21 may promote
proliferation, migration and invasion of breast cancer cells.
Therefore, USP21 may have a potential role in the prognosis of and
be a relevant target in breast cancer.
Acknowledgements
The present research was supported by the China
National Nature Science Youth Fund (grant no. 30600613).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zang QQ, Zhang L, Gao N and Huang C:
Ophiopogonin D inhibits cell proliferation, causes cell cycle
arrest at G2/M, and induces apoptosis in human breast carcinoma
MCF-7 cells. J Integr Med. 14:51–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Foulkes WD, Smith IE and Reis-Filho JS:
Triple-negative breast cancer. N Engl J Med. 363:1938–1948. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lehmann BD, Bauer JA, Chen X, Sanders ME,
Chakravarthy AB, Shyr Y and Pietenpol JA: Identification of human
triple-negative breast cancer subtypes and preclinical models for
selection of targeted therapies. J Clin Invest. 121:2750–2767.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liang Y, Zhu F, Zhang H, Chen D, Zhang X,
Gao Q and Li Y: Conditional ablation of TGF-β signaling inhibits
tumor progression and invasion in an induced mouse bladder cancer
model. Sci Rep. 6:294792016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun S, Xu A, Yang G and Cheng Y: Galanin
is a novel epigenetic silenced functional tumor suppressor in renal
cell carcinoma. Cancer Translational Medicine. 1:183–187. 2015.
View Article : Google Scholar
|
|
7
|
Cheng Y, Tu Y and Liang P: Promoter
methylated tumor suppressor genes in glioma. Cancer Translational
Medicine. 1:123–130. 2015. View Article : Google Scholar
|
|
8
|
Mani A and Gelmann EP: The
ubiquitin-proteasome pathway and its role in cancer. J Clin Oncol.
23:4776–4789. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pal A and Donato NJ: Ubiquitin-specific
proteases as therapeutic targets for the treatment of breast
cancer. Breast Cancer Res. 16:4612014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen D, Dai C and Jiang Y: Histone H2A and
H2B deubiquitinase in developmental disease and cancer. Cancer
Translational Med. 1:170–175. 2015. View Article : Google Scholar
|
|
11
|
Reyes-Turcu FE, Ventii KH and Wilkinson
KD: Regulation and cellular roles of ubiquitin-specific
deubiquitinating enzymes. Annu Rev Biochem. 78:363–397. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakagawa T, Kajitani T, Togo S, Masuko N,
Ohdan H, Hishikawa Y, Koji T, Matsuyama T, Ikura T, Muramatsu M and
Ito T: Deubiquitylation of histone H2A activates transcriptional
initiation via trans-histone cross-talk with H3K4 di- and
trimethylation. Genes Dev. 22:37–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Chen C, Hou X, Gao Y, Lin F, Yang
J, Gao Z, Pan L, Tao L, Wen C, et al: Identification of the E3
deubiquitinase ubiquitin-specific peptidase 21 (USP21) as a
positive regulator of the transcription factor GATA3. J Biol Chem.
288:9373–9382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sippl W, Collura V and Colland F:
Ubiquitin-specific proteases as cancer drug targets. Future Oncol.
7:619–632. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saxena M and Yeretssian G: NOD-like
receptors: Master regulators of inflammation and cancer. Front
Immunol. 5:3272014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yimam M, Lee YC, Moore B, Jiao P, Hong M,
Nam JB, Kim MR, Hyun EJ, Chu M, Brownell L and Jia Q: Analgesic and
anti-inflammatory effects of UP1304, a botanical composite
containing standardized extracts of Curcuma longa and Morus alba. J
Integr Med. 14:60–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wertz IE, O'Rourke KM, Zhou H, Eby M,
Aravind L, Seshagiri S, Wu P, Wiesmann C, Baker R, Boone DL, et al:
De-ubiquitination and ubiquitin ligase domains of A20 downregulate
NF-kappaB signalling. Nature. 430:694–699. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tzimas C, Michailidou G, Arsenakis M,
Kieff E, Mosialos G and Hatzivassiliou EG: Human ubiquitin specific
protease 31 is a deubiquitinating enzyme implicated in activation
of nuclear factor-kappaB. Cell Signal. 18:83–92. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fan YH, Yu Y, Mao RF, Tan XJ, Xu GF, Zhang
H, Lu XB, Fu SB and Yang J: USP4 targets TAK1 to downregulate
TNFα-induced NF-κB activation. Cell Death Differ. 18:1547–1560.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun W, Tan X, Shi Y, Xu G, Mao R, Gu X,
Fan Y, Yu Y, Burlingame S, Zhang H, et al: USP11 negatively
regulates TNFalpha-induced NF-kappaB activation by targeting on
IkappaBalpha. Cell Signal. 22:386–394. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu G, Tan X, Wang H, Sun W, Shi Y,
Burlingame S, Gu X, Cao G, Zhang T, Qin J and Yang J:
Ubiquitin-specific peptidase 21 inhibits tumor necrosis factor
alpha-induced nuclear factor kappaB activation via binding to and
deubiquitinating receptor-interacting protein 1. J Biol Chem.
285:969–978. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pei M, Chen D, Li J and Wei L: Histone
deacetylase 4 promotes TGF-beta1-induced synovium-derived stem cell
chondrogenesis but inhibits chondrogenically differentiated stem
cell hypertrophy. Differentiation. 78:260–268. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu XX, Yan YW, Chen D, Ai CZ, Lu X, Xu
SS, Jiang S, Zhong GS, Chen DB and Jiang YZ: Long non-coding RNA
HoxA-AS3 interacts with EZH2 to regulate lineage commitment of
mesenchymal stem cells. Oncotarget. 2016.(Epub ahead of print).
doi: 10.18632/oncotarget.11538.
|
|
24
|
Chen Y, Wang M, Chen D, Wang J and Kang N:
Chromatin remodeling enzyme CHD7 is necessary for osteogenesis of
human mesenchymal stem cells. Biochem Biophys Res Commun.
478:1588–1593. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen D, Jarrell A, Guo C, Lang R and Atit
R: Dermal β-catenin activity in response to epidermal Wnt ligands
is required for fibroblast proliferation and hair follicle
initiation. Development. 139:1522–1533. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Budnick I, Hamburg-Shields E, Chen D,
Torre E, Jarrell A, Akhtar-Zaidi B, Cordovan O, Spitale RC,
Scacheri P and Atit RP: Defining the identity of mouse embryonic
dermal fibroblasts. Genesis. 54:415–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peng L, Hu Y, Chen D, Jiao S and Sun S:
Ubiquitin specific peptidase 21 regulates interleukin-8 expression,
stem-cell like property of human renal cell carcinoma. Oncotarget.
2016.(Epub ahead of print). doi: 10.18632/oncotarget.9751.
View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Johansson P, Pavey S and Hayward N:
Confirmation of a BRAF mutation-associated gene expression
signature in melanoma. Pigment Cell Res. 20:216–221. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Anders C and Carey LA: Understanding and
treating triple-negative breast cancer. Oncology (Williston Park).
22:1233–1240, 1243. 2008.PubMed/NCBI
|
|
34
|
Hartman ZC, Poage GM, den Hollander P,
Tsimelzon A, Hill J, Panupinthu N, Zhang Y, Mazumdar A, Hilsenbeck
SG, Mills GB and Brown PH: Growth of triple-negative breast cancer
cells relies upon coordinate autocrine expression of the
proinflammatory cytokines IL-6 and IL-8. Cancer Res. 73:3470–3480.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Singel SM, Batten K, Cornelius C, Jia G,
Fasciani G, Barron SL, Wright WE and Shay JW: Receptor-interacting
protein kinase 2 promotes triple-negative breast cancer cell
migration and invasion via activation of nuclear factor-kappaB and
c-Jun N-terminal kinase pathways. Breast Cancer Res. 16:R282014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tao L, Chen C, Song H, Piccioni M, Shi G
and Li B: Deubiquitination and stabilization of IL-33 by USP21. Int
J Clin Exp Pathol. 7:4930–4937. 2014.PubMed/NCBI
|
|
37
|
Fan Y, Mao R, Yu Y, Liu S, Shi Z, Cheng J,
Zhang H, An L, Zhao Y, Xu X, et al: USP21 negatively regulates
antiviral response by acting as a RIG-I deubiquitinase. J Exp Med.
211:313–328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yamamoto M, Taguchi Y, Ito-Kureha T, Semba
K, Yamaguchi N and Inoue J: NF-κB non-cell-autonomously regulates
cancer stem cell populations in the basal-like breast cancer
subtype. Nat Commun. 4:22992013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bhola NE, Balko JM, Dugger TC, Kuba MG,
Sánchez V, Sanders M, Stanford J, Cook RS and Arteaga CL: TGF-β
inhibition enhances chemotherapy action against triple-negative
breast cancer. J Clin Invest. 123:1348–1358. 2013. View Article : Google Scholar : PubMed/NCBI
|