Introduction
Lung cancer is one of the most prevalent types of
cancer among women and men worldwide (1). Non-small cell lung cancer (NSCLC)
accounts for >85% of all cases of lung cancer. NSCLC is
primarily treated by surgery (2),
however, patients diagnosed in the advanced stages of the disease
often may not qualify for surgery. In such cases, immunotherapy may
be a promising therapeutic strategy. Although, systemic immune
suppression is frequently observed in patients with advanced NSCLC
(3). Therefore, researchers are
investigating the tumor microenvironment with the aim of making
lung cancer treatment less invasive and more effective.
A number of studies have demonstrated that
myeloid-origin cells are potent suppressors of tumor immunity,
therefore this cell population may significantly impede the
application of immunotherapy in cancer (4–7).
Researchers have endeavored to determine the best possible
phenotype of myeloid-derived suppressor cells (MDSCs) (8,9).
Identification of characteristic surface markers of this population
is complicated, particularly due to its high plasticity and effect
on the tumor microenvironment (9).
With regards to murine models, the most universal is a combination
of Gr-1 protein isoforms and cluster of differentiation (CD) 11b
(10,11). However, the Gr-1 molecule is not
expressed in human MDSCs (12), and
researchers must therefore search for a different characteristic
surface marker.
Human MDSCs are divided into granulocytic and
monocytic (Mo) populations (13). The
first group of cells has been described as
CD14−CD11b+CD33+CD15+
and the second group as CD14+ human leukocyte antigen-D
related (HLA-DR)−/low (13). The absence or low expression of HLA-DR
is characteristic in Mo-MDSCs, thus the
CD14+HLA-DR−/low phenotype has been used in
studies concerning the identification of Mo-MDSCs in NSCLC
(5,14).
Several immunosuppressive functions of MDSCs have
been suggested, including the inhibition of dendritic cell
differentiation and natural killer cell cytotoxicity, inhibition of
T-cell (CD8+ and CD4+) activation, B-cell
impairment and promotion of regulatory T-cell (Treg) expansion.
Furthermore, MDSCs may be involved in the promotion of metastasis
and angiogenesis (15,16). The presence of MDSCs was confirmed in
several forms of cancer, including prostate cancer, glioblastoma,
hepatocellular carcinoma, melanoma, squamous cell carcinoma of the
head and neck, and NSCLC (5,12,17,18).
However, the phenotype and function of MDSCs in NSCLC requires
further clarification.
The present study evaluated the distribution of
Mo-MDSCs (CD14+HLA-DR−/low) and
monocytes/macrophages (CD14+HLA-DR+) in
peripheral blood, lymph nodes and tumor tissue of patients with
NSCLC. Furthermore, the profiles of cytokines produced by Mo-MDSCs
and monocytes/macrophages were compared in order to investigate
their role in NSCLC. The present study focused on interleukin
(IL)-1β, IL-10, IL-12/23p40, tumor necrosis factor (TNF) and
transforming growth factor-β (TGF-β) as they are frequently present
in numerous solid tumors and serve important roles in tumor
progression. The data obtained during the study identified
statistically significant differences between the expression of
Mo-MDSCs and monocytes/macrophages in several tissues and the
profiles of cytokines secreted by these cell populations.
Materials and methods
Blood, lymph node and primary tumor
samples
Samples were taken from 12 patients with NSCLC, who
underwent surgical procedures in the Department of Thoracic
Surgery, Independent Public Teaching Hospital No. 4, Medical
University of Lublin (Lublin, Poland). Whole blood samples and
fragments of lymph nodes and tumor tissue were collected from
October 2013 to September 2014. The mean age of patients enrolled
in the study was 62.25±8.44 years (range, 48–77 years). Of these 12
patients with histologically confirmed cancer (according to the 7th
edition of the Lung Cancer Tumor-Node-Metastasis classification and
staging system) (19), 4 were
classified as stage IIIA, 5 as stage IIA, 1 as stage IIB and 2 as
stage IA or IB (Table I). The number
of patients studied was small due to the very strict inclusion
criteria, which was as follows: Patients were not allowed to have
taken any medication with an immunomodulatory effect in the last
month, they were not allowed to have a previous medical history of
other autoimmune diseases or cancer, and they must not have
presented with any signs of inflammation in the last month. All
patients underwent thoracotomy performed by the same surgical team.
Blood was drawn immediately prior to surgery, while lymph node and
tumor samples were obtained during the surgery. From the whole
blood samples, peripheral blood mononuclear cells (PBMCs) were
isolated by density gradient centrifugation (700 × g) for 20
min at 20°C using Gradisol L (Aqua-Med, Łódź, Poland). Lymph node
and tumor samples were cut into small pieces and subsequently
homogenized using a gentleMACS™ Dissociator (Miltenyi Biotec GmbH,
Bergisch Gladbach, Germany). Suspensions were subsequently passed
through 70 µm mesh strainers and separated cells were washed twice
with phosphate-buffered saline (PAA Laboratories; GE Healthcare
Life Sciences, Chalfont, UK). The study was approved by the ethics
committee of the Medical University of Lublin, and written informed
consent was obtained from all patients enrolled in the study.
 | Table I.Clinicopathological features of
patients with non-small cell lung cancer. |
Table I.
Clinicopathological features of
patients with non-small cell lung cancer.
| Patient | Age, years | Gender | Smoker | Type of
surgery | Stage | Histopathology |
|---|
| 1 | 77 | Male | Yes | Thoracotomy,
lymphadenectomy | IIIA | Carcinoma
planoepitheliale |
| 2 | 55 | Female | Yes | Thoracotomy,
lobectomy and lymphadenectomy | IIA | Adenocarcinoma |
| 3 | 60 | Male | Yes | Lobectomy and
lymphadenectomy | IIIA | Adenocarcinoma |
| 4 | 64 | Male | Yes | Thoracotomy and
lobectomy | IIA | Adenocarcinoma |
| 5 | 73 | Male | Yes | Thoracotomy and
lobectomy | IIIA | Carcinoma
planoepitheliale |
| 6 | 71 | Male | Yes | Thoracotomy,
lobectomy and lymphadenectomy | IB | Carcinoma
planoepitheliale |
| 7 | 64 | Male | Yes | Thoracotomy,
pulmonectomy and lymphadenectomy | IIB | Carcinoma
planoepitheliale |
| 8 | 48 | Male | Yes | Pulmonectomy | IIIA | Carcinoma
planoepitheliale |
| 9 | 62 | Male | Yes | Lobectomy | IA | Adenocarcinoma |
| 10 | 63 | Male | Yes | Lobectomy | IIA | Carcinoma
macrocellulare |
| 11 | 49 | Male | Yes | Lobectomy | IIA | Carcinoma
macrocellulare |
| 12 | 61 | Female | Yes | Thoracotomy,
lobectomy and | IIA | Adenocarcinoma |
Cell culture
PBMCs, lymph node and tumor cells were cultured in
AIM-V® medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) for 4 h with the addition of ionomycin, phorbol
myristate acetate, lipopolysaccharide and Brefeldin A
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany). Cells were
cultured at 37°C in a humidified atmosphere (95%) with 5%
CO2.
Flow cytometry
The following fluorochrome-labeled monoclonal
antibodies (mAbs), which were purchased from BioLegend, Inc. (San
Diego, CA, USA), were used: Phycoerythrin cyanine 7 anti-human CD14
(200 µg/ml; #325618) and Pacific Blue anti-human HLA-DR (200 µg/ml;
#307633). In addition, the following monoclonal antibodies
conjugated with Alexa Fluor 647 were used: Anti-human IL-1β (50
µg/ml; #508208), anti-human IL-10 (50 µg/ml; #501412), anti-human
IL-12/23p40 (25 µg/ml; #501818), anti-human TNF (25 µg/ml; #502916)
and anti-human latency-associated peptide (TGF-β1) (400 µg/ml;
#341801). Non-specific Fc receptor-mediated antibody binding was
blocked with the use of Human TruStain FcX™ (BioLegend, Inc.).
Subsequently, PBMCs, lymph node and tumor cells were incubated with
anti-CD14 and anti-HLA-DR mAbs in the dark for 20 min at room
temperature. Following surface staining, cells were fixed with
Fixation Buffer (BioLegend, Inc.) and permeabilized with
Intracellular Staining Permeabilization Wash Buffer (BioLegend,
Inc.). Intracellular cytokine staining was then performed using
anti-IL-1β, anti-IL-10, anti-IL-12/23p40, anti-TNF or anti-TGF-β1
mAbs. Data acquisition and analysis were performed on a BD
FACSCanto™ II Flow Cytometer (BD Biosciences, Franklin Lakes, NJ,
USA). Representative examples of cytometric analysis are presented
in Fig. 1. Results were generated
using FACSDiva v8.0.1 software (BD Biosciences).
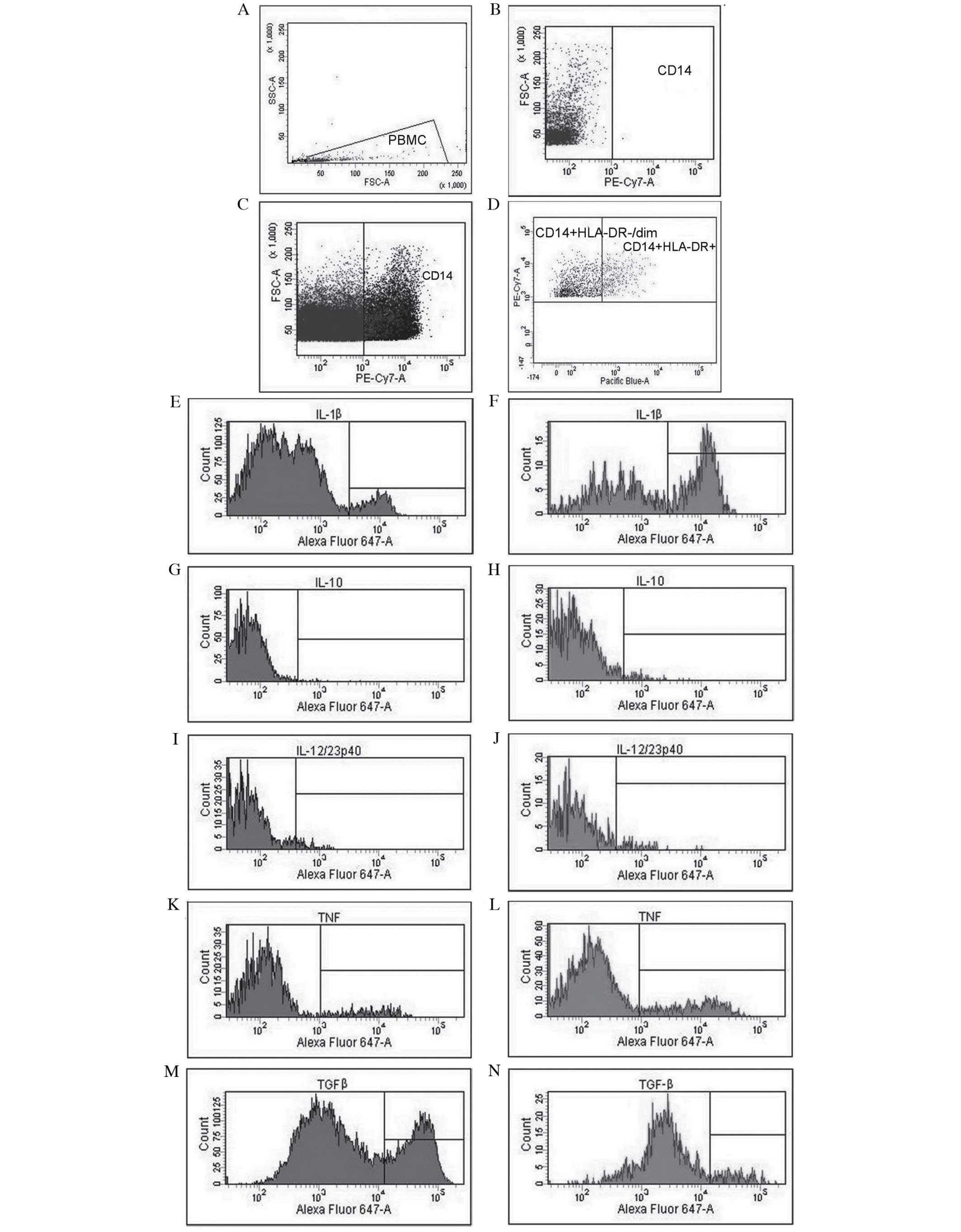 | Figure 1.Analysis of cytokine profiles and
evaluation of the expression of Mo-MDSCs and monocytes/macrophages
in patients with non-small cell lung cancer with the use of flow
cytometry. (A) Gate for PBMCs, (B) CD14 negative control, (C)
identification of CD14+ cells and (D) differentiation of
CD14+HLA−DR−/low cells (Mo-MDSCs)
and CD14+HLA−DR+ cells
(monocytes/macrophages). Evaluation of percentage of Mo-MDSCs
producing (E) IL-1β, (G) IL-10, (I) IL-12/23p40, (K) TNF and (M)
TGF-β. Assessment of percentage of monocytes/macrophages producing
(F) IL-1β, (H) IL-10, (J) IL-12/23p40, (L) TNF and (N) TGF-β.
Mo-MDSCs, monocytic myeloid-derived suppressor cells; PBMC,
peripheral blood mononuclear cells; CD, cluster of differentiation;
IL, interleukin; TNF, tumor necrosis factor; TGF-β, transforming
growth factor-β. |
Statistical analysis
Statistical analyses were performed using STATISTICA
v10.0 software (StatSoft, Inc., Tulsa, OK, USA). The data collected
in the current study did not meet with assumptions of Gaussian
distribution, therefore non-parametric tests were used to analyze
the results. Differences between particular tissues were determined
by the analysis of variance (ANOVA) Friedman test and Kendall's
W-normalization of the statistic of the Friedman test followed by
post-hoc tests. Comparisons between two variables were
prepared with the use of Wilcoxon signed-rank test. Results were
presented as the median and interquartile range, and P<0.05 was
considered to indicate a statistically significant result.
Results
Distribution of Mo-MDSCs and
monocytes/macrophages in the peripheral blood, lymph nodes and
tumor tissues
Using flow cytometry analysis, two cell populations
were detected with the following immunophenotypes:
CD14+HLA-DR−/low and
CD14+HLA-DR+. While
CD14+HLA-DR+ is characteristic for
populations of monocytes/macrophages, in the current study, low
expression of major histocompatibility complex class II among
CD14+ cells was observed on the surface of the Mo-MDSCs
(CD14+HLA-DR−/low). The population of
Mo-MDSCs was significantly higher than the population of
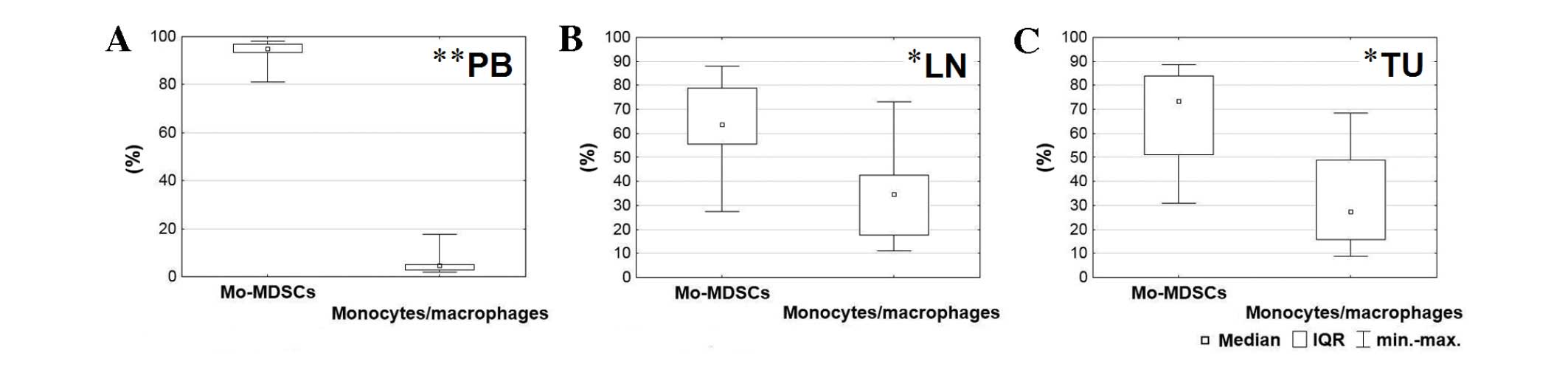
monocytes/macrophages in all tissues (blood, P=0.005; lymph nodes,
P=0.025; tumor, P=0.017; Fig. 2). The
highest percentage of Mo-MDSCs was identified in the peripheral
blood and the lowest was in the lymph nodes. According to
post-hoc tests, which followed the Friedman ANOVA test, the
level of Mo-MDSCs was significantly higher in the blood than the
lymph nodes (P=0.001) and tumor tissue (P=0.001). By contrast, the
highest percentage of monocytes/macrophages was detected in lymph
nodes and the lowest in peripheral blood. The prevalence of
monocytes/macrophages in lymph nodes and tumor tissue in comparison
with peripheral blood was also statistically significant (P=0.001;
Fig. 2).
Comparison of profiles of cytokines
produced by Mo-MDSCs and monocytes/macrophages in the peripheral
blood, lymph nodes and tumor tissues
The present study demonstrated that each population,
CD14+HLA-DR-/low and CD14+HLA-DR+, produced IL-1β, IL-10,
IL-12/23p40, TNF and TGF-β in all examined tissues of patients with
NSCLC. However, significant differences between particular tissues
were identified in the proportion of Mo-MDSCs and
monocytes/macrophages producing these cytokines (Table II).
| Table II.Percentage of cells expressing
cytokines in the peripheral blood, lymph nodes and tumor
tissue. |
Table II.
Percentage of cells expressing
cytokines in the peripheral blood, lymph nodes and tumor
tissue.
|
| Peripheral
blood | Lymph nodes | Tumor tissue |
|---|
|
|
|
|
|
|---|
| Cytokines, % | Mo-MDSCs, median
(IQR) |
Monocytes/macrophages, median (IQR) | Mo-MDSCs, median
(IQR) |
Monocytes/macrophages, median (IQR) | Mo-MDSCs, median
(IQR) |
Monocytes/macrophages, median (IQR) |
|---|
| IL-1β | 5.30 (18.93) | 36.75
(37.00)d | 0.90 (0.85) | 1.70 (1.45) | 1.10 (2.05) | 5.50
(5.50)d |
| IL-10 | 0.20 (0.50) | 0.15 (1.28) | 0.65 (1.00) | 1.90 (2.65) | 0.20 (0.45) | 0.20 (0.35) |
| IL-12/23p40 | 0.00 (0.50) | 0.00 (0.70) | 1.00 (1.00) | 1.30 (2.20) | 1.00 (0.75) | 1.50 (2.50) |
| TNF | 12.50 (17.80) | 34.00 (28.00) | 18.30 (31.53) | 35.75 (32.55) | 2.90 (11.85) | 6.40 (13.25) |
| TGF-β | 38.55
(28.48)a,b | 19.70
(0.70)c | 2.15
(3.95)a | 0.70
(0.90)c | 5.80
(23.40)b | 3.80 (9.50) |
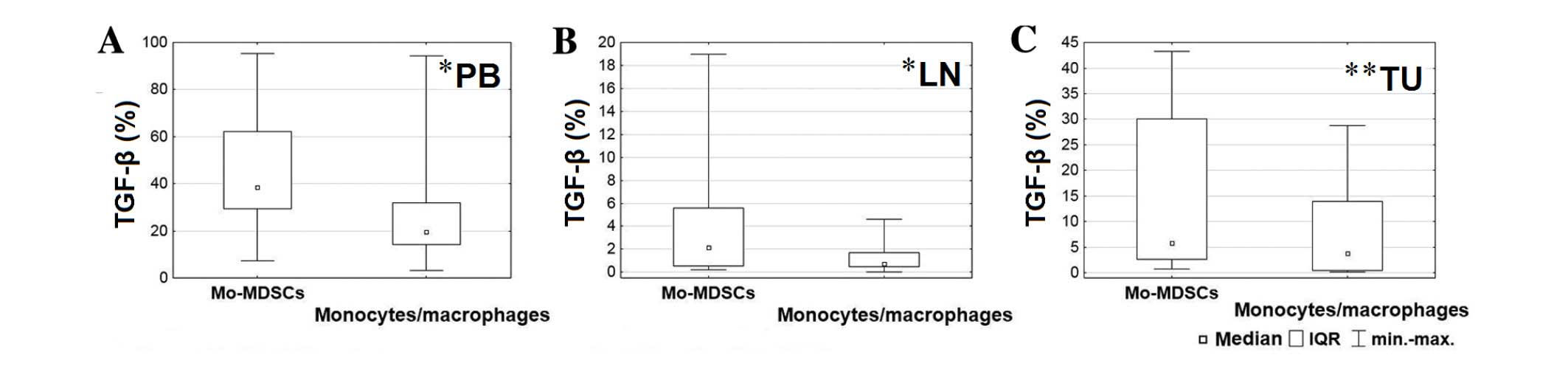
The percentage of Mo-MDSCs producing
TGF-β was significantly higher in the blood than in other tissues
(P=0.001)
Peripheral blood was characterized by a higher
proportion of monocytes/macrophages producing TGF-β (P=0.002) than
lymph nodes, and a higher proportion of monocytes/macrophages
producing IL-1β than tumor tissue (P=0.002) (Table II). The post-hoc test did not
identify a significant difference between blood and lymph nodes in
the proportion of monocytes/macrophages producing TNF. However, the
mean fluorescence intensity (MFI) value for this population
producing TNF was significantly higher in blood (P=0.008; data not
shown). No statistically significant differences were observed
among the level of Mo-MDSCs and monocytes/macrophages or in the
expression of cytokines in the post-hoc tests between lymph
nodes and tumor tissue.
Analysis of differences between the
levels of cytokines produced by the Mo-MDSCs and
monocytes/macrophages in the peripheral blood, lymph nodes and
tumor tissues
In order to analyze the differences between levels
of cytokines produced by the Mo-MDSCs and monocytes/macrophages in
several tissues, the Wilcoxon signed-rank test was used. A
significantly lower proportion of monocytes/macrophages produced
TGF-β in the blood (P=0.028), lymph nodes (P=0.015) and tumor
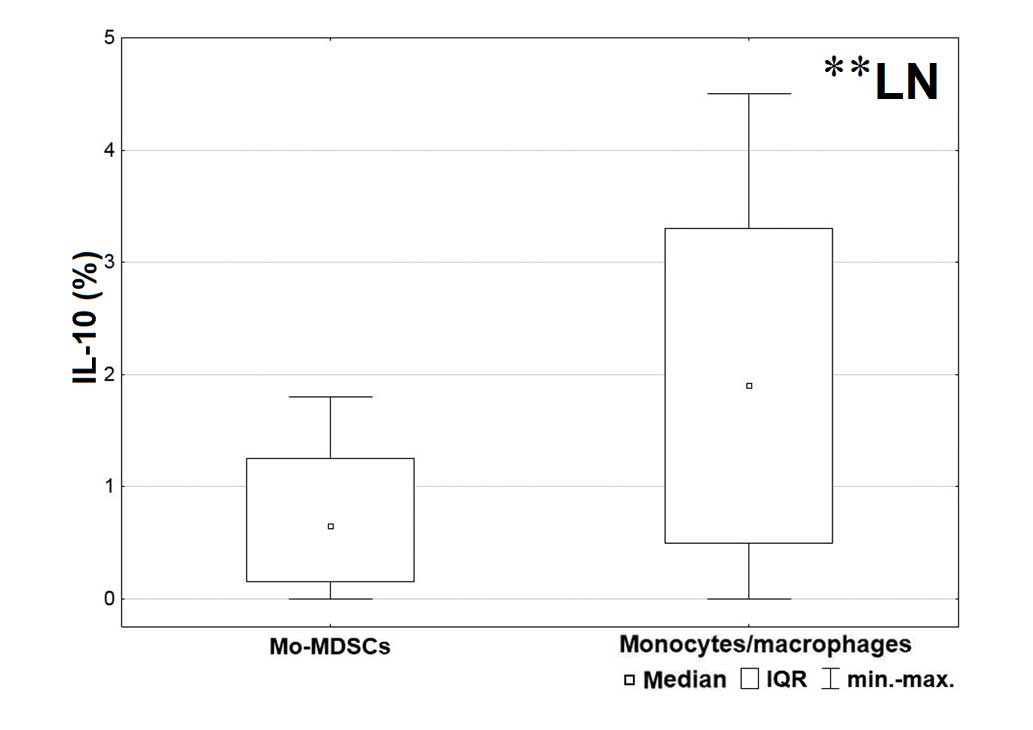
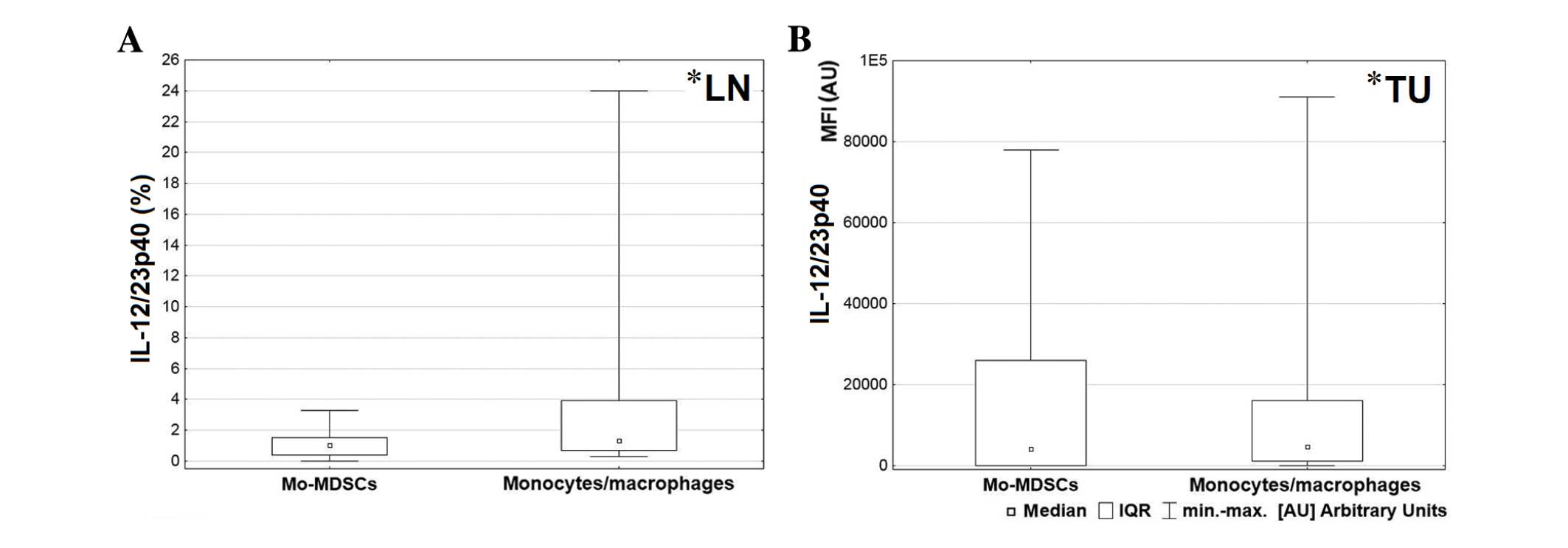
tissue (P=0.006) compared with the Mo-MDSCs (Fig. 3). Furthermore, in the lymph nodes, a
significantly higher proportion of IL-10 was produced by
monocytes/macrophages than the Mo-MDSCs (P=0.003; Fig. 4).
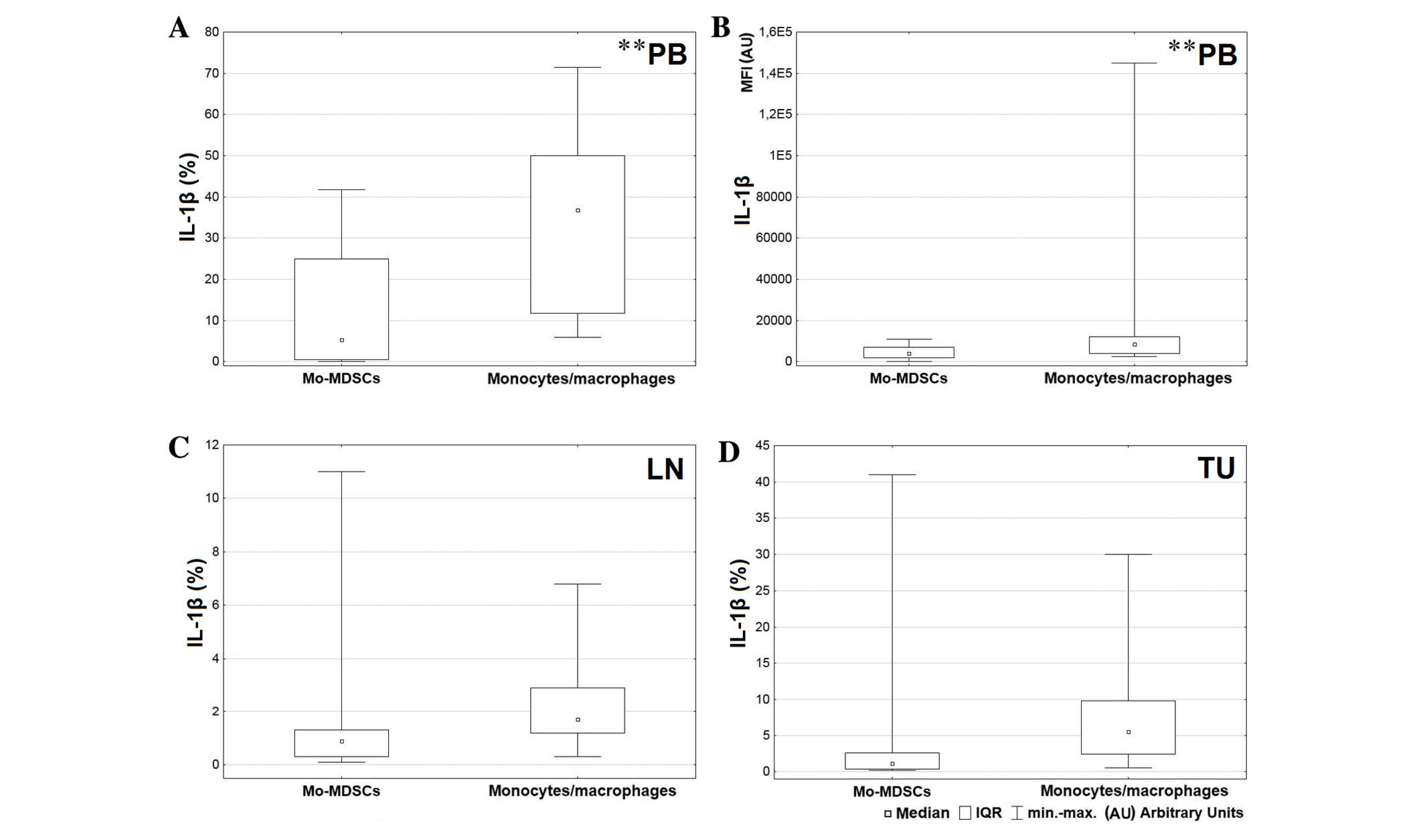
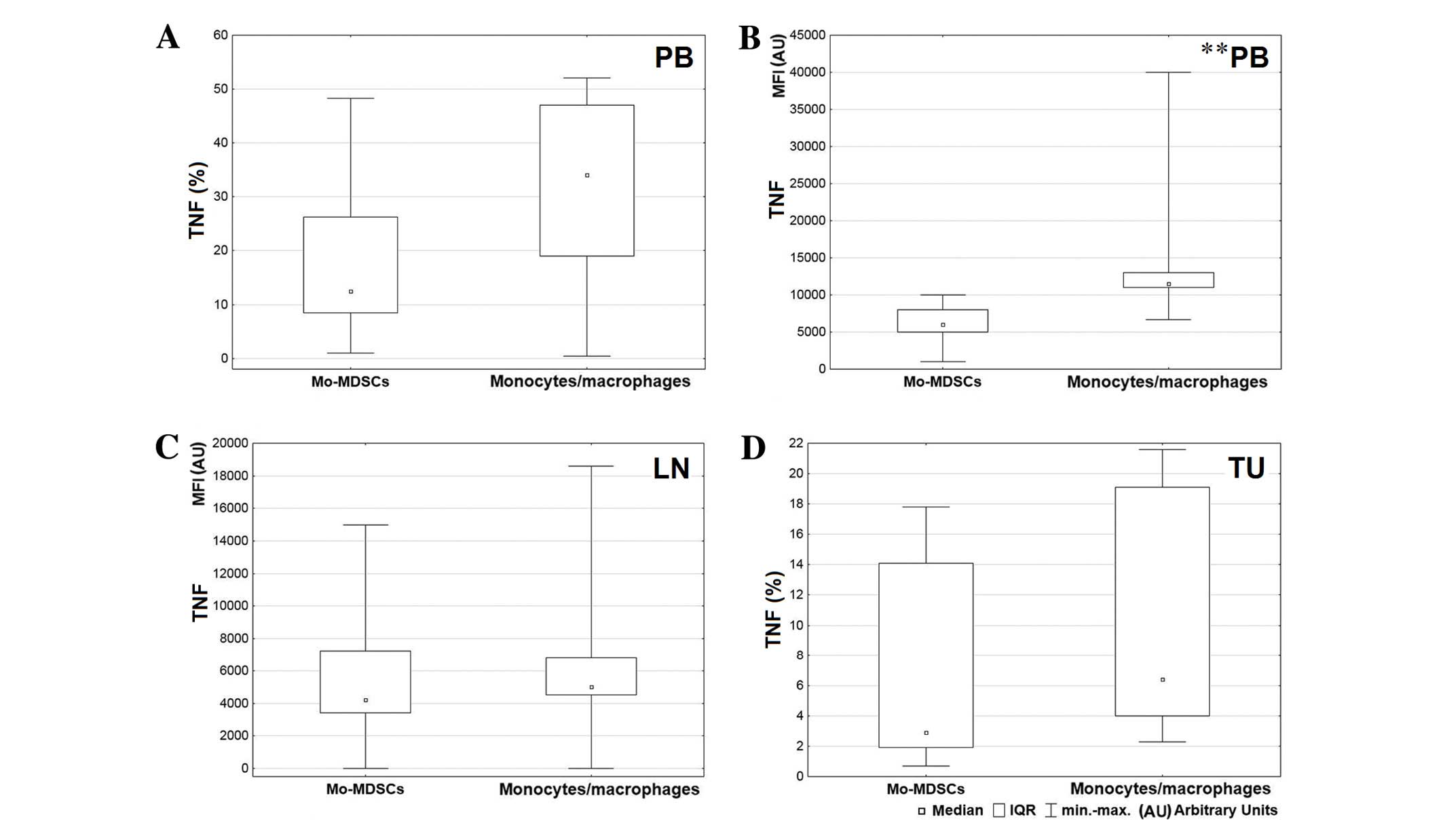
A greater level of IL-1β was produced by the
CD14+HLA-DR+ population in comparison with
Mo-MDSCs in all tissues, but the only significant result was
observed in the blood (P=0.004; Fig.
5A). Also the MFI value for monocytes/macrophages producing
IL-1β in peripheral blood was significantly higher than the MFI
value for Mo-MDSCs producing IL-1β (P=0.005; Fig. 5B). Differences in expression of IL-1β
in the tumor and lymph nodes were not significant (lymph nodes
P=0.074; tumor P=0.062; Fig. 5C and
D, respectively). Statistical analysis demonstrated a higher
percentage of monocytes/macrophages producing TNF in peripheral
blood and tumor tissue (Fig. 6A and
D, respectively) and a higher MFI value for the mentioned
population in the blood and lymph nodes (Fig. 6B and C, respectively). However, a
significant difference was only observed between MFI values
measured for monocytes/macrophages and Mo-MDSCs producing TNF in
the blood (P=0.008; Fig. 6B). Data
analysis indicated that in lymph nodes and tumor tissue,
monocytes/macrophages produced a higher proportion of IL-12/23p40
than Mo-MDSCs (P=0.047 and P=0.041, respectively; Fig. 7).
Discussion
Lung cancer is the most prevalent cause of
cancer-associated mortality in the world (20). The most common subtype is NSCLC, which
is characterized by poor prognosis and high mortality, particularly
in cases that are unsuitable for surgery (1). A number of treatment options are
currently available to treat patients with cancer, however, therapy
is often ineffective. A growing body of evidence suggests that
cancer initiation and progression depends on the ability of cancer
cells to evade host immunosurveillance (21,22).
Cancer cells regulate and modify immune responses and therefore
contribute to an imbalance in type 1 T-helper (Th)/Th2 responses
(21) and the enhancement of
immunosuppressive cells, including Treg cells (23), tumor associated macrophages (24), type 2 natural killer T-cells (25) and myeloid-derived suppressor cells
(26). Therapeutic strategies
targeting these cell populations are emerging (27–31) and
may benefit patients in the future. However, prior to the
therapeutic exploitation of these cell populations, it is important
to evaluate their role and interactions in the tumor
microenvironment. The current study therefore aimed to investigate
the distribution of Mo-MDSCs and monocytes/macrophages in NSCLC
tissues, and additionally assess and compare the profiles of
cytokines produced by these cell populations. To the best of our
knowledge, this is the first study to examine the distribution of
Mo-MDSCs and monocytes/macrophages simultaneously in the peripheral
blood, tumor tissues and lymph nodes of patients with NSCLC.
In the present study, Mo-MDSCs and
monocytes/macrophages were detected in all examined tissues.
Mo-MDSCs were more abundant than monocytes/macrophages throughout
all tissues, particularly in peripheral blood. Vetsika et al
(15) previously identified an
inverse correlation between CD14+HLA-DR−/low
and CD14+HLA-DR+ cells in the peripheral
blood of patients with NSCLC. This suggests that
CD14+HLA-DR+ cells are effector cells, which
are responsible for activating T lymphocytes (15). However, Feng et al (32) demonstrated that in patients with
NSCLC, the mixture of CD14+HLA-DR−/low cells
and CD14+HLA-DR+ cells resulted in a more
powerful suppressive activity on T-cells than the
CD14+HLA-DR−/low population alone (32). The immunosuppressive activity of
Mo-MDSCs and CD14+HLA-DR+ cells was confirmed
in multiple myeloma (33). By
contrast, Schilling et al (14) demonstrated that
CD14+HLA-DR+ cells did not exhibit
suppressive properties in melanoma.
In the present study, it was observed that
monocytes/macrophages were significantly more prevalent in the
lymph nodes and tumor tissues than in blood. This result supports
the statement that Mo-MDSCs may differentiate into
monocytes/macrophages in the tumor microenvironment (34). This process may be regulated by
hypoxia inducible factor 1α, which is induced in the tumor
microenvironment by hypoxia (34,35).
TGF-β may stimulate tumor progression in advanced
stages of tumorigenesis, and is described as an immunosuppression
and angiogenesis promoter in the tumor microenvironment (36). Beury et al (37) and De Keersmaecker et al
(33) reported that TGF-β is produced
by Mo-MDSCs and monocytes/macrophages in the peripheral blood.
However, it may be necessary to closely analyze the cross-talk and
differences between MDSCs and monocytes/macrophages, and
investigate the distribution of TGF-β in the tissues of patients
with cancer. At the protein level, FACS data obtained during the
present study indicated a significantly higher number of Mo-MDSCs
producing TGF-β in all tissues. However, a previous study by Huang
et al (5) did not observe any
differences in the transcription of this cytokine between Mo-MDSCs
and CD14+HLA-DR+ cells in the peripheral
blood of patients with NSCLC.
The presence of IL-10 has been confirmed in various
types of cancer, including B-cell non-Hodgkin lymphoma (NHL),
prostate cancer, melanoma, squamous cell carcinoma of the head and
neck, hepatocellular carcinoma, multiple myeloma, glioblastoma and
bladder carcinoma (5,38–40).
According to Feng et al (32),
IL-10 is responsible for upregulating arginase 1 and therefore
inhibits T-cell activation in patients with NSCLC. Furthermore, Xiu
et al (40) demonstrated that
IL-10 is responsible for the development of Mo-MDSCs in B-cell NHL.
Elevated production of IL-10 by macrophages associated with the
tumor microenvironment correlates with poor prognosis, stage of
disease, tumor size and lymph node metastasis (38,41). This
correlation was confirmed in patients with NSCLC (42,43).
However, there was no clear division of cells into
CD14+HLA-DR−/low and HLA-DR+
populations in the aforementioned studies. The results of the
current study demonstrated that Mo-MDSCs and monocytes/macrophages
produced IL-10 in all examined tissues of patients with NSCLC.
Furthermore, a significantly higher proportion of
monocytes/macrophages producing IL-10 than Mo-MDSCs were observed
in lymph nodes; this is important as lymph nodes are sites of
lymphocyte activation and immune response induction.
According to the results of the present study,
monocytes/macrophages have the ability to produce higher levels of
IL-1β, TNF and IL-12/23p40 than Mo-MDSCs. A significantly higher
prevalence of CD14+HLA-DR+ producing IL-1β
and TNF was observed in peripheral blood and a significantly higher
prevalence of CD14+HLA-DR+ cells producing
IL-12/23p40 was observed in lymph nodes and tumor tissue. These
observations correspond to the cytokine functions: IL-1β and TNF
are responsible for generating MDSCs in the bone marrow (44), and both cytokines have a proangiogenic
role in patients with NSCLC (45).
The p40 subunit is common for IL-12 and IL-23, therefore, both
cytokines had to be considered in the analysis. IL-12 promotes Th 1
immunity and inhibits angiogenesis in the tumor microenvironment,
while IL-23 exhibits tumor-promoting effects (46). According to the results of research on
cross-talk between MDSCs and macrophages in solid tumors, each
population produces IL-12 (37,47).
However, in the current in vitro study, IL-23p40 was not
produced. By contrast, Baird et al (48) confirmed the presence of IL-23 in the
tumor microenvironment and demonstrated that IL-23 induced
proliferation in an NSCLC cell line and promoted proliferation in
primary NSCLC tumors (48). In the
present study, a higher prevalence of IL-12/23p40 positive cells in
lymph nodes and in tumor tissue was observed. This may be
associated with a more aggressive tumor, when the results of Baird
et al (48) are
considered.
In conclusion, the present study demonstrated that
Mo-MDSCs were more abundant than monocytes/macrophages in all NSCLC
tissue examined and the highest percentage of Mo-MDSCs was detected
in the blood. In all tissues, Mo-MDSCs produced higher levels of
TGF-β than CD14+HLA-DR+ cells. Furthermore,
the proportion of Mo-MDSCs producing TGF-β was higher in the blood
compared with the lymph nodes and tumor tissues. These observations
suggest that Mo-MDSCs and monocytes/macrophages participate in
NSCLC induced immunosuppression. In addition, higher levels of
Mo-MDSCs and TGF-β identified in the blood may correspond with
results from previous studies regarding associations between the
TGF-β signaling pathway and tumor cell invasion, motility and
metastasis, and also with the notion that Mo-MDSCs promote tumor
growth through their immunosuppressive activity. Lymph nodes and
tumor tissue were characterized by a higher prevalence of
monocytes/macrophages than peripheral blood. This population
produced significantly more immunosuppressive IL-10 in lymph nodes
than Mo-MDSCs. CD14+HLA-DR+ cells secreted
more IL-1β and TNF than Mo-MDSCs in all tissues. Furthermore, a
higher proportion of monocytes/macrophages producing cytokines was
observed in the peripheral blood. This corresponds with the fact
that IL-1β and TNF produced by monocytes/macrophages are associated
with metastasis formation and angiogenesis promotion. A higher
percentage of IL-12/23p40 produced by
CD14+HLA-DR+ cells was detected in lymph
nodes and in tumor tissue. This molecule may represent IL-23, which
promotes proliferation in primary NSCLC tumors. Analyzing the
distribution of Mo-MDSCs and monocytes/macrophages in NSCLC tissues
and profiling the cytokines they secrete may allow researchers to
investigate the role of these populations in the tumor
microenvironment and subsequently enable modifications to their
functions. In the future, modulating the activities of MDSCs may be
a useful additional tool in anticancer therapy.
Acknowledgements
The present study was performed within the project
‘Centre for Innovative Research in Medical and Natural Sciences’
initiated by the University of Rzeszów (Rzeszów, Poland), and was
co-financed by the Regional Operational Programme for the
Podkarpackie Province between 2007 and 2013 (contract number,
UDA-RPPK.01.03.00–18-004/12-00).
Glossary
Abbreviations
Abbreviations:
|
IL-1β
|
interleukin-1β
|
|
IL-10
|
interleukin-10
|
|
mAbs
|
monoclonal antibodies
|
|
MDSCs
|
myeloid-derived suppressor cells
|
|
Mo-MDSCs
|
monocytic myeloid-derived suppressor
cells
|
|
Treg
|
regulatory T-cell
|
|
NHL
|
non-Hodgkin lymphoma
|
|
NSCLC
|
non-small cell lung cancer
|
|
PBMCs
|
peripheral blood mononuclear cells
|
|
TGF-β
|
transforming growth factor-β
|
|
TNF
|
tumor necrosis factor
|
References
|
1
|
Nur U, Quaresma M, De Stavola B, Peake M
and Rachet B: Inequalities in non-small cell lung cancer treatment
and mortality. J Epidemiol Community Health. 69:985–992. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thomas A, Chen Y, Yu T, Jakopovic M and
Giaccone G: Trends and characteristics of young non-small cell lung
cancer patients in the United States. Front Oncol. 5:1132015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aerts JG, Lievense LA, Hoogsteden HC and
Hegmans JP: Immunotherapy prospects in the treatment of lung cancer
and mesothelioma. Transl Lung Cancer Res. 3:34–45. 2014.PubMed/NCBI
|
|
4
|
Ohki S, Shibata M, Gonda K, Machida T,
Shimura T, Nakamura I, Ohtake T, Koyama Y, Suzuki S, Ohto H and
Takenoshita S: Circulating myeloid-derived suppressor cells are
increased and correlate to immune suppression, inflammation and
hypoproteinemia in patients with cancer. Oncol Rep. 28:453–458.
2012.PubMed/NCBI
|
|
5
|
Huang A, Zhang B, Wang B, Zhang F, Fan KX
and Guo YJ: Increased CD14(+)HLA-DR (−/low) myeloid-derived
suppressor cells correlate with extrathoracic metastasis and poor
response to chemotherapy in non-small cell lung cancer patients.
Cancer Immunol Immunother. 62:1439–1451. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ostrand-Rosenberg S: Myeloid-derived
suppressor cells: More mechanisms for inhibiting antitumor
immunity. Cancer Immunol Immunother. 59:1593–1600. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Parker KH, Beury DW and Ostrand-Rosenberg
S: Myeloid-Derived suppressor cells: Critical cells driving immune
suppression in the tumor microenvironment. Adv Cancer Res.
128:95–139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marigo I, Dolcetti L, Serafini P,
Zanovello P and Bronte V: Tumor-induced tolerance and immune
suppression by myeloid derived suppressor cells. Immunol Rev.
222:162–179. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Damuzzo V, Pinton L, Desantis G, Solito S,
Marigo I, Bronte V and Mandruzzato S: Complexity and challenges in
defining myeloid-derived suppressor cells. Cytometry B Clin Cytom.
88:77–91. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ortiz ML, Lu L, Ramachandran I and
Gabrilovich DI: Myeloid-derived suppressor cells in the development
of lung cancer. Cancer Immunol Res. 2:50–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dolcetti L, Peranzoni E, Ugel S, Marigo I,
Gomez A Fernandez, Mesa C, Geilich M, Winkels G, Traggiai E, Casati
A, et al: Hierarchy of immunosuppressive strength among
myeloid-derived suppressor cell subsets is determined by GM-CSF.
Eur J Immunol. 40:22–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fujimura T, Mahnke K and Enk AH: Myeloid
derived suppressor cells and their role in tolerance induction in
cancer. J Dermatol Sci. 59:1–6. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qu P, Wang LZ and Lin PC: Expansion and
functions of myeloid-derived suppressor cells in the tumor
microenvironment. Cancer Lett. 380:253–256. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schilling B, Sucker A, Griewank K, Zhao F,
Weide B, Görgens A, Giebel B, Schadendorf D and Paschen A:
Vemurafenib reverses immunosuppression by myeloid derived
suppressor cells. Int J Cancer. 133:1653–1663. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vetsika EK, Koinis F, Gioulbasani M,
Aggouraki D, Koutoulaki A, Skalidaki E, Mavroudis D, Georgoulias V
and Kotsakis A: A circulating subpopulation of monocytic
myeloid-derived suppressor cells as an independent
prognostic/predictive factor in untreated non-small lung cancer
patients. J Immunol Res. 2014:6592942014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schmid MC and Varner JA: Myeloid cells in
the tumor microenvironment: Modulation of tumor angiogenesis and
tumor inflammation. J Oncol. 2010:2010262010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Solito S, Marigo I, Pinton L, Damuzzo V,
Mandruzzato S and Bronte V: Myeloid-derived suppressor cell
heterogeneity in human cancers. Ann N Y Acad Sci. 1319:47–65. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mandruzzato S, Solito S, Falisi E,
Francescato S, Chiarion-Sileni V, Mocellin S, Zanon A, Rossi CR,
Nitti D, Bronte V and Zanovello P: IL4Ralpha+ myeloid-derived
suppressor cell expansion in cancer patients. J Immunol.
182:6562–6568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mirsadraee S, Oswal D, Alizadeh Y, Caulo A
and van Beek E Jr: The 7th lung cancer TNM classification and
staging system: Review of the changes and implications. World J
Radiol. 4:128–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ridge CA, McErlean AM and Ginsberg MS:
Epidemiology of lung cancer. Semin Intervent Radiol. 30:93–98.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vinay DS, Ryan EP, Pawelec G, Talib WH,
Stagg J, Elkord E, Lichtor T, Decker WK, Whelan RL, Kumara HM, et
al: Immune evasion in cancer: Mechanistic basis and therapeutic
strategies. Semin Cancer Biol. 35:(Suppl). S185–S198. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Parcesepe P, Giordano G, Laudanna C,
Febbraro A and Pancione M: Cancer-associated immune resistance and
evasion of immune surveillance in colorectal cancer. Gastroenterol
Res Pract. 2016:62617212016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Curiel TJ: Regulatory T cells and
treatment of cancer. Curr Opin Immunol. 20:241–246. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stewart TJ and Abrams SI: How tumours
escape mass destruction. Oncogene. 27:5894–5903. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Berzofsky JA and Terabe M: NKT cells in
tumor immunity: Opposing subsets define a new immunoregulatory
axis. J Immunol. 180:3627–3635. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marvel D and Gabrilovich DI:
Myeloid-derived suppressor cells in the tumor microenvironment:
Expect the unexpected. J Clin Invest. 125:3356–3364. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Adams JL, Smothers J, Srinivasan R and
Hoos A: Big opportunities for small molecules in immuno-oncology.
Nat Rev Drug Discov. 14:603–622. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mahoney KM, Rennert PD and Freeman GJ:
Combination cancer immunotherapy and new immunomodulatory targets.
Nat Rev Drug Discov. 14:561–584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sinha P and Ostrand-Rosenberg S:
Myeloid-derived suppressor cell function is reduced by Withaferin
A, a potent and abundant component of Withania somnifera root
extract. Cancer Immunol Immunother. 62:1663–1673. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vincent J, Mignot G, Chalmin F, Ladoire S,
Bruchard M, Chevriaux A, Martin F, Apetoh L, Rébé C and
Ghiringhelli F: 5-Fluorouracil selectively kills tumor-associated
myeloid-derived suppressor cells resulting in enhanced T
cell-dependent antitumor immunity. Cancer Res. 70:3052–3061. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang WC, Ma G, Chen SH and Pan PY:
Polarization and reprogramming of myeloid-derived suppressor cells.
J Mol Cell Biol. 5:207–209. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Feng PH, Lee KY, Chang YL, Chan YF, Kuo
LW, Lin TY, Chung FT, Kuo CS, Yu CT, Lin SM, et al:
CD14(+)S100A9(+) monocytic myeloid-derived suppressor cells and
their clinical relevance in non-small cell lung cancer. Am J Respir
Crit Care Med. 186:1025–1036. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
De Keersmaecker B, Fostier K, Corthals J,
Wilgenhof S, Heirman C, Aerts JL, Thielemans K and Schots R:
Immunomodulatory drugs improve the immune environment for dendritic
cell-based immunotherapy in multiple myeloma patients after
autologous stem cell transplantation. Cancer Immunol Immunother.
63:1023–1036. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Corzo CA, Condamine T, Lu L, Cotter MJ,
Youn JI, Cheng P, Cho HI, Celis E, Quiceno DG, Padhya T, et al:
HIF-1α regulates function and differentiation of myeloid-derived
suppressor cells in the tumor microenvironment. J Exp Med.
207:2439–2453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kumar V and Gabrilovich DI:
Hypoxia-inducible factors in regulation of immune responses in
tumour microenvironment. Immunology. 143:512–519. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Krstic J and Santibanez JF: Transforming
growth factor-beta and matrix metalloproteinases: Functional
interactions in tumor stroma-infiltrating myeloid cells. Scientific
World Journal. 2014:5217542014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Beury DW, Parker KH, Nyandjo M, Sinha P,
Carter KA and Ostrand-Rosenberg S: Cross-talk among myeloid-derived
suppressor cells, macrophages, and tumor cells impacts the
inflammatory milieu of solid tumors. J Leukoc Biol. 96:1109–1118.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
O'Garra A, Barrat FJ, Castro AG, Vicari A
and Hawrylowicz C: Strategies for use of IL-10 or its antagonists
in human disease. Immunol Rev. 223:114–131. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eruslanov E, Neuberger M, Daurkin I,
Perrin GQ, Algood C, Dahm P, Rosser C, Vieweg J, Gilbert SM and
Kusmartsev S: Circulating and tumor-infiltrating myeloid cell
subsets in patients with bladder cancer. Int J Cancer.
130:1109–1119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiu B, Lin Y, Grote DM, Ziesmer SC,
Gustafson MP, Maas ML, Zhang Z, Dietz AB, Porrata LF, Novak AJ, et
al: IL-10 induces the development of immunosuppressive
CD14(+)HLA-DR (low/-) monocytes in B-cell non-Hodgkin lymphoma.
Blood Cancer J. 5:e3282015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang R, Lu M, Zhang J, Chen S, Luo X, Qin
Y and Chen H: Increased IL-10 mRNA expression in tumor-associated
macrophage correlated with late stage of lung cancer. J Exp Clin
Cancer Res. 30:622011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lievense LA, Bezemer K, Aerts JG and
Hegmans JP: Tumor-associated macrophages in thoracic malignancies.
Lung Cancer. 80:256–262. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zeni E, Mazzetti L, Miotto D, Lo Cascio N,
Maestrelli P, Querzoli P, Pedriali M, De Rosa E, Fabbri LM, Mapp CE
and Boschetto P: Macrophage expression of interleukin-10 is a
prognostic factor in nonsmall cell lung cancer. Eur Respir J.
30:627–632. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wesolowski R, Markowitz J and Carson WE
III: Myeloid derived suppressor cells-a new therapeutic target in
the treatment of cancer. J Immunother Cancer. 1:102013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Quatromoni JG and Eruslanov E:
Tumor-associated macrophages: Function, phenotype, and link to
prognosis in human lung cancer. Am J Transl Res. 4:376–389.
2012.PubMed/NCBI
|
|
46
|
Ngiow SF, Teng MW and Smyth MJ: A balance
of interleukin-12 and −23 in cancer. Trends Immunol. 34:548–555.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ostrand-Rosenberg S, Sinha P, Beury DW and
Clements VK: Cross-talk between myeloid-derived suppressor cells
(MDSC), macrophages and dendritic cells enhances tumor-induced
immune suppression. Semin Cancer Biol. 22:275–281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Baird AM, Leonard J, Naicker KM, Kilmartin
L, O'Byrne KJ and Gray SG: IL-23 is pro-proliferative,
epigenetically regulated and modulated by chemotherapy in non-small
cell lung cancer. Lung Cancer. 79:83–90. 2013. View Article : Google Scholar : PubMed/NCBI
|