Introduction
Tumors and metastases usually arise as small
avascular masses that subsequently induce neovascularization in
order to acquire nutrients for further growth and metastatic spread
(1–3).
This angiogenic switch is induced by several factors such as
vascular endothelial growth factor (VEGF), which is secreted by
tumor cells (4). VEGF was originally
identified and isolated as an endothelial cell-specific mitogen
that is able to induce physiological and pathological angiogenesis
(5,6).
Alternative splicing of VEGF leads to several different isoforms,
which are differentially expressed in various tumor types and have
distinct functions in tumor blood vessel formation (7). The angiogenic isoforms of VEGF are known
as VEGF121, VEGF145, VEGF165 and VEGF189 in humans (8). VEGF165 is one of the most abundant and
potent angiogenic agents among all the VEGF isoforms (9). Cellular responses to VEGF165 are
mediated by two high-affinity type III tyrosine kinase receptors,
VEGF receptor (VEGF-R)2 (also known as kinase insert domain
receptor and fetal liver kinase 1) and VEGF-R1 (also known as
Fms-related tyrosine kinase 1), and two receptors of the semaphorin
receptor family, neuropilin-1 and neuropilin-2 (10). In addition to its functions in
endothelial cells, the role of VEGF in tumor cells is currently an
emerging area of importance; therefore, an increasing number of
studies have focused on the biology of VEGF in tumor cells
(11–13).
Vector-mediated gene expression is one of the most
important tools for studying gene functions in vitro and
in vivo (14). To date,
several viral vectors have been applied in laboratory experiments
or clinical trials and have achieved good results (15–17). In
the majority of cases, the efficiency of gene transfer represents
the most relevant obstacle, since it limits the success of gene
overexpression or gene silencing (18). Compared with non-viral methods,
viruses are highly-evolved, natural delivery agents for genetic
materials (16,19). In addition to their remarkable
transduction efficiency, previous laboratory experiments have
suggested that lentivirus vectors (LVs) exhibit low immunogenicity
and durable expression (14).
Furthermore, one of the most striking characteristics
distinguishing the lentivirus genus from gammaretroviruses is their
ability to infect non-replicating cells (20). These advantages make lentiviruses a
powerful tool in the studies of gene functions.
The present study intended to construct a
recombinant LV containing the VEGF165-enhanced green fluorescent
protein (EGFP) fusion gene, and investigated the feasibility of
using LV to express the fused VEGF165-EGFP gene in the breast
cancer cell line MCF-7. The present study lays the foundation for
future in vitro and in vivo studies on tumor cell
derived-VEGF.
Materials and methods
Cell lines and cell culture
The breast carcinoma cell line MCF-7 and the human
embryonic kidney epithelial cell line 293T were obtained from
Shanghai Cell Bank, Chinese Academy of Sciences (Shanghai, China).
All cells were cultured in Dulbecco's modified Eagle medium (DMEM;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin, and were
maintained at 37°C under 5% CO2. Cells used in the
experiments were in logarithmic growth phase.
Construction and sequencing of
pLVX-VEGF165-EGFP- 3FLAG recombinant plasmid expression vector
The VEGF165 (NM_001171626) coding region was
amplified by polymerase chain reaction (PCR) using the PrimeScript™
RT-PCR kit (Takara Biotechnology Co., Ltd., Dalian, China) and the
following cycling conditions: Pre-denaturation at 98°C for 3 min;
30 cycles at 94°C for 10 sec, 55°C for 15 sec and 72°C for 1 min;
and 72°C for a final 10 min. The VEGF165 fragment DNA was extracted
and purified using the QIAquick Gel Extraction kit and the QIAquick
PCR Purification kit, respectively (Qiagen GmbH, Hilden, Germany).
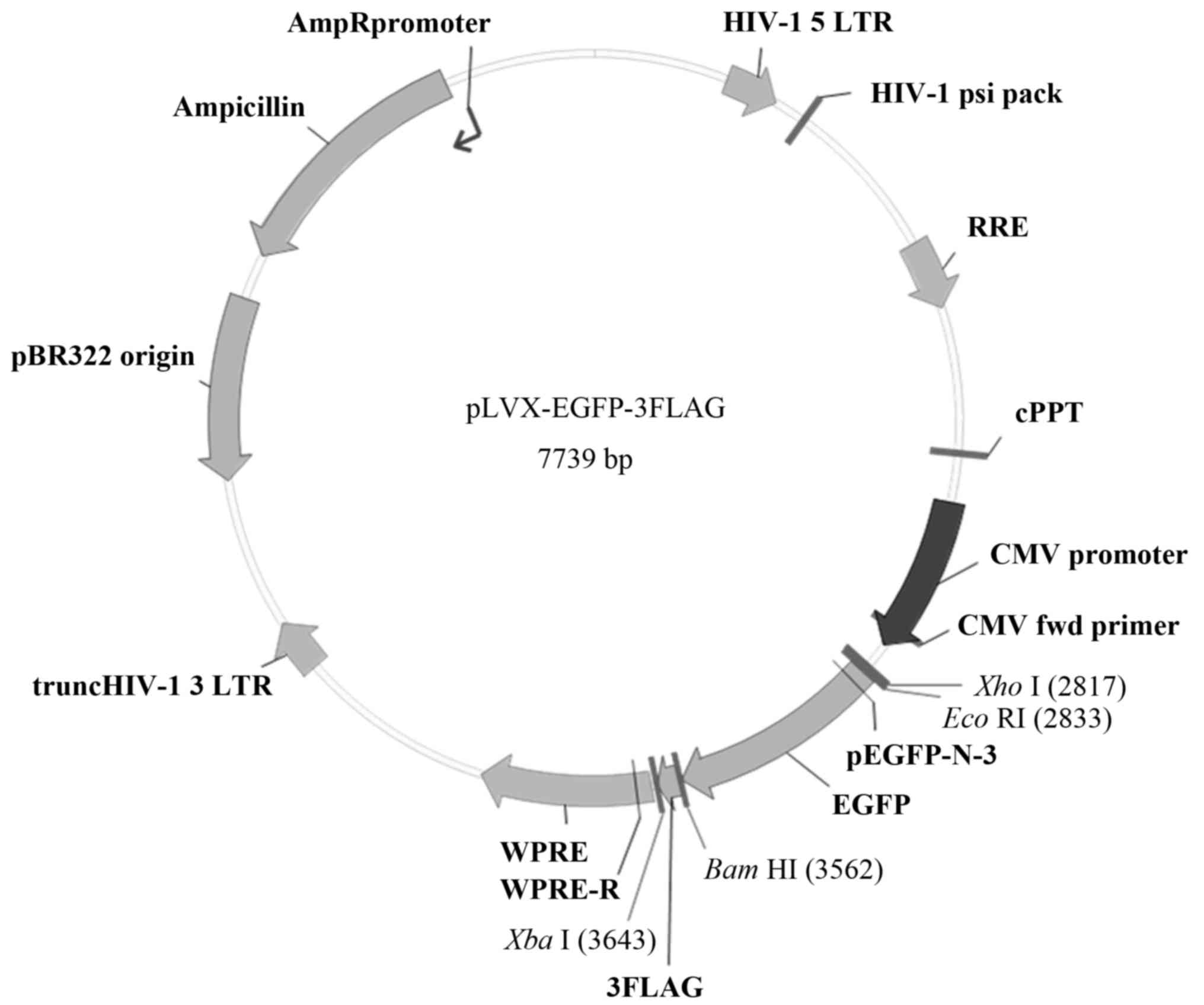
The purified VEGF165 fragment and the pLVX-EGFP-3FLAG plasmid
(Shanghai Sbo-bio Biotechnology, Shanghai, China) (Fig. 1) were digested separately using
EcoRI restriction endonuclease. Upon digestion, the mixture
was incubated at 65°C for 10 min to stop the reaction, after which
the products were separated by 1.5% agarose gel electrophoresis.
The bands were visualized using a gel scanning analysis system
(ChemiDoc MP imaging system; Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Then, the target fragment of about 7.7 kb in size was
recycled by 1% agarose gel electrophoresis.
Using the In-Fusion® HD Cloning kit
(Clontech Laboratories, Inc., Mountainview, CA, USA), the VEGF165
fragment was ligated into the pLVX-EGFP-3FLAG expression vector,
which had been digested with EcoRI. Two control groups were
used, including a blank control and a negative control group. Next,
the recombinant plasmid vector was transformed into Escherichia
coli DH5α competent bacteria (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocol, and Luria broth (LB)-agar
plates containing 100 µg/ml ampicillin were employed to select the
positive clones. Upon transformation, the positive clones were
identified by PCR using the PrimeScript™ RT-PCR kit and the
following cycling conditions: Pre-denaturation at 94°C for 5 min;
30 cycles at 94°C for 30 sec, 55°C for 30 sec and 72°C for 1 min;
and 72°C for 10 min.
The E. coli clones were cultured in LB medium
at 37°C, and the plasmid was extracted using the QIAprep Spin
Miniprep kit according to the manufacturer's protocol (Qiagen
GmbH). The VEGF165 gene was then amplified by PCR using the Prime
Script™ RT-PCR kit to identify the correct recombinant plasmids.
The cycling conditions were: Pre-denaturation at 94°C for 5 min; 30
cycles at 94°C for 30 sec, 55°C for 30 sec and 72°C for 1 min; and
72°C for 10 min. The CMV-F and pEGFP-N-3 primers were used to set
up the PCR reaction, and their sequences are indicated in Table I. The positive bacteria colonies were
inoculated into LB medium at 37°C for 16 h, and upon adding
glycerol, they were stored at −80°C. Aliquots (200 µl) of each were
sent for DNA sequencing by Shanghai Sbio-bio Biotechnology Co. Ltd.
(Shanghai, China).
 | Table I.Primers used for VEGF165 gene fragment
amplification and RT-qPCR. |
Table I.
Primers used for VEGF165 gene fragment
amplification and RT-qPCR.
| Primer | Sequence |
|---|
|
VEGF165-EcoRI-Forward |
5′-CTCAAGCTTCGAATTCGCCACCATGAACTTTCTGCTGTCTTGG-3′ |
|
VEGF165-EcoRI-Reverse |
5′-CATGGTGGCGAATTCCCGCCTCGGCTTGTCACA-3′ |
| CMV-F |
5′-CGCAAATGGGCGGTAGGCGTG-3′ |
| pEGFP-N3 |
5′-CGTCGCCGTCCAGCTCGACCAG-3′ |
| EGFP-Forward |
5′-CCTTTCCGGGACTTTCGCTTT-3′ |
| EGFP-Reverse |
5′-GCAGAATCCAGGTGGCAACA-3′ |
|
β-actin-Forwarda |
5′-AAGAGAGGCATCCTCACCCT-3′ |
|
β-actin-Reversea |
5′-TACATGGCTGGGGTGTTGAA-3′ |
|
β-actin-forwardb |
5′-GGAGATTACTGCCCTGGCTCCTA-3′ |
|
β-actin-Reverseb |
5′-GACTCATCGTACTCCTGCTTGCTG-3′ |
| VEGF165-Forward |
5′-ATCTTCAAGCCATCCTGTGTGC-3′ |
| VEGF165-Reverse |
5′-CAAGGCCCACAGGGATTTTC-3′ |
Transfection of the VEGF165
recombinant lentiviral plasmid in 293T cells
At 24 h prior to transfection, 293T cells growing in
the logarithmic phase were selected and propagated. The cell number
was adjusted to 6×105 cells/ml with 10% FBS DMEM. The
cells were transfected at 60–70% confluence, and 2 h prior to
transfection, the medium was changed to serum-free medium. The
recombinant pLVX-VEGF165-EGFP-3FLAG vector and three packaging
components (pRSV-REV, pMDLg-pRRE and pMD2.G) DNAs (Addgene, Inc.,
Cambridge, MA, USA) were added into a sterile centrifuge tube and
mixed with an appropriate volume of Gibco™ Opti-MEM™ (Thermo Fisher
Scientific, Inc.). The 293T cells were then co-transfected with the
above DNAs using Lipofectamine 2000 according to the manufacturer's
protocol (Thermo Fisher Scientific, Inc.). At 8 h
post-transfection, the medium was replaced with complete culture
medium, and the cells were continuously cultured for 48 h.
Subsequently, the supernatant containing lentivirus particles was
harvested and concentrated by ultracentrifugation at 4,000 ×
g for 20 min at 4°C to obtain a high-titer lentivirus
concentration. 293T cells were used to measure the supernatant
virus titer.
Western blot analysis of VEGF165
expression in 293T cells
Transfected and non-transfected 293T cells were
harvested, and proteins were extracted using
radioimmunoprecipitation assay lysis buffer (Pioneer Biotech, Co.,
Ltd., Shaanxi, China) containing protease inhibitors for 30 min on
ice, and then cleared at 25,000 × g for 20 min at 4°C. The
protein concentration was determined using the Bradford assay
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany). Upon
denaturation by heating at 100°C for 5–10 min, equivalent amounts
of total cellular protein were subjected to reducing sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (8–12%), followed by
blotting on a polyvinylidene difluoride membrane. After blocking in
5% non-fat dry milk for 2 h, membranes were incubated with the
mouse anti-VEGF monoclonal antibody (ab1316, 1:100; Abcam,
Cambridge, UK), the mouse anti-GFP polyclonal antibody (sc-9996,
1:3,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and the
mouse anti-β-actin monoclonal antibody (sc-69879, 1:1,000; Santa
Cruz Biotechnology, Inc.) overnight at 4°C. The membranes were then
washed with TBST three times for 5 min each, and incubated with a
horseradish peroxidase (HRP)-conjugated secondary antibody (Santa
Cruz Biotechnology, Inc.) for 2 h at room temperature and
visualized by enhanced chemiluminescence (GE Healthcare Life
Sciences, Chalfont, UK). Images were documented by a scanner,
quantified using Pro-Plus 6.0 software (Media Cybernetics, Inc.,
Rockville, MD, USA) and analyzed using ImageJ 1.49 software
(National Institutes of Health, Bethesda, MD, USA).
Detection of recombinant lentiviral
titer by quantitative (q)PCR
Viral titers were measured by qPCR, using β-actin as
an internal control. The quantification cycle (Cq) value was
defined as the number of cycles when the fluorescent signal reached
a specified threshold, as described previously (21). The sequences of the primers used are
presented in Table I.
Briefly, 293T cells (1×105) were placed
into each well of a 6-well plate for 24 h culture, and 10 µl of a
virus stock solution was added into an Eppendorf tube containing 90
µl of cell culture medium, mixed and diluted 10X. Three different
concentrations were added separately to the wells where the 293T
cells were cultured. After 48-h culture, complete culture medium
was added, and 4 days later, the fluorescence expression was
examined. Total RNA was extracted from the cells with TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). The reverse
transcription (RT) reaction was then performed for complementary
(c)DNA synthesis using the PrimeScript First Strand cDNA Synthesis
kit (Takara Biotechnology Co., Ltd.). qPCR was performed using the
SYBR Green PCR kit (Takara Biotechnology Co., Ltd.) and the
following cycling conditions: Pre-denaturation at 95°C for 15 sec,
followed by 40 cycles at 95°C for 5 sec, 60°C for 30 sec and 60°C
for 30 sec.
Lentivirus infection of MCF-7
cells
Concentrated lentivirus solutions of LV-EGFP and
LV-VEGF165-EGFP were added separately into two wells of cultured
MCF-7 cells once the cells reached 40% confluence. Enhanced
infection solution (Shanghai Sbo-bio Biotechnology) was then added
to reach a total incubation volume of 2 ml. After 12 h of
incubation, the cell culture medium was changed, and 72 h later,
cells expressing EGFP were imaged under an inverted fluorescence
microscope (Eclipse Ti-E; Nikon Corporation, Tokyo, Japan). Images
were processed using NIS-Elements Basic Research Imaging Software
(Nikon Corporation).
RT-qPCR and western blot analysis of
VEGF165 expression in vitro
Stably transfected MCF-7 cells were selected by
limiting dilution, and the expression levels of VEGF165 in the
cells were measured by RT-qPCR and western blotting. Total RNA and
total protein were extracted from MCF-7 cells transfected with
LV-EGFP and LV-VEGF165-EGFP. Western blotting was performed as
described above, and the target proteins were hybridized with mouse
anti-VEGF monoclonal antibody (1:200, Abcam), mouse anti-GFP
(1:1,000, ProteinTech Group, Inc., Chicago, IL, USA), mouse
anti-β-actin antibody (1:5,000, ProteinTech Group, Inc.) and
HRP-conjugated goat anti-mouse immunoglobulin G (1:10,000, CWBIO,
Beijing, China). Enhanced chemiluminescent substrates (SuperSignal™
West Femto Maximum Sensitivity Substrate; Pierce; Thermo Fisher
Scientific, Inc.) were used to detect the signals of targeted
proteins.
Statistical analyses
All statistical analyses were performed using SPSS
16.0 software (SPSS, Inc., Chicago, IL, USA). Data are expressed as
the mean ± standard error of the mean. One way analysis of variance
was used for comparisons among three groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
pLVX-VEGF165-EGFP-3FLAG plasmid
construction
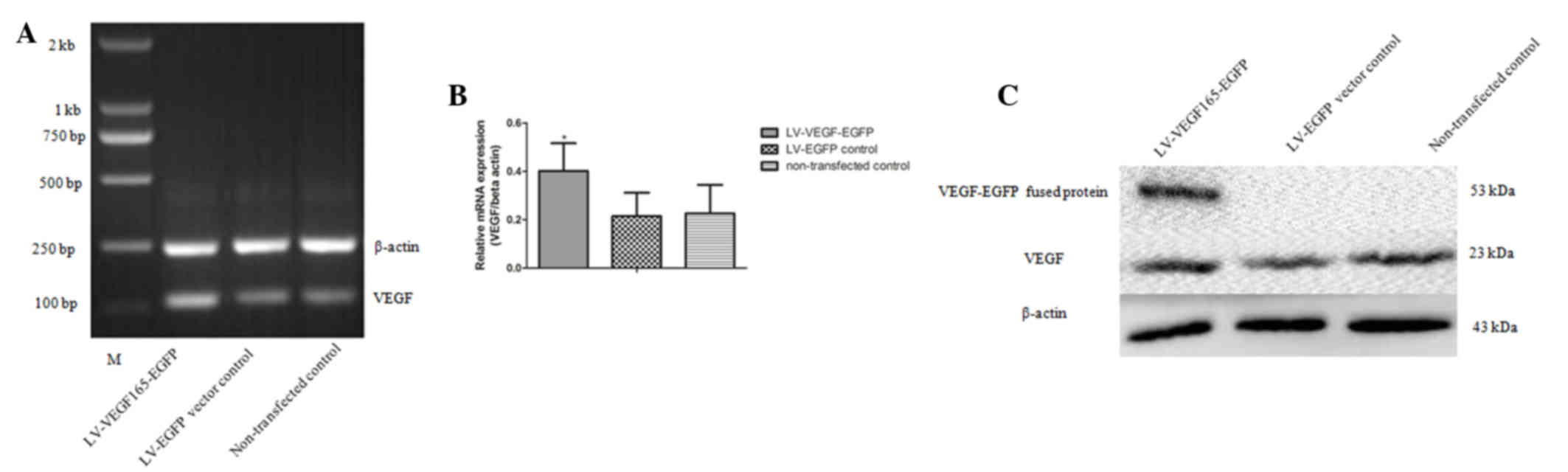
The expected PCR product size of 611 bp for the
VEGF165 coding sequence was obtained (Fig. 2A), and the PCR products of VEGF165 and
the pLVX-EGFP-3FLAG plasmid were then digested separately with
EcoRI (Fig. 2B), followed by
construction of the pLVX-VEGF-EGFP-3FLAG vector via In-Fusion™
Enzyme ligation (Clontech Laboratories, Inc.). Competent E.
coli DH5α cells were transformed with the above plasmid, and
three positive bacteria clones containing a fusion gene composed of
the VEGF165 coding sequence and a fragment of the EGFP gene
sequence were identified using PCR, which detected a product of 837
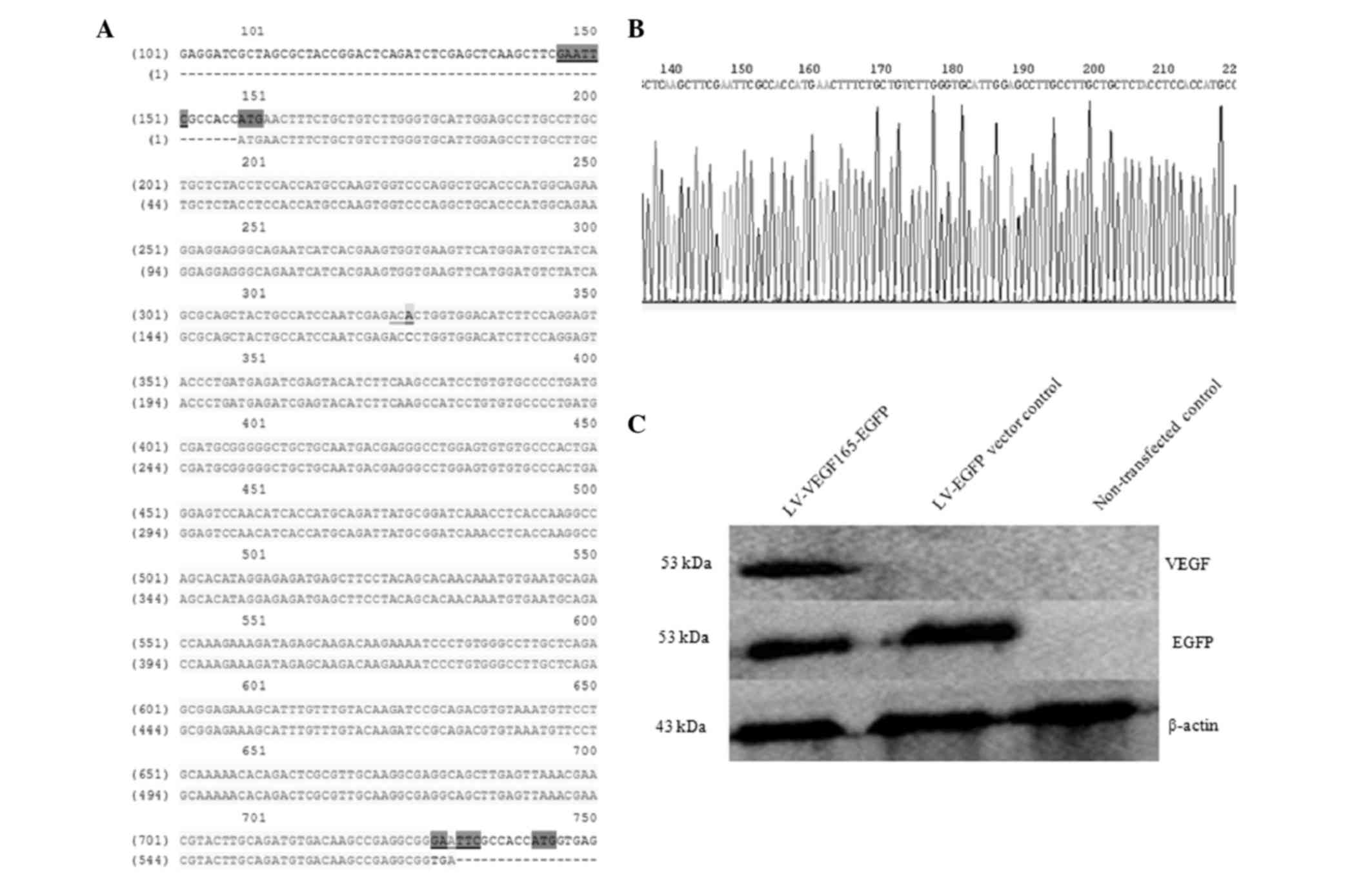
bp in size (Fig. 2C). The sequence of
the recombinant plasmid pLVX-VEGF165-EGFP-3FLAG was confirmed by
sequencing, and the VEGF165 sequence length was 611 bp. This
sequence was consistent with that of the human VEGF165 gene
published in the GenBank® database (http://www.ncbi.nlm.nih.gov/nuccore/NM_001171626.1),
with the exception of a synonymous mutation from ACC to ACA, which
is not expected to influence the protein synthesis (Fig. 3A and B).
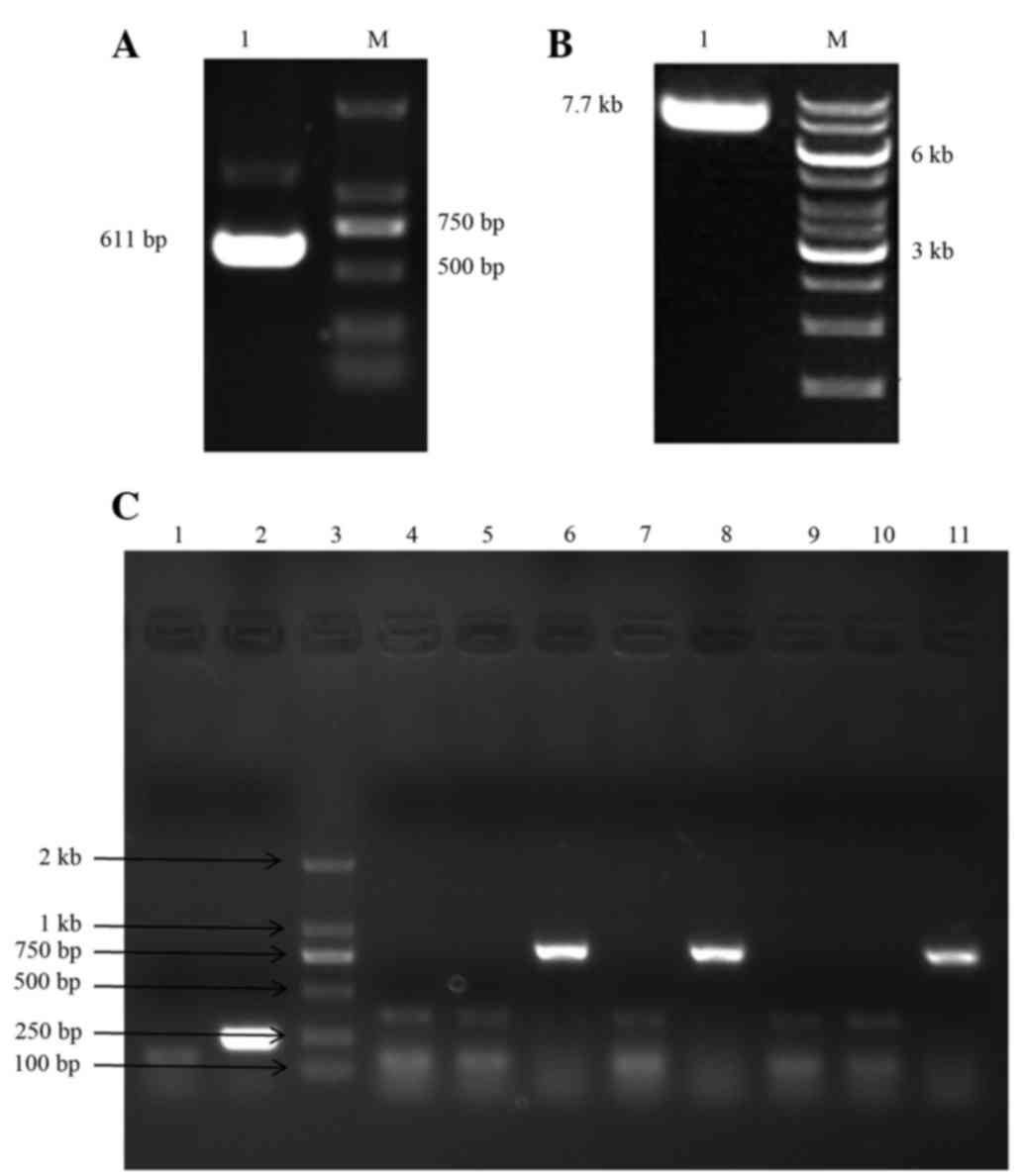 | Figure 2.(A) PCR product for the VEGF165 coding
sequence (611 bp). (B) Linearized pLVX-EGFP-3FLAG plasmid upon
digestion with EcoRI (7.7 kb). (C) The
pLVX-VEGF165-EGFP-3FLAG recombinant expression vector was
identified in bacterial clones by PCR. Lane 1, blank control group;
lane 2, negative control group (LV-EGFP vector, 252 bp); lane 3,
DL2000 DNA Marker; lanes 4–11: eight
pLVX-VEGF165-EGFP-3FLAG-transformed clones, of which, clones 6, 8
and 11 were positive clones (837-bp band). M, marker; LV,
lentivirus vector; VEGF, vascular endothelial growth factor; EGFP,
enhanced green fluorescent protein; PCR, polymerase chain
reaction. |
Transfection of the VEGF165
recombinant lentiviral plasmid in 293T cells, and expression of
EGFP and VEGF165
The recombinant pLVX-VEGF165-EGFP-3FLAG vector and
three packaging components, pRSV-REV, pMDLg-pRRE and pMD2.G DNAs,
were co-transfected in 293T cells. The expression of EGFP and
VEGF165 was detected by western blotting. When incubated with an
anti-VEGF antibody, the VEGF165 fusion protein at a specific band
size of 53 kDa was uniquely observed in VEGF165 recombinant
lentiviral-transfected 293T cells, while expression of the EGFP
fusion protein (53 kDa) was observed in both the LV-VEGF165-EGFP
group and the LV-EGFP group if incubated with an anti-GFP antibody.
This band was consistent with the expected size of the VEGF165-EGFP
fusion protein, indicating that the VEGF165 gene fused with the
EGFP gene, and both could be co-expressed in 293T cells.
Furthermore, the aforementioned band was not detected in
non-transfected cells (Fig. 3C).
VEGF165 recombinant lentiviral
packaging and determination of virus titer
To examine the virus titer of each group, qPCR was
performed. The difference of Cq values was compared between the
control and the test group in order to determine the titer
concentration. Cq values >2.0 were considered to be
significantly different. The titer of recombinant lentivirus was
5.44×107 TU/ml in the LV-VEGF165-EGFP group and
5.00×108 TU/ml in the LV-EGFP negative control group, as
calculated according to the following formula: Virus titer (IU/ml)
= (CxNxDx1,000)/V, where V is the volume of the diluted virus
solution in µl, C is the number of virus copies integrated in each
cell genome, N is the cell number and D is the dilution of the
virus solution (Table II).
| Table II.Virus titers in the LV-VEGF165-EGFP
and LV-EGFP groups. |
Table II.
Virus titers in the LV-VEGF165-EGFP
and LV-EGFP groups.
| Group | V | C | N | D | Virus
titera | Average titer |
|---|
| LV-VEGF165-EGFP
group |
|
|
|
|
|
|
| 10
µl | 10.0 | 0.60 |
2×105 | 1 |
1.20×107 |
5.44±4.20×107 |
| 1
µl |
1.0 | 0.28 |
2×105 | 1 |
5.54×107 |
|
|
10−1 µl |
0.1 | 0.05 |
2×105 | 1 |
9.60×107 |
|
| LV-EGFP group |
|
|
|
|
|
|
| 10
µl | 10.0 | 15.56 |
2×105 | 1 |
3.12×108 |
5.00±1.85×108 |
| 1
µl |
1.0 | 3.41 |
2×105 | 1 |
6.82×108 |
|
|
10−1 µl |
0.1 | 0.25 |
2×105 | 1 |
5.06×108 |
|
Observation of cellular localization
of VEGF165 expression in MCF-7 cells by fluorescence
microscopy
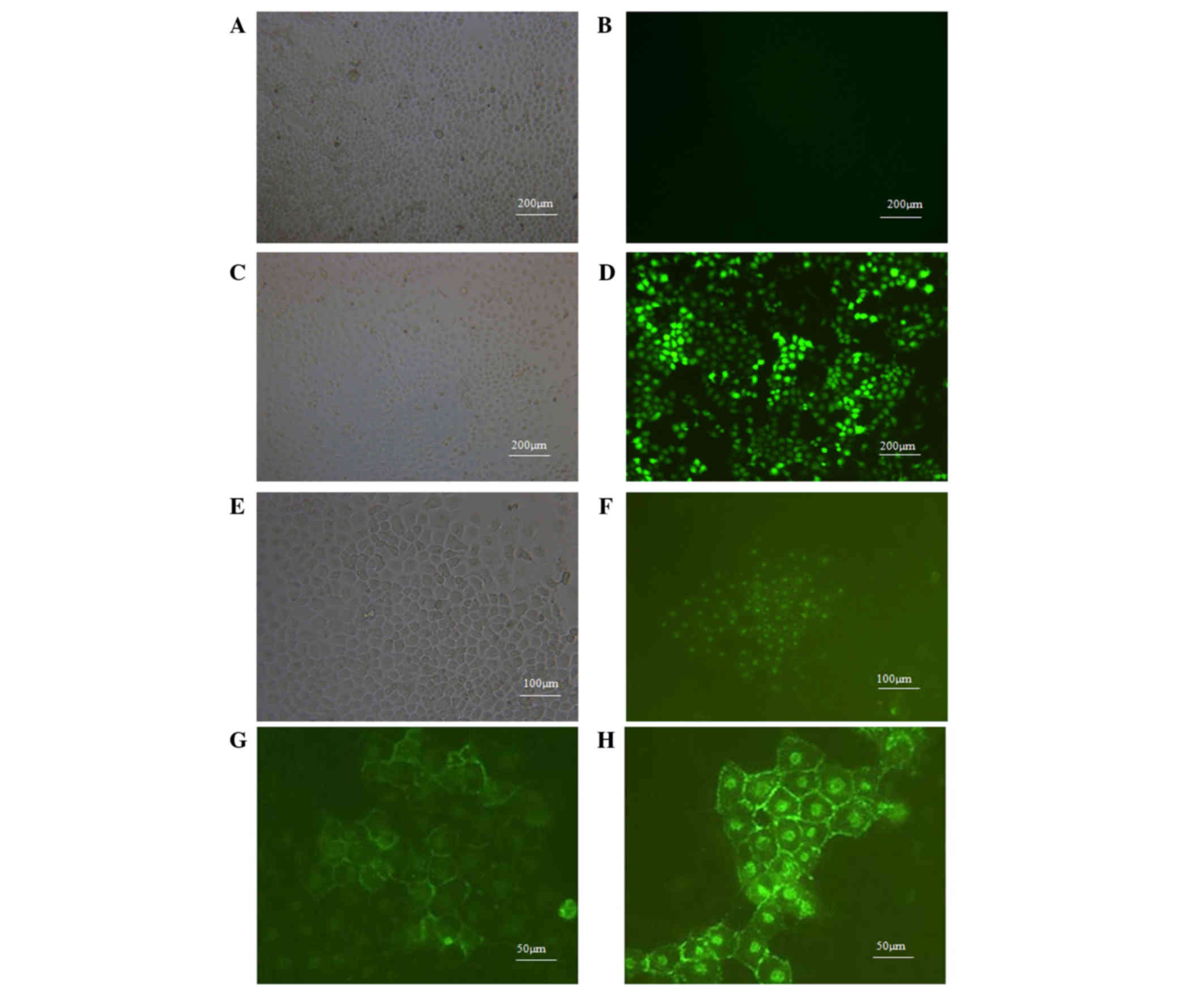
After 72 h of transfection, MCF-7 cells were
observed under a fluorescence microscope to assess the expression
of EGFP. Both LV-EGFP and LV-VEGF165-EGFP groups demonstrated
transfection efficiencies >80%. As represented in Fig. 4A-F, EGFP was highly expressed in MCF-7
cells transfected with LV. By contrast, no EGFP signal was detected
in non-transfected cells. Fluorescence microscopy was used to
observe the cellular localization of VEGF165 in MCF-7 cells. As
represented in Fig. 4G and H,
VEGF165-EGFP was mainly expressed in the cell membrane and nucleus
of MCF-7 cells, and certain expression was also noticed outside the
cell membrane. This indicates that the VEGF165-EGFP fusion protein
could be expressed and secreted normally.
Detection of the VEGF165 expression in
MCF-7 cell lines by RT-qPCR and western blotting
RT-qPCR and western blotting were applied to detect
the RNA and protein expression levels of VEGF165-EGFP,
respectively. RT-qPCR demonstrated that the expression of VEGF165
in the LV-VEGF165-EGFP group was significantly increased (relative
gray value of PCR, 0.4019±0.1143 and 0.2147±0.0965 in the
LV-VEGF165-EGFP and LV-EGFP groups, respectively; Fig. 5A and B). Similarly, compared with
non-transfected cells, a characteristic band of 53 kDa was observed
by western blotting in MCF-7 cells transfected with
LV-VEGF165-EGFP, whose size was consistent with that of the
VEGF165-EGFP fusion protein (Fig.
5C), indicating that the VEGF165-EGFP fusion gene in the
recombinant LV could be expressed following transfection into MCF-7
cells.
Discussion
It has been reported that different isoforms of VEGF
are detected in individual tissues; however, VEGF165 is the most
abundant one in almost all human organs with the exception of the
lung (22). Several studies revealed
that VEGF165 was highly expressed in breast cancer tissues compared
with adjacent tissues (23,24). RT-qPCR experiments demonstrated that
four isoforms of VEGF were expressed in human breast cancer
tissues, including VEGF121, VEGF145, VEGF165 and VEGF189, being
VEGF165 the most abundant one (25).
In addition, previous studies demonstrated that VEGF165 fused to
GFP at its C-terminus was secreted and biologically active
(26). Therefore, in order to
elucidate the function of tumor cell derived-VEGF in breast cancer
biology, VEGF165 was selected as the target gene in the present
study, and the VEGF165-EGFP fusion gene expression vector was
constructed.
As a newly developed technology, recombinant
expression through viral vectors have been used in numerous
laboratory experiments and clinical trials (27–29). There
are mainly five types of viral vectors that are commonly used for
recombinant expression, namely adenovirus, adeno-associated virus,
herpes simplex virus, retrovirus and lentivirus (14). Due to the wide range of host cells,
the ability to infect both replicating and non-replicating cells,
the easy integration of the exogenous gene in the host cells, and
the long-lasting and stable expression of exogenous genes,
lentivirus was the most suitable type of vector for our experiments
(14,30). In the present study, a VEGF165-EGFP
fusion gene recombinant lentiviral expression vector was first
constructed. Using molecular cloning techniques such as PCR and DNA
sequencing, the human VEGF165-EGFP fusion gene lentiviral
expression plasmid was successfully constructed, and high-titer
viral particles were obtained following plasmid transfection in
packaging cells. Due to its safety and high transfection efficiency
(31), a four-plasmid lentivirus
packaging system was employed in the current study, including a
carrier plasmid containing the VEGF165-EGFP fusion gene, a
packaging plasmid pRSV-Rev encoding Rev response element (RRE), a
packaging plasmid pMDLg-pRRE containing the Gag-Pol coding
sequence, and a packaging plasmid pMD2.G encoding the vesicular
stomatitis virus (VSV)-G envelope protein. In this packaging
system, the RRE and Gag-Pol coding sequences are located in two
different plasmids, which greatly reduces the possibility of
autonomous viral replication (31,32).
Furthermore, the human immunodeficiency virus-derived Env gene was
replaced by the VSV-G gene in our packaging system, which
significantly increased the range of host cell types that could be
infected.
The VEGF165-EGFP expression rate was >80% in
infected human breast cancer cells MCF-7. Furthermore, it was
observed that VEGF165 was mainly located on the cell membrane and
nucleus of MCF-7 cells by fluorescence microscopy. The constructed
VEGF165 lentiviral recombinant vector will be used in future
studies to evaluate the function of the target gene, following its
stable transfection in tumor cell lines. This will enable a
detailed examination of the mechanism of tumor cell derived-VEGF165
in tumor cells and in the tumor microenvironment. Besides, this
fusion gene expression vector may also provide a potential approach
for gene therapy in diseases that require regulation of
angiogenesis.
Acknowledgements
The present study was supported by Specialized
Research Fund for the Doctoral Program of Higher Education of China
(Beijing, China; grant no. 20100201110059).
References
|
1
|
Lichtenberger BM, Tan PK, Niederleithner
H, Ferrara N, Petzelbauer P and Sibilia M: Autocrine VEGF signaling
synergizes with EGFR in tumor cells to promote epithelial cancer
development. Cell. 140:268–279. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ribatti D, Nico B, Crivellato E, Roccaro
AM and Vacca A: The history of the angiogenic switch concept.
Leukemia. 21:44–52. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roskoski R Jr: Vascular endothelial growth
factor (VEGF) signaling in tumor progression. Crit Rev Oncol
Hematol. 62:179–213. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ferrara N and Davis-Smyth T: The biology
of vascular endothelial growth factor. Endocr Rev. 18:4–25. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
MacGabhann F and Popel AS: Systems biology
of vascular endothelial growth factors. Microcirculation.
15:715–738. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vempati P, Popel AS and Mac GF:
Extracellular regulation of VEGF: Isoforms, proteolysis and
vascular patterning. Cytokine Growth Factor Rev. 25:1–19. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Otrock ZK, Makarem JA and Shamseddine AI:
Vascular endothelial growth factor family of ligands and receptors:
Review. Blood Cells Mol Dis. 38:258–268. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Knizetova P, Ehrmann J, Hlobilkova A,
Vancova I, Kalita O, Kolar Z and Bartek J: Autocrine regulation of
glioblastoma cell-cycle progression, viability and radioresistance
through the VEGF-VEGFR2 (KDR) interplay. Cell Cycle. 7:2553–2561.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chakraborty G, Jain S and Kundu GC:
Osteopontin promotes vascular endothelial growth factor-dependent
breast tumor growth and angiogenesis via autocrine and paracrine
mechanisms. Cancer Res. 68:152–161. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hamerlik P, Lathia JD, Rasmussen R, Wu Q,
Bartkova J, Lee M, Moudry P, Bartek J Jr, Fischer W, Lukas J, et
al: Autocrine VEGF–VEGFR2–Neuropilin-1 signaling promotes glioma
stem-like cell viability and tumor growth. J Exp Med. 209:507–520.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vannucci L, Lai M, Chiuppesi F,
Ceccherini-Nelli L and Pistello M: Viral vectors: A look back and
ahead on gene transfer technology. New Microbiol. 36:1–22.
2013.PubMed/NCBI
|
|
15
|
Nathwani AC, Tuddenham EGD, Rangarajan S,
Rosales C, McIntosh J, Linch DC, Chowdary P, Riddell A, Pie AJ,
Harrington C, et al: Adenovirus-associated virus vector-mediated
gene transfer in hemophilia b. N Engl J Med. 365:2357–2365. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Giacca M and Zacchigna S: Virus-mediated
gene delivery for human gene therapy. J Control Release.
161:377–388. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang S and Kamihira M: Development of
hybrid viral vectors for gene therapy. Biotechnol Adv. 31:208–223.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lehto T, Simonson OE, Mager I, Ezzat K,
Sork H, Copolovici D, Viola JR, Zaghloul EM, Lundin P, Moreno PM,
et al: A peptide-based vector for efficient gene transfer in vitro
and in vivo. Mol Ther. 19:1457–1467. 2011. View Article : Google Scholar
|
|
19
|
Mingozzi F and High KA: Therapeutic in
vivo gene transfer for genetic disease using AAV: Progress and
challenges. Nat Rev Genet. 12:341–355. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heilbronn R and Weger S: Viral vectors for
gene transfer: Current status of gene therapeutics. Handb Exp
Pharmacol. 143–170. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang S, Zeng X, Liu Y, Liang C, Zhang H,
Liu C, Du W and Zhang Z: Construction and characterization of a
PDCD5 recombinant lentivirus vector and its expression in tumor
cells. Oncol Rep. 28:91–98. 2012.PubMed/NCBI
|
|
22
|
Robinson CJ and Stringer SE: The splice
variants of vascular endothelial growth factor (VEGF) and their
receptors. J Cell Sci. 114:853–865. 2001.PubMed/NCBI
|
|
23
|
Ghosh S, Sullivan CA, Zerkowski MP,
Molinaro AM, Rimm DL, Camp RL and Chung GG: High levels of vascular
endothelial growth factor and its receptors (VEGFR-1, VEGFR-2,
neuropilin-1) are associated with worse outcome in breast cancer.
Hum Pathol. 39:1835–1843. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoshiji H, Gomez DE, Shibuya M and
Thorgeirsson UP: Expression of vascular endothelial growth factor,
its receptor, and other angiogenic factors in human breast cancer.
Cancer Res. 56:2013–2016. 1996.PubMed/NCBI
|
|
25
|
Goel HL and Mercurio AM: VEGF targets the
tumour cell. Nat Rev Cancer. 13:871–882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guzmán-Hernández ML, Potter G, Egervári K,
Kiss JZ and Balla T: Secretion of VEGF-165 has unique
characteristics, including shedding from the plasma membrane. Mol
Biol Cell. 25:1061–1072. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kotterman MA and Schaffer DV: Engineering
adeno-associated viruses for clinical gene therapy. Nat Rev Genet.
15:445–451. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mahn M, Ron S and Yizhar O: Viral
vector-based techniques for optogenetic modulation in vivoViral
Vector Approaches in Neurobiology and Brain Diseases. Brambilla R:
Humana Press; Totowa, NJ: pp. 289–310. 2014, View Article : Google Scholar
|
|
29
|
Barrett DM, Singh N, Porter DL, Grupp SA
and June CH: Chimeric antigen receptor therapy for cancer. Annu Rev
Med. 65:333–347. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Miller AD and Rosman GJ: Improved
retroviral vectors for gene transfer and expression. Biotechniques.
7:980–990. 1989.PubMed/NCBI
|
|
31
|
Maetzig T, Galla M, Baum C and Schambach
A: Gammaretroviral vectors: Biology, technology and application.
Viruses. 3:677–713. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Burns JC, Friedmann T, Driever W,
Burrascano M and Yee JK: Vesicular stomatitis virus G glycoprotein
pseudotyped retroviral vectors: concentration to very high titer
and efficient gene transfer into mammalian and nonmammalian cells.
Proc Natl Acad Sci U S A. 90:8033–8037. 1993. View Article : Google Scholar : PubMed/NCBI
|