Introduction
Protein arginine methyltransferases (PRMTs) are one
class of epigenetic regulator that have come under investigation
recently for their potential roles as diagnostic biomarkers and
therapeutic targets in various cancer types and subtypes. PRMT1, in
particular, has been investigated for the correlation of its
expression with various cancer types, its role in oncogenesis and
its ability to act as a prognostic biomarker (1–6). For
example, prostate tumor grade has been identified to correlate
positively with histone 4 arginine 3 (H4R3) methylation, which is
mediated by PRMT1 (7–9). In addition to PRMT1, PRMT4 (also termed
coactivator-associated arginine methyltransferase 1) (10,11), PRMT5
(12) and PRMT6 (2) have also been studied with respect to
their presence and activities in various cancer types. PRMT8, which
was first described in 2005, has been the subject of limited study,
despite the understanding that it contains a high level of
structural homology (>80% sequence identity) to PRMT1, a
well-studied PRMT family member with strong correlative and
mechanistic ties to cancer (13).
Particular PRMT protein isoforms have been
investigated in order to determine the specific roles of their
protein products in cancer, suggesting the possibility for the use
of PRMT transcript variants as cancer biomarkers (3,14–16). Our previous study (17) demonstrated the endogenous expression
of a novel transcript variant of PRMT8 in long-lived,
fibroblast growth factor 2 (FGF2)-treated primary human dermal
fibroblasts cultured in low oxygen. This study also demonstrated
that knockdown of PRMT8 was sufficient to halt proliferation and
cause cell death in primary human dermal fibroblasts and U87MG
glioblastoma cells, suggesting that PRMT8 expression is necessary
for cellular viability and/or proliferation in healthy and
neoplastic cells. Given the absence of studies investigating PRMT8
expression in association with cancer diagnosis or prognosis, the
high homology of PRMT8 to the comparatively well-studied PRMT1, and
the identification of a novel PRMT8 variant that brings to mind the
isoform-specific effects of PRMT1 that are currently being teased
out in various biological contexts, the present study set out to
explore the potential for use of PRMT8 as a cancer biomarker, and
to investigate the possibility of variant-specific expression and
the effects of PRMT8 in distinct cancer subtypes.
Materials and methods
Bioinformatics analysis
The cancer tissue atlas of the Human Protein Atlas
(HPA; proteinatlas.org) (18) was utilized to generate a list of
cancer types detailing the expression level of PRMT8. PRMT8
staining was performed with the rabbit anti-PRMT8 primary antibody
(cat. no. HPA039747; Sigma-Aldrich; Merck Millipore, Darmstadt,
Germany) (18). HPA is a public
database that curates histological images of 44 normal human
tissues and the 20 most common types of cancer. In total, 216
cancer samples were used to generate profiles of various proteins
using immunohistochemistry. Protein expression profiles were
generated by staining samples from 44 normal human tissues, 20
different cancer types, 46 human cell lines and 6 patient-derived
hematopoietic cell types. Pathologists annotated individual image
files using internal annotation software by scoring the intensity
of staining, percentage positivity and staining localization. The
HPA was accessed in October 2015 using version 13.
Kaplan-Meier plots were used to assess survival
differences at the gene expression level using the Kaplan-Meier
Plotter (KM Plotter; kmplot.com)
(19). KM Plotter is an integrative
data analysis tool that curates gene expression data from
Affymetrix microarrays from the Gene Expression Omnibus (National
Center for Biotechnology Information, Bethesda, MD, USA), the
European Genome-phenome Archive (European Bioinformatics Institute,
Hinxton, UK) and The Cancer Genome Atlas (National Cancer Institute
and National Human Genome Research Institute, Bethesda, MD, USA).
KM Plotter is capable of assessing the effect of 70,632 genes on
the survival of 4,142 breast, 2,437 lung, 1,648 ovarian and 1,065
gastric cancer patients. Gene expression and clinical data are
managed on a PostgreSQL server and each database is updated twice
yearly. Prognostic values for specific genes are split into two
groups according to the quartile expression of a proposed
biomarker. Patient cohorts are then compared using a Kaplan-Meier
survival plot and P-values; hazard ratios with 95% confidence
intervals are also calculated. PRMT8 was analyzed by selecting the
median value of PRMT8 expression as the cut off for high and low
PRMT8 groups. A univariate Cox regression analysis was performed to
calculate hazard ratios and P-values. KM Plotter was accessed in
September 2015 for all analyses.
Cell culture
The CRL-2073 human teratocarcinoma cell line was
obtained from American Tissue Culture Collection (Manassas, VA,
USA). The U-2OS human bone osteosarcoma cell line was a gift from
the Billiar lab at the Worcester Polytechnic Institute (Worcester,
MA, USA). The adult human Caco-2 colorectal adenocarcinoma cell
line was a gift from the Weathers lab at the Worcester Polytechnic
Institute. The adult human A172 and U87MG glioblastoma cell lines,
and the MCF-10A, SK-BR-3 and MDA-MB-231 mammary tissue cell lines
were gifts from the Jain lab at the Worcester Polytechnic
Institute. Cells were cultured in medium consisting of Dulbecco's
modified Eagle's medium (DMEM) Ham's F12 (50:50; MediaTech, Inc.,
Manassas, VA, USA) with 10% Fetal Clone III (HyClone; GE Healthcare
Life Sciences, Logan, UT, USA). The DMEM [without L-Glutamine
(L-Gln) or phenol red] was supplemented with 4 mM fresh L-Gln
(MediaTech) prior to use. Cultures were performed in a 37°C
incubator in a humidified environment of 5% CO2 and 19%
O2. Human WA09 embryonic stem cells (WiCell, Madison,
WI, USA) were cultured on mitomycin C-treated mouse embryonic
fibroblasts seeded at 10,000 cells/cm2 onto 0.1% gelatin
coated 6-well plates using 80% Knockout™ DMEM (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 20% Knockout™ serum
replacement supplemented with 2.0 mM L-Gln, 0.055 mM
2-mercaptoethanol and 4 ng/ml FGF2, as recommended by the supplier.
All cells were harvested on day 7.
Reverse transcription-polymerase chain
reaction (RT-PCR)
RNA was isolated using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol and quantified by spectrophotometry
(NanoDrop 2000; NanoDrop Technologies, Thermo Fisher Scientific,
Inc.). In total, 1 µg total RNA was used to perform first strand
cDNA synthesis using qScript™ cDNA SuperMix (Quanta Biosciences,
Beverly, MA, USA). For PCR, 50 ng first-strand cDNA was used as a
template for each reaction. PCR was performed using 12.5 µl GoTaq
(Promega Corporation, Madison, WI, USA) and 0.2 mM each of forward
and reverse primers (primer sequences in Table I). PCR cycling for PRMT8 was
performed as follows: Initial denaturation at 95°C for 2 min,
followed by 35 cycles of denaturation at 94°C for 15 sec, annealing
at primer-specific annealing temperature for 30 sec and extension
at 72°C for 1 min. Final extension was performed at 72°C for 10 min
and samples were held at 4°C until use. PCR cycling for β-actin was
performed as follows: Initial denaturation at 95°C for 5 min,
followed by 30 cycles of denaturation at 95°C for 15 sec, annealing
at primer-specific annealing temperature for 15 sec and extension
at 72°C for 15 sec. Final extension was performed at 72°C for 7 min
and the samples held at 4°C until use. PCR cycling for PRMT8
variant 1: Initial denaturation at 95°C for 5 min, followed by 35
cycles of denaturation at 95°C for 30 sec, annealing at a
primer-specific annealing temperature for 30 sec and extension at
72°C for 30 sec. Final extension was performed at 72°C for 10 min
and the samples held at 4°C until use. PCR cycling for PRMT8
variant 2: Initial denaturation at 95°C for 5 min, followed by 30
cycles of denaturation at 95°C for 15 sec, annealing at
primer-specific annealing temperature for 15 sec and extension at
72°C for 15 sec. Final extension was performed at 72°C for 10 min
and the samples held at 4°C until use. Amplification products were
resolved on 2% agarose gels containing 0.5 µg/ml ethidium bromide
in 1X Tris base, acetic acid and EDTA buffer, and images were
captured using a Gel Doc XR System (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
 | Table I.RT-PCR primer sequences. |
Table I.
RT-PCR primer sequences.
| Primer | Forward sequence
(5–3′) | Reverse sequence
(5–3′) | Amplicon (bp) |
|---|
| PRMT8 |
GACTACGTCCACGCCCTGGTCACCTATTTTATT |
GGTCTCGCACATTTTTGGCATTTGGCTTCATGG | 205 |
| PRMT8 v1 |
AAGGAATCCGGAGCAGATGAGAAG |
GGCATAGGAGTCGAAGTAATAATCTCTC | 458 |
| PRMT8 v2 |
CTGTTTGAATGTGTGCCAGGTTG |
GGCATAGGAGTCGAAGTAATAATCTCTC | 240 |
| β-actin |
TCTGGCACCACACCTTCTACAA |
CTTCTCCTTAATGTCACGCACG | 392 |
Results
PRMT8 is highly expressed in various
types of cancer
To explore the possibility of PRMT8 as a cancer
biomarker, the cancer proteome, as curated by the HPA (18), was assessed for PRMT8 expression.
Protein expression was measured by the HPA using
immunohistochemical staining in patient-derived primary cancer
tissue samples. The HPA analyzed 216 independent cancer samples
from 20 of the most common types of cancer for 16,613 genes. PRMT8
expression was evaluated using a single primary antibody. Of all
the types of human cancer tested, PRMT8 expression was detected in
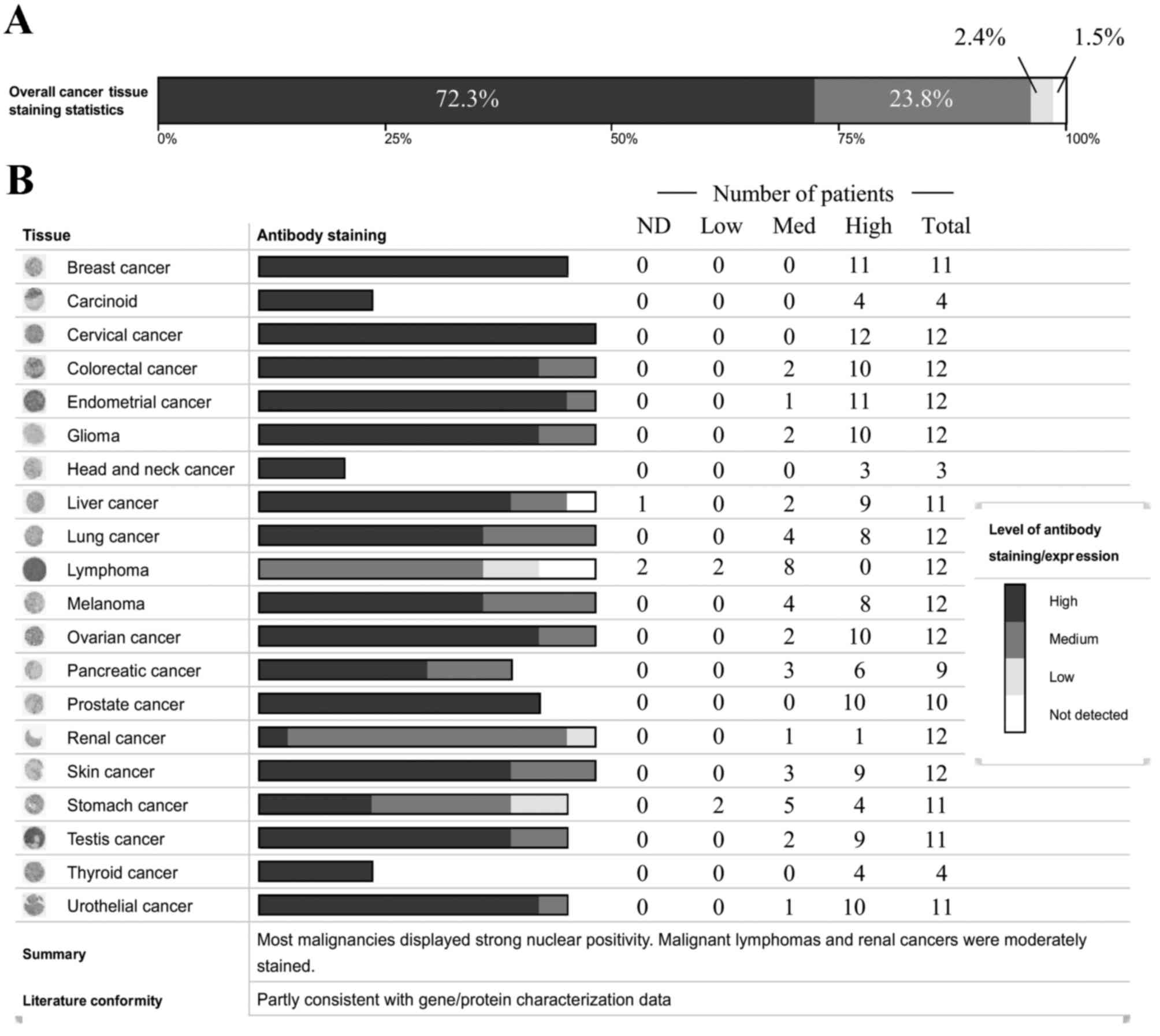
98.5% of the tissue samples (Fig.
1A). PRMT8 is highly expressed in 72.3% of tissues, at moderate
levels in 23.8% and at low-levels in 2.4% of patients analyzed.
Fig. 1B depicts PRMT8 expression
levels categorized by cancer type. From these data, PRMT8 is highly
expressed in breast, glandular, cervical, head and neck, prostate
and thyroid cancer. PRMT8 is expressed at moderate-high levels in
colorectal, endometrial, brain, lung, ovarian, pancreatic, skin,
testicular and urothelial cancer. PRMT8 is expressed at low-high
levels in renal and stomach cancers. PRMT8 expression ranges from
undetectable to high or from undetectable to moderate in liver and
lymphatic cancer, respectively.
Expression of PRMT8 correlates with
patient survival in several cancer types
Ideally, a good biomarker correlates with a
measurable outcome, a certain metric that can be used to
characterize the disease state, such as patient survival. KM
Plotter was used to analyze microarray data from 10,188 cancer
samples (19) for the purpose of
correlating PRMT8 expression with patient survival. A total
of 2 probe sets for PRMT8 were included in the microarray
data curated by the KM Plotter. Probe sets 1 and 2 were used to
test PRMT8 expression levels in 1,660 and 3,554 patients
with breast cancer, respectively. Patients with high levels of
PRMT8 expression were revealed to survive significantly
longer, compared with patients with low PRMT8 expression
(P=8.2×10-3 and P=9.6×10-8, respectively; Fig. 2A and B). This was also the case when
probe set 2 was used to measure PRMT8 expression in patients
with ovarian cancer, where high levels of PRMT8 expression
correlated significantly with increased patient survival (n=1,305;
P=0.014; Fig. 2C). Conversely, when
probe set 2 was used to measure PRMT8 expression in patients
with gastric cancer, high levels of PRMT8 expression were
significantly correlated with decreased patient survival (n=876;
P=1.1×10−9; Fig. 2D). When
probe set 2 was used to measure PRMT8 expression in patients
with non-small-cell lung cancer, no significant correlation was
identified between patient survival and PRMT8 expression
levels (n=1,926; P=0.84; Fig.
2E).
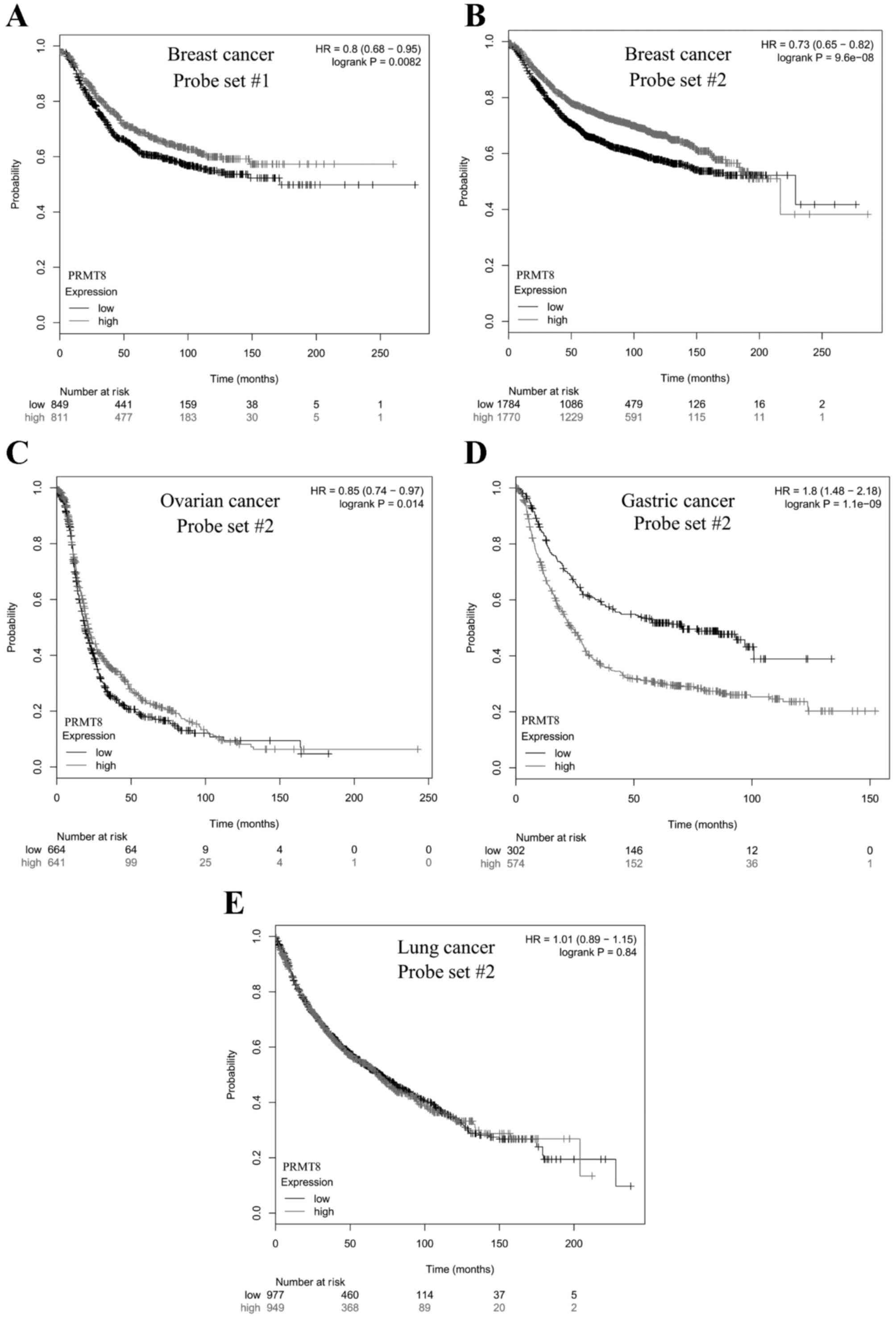 | Figure 2.Kaplan-Meier survival plot of cancer
patients with varying PRMT8 expression. Differences in
PRMT8 expression in breast cancer patients were determined
with Affymetrix (A) probe set #1 or (B-E) #2 using a microarray,
and patient survival was plotted over time. The gray curve consists
of all patients with high PRMT8 expression. The black curve
consists of all patients with low PRMT8 expression. (A) In
1,660 patients with breast cancer analyzed with probe set #1, there
was a significant positive correlation between high PRMT8
expression and patient survival (P=8.2e-03). (B) In 3,554 patients
with breast cancer analyzed with probe set #2, there was a
significant positive correlation between high PRMT8
expression and patient survival (P=9.6e-08). (C) In 1,305 patients
with ovarian cancer analyzed with probe set #2, there was a
significant positive correlation between high PRMT8
expression and patient survival (P=0.014). (D) In 876 patients with
gastric cancer, there was a significant negative correlation
between high PRMT8 expression and patient survival
(P=1.1e-09). (E) In 1,926 patients with non-small cell lung cancer,
there was no significant correlation between high PRMT8
expression and patient survival (P=0.84). Data obtained from
Kaplan-Meier Plotter (kmplot.com)
(19). PRMT8, protein arginine
methyltransferase 8; HR, hazard ratio. |
PRMT8 variant 2 is expressed in the
U87MG tumorigenic glioblastoma cell line
While data from the KM Plotter is useful for
determining whether PRMT8 is a potentially useful biomarker
for specific types of cancer, the effect of variant-specific
expression on patient survival cannot be determined from these
datasets. To gain insight into the variant-specific expression and
function that PRMT8 may have in various types of cancer, the
present study evaluated individual cell lines for specific
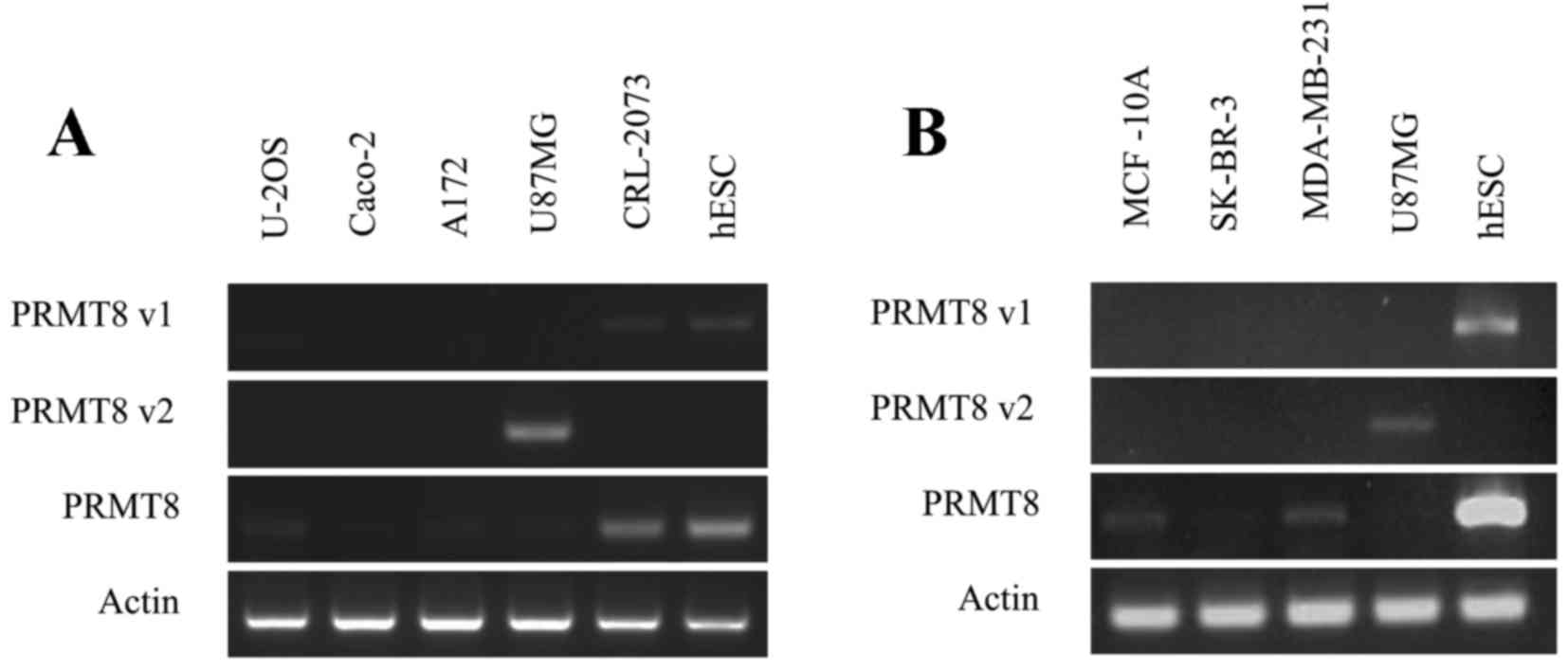
PRMT8 variants using RT-PCR. This preliminary assay revealed
that the U-2OS osteosarcoma, the A172 and U87MG glioblastoma, the
CRL-2073 teratocarcinoma line and the three MCF-10A, SK-BR-3, and
MDA-MB-231 mammary gland cell lines, all express PRMT8 at
varying levels (Fig. 3A and B).
Certain cell lines, including SK-BR-3, expressed PRMT8 at
barely detectable levels under the conditions tested. The Caco-2
colorectal adenocarcinoma line does not express detectable levels
of PRMT8 (Fig. 3A). Of the
cell lines tested, and under the conditions used, only the U87MG
glioblastoma cell line expressed detectable levels of PRMT8
variant 2 (Fig. 3A). The U-2OS
osteosarcoma and the CRL-2073 teratocarcinoma cell line express
detectable levels of PRMT8 variant 1, and the A172
glioblastoma cell line and the aforementioned three mammary gland
lines do note express detectable levels of PRMT8 variant 1
or PRMT8 variant 2 (Fig. 3B).
In Fig. 3A and B, WA09 human
embryonic stem cells (hESCs) were used as a positive control for
PRMT8 expression, as well as a positive control for
PRMT8 variant 1 expression. U87MG cells were used in
Fig. 3B as a positive control for
PRMT8 variant 2 expression.
Discussion
Correlations of PRMT expression with oncogenic
disease states, and the activity of PRMT family members in cancer
cell phenotypes, have been previously studied, generally and in
isoform-specific manners (3,14–16). This
is particularly true of PRMT1, the PRMT family member most
homologous to PRMT8 (13), the
expression of which is amplified in numerous breast cancer tissues
and has been demonstrated to correlate with patient age, tumor
grade and menopausal status (2,3). In lung
and bladder cancer, PRMT1 and 6 demonstrated elevated expression
and regulated cancer cell growth, as knockdown leads to decreased
proliferation in each type of cancer cell line (2). PRMT4 was revealed to be highly expressed
in prostatic intraepithelial neoplasia and in prostatic
adenocarcinoma, compared with benign prostate tissue (10), as well as in colorectal cancer, where
PRMT4 was identified to regulate p53 and nuclear factor-κB target
gene transcription in colorectal adenocarcinoma cells (11).
The present study reports that PRMT8 is expressed at
high levels in numerous types of cancer, which is notable given
that, in mature organisms, PRMT8 tends to be localized to brain
tissue (13), suggesting a role for
PRMT8 dysregulation in oncogenesis or the maintenance of cancer
cell phenotypes. This appears particularly provocative given the
increasing importance attributed to epigenetics in cancer and
cancer therapies (20,21), and in light of our recent study
demonstrating that the knockdown of PRMT8 is sufficient to
kill U87MG glioblastoma cells in vitro (17). The HPA curation of the cancer proteome
demonstrated that PRMT8 is moderately-highly expressed in
various cancerous tissues, including those of non-neural origin
that would not be otherwise expected to express high levels of
PRMT8, comprising breast, glandular, prostate, and thyroid
tissues.
Additionally, the present study reports that high
PRMT8 expression correlates significantly with patient
survival in several different types of cancer, demonstrating a
positive association in patients with breast and ovarian cancer, a
negative correlation in patients with gastric cancer and no
significant correlation in patients with non-small cell lung
cancer. These varied tissue-specific effects may be due to
expression of different PRMT8 transcript variants, as the
probe set data were not variant-specific, preventing uncoupling of
the effects of specific PRMT8 transcript variants on patient
survival. The present study believes that this possibility merits
future investigation into the use of variant-specific PRMT8
expression as a cancer biomarker; much in the same way that
variant-specific expression of PRMT1 has been explored
(3,14–16). It is
worth noting that, even in the absence of available data to
uncouple variant-specific PRMT8 expression and correlation
with patient survival, the potential for total PRMT8
expression to be used as a prognostic biomarker and for
consideration as a possible therapeutic target stands on its own.
This is particularly true in gastric cancer, where the negative
correlation between PRMT8 expression and patient survival
time is considerable: Half of patients with low levels of
PRMT8 expression had succumbed to the disease by ~8 years
post-analysis; whereas half of patients with high levels of
PRMT8 were deceased by ~2 years post-analysis.
The novel transcript variant of PRMT8 that
was recently identified (17),
PRMT8 variant 2, is unique in that its protein product lacks
a portion of the N-terminus of the full-length PRMT8 isoform, which
normally harbors a hydrophobic myristoylation motif that confers
localization to the plasma membrane (13). As the protein product of PRMT8
variant 2 lacks the glycine residue near the N-terminus that is
able to be myristoylated, the protein product of PRMT8
variant 2 instead localizes to the nucleus (Hernandez et al,
unpublished data). This finding is consistent with a previous study
(22), which demonstrated that, while
the full-length isoform of PRMT8 localizes to the plasma membrane,
two truncated isoforms translated from in-frame methionine codons
and lacking the N-terminal glycine residue that may be
myristoylated, demonstrated strong patterns of nuclear
localization. Nuclear localization, rather than anchorage of the
protein into a membrane, appears more fitting for a protein with
suspected epigenetic activity, particularly given the demonstrated
ability of full-length PRMT8 to bind histone H4 and
nucleosome assembly protein 3 in vitro (13). That the protein product of
PRMT8 variant 2 exhibits nuclear localization suggests that
the expression of PRMT8 variant 2 in various types of
cancer, including the demonstrated expression in U87MG glioblastoma
cells, may contribute to the development of the cancer cell
phenotype through epigenetic mechanisms due to its arginine
methyltransferase activity. This hypothesis is supported by a
recent report that PRMT8 positively regulates p53 expression due to
etopside-induced DNA damage in U-2OS osteosarcoma cells (23). Additional investigation into the role
of PRMT8 variants and their protein products in various
cancer cells may reveal information about cancer cell phenotypes,
and may propose other uses for PRMT8 as a cancer biomarker
or even as a therapeutic target.
Acknowledgements
The authors would like to thank Kris Billiar, Anjana
Jain and Pam Weathers at Worcester Polytechnic Institute for their
generous gifts of U-2OS; A172, U87MG, MCF-10A, SK-BR-3 and
MDA-MB-231; and Caco-2 cells, respectively. The present study was
supported by the National Institutes of Health award (grant no.
R01GM85456) and the National Science Foundation Integrative
Graduate Education and Research Traineeship Training Program (grant
no. DGE 1144804).
References
|
1
|
Cheung N, Chan L, Thompson A, Cleary ML
and So C: Protein arginine-methyltransferase-dependent oncogenesis.
Nat Cell Biol. 9:1208–1215. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoshimatsu M, Toyokawa G, Hayami S, Unoki
M, Tsunoda T, Field HI, Kelly JD, Neal DE, Maehara Y, Ponder BA, et
al: Dysregulation of PRMT1 and PRMT6, Type I arginine
methyltransferases, is involved in various types of human cancers.
Int J Cancer. 128:562–573. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mathioudaki K, Scorilas A, Ardavanis A,
Lymberi P, Tsiambas E, Devetzi M, Apostolaki A and Talieri M:
Clinical evaluation of PRMT1 gene expression in breast cancer.
Tumor Biol. 32:575–582. 2011. View Article : Google Scholar
|
|
4
|
Mathioudaki K, Papadokostopoulou A,
Scorilas A, Xynopoulos D, Agnanti N and Talieri M: The PRMT1 gene
expression pattern in colon cancer. Br J Cancer. 99:2094–2099.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Papadokostopoulou A, Mathioudaki K,
Scorilas A, Xynopoulos D, Ardavanis A, Kouroumalis E and Talieri M:
Colon cancer and protein arginine methyltransferase 1 gene
expression. Anticancer Res. 29:1361–1366. 2009.PubMed/NCBI
|
|
6
|
Li B, Liu L, Li X and Wu L: miR-503
suppresses metastasis of hepatocellular carcinoma cell by targeting
PRMT1. Biochem Biophys Res Commun. 464:982–987. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seligson DB, Horvath S, Shi T, Yu H, Tze
S, Grunstein M and Kurdistani SK: Global histone modification
patterns predict risk of prostate cancer recurrence. Nature.
435:1262–1266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Huang ZQ, Xia L, Feng Q,
Erdjument-Bromage H, Strahl BD, Briggs SD, Allis CD, Wong J, Tempst
P and Zhang Y: Methylation of histone H4 at arginine 3 facilitating
transcriptional activation by nuclear hormone receptor. Science.
293:853–857. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Strahl BD, Briggs SD, Brame CJ, Caldwell
JA, Koh SS, Ma H, Cook RG, Shabanowitz J, Hunt DF, Stallcup MR and
Allis CD: Methylation of histone H4 at arginine 3 occurs in vivo
and is mediated by the nuclear receptor coactivator PRMT1. Curr
Biol. 11:996–1000. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hong H, Kao C, Jeng MH, Eble JN, Koch MO,
Gardner TA, Zhang S, Li L, Pan CX, Hu Z, et al: Aberrant expression
of CARM1, a transcriptional coactivator of androgen receptor, in
the development of prostate carcinoma and androgen-independent
status. Cancer. 101:83–89. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim YR, Lee BK, Park RY, Nguyen NT, Bae
JA, Kwon DD and Jung C: Differential CARM1 expression in prostate
and colorectal cancers. BMC Cancer. 10:1972010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pal S, Baiocchi RA, Byrd JC, Grever MR,
Jacob ST and Sif S: Low levels of miR-92b/96 induce PRMT5
translation and H3R8/H4R3 methylation in mantle cell lymphoma. EMBO
J. 26:3558–3569. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee J, Sayegh J, Daniel J, Clarke S and
Bedford MT: PRMT8, a new membrane-bound tissue-specific member of
the protein arginine methyltransferase family. J Biol Chem.
280:32890–32896. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Goulet I, Gauvin G, Boisvenue S and Côté
J: Alternative splicing yields protein arginine methyltransferase 1
isoforms with distinct activity, substrate specificity, and
subcellular localization. J Biol Chem. 282:33009–33021. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong J, Cao RX, Zu XY, Hong T, Yang J,
Liu L, Xiao XH, Ding WJ, Zhao Q, Liu JH and Wen GB: Identification
and characterization of novel spliced variants of PRMT2 in breast
carcinoma. FEBS J. 279:316–335. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scorilas A, Black MH, Talieri M and
Diamandis EP: Genomic organization, physical mapping, and
expression analysis of the human protein arginine methyltransferase
1 gene. Biochem Biophys Res Commun. 278:349–359. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hernandez S and Dominko T: Novel protein
arginine methyltransferase 8 isoform is essential for cell
proliferation. J Cell Biochem. 117:2056–2066. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Györffy B, Lanczky A, Eklund AC, Denkert
C, Budczies J, Li Q and Szallasi Z: An online survival analysis
tool to rapidly assess the effect of 22,277 genes on breast cancer
prognosis using microarray data of 1,809 patients. Breast Cancer
Res Treat. 123:725–731. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dawson MA and Kouzarides T: Cancer
epigenetics: From mechanism to therapy. Cell. 150:12–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sharma S, Kelly TK and Jones PA:
Epigenetics in cancer. Carcinogenesis. 31:27–36. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kousaka A, Mori Y, Koyama Y, Taneda T,
Miyata S and Tohyama M: The distribution and characterization of
endogenous protein arginine N-methyltransferase 8 in mouse CNS.
Neuroscience. 163:1146–1157. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sammons MA, Zhu J and Berger SL: A
chromatin-focused siRNA screen for regulators of p53-dependent
transcription. G3 (Bethesda). 6:2671–2678. 2016. View Article : Google Scholar : PubMed/NCBI
|