Introduction
Pancreatic cancer, one of the most common cancers
globally, is the fourth most common cause of cancer-associated
mortality in the USA (1). In 2015, it
was estimated that there would be 48,960 new pancreatic cancer
cases and 40,560 mortalities due to pancreatic cancer in the USA
(2). Epidemiological studies have
demonstrated that the likelihood of pancreatic cancer is associated
with numerous factors, including smoking, high-fat high-protein
diets, alcoholism, coffee drinking, exposure to certain chemical
carcinogens, diabetes mellitus and chronic pancreatitis (3,4).
Pancreatic ductal adenocarcinoma is the most common subtype of
pancreatic cancer, accounting for 85–90% of all pancreatic cancer
cases (5). Although progress has been
made in surgery and perioperative management, the 5-year relative
survival rate for patients with pancreatic cancer remains at ~5%
and the median survival time is only 6 months (6). The poor prognosis is mainly a result of
early and aggressive local invasion and metastasis, as well as
dissemination of the pancreatic cancer cells (7). Therefore, it is urgent to improve
understanding of the molecular mechanisms underlying the metastasis
of pancreatic cancer, and to investigate more effective therapeutic
treatments for patients with pancreatic cancer to block cancer
metastasis.
MicroRNAs (miRNAs/miRs) are considered to be useful
biomarkers for the early diagnosis, therapy and prognosis of human
cancers, including pancreatic cancer (8–10). They
are an evolutionarily conserved group of non-protein-coding and
short RNAs, 18–22 nucleotides in length, whose function is to
regulate the expression of their target protein-coding genes either
by translational suppression or gene degradation through binding to
the 3′ untranslated regions (3′UTRs) of target genes in a
base-pairing manner (11,12). An increasing number of studies have
demonstrated that miRNAs regulate numerous biological processes,
including cell growth, apoptosis, the cell cycle, metastasis and
metabolism, and consequently, their alterations are considered to
perform important functions in carcinogenesis and progression of
cancers (13). Abnormal expression of
miRNAs is associated with various types of diseases, including
cancer, and tumor-associated miRNAs act as tumor suppressors or
oncogenes depending on their target mRNAs (14). Therefore, elucidating the expression,
function and molecular mechanism of miRNAs will be particularly
useful in investigating novel therapeutic treatments for
cancers.
miR-452 has been studied in several types of cancer,
including non-small cell lung cancer (15), breast cancer (16), bladder cancer (17) and hepatocellular carcinoma (18). However, the expression, biological
function and molecular mechanism of miR-452 in pancreatic cancer
remains to be fully elucidated. The present study revealed that
miR-452 was significantly downregulated in pancreatic cancer
tissues, particularly in metastatic tumors and pancreatic cancer
cell lines. Notably, migration and invasion assays indicated that
overexpression of miR-452 decreased the migration and invasion of
pancreatic cancer cells. Subsequent experiments demonstrated that
BMI1 was a direct target gene of miR-452 in pancreatic cancer.
Collectively, miR-452 suppressed the migration and invasion of
pancreatic cancer cells by directly targeting BMI1.
Materials and methods
Human tissue specimens and ethics
statement
A total of 32 pancreatic cancer tissues with matched
adjacent normal tissues were obtained from patients (male, 19;
female, 13; age, 46–69 years; mean, 57 years) during surgery at
Weifang People's Hospital (Weifang, China) between June 2013 and
March 2015. None of patients underwent chemotherapy or radiotherapy
prior to surgery. All the tissues were immediately snap-frozen and
stored in liquid nitrogen until use. For the analyzed tissue
specimens, written informed consent was obtained from all patients
to use excess pathological specimens for study purposes. The
present study was approved by the Ethics Committee of Weifang
People's Hospital and performed according to the ethical
principles.
Cell culture and cell
transfection
The human pancreatic cancer cell lines (PANC-1,
SW1990, ASPC-1 and CFPAC-1) and the human normal pancreatic cell
line (HPDE6c7) were obtained from American Type Culture Collection
(Manassas, VA, USA) and maintained in Dulbecco's modified Eagle's
medium supplemented with 10% fetal bovine serum (FBS), 2 mM
glutamine, 100 IU/ml penicillin and 100 µg/ml streptomycin (all
from Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA). All
cell lines were cultured in a humidified cell incubator, with an
atmosphere of 5% CO2 and 95% air, at 37°C.
miR-452 mimic and negative control (NC) were
obtained from Shanghai GenePharma Co., Ltd. (Shanghai, China). BMI1
small interfering RNA (siRNA) and NC siRNA were purchased from
Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The miR-452 mimic
sequence was 5′-AACUGUUUGCAGAGGAAACUGA-3′ and the NC sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. The BMI1 siRNA sequence was
5′-GUUCACAAGACCAGACCAC-3′ and the NC siRNA sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. miRNA or siRNA were transfected into
cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's protocol.
The medium was replaced with fresh culture medium 6–8 h after
transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from tissues and cell lines
using TRIzol regent (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. For miRNA expression,
RT-qPCR assays were performed using a TaqMan miRNA Assay in an
Applied Biosystems 7500 Real-time PCR system (both from Applied
Biosystems; Thermo Fisher Scientific, Inc.). For mRNA expression,
cDNA was generated from total RNA through reverse transcription
using M-MLV reverse transcriptase (Promega Corporation, Madison,
WI, USA). BMI1 mRNA expression was evaluated with SYBR-Green Master
Mix (Takara Biotechnology Co., Ltd., Dalian, China). The
thermocycling conditions were as follows: 95°C for 10 min, followed
by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The primers
were designed as follows: miR-452, 5′-GCGAACTGTTTGCAGAGG-3′
(forward) and 5′-CAGTGCGTGTCGTGGAGT-3′ (reverse); U6,
5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-ACGCTTCACCGAATTTGAGT-3′
(reverse); BMI1, 5′-GTGCTTTGTGGAGGGTACTTCAT-3′ (forward) and
5′-TTGGACATCACAAATAGGACAATACTT-3′ (reverse); and GAPDH,
5′-GAAGGTGAAGGTCGGAGTC-3′ (forward) and 5′-GAAGATGGTGATGGGATTTC-3′
(reverse). Each sample was analyzed in triplicate. U6 and GADPH
were used as internal controls for miR-452 and BMI1 mRNA
expression, respectively. Relative expression levels were
calculated using the 2−∆∆Cq method (19).
Migration and invasion assay
Transwell chambers (Costar; Corning Incorporated,
Corning, NY, USA) with 8-mm pore size polycarbonate membranes were
applied to examine cell migration and invasion capacity. For the
invasion assay, Transwell chambers were coated with Matrigel (40
µg/well; BD Biosciences, San Jose, CA, USA) prior to the
experiment. In brief, 1×105 transfected cells suspended in 200 µl
serum-free medium were placed in the top chambers and 500 µl
complete culture medium containing 10% FBS was added to the lower
chambers.
Following incubation at 37°C in a cell incubator for
48 h, the cells on the top surface of the Transwell chamber were
gently removed from the top chambers with a cotton swab. Cells on
the lower surface of the membrane were then fixed with 90% methanol
for 10 min and stained with 0.1% crystal violet for 10 min, and
averaged across five random fields at ×100 magnification using an
inverted microscope (Olympus Corporation, Tokyo, Japan).
Target prediction of miRNAs
TargetScan (http://www.targetscan.org/) was adopted to identify
the potential targets of miR-452.
Western blot analysis
Following incubation at 37°C for 72 h
post-transfection, cells were lysed using radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology, Haimen,
China) supplemented with 0.1 mg/ml phenylmethylsulfonyl fluoride, 1
mM sodium orthovanadate and 1 mg/ml aprotinin. Protein
concentration was determined using a bicinchoninic acid assay kit
(Beyotime Institute of Biotechnology). Equal amounts of protein (20
mg) were separated by 10% SDS-PAGE and transferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA) using a Bio-Rad semidry transfer system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). The membranes were then blocked in 5%
skim milk in TBS-Tween-20 (TBST) for 2 h at room temperature,
followed by incubation with the following primary antibodies: Mouse
anti-human monoclonal BMI1 (1:1,000 dilution; cat no. sc-13519) and
mouse anti-human GADPH (1:1,000 dilution; cat no. sc-166574) (both
from Santa Cruz Biotechnology, Inc., Dallas, TX, USA) overnight at
4°C. The membranes were washed three times with TBST and probed
with goat anti-mouse horseradish peroxidase-conjugated secondary
antibody (1:5,000 dilution; cat no. sc-2005; Santa Cruz
Biotechnology, Inc.) at room temperature for 1 h. Membranes were
then washed three times with TBST and visualized with enhanced
chemiluminescence plus reagents (Pierce; Thermo Fisher Scientific,
Inc.). Protein expression was quantified using Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc.). GADPH was used
as an internal control for BMI1 protein expression.
Dual-luciferase reporter assay
PGL3-BMI1-3′UTR wild-type (Wt) and PGL3-BMI1-3′UTR
mutant (Mut) were purchased from Shanghai GenePharma Co., Ltd.
Cells were seeded onto 96-well plates and transfected with miR-452
mimics or NC, and co-transfected with PGL3-BMI1-3′UTR Wt or
PGL3-BMI1-3′UTR Mut using Lipofectamine 2000 according to the
manufacturer's protocol. Following incubation at 37°C for 48 h,
cells were collected and luciferase activities were measured using
a Dual-Luciferase Reporter Assay system (Promega Corporation). The
relative luciferase activities were normalized to that of
Renilla activity.
Statistical analysis
Data are expressed as the mean ± standard deviation,
and compared using SPSS 17.0 software (SPSS, Inc., Chicago, IL,
USA). Groups were compared using two-tailed Student's t-tests or
one-way analysis of variance with Student-Newman-Keuls post hoc
tests. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-452 is frequently downregulated in
pancreatic cancer tissues and cell lines
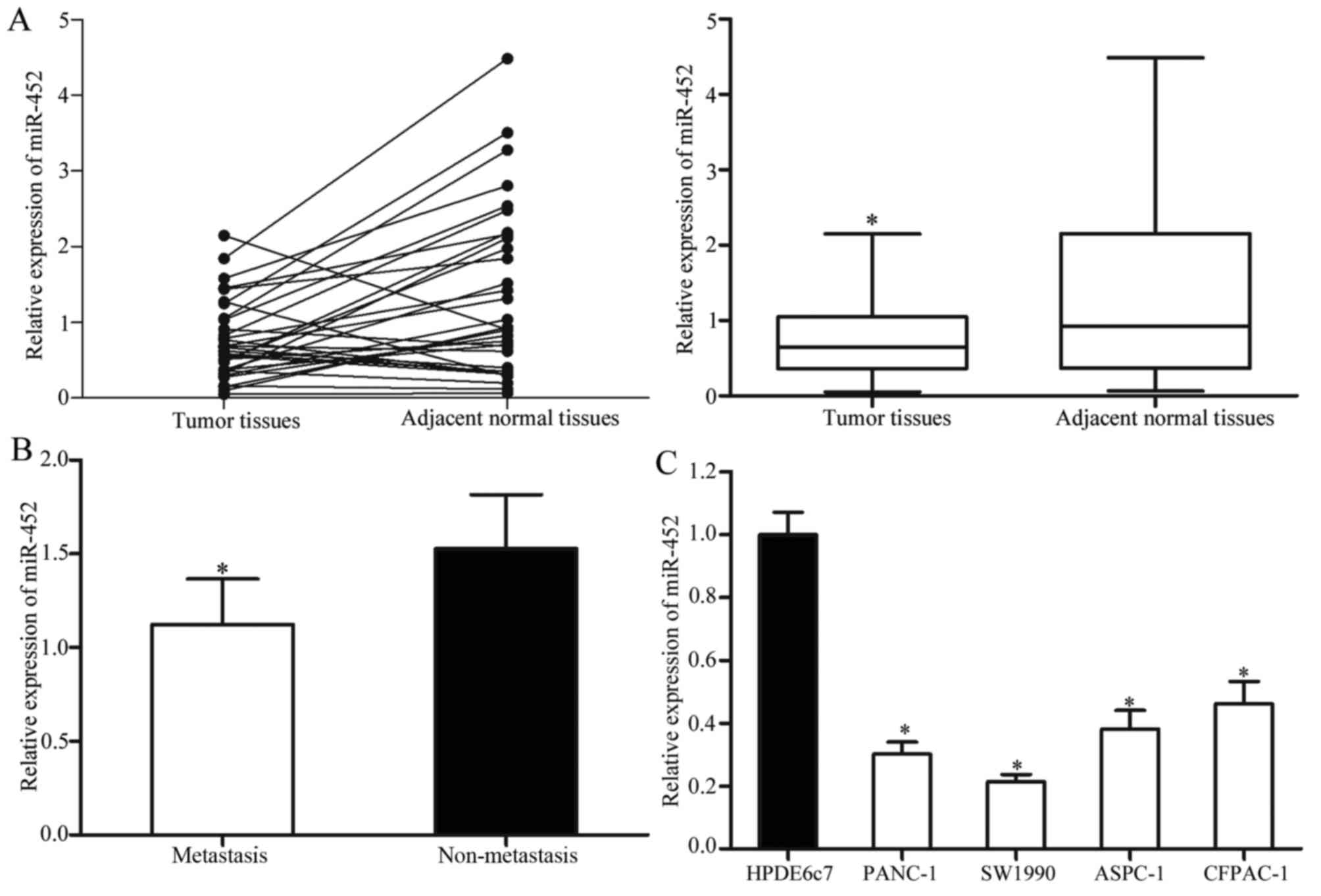
The expression levels of mature miR-452 in the
pancreatic cancer tissues and matched adjacent normal tissues were
measured using RT-qPCR. The results revealed that miR-452 was
significantly decreased in the pancreatic cancer tissues compared
with that in matched adjacent normal tissues (Fig. 1A). In addition, the levels of miR-452
in metastatic pancreatic cancer tissues were significantly lower
compared with that in pancreatic cancer without metastasis
(Fig. 1B).
miR-452 expression levels were also explored in four
pancreatic cancer cell lines (PANC-1, SW1990, ASPC-1 and CFPAC-1)
and the human normal pancreatic cell line (HPDE6c7). Lower
expression levels of miR-452 were observed in all four pancreatic
cancer cell lines compared with that in the HPDE6c7 cell line
(Fig. 1C). Among these cell lines,
PANC-1 and SW1990 cells were selected for subsequent functional
studies as they expressed relatively lower levels of miR-452.
Overexpression of miR-452 inhibits
pancreatic cancer migration and invasion
Since miR-452 was downregulated in patients with
metastatic pancreatic cancer, whether overexpression of miR-452
decreased migration and invasion capacity of pancreatic cancer was
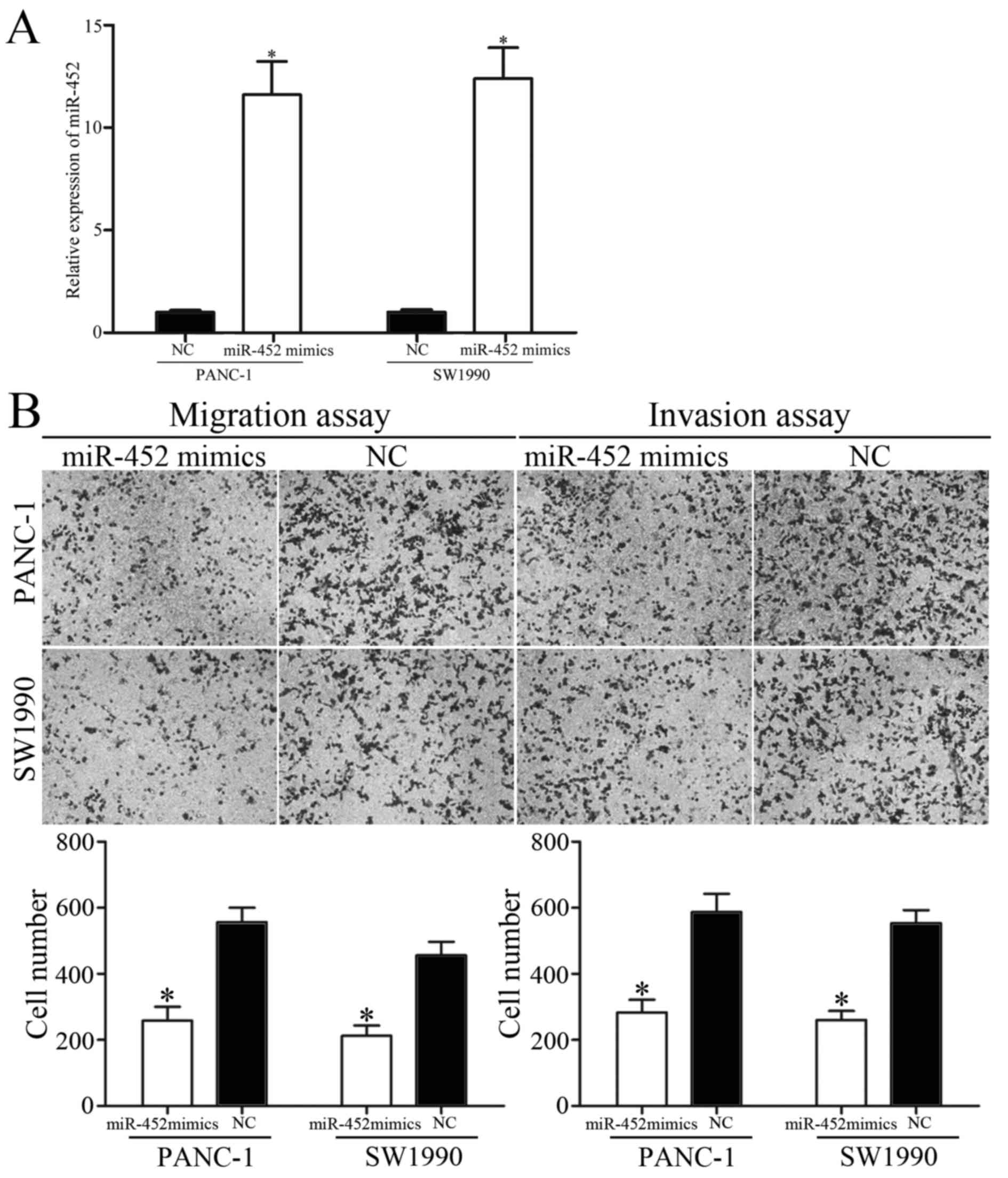
subsequently explored. PANC-1 and SW1990 cells were transfected
with miR-452 mimics or NC. The ectopic expression efficiency of
miR-452 was validated by RT-qPCR (Fig.
2A).
To clarify the effect of miR-452 on tumor
metastasis, migration and invasion assays were performed with
Transwell chambers. The results revealed that the migratory and
invasive abilities of PANC-1 and SW1990 cells were suppressed by
miR-452 overexpression (Fig. 2B).
Collectively, these results indicated that miR-452 functioned as a
tumor suppressor and contributed to suppression of metastasis of
pancreatic cancer.
BMI1 is a direct target gene of
miR-452 in pancreatic cancer
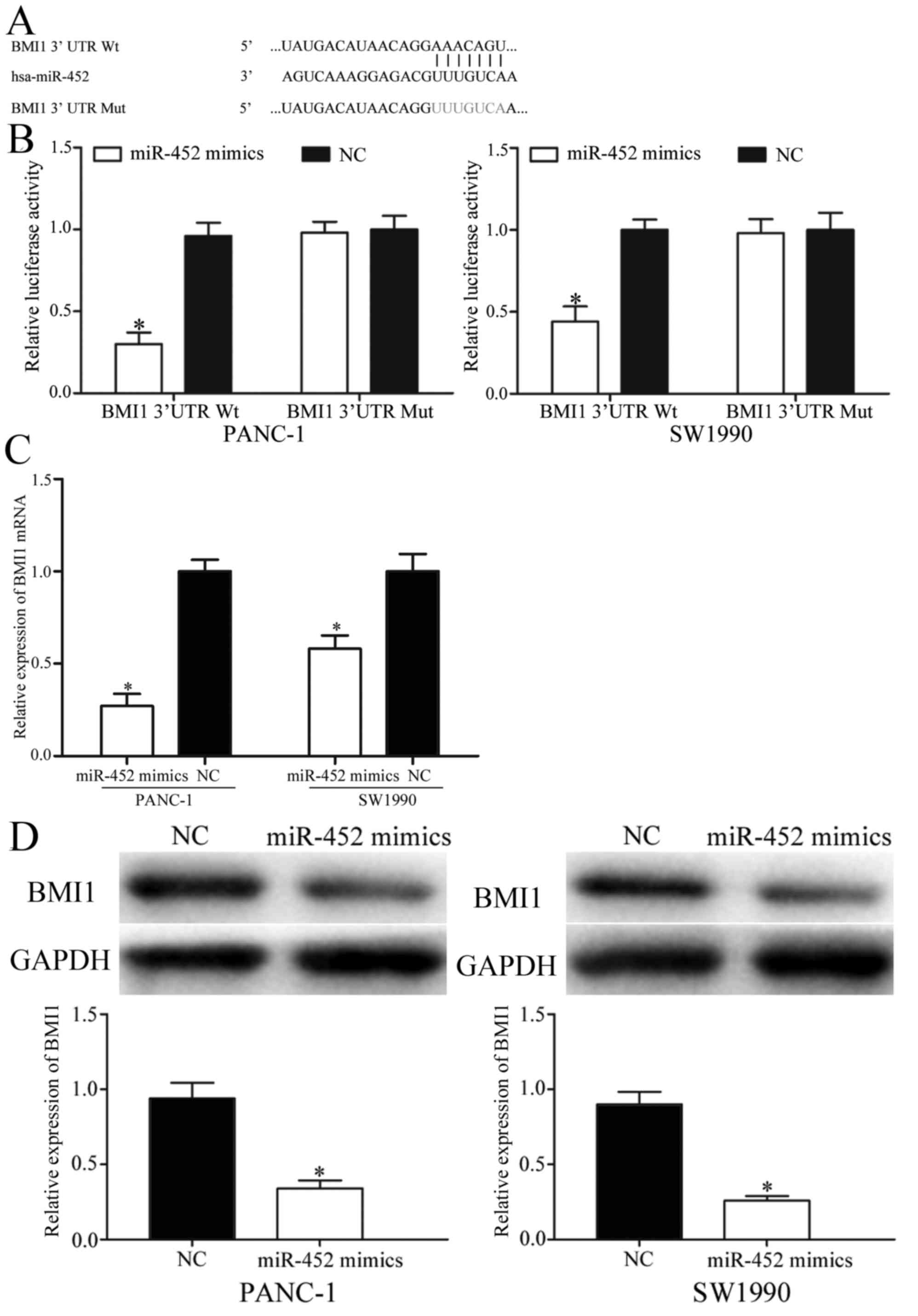
To further elucidate the molecular mechanisms
underlying the inhibitory functions of miR-452 in pancreatic
cancer, TargetScan was applied to identify predicated targets. As
presented in Fig. 3A, BMI1 contained
the predicted binding sites for miR-452. Dual-luciferase reporter
assays were then performed to investigate whether miR-452 directly
targeted the 3′UTR of BMI1. The results revealed that
co-transfection of miR-452 significantly decreased the activities
of the firefly luciferase reporter with BMI1-3′UTR Wt, whereas the
inhibition effect was abolished when the predicted 3′UTR binding
sites of BMI1 were mutated (Fig.
3B).
Furthermore, RT-qPCR and western blot analysis were
adopted to determine the regulatory effects of miR-452 in BMI1
expression. The results indicated that overexpression of miR-452
markedly suppressed BMI1 mRNA and protein levels in PANC-1 and
SW1990 cells (Fig. 3C and D). Taken
together, BMI1 was a direct target gene of miR-452 in pancreatic
cancer.
Overexpression of miR-452 inhibits
migration and invasion of pancreatic cancer cells via knockdown of
BMI1
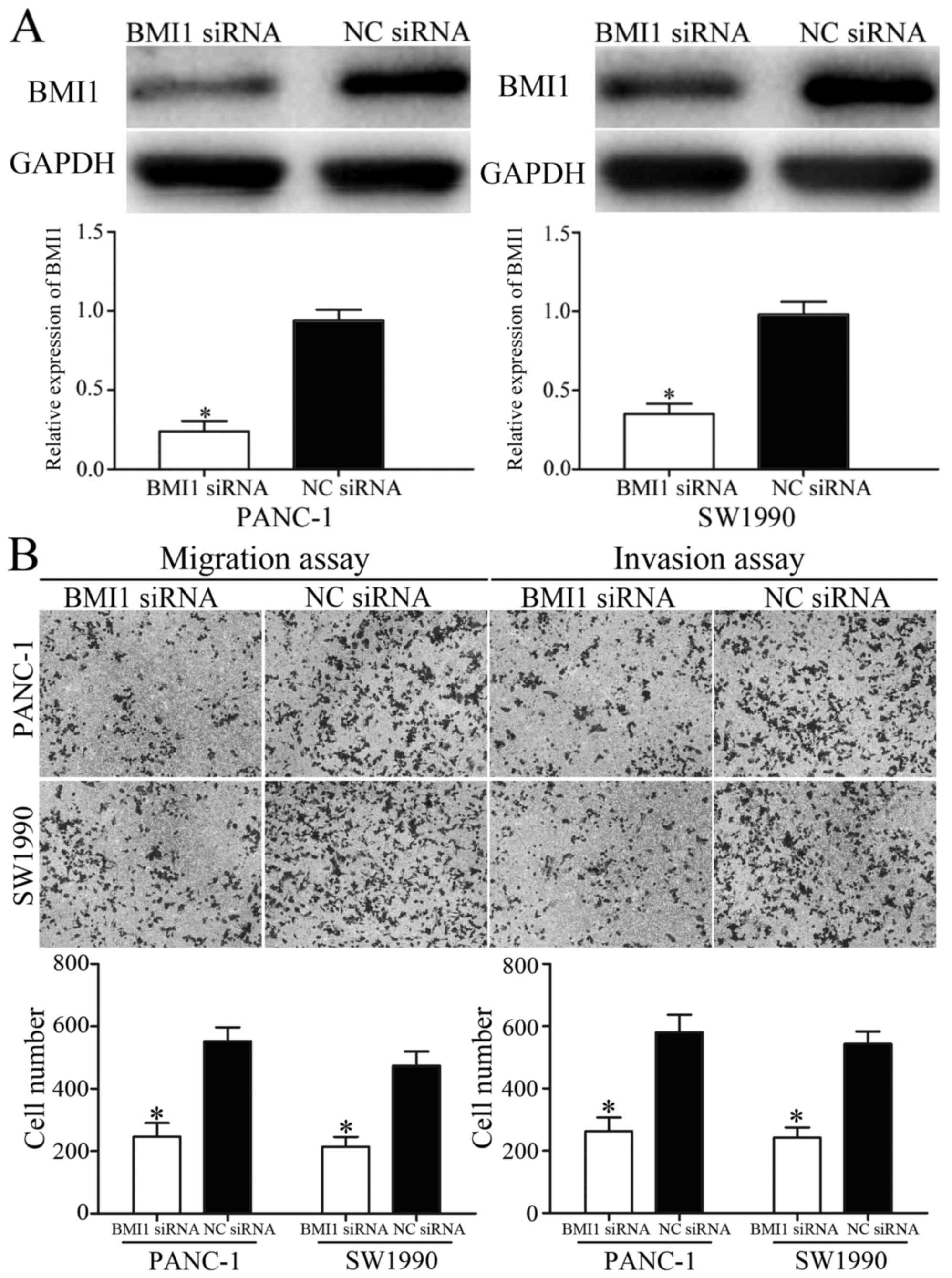
BMI1 was verified as a direct target gene of miR-452
in pancreatic cancer. To explore whether the inhibition effects of
miR-452 on migration and invasion of pancreatic cancer was
achievable by knockdown of BMI1, BMI1 siRNA or NC siRNA were
transfected into PANC-1 and SW1990 cells. The transfection
efficiency was measured using western blot analysis (Fig. 4A). The effect of BMI1 siRNA in
metastasis of pancreatic cancer was also measured. The migration
and invasion assays indicated that BMI1 siRNA inhibited migration
and invasion capacity of PANC-1 and SW1990 cells compared with that
in NC siRNA groups (Fig. 4B). These
results suggested that overexpression of miR-452 inhibited
migration and invasion of pancreatic cancer, at least partially by
knockdown of BMI1 expression.
Discussion
Pancreatic cancer is an aggressive tumor
characterized by early and aggressive metastasis, as well as
dissemination of pancreatic cancer (7). Successful management and therapy of
patients with pancreatic cancer remains one of the major challenges
in clinical oncology. It is possible to cure patients with early
stage pancreatic cancer with surgical operation (20). However, the majority of patients are
diagnosed at advanced stages and surgery is not possible (21). Therefore, there is an urgent
requirement for novel therapeutic treatments for patients with
pancreatic cancer. A number of studies have indicated that miRNAs
contribute to the carcinogenesis and progression of numerous types
of human cancer via targeting multiple mRNAs (22–24).
Therefore, identification of specific miRNAs may provide
therapeutic implications and may be exploited to improve treatments
for patients with pancreatic cancer.
The present study is the first to investigate the
expression, biological functions and molecular mechanism of miR-452
in pancreatic cancer. It was demonstrated that miR-452 was
significantly downregulated in pancreatic cancer tissues and cell
lines. In addition, the levels of miR-452 in metastatic pancreatic
cancer tissues were significantly lower compared with those in
pancreatic cancer without metastasis. Functional studies indicated
that overexpression of miR-452 suppressed the migration and
invasion of pancreatic cancer. In addition, BMI1 was identified as
a direct target gene of miR-452 in pancreatic cancer. Thus, miR-452
acted as a tumor suppressor in pancreatic cancer and its
downregulation enhanced metastasis of pancreatic cancer.
miR-452 has been studied in numerous types of human
cancer (15,16,25). For
example, He et al (15)
reported that the expression levels of miR-452 were decreased in
non-small-cell lung cancer, and low expression levels of miR-452
were associated with advanced tumor stage and more extent of lymph
nodes metastasis. Functional studies have revealed that
under-expression of miR-452 may induce cell invasion of
non-small-cell lung cancer (15). In
addition, miR-452 was revealed to negatively regulate BMI1
expression by binding to its 3′UTR, and the enhanced cell invasion
induced by downregulated miR-452 may be eliminated by knockdown of
BMI1 (15). In breast cancer, miR-452
was downregulated in adriamycin-resistant MCF-7 cells compared with
in parental MCF-7 cells. miR-452 mediated chemosensitivity to
adriamycin in human breast cancer cells (16). Furthermore, bioinformatics analysis,
RT-qPCR and western blot analysis have demonstrated that miR-452
may be at least in part be involved in adriamycin-resistance of
breast cancer via the targeting of insulin-like growth factor-1
(16). Liu et al (25) revealed that miR-452 was significantly
downregulated in glioma tissues and cell lines, and miR-452 levels
were associated with World Health Organization grades and patient
survival. Additional studies revealed that miR-452 targeted BMI1,
lymphoid enhancer-binding factor 1 and T cell factor 4 to decrease
glioma stem-like phenotypes in vitro and inhibit glioma
carcinogenesis in vivo (21).
miR-452 is regarded as a tumor suppressor in
non-small cell lung cancer (15),
breast cancer (16) and glioma
(25). However, in hepatocellular
carcinoma, miR-452 was markedly upregulated in tumor tissues and
cell lines (18). Ectopic expression
of miR-452 enhanced cell proliferation, promoted transition from
phase G1 to S in the cell cycle and suppressed apoptosis, migration
and invasion of hepatocellular carcinoma. In addition, miR-452
decreased cyclin-dependent kinase inhibitor 1B (CDKN1B) mRNA and
protein expression by directly targeting the 3′UTR of CDKN1B; the
knockdown of CDKN1B resembled the phenotype resulting from
overexpression of miR-452 expression (18). miR-452 was also demonstrated to be an
oncogene in esophageal cancer (26)
and urothelial carcinoma (27). These
conflicting studies indicated that the expression and functions of
miR-452 in human cancers have tissue specificity. It may be
explained by distinct context of various microenvironments, and the
imperfect complementarity of the interactions between miRNAs and
target mRNAs (28). In the present
study, miR-452 was demonstrated to act as a tumor suppressor in
pancreatic cancer by inhibiting cell migration and invasion. The
present study expanded the expression and functions of miR-452 in
human cancers.
Identifying the target genes of miRNA is essential
for understanding its functions in carcinogenesis and progression.
In the present study, BMI1 was revealed to be the direct target
gene of miR-452 in pancreatic cancer. BMI1, a member of the
Polycomb group genes, was first demonstrated as an oncogene in
murine lymphoma (29). Previous
studies have suggested that BMI1 is highly expressed in numerous
human cancers, including lung cancer (30), prostate cancer (31) and colorectal cancer (32). Additional studies have demonstrated
that BMI1 serves important functions in a number of biological
processes, including the cell cycle, apoptosis, senescence and
proliferation (33–35). In pancreatic cancer, BMI1 is
upregulated and expression levels of BMI1 are associated with
proliferation, survival and poor prognosis of patients with
pancreatic cancer (36). Furthermore,
BMI1 functioned as an oncogene in pancreatic cancer by promoting
chemoresistance, invasion and tumorigenesis of pancreatic cancer
(37). The results of the present
study were in accordance with the aforementioned results. The
present study revealed that knockdown of BMI1 suppressed migration
and invasion of pancreatic cancer cells, which is similar to the
effects of miR-452 in pancreatic cancer.
In summary, the present study demonstrated that
miR-452 was downregulated in pancreatic cancer tissues,
particularly in metastatic tumors and pancreatic cancer cell lines.
Ectopic expression of miR452 may suppress migration and invasion of
pancreatic cancer cells through directly targeting BMI1. These
results indicated that miR-452 may be useful as a novel potential
therapeutic treatment for pancreatic cancer to block
metastasis.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moir J, White SA, French JJ, Littler P and
Manas DM: Systematic review of irreversible electroporation in the
treatment of advanced pancreatic cancer. Eur J Surg Oncol.
40:1598–1604. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burkey MD, Feirman S, Wang H, Choudhury
SR, Grover S and Johnston FM: The association between smokeless
tobacco use and pancreatic adenocarcinoma: A systematic review.
Cancer Epidemiol. 38:647–653. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hou BH, Jian ZX, Cui P, Li SJ, Tian RQ and
Ou JR: miR-216a may inhibit pancreatic tumor growth by targeting
JAK2. FEBS Lett. 589:2224–2232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang C and Xie K: Analysis of the
potential for pancreatic cancer metastasis in vitro and in vivo.
Methods Mol Biol. 980:301–319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Su D, Yamaguchi K and Tanaka M: The
characteristics of disseminated tumor cells in pancreatic cancer: A
black box needs to be explored. Pancreatology. 5:316–324. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qin Y, Dang X, Li W and Ma Q: miR-133a
functions as a tumor suppressor and directly targets FSCN1 in
pancreatic cancer. Oncol Res. 21:353–363. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu Y, Ou Y, Wu K, Chen Y and Sun W:
miR-143 inhibits the metastasis of pancreatic cancer and an
associated signaling pathway. Tumour Biol. 33:1863–1870. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tréhoux S, Lahdaoui F, Delpu Y, Renaud F,
Leteurtre E, Torrisani J, Jonckheer N and Van Seuningen I:
Micro-RNAs miR-29a and miR-330-5p function as tumor suppressors by
targeting the MUC1 mucin in pancreatic cancer cells. Biochim
Biophys Acta. 1853:2392–2403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guled M and Knuutila S: MicroRNAs and
cancer. Duodecim. 129:1661–1669. 2013.(In Finnish). PubMed/NCBI
|
|
12
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ryan BM, Robles AI and Harris CC: Genetic
variation in microRNA networks: The implications for cancer
research. Nat Rev Cancer. 10:389–402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cimmino A, Calin GA, Fabbri M, Iorio MV,
Ferracin M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M, et
al: miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl
Acad Sci USA. 102:13944–13949. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He Z, Xia Y, Pan C, Ma T, Liu B, Wang J,
Chen L and Chen Y: Up-regulation of miR-452 inhibits metastasis of
non-small cell lung cancer by regulating BMI1. Cell Physiol
Biochem. 37:387–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu Q, Gong JP, Li J, Zhong SL, Chen WX,
Zhang JY, Ma TF, Ji H, Lv MM, Zhao JH and Tang JH: Down-regulation
of miRNA-452 is associated with adriamycin-resistance in breast
cancer cells. Asian Pac J Cancer Prev. 15:5137–5142. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Puerta-Gil P, Garcia-Baquero R, Jia AY,
Ocaña S, Alvarez-Múgica M, Alvarez-Ossorio JL, Cordon-Cardo C, Cava
F and Sánchez-Carbayo M: miR-143, miR-222, and miR-452 are useful
as tumor stratification and noninvasive diagnostic biomarkers for
bladder cancer. Am J Pathol. 180:1808–1815. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng Q, Sheng Q, Jiang C, Shu J, Chen J,
Nie Z, Lv Z and Zhang Y: MicroRNA-452 promotes tumorigenesis in
hepatocellular carcinoma by targeting cyclin-dependent kinase
inhibitor 1B. Mol Cell Biochem. 389:187–195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Link KH, Leder G, Formentini A, Fortnagel
G, Kornmann M, Schatz M and Beger HG: Surgery and multimodal
treatments in pancreatic cancer-a review on the basis of future
multimodal treatment concepts. Gan To Kagaku Ryoho. 26:10–40.
1999.PubMed/NCBI
|
|
21
|
Vychytilova-Faltejskova P, Kiss I, Klusova
S, Hlavsa J, Prochazka V, Kala Z, Mazanec J, Hausnerova J, Kren L,
Hermanova M, et al: miR-21, miR-34a, miR-198 and miR-217 as
diagnostic and prognostic biomarkers for chronic pancreatitis and
pancreatic ductal adenocarcinoma. Diagn Pathol. 10:382015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osada H and Takahashi T: MicroRNAs in
biological processes and carcinogenesis. Carcinogenesis. 28:2–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yonemori K, Kurahara H, Maemura K and
Natsugoe S: MicroRNA in pancreatic cancer. J Hum Genet. 62:33–40.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu M, Xu Z, Wang K, Wang N and Li Y:
microRNA and gene networks in human pancreatic cancer. Oncol Lett.
6:1133–1139. 2013.PubMed/NCBI
|
|
25
|
Liu L, Chen K, Wu J, Shi L, Hu B, Cheng S,
Li M and Song L: Downregulation of miR-452 promotes stem-like
traits and tumorigenicity of gliomas. Clin Cancer Res.
19:3429–3438. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu SG, Qin XG, Zhao BS, Qi B, Yao WJ,
Wang TY, Li HC and Wu XN: Differential expression of miRNAs in
esophageal cancer tissue. Oncol Lett. 5:1639–1642. 2013.PubMed/NCBI
|
|
27
|
Veerla S, Lindgren D, Kvist A, Frigyesi A,
Staaf J, Persson H, Liedberg F, Chebil G, Gudjonsson S, Borg A, et
al: miRNA expression in urothelial carcinomas: Important roles of
miR-10a, miR-222, miR-125b, miR-7 and miR-452 for tumor stage and
metastasis and frequent homozygous losses of miR-31. Int J Cancer.
124:2236–2242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu Z, Ni L, Chen D, Zhang Q, Su Z, Wang Y,
Yu W, Wu X, Ye J, Yang S, et al: Identification of miR-7 as an
oncogene in renal cell carcinoma. J Mol Histol. 44:669–677. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van Lohuizen M, Verbeek S, Scheijen B,
Wientjens E, van der Gulden H and Berns A: Identification of
cooperating oncogenes in E mu-myc transgenic mice by provirus
tagging. Cell. 65:737–752. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiong D, Ye Y, Fu Y and Wang J, Kuang B,
Wang H, Wang X, Zu L, Xiao G, Hao M and Wang J: Bmi-1 expression
modulates non-small cell lung cancer progression. Cancer Biol Ther.
16:756–763. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wolters T, Vissers KJ, Bangma CH, Schröder
FH and van Leenders GJ: The value of EZH2, p27(kip1), BMI-1 and
MIB-1 on biopsy specimens with low-risk prostate cancer in
selecting men with significant prostate cancer at prostatectomy.
BJU Int. 106:280–286. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang X, Yang X, Zhang Y, Liu X, Zheng G,
Yang Y, Wang L, Du L and Wang C: Direct serum assay for cell-free
bmi-1 mRNA and its potential diagnostic and prognostic value for
colorectal cancer. Clin Cancer Res. 21:1225–1233. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park IK, Morrison SJ and Clarke MF: Bmi1,
stem cells, and senescence regulation. J Clin Invest. 113:175–179.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee K, Adhikary G, Balasubramanian S,
Gopalakrishnan R, McCormick T, Dimri GP, Eckert RL and Rorke EA:
Expression of Bmi-1 in epidermis enhances cell survival by altering
cell cycle regulatory protein expression and inhibiting apoptosis.
J Invest Dermatol. 128:9–17. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pasini D, Bracken AP and Helin K: Polycomb
group proteins in cell cycle progression and cancer. Cell Cycle.
3:396–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Song W, Tao K, Li H, Jin C, Song Z, Li J,
Shi H, Li X, Dang Z and Dou K: Bmi-1 is related to proliferation,
survival and poor prognosis in pancreatic cancer. Cancer Sci.
101:1754–1760. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin T, Wei H, Leng Z, Yang Z, Gou S, Wu H,
Zhao G, Hu X and Wang C: Bmi-1 promotes the chemoresistance,
invasion and tumorigenesis of pancreatic cancer cells.
Chemotherapy. 57:488–496. 2011. View Article : Google Scholar : PubMed/NCBI
|