Introduction
Glioblastoma multiforme (GBM) is the most common
malignant primary brain tumors in humans (1). A notable characteristic of GBM is the
ability to invade normal, healthy brain tissue, which creates new
malignant foci and results in low total resection and a high
recurrence rate, which is problematic in terms of treatment. Due to
the high proliferation rate and malignant and invasive
characteristics, GBM one of the most aggressive brain tumors, and
under the current standard of care the median survival time from
diagnosis is 15 months (2). One way
to solve this problem would be to effectively inhibit signaling
pathways that control cell migration and invasion.
Luteolin (3′,4′,5,7-tetrahydroxyflavone) is a common
dietary flavonoid, which is present at high concentrations in food
plants and vegetables (3). Flavonoids
are well-known to have effects on xenobiotic and carcinogen
metabolism (4). Previous research has
demonstrated that plants rich in luteolin have a wide range of
antioxidant, anti-inflammatory and anticancer effects (5). Luteolin demonstrates strong anticancer
activity against a series of solid tumors, including colonic HT-29,
HCT116, hepatic HepG2 and pulmonic A549 (6–9). Although
the preclinical anticancer efficacy of luteolin has been
demonstrated in various cancer models, its effect on glioblastoma
cells has rarely been studied. As a flavonoid, previous studies
have revealed that luteolin is able to cross the blood-brain
barrier (BBB) (10,11). Furthermore, luteolin is potentially
beneficial for the central nervous system (CNS), as it is able to
decrease inflammation and axonal damage by preventing monocyte
migration across the BBB (12). In
the present study, the effects of luteolin on the migration of
human glioblastoma cell lines was investigated, and the potential
underlying mechanisms were investigated.
Materials and methods
Cell culture procedures
Human glioblastoma U251MG and U87MG cell lines were
obtained from the American Type Culture Collection (Manassas, VA,
USA) and cultured in Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA). The cells were incubated
at 37°C in a humidified atmosphere of 95% air and 5%
CO2.
Reagents and antibodies
Luteolin and phalloidin were purchased from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). Insulin-like growth
factor-1 (IGF-1) was purchased from PeproTech, Inc. (Rocky Hill,
NJ, USA) Anti-phosphorylated (p-)insulin-like growth factor-1
receptor (IGF-1R) (cat. no. sc-81499; 1:500) was purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). Anti-matrix
metalloproteinase (MMP)-2 (cat. no. ab-7033; 1:1,000), anti-MMP-9
(cat. no. ab-76003; 1:1,000), anti-tissue inhibitor of
metalloproteinase (TIMP)-1 (cat. no. ab-109125; 1:1,000) and
anti-TIMP-2 (cat. no. ab-157386; 1:1,000) were purchased from Abcam
(Cambridge, UK). Anti-E-cadherin, anti-N-cadherin, anti-vimentin,
anti-β-catenin, anti-vimentin (EMT kit; cat. no. cst-9782;
1:1,000), anti-p-protein kinase B (AKT) (cst-4060; 1:1,000),
anti-AKT (cat. no. cst-9272; 1:1,000), anti-p-mammalian target of
rapamycin (mTOR) (cat. no. cst-2971; 1:1,000), anti-mTOR (cat. no.
cst-2983; 1:1,000), anti-β-actin (cat. no. cst-4970; 1:1,000), and
horseradish peroxidase (HRP)-conjugated goat anti-rabbit
immunoglobulin G (IgG; heavy and light chain; cat. no. cst-7074;
1:5,000) secondary antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The goat anti-mouse IgG-HRP
were purchased from BioworldTechnology (cat. no. BS12478; 1:10,000;
St. Louis Park, MN, USA).
Cell proliferation assay
Cell proliferation was assessed by Cell Counting
Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Briefly, U251MG and U87MG cells were seeded into
96-well plates at a density of 2×103 cells per well, and
cultured for at least 24 h to adhere. Then, the cells were treated
with different concentrations of luteolin (0, 5, 10, 20, 40, 80 and
100 µM). Following treatment for 24 h at 37°C, 10 µl CCK-8 reagent
was added to the cells followed by incubation for 2 h at 37°C. Then
the absorbance was measured at a wavelength of 450 nm using a
Bio-Rad ELISA microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The proliferation rate was calculated as
follows: [1-optical density (OD) 450 values of treated groups/OD450
values of control group] × 100%.
Scratch-induced migration assay
This was performed as previously described by
Etienne-Manneville (13). U251MG and
U87MG cells were seeded on collagen-covered 6-well plates at a
density of 0.3×106 (U251MG) and 0.4×106
(U87MG) per plate in DMEM containing 10% FBS. Following incubation
for 24 h, the medium was replaced by DMEM containing 0.5% FBS, and
the cells were treated for 24 h with 0, 5, 10 or 20 µM luteolin at
37°C. In each plate, three areas were scratched, creating three
gaps of similar widths with a 200 µl standard pipette tip. At the
indicated time points (0, 6, 12 and 24 h), phase-contrast images of
the plates were obtained using a ZEISS inverted microscope (Zeiss
GmbH, Jena, Germany; magnification, ×4). The widths of gaps treated
with the different concentrations of luteolin and at different time
points were measured by Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc., Rockville, MD, USA). The widths of the three
scratches on each plate were averaged to obtain the mean gap width
at a given time. Statistical analysis disclosed the mean gap width
(in arbitrary units) of luteolin-treated cells relative to the
control (DMSO) at different time points (mean ± standard error of
the mean; n=3).
Western blot analysis
U251MG and U87MG cells were plated at a density of
0.3×106 (U251MG) and 0.4×106 (U87MG) cells in
6-well plates or 35 mm dishes, respectively, and were allowed to
grow overnight in DMEM containing 10% FBS. The medium was then
replaced by medium without FBS, and the cells were treated for 24 h
with different concentrations of luteolin (0, 5, 10 or 20 µM) with
or without 100 ng/ml IGF-1. The cells were then lysed with
solubilization buffer [50 mmol/l Tris-HCl (pH 7.6), 20 mmol/l
MgCl2, 200 mmol/l NaCl, 0.5% NP40, 1 mmol/l DTT and
protease inhibitors] on ice for 10 min, and the lysate (20–100 µg)
was subjected to 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred to polyvinylidene fluoride
membranes (Merck KGaA). Following blocking with 5% nonfat dried
milk for 1 h at room temperature, the membrane was incubated with
the appropriate primary antibodies overnight at 4°C. Then, the
immunoreactive bands were visualized using an Enhanced
Chemiluminescence kit (EMD Millipore, Billerica, MA, USA) using
horseradish peroxidase-conjugated IgG secondary antibodies at room
temperature for 2 h.
Cytoskeleton staining
U251MG cells were seeded on glass coverslips in DMEM
containing 10% FBS, placed in 6-well plates at a density of
1×105 cells per well, for 24 h. The medium was then
replaced by DMEM containing 0.5% FBS, and 20 µM luteolin was added.
Following further incubation for 24 h at 37°C, the cells were fixed
with 4% paraformaldehyde for 30 min at room temperature, and were
stained using rhodamine-conjugated phalloidin (1:1,000;
Sigma-Aldrich; Merck KGaA) for 30 min at room temperature. Slides
were washed with PBS, mounted, imaged with 20 fields and counted
using a ZEISS fluorescence microscope (Zeiss GmbH) (magnification,
×40). Cells with stress fibers (mean ± standard error of the mean;
n=3) were expressed as a percentage of 100 cells counted from each
slide. Cell area (mean ± standard error of the mean; n=3) was
measured using ZEN2 software (Zeiss GmbH).
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM SPSS, Armonk, NY, USA). Data are presented as the
mean ± standard deviation and evaluated using one-way analysis of
variance followed with Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Luteolin inhibits the proliferation of
glioblastoma cells
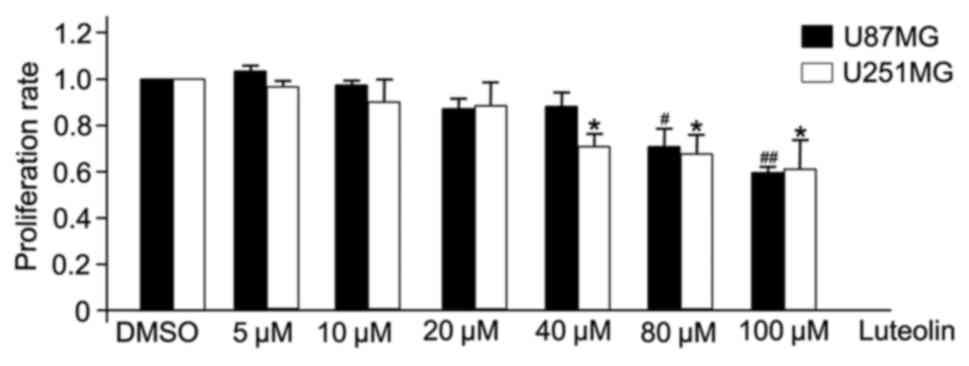
Luteolin demonstrated a clear anti-proliferative
effect on U251MG and U87MG cells (Fig.
1). U251MG and U87MG glioblastoma cells were treated with
various concentrations of luteolin (0, 5, 10, 20, 40, 80 and 100
µM) for 24 h, and the effect was examined by CCK-8 assay. At
concentrations >40 µM, luteolin significantly inhibited the
proliferation of U251MG cells, and significantly inhibited the
proliferation of U87MG cells at 80 µM. However, at concentrations
<20 µM, there was no apparent anti-proliferative effect.
Therefore, to exclude the cytotoxic effects of excess luteolin, the
following experiments selected a concentration range of luteolin
<40 µM to determine the associated effects on glioblastoma
cells.
Luteolin disrupts the migration of
glioblastoma cells
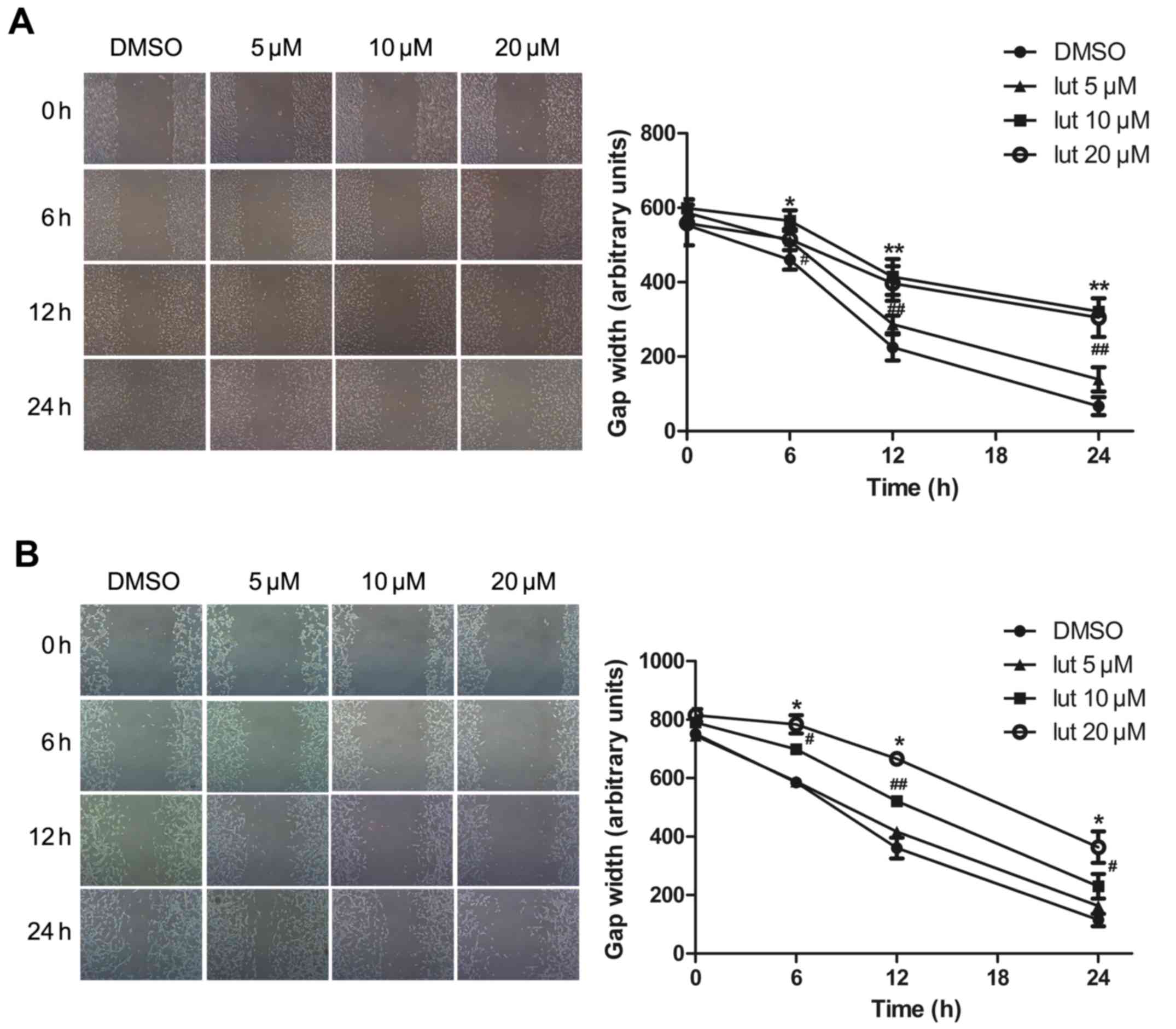
Next, the effect of luteolin on the motility of
glioblastoma was assessed. This was performed using a
scratch-induced migration assay in which the width of the gap
formed by a scratch was monitored at different times following the
infliction of the wound (13). The
U251MG and U87MG cells were pretreated with luteolin (5, 10 or 20
µM) or with DMSO (as a control) for 24 h, and maintained in DMEM
containing 0.5% FBS to block cell proliferation, which would
otherwise account for gap closure. The gap width was then monitored
at the indicated time points of 0, 6, 12 and 24 h. Luteolin
treatment significantly decreased the migration ability of
glioblastoma cells in a time- and concentration-dependent manner
(Fig. 2). There was no difference in
width length at the lowest concentration of 5 µM, whilst at
concentrations of 10 and 20 µM the gap closure was attenuated. It
appeared to move at a slower pace compared with the control cells,
closing a smaller portion of the wound gap. Statistical analysis of
the results indicated that luteolin caused a significant decrease
in U251MG and U87MG cell migration.
Effects of luteolin on the expression
of MMP-2, MMP-9, TIMP-1 and TIMP-2 in glioblastoma cells
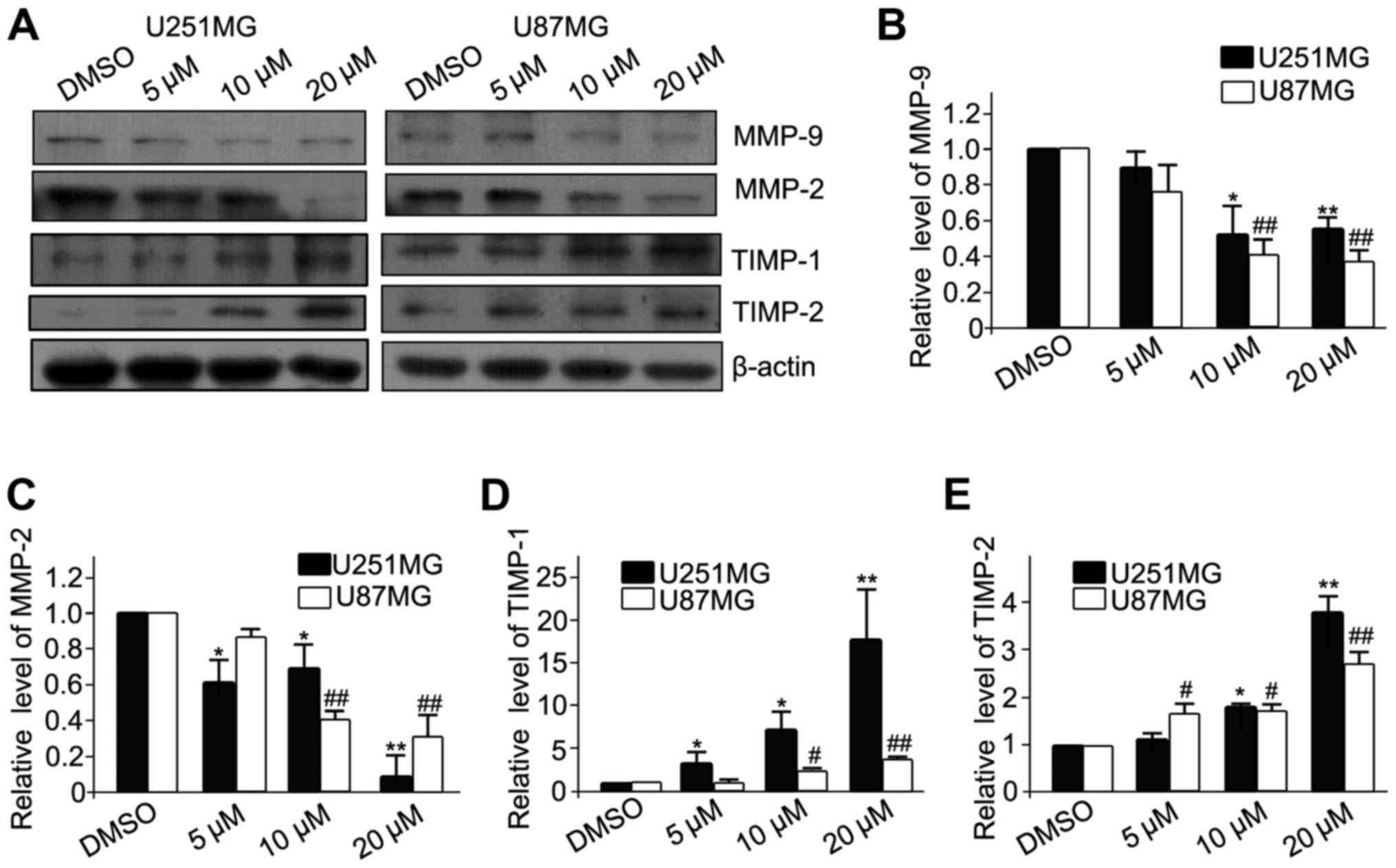
MMPs are gelatinizes which are capable of degrading
the extracelluar matrix (ECM) and facilitating the migration and
invasion of cancer cells. In contrast, TIMPs are the endogenous
inhibitors of MMPs, and prevent the breakdown of the ECM.
Therefore, the protein expression of MMPs (MMP-2 and MMP-9) and
TIMPs (TIMP-1 and TIMP-2) was analyzed (Fig. 3A). The results revealed that luteolin
significantly decreased the expression of MMP-2 and MMP-9 (Fig. 3B and C, respectively), and
significantly increased the expression of TIMP-1 and TIMP-2
(Fig. 3D and E, respectively). These
results demonstrated that luteolin inhibited the migration of
glioblastoma cells partially via downregulation of MMPs and
upregulation of TIMPs.
Luteolin prevents
epithelial-mesenchymal transition (EMT) progression in glioblastoma
cells
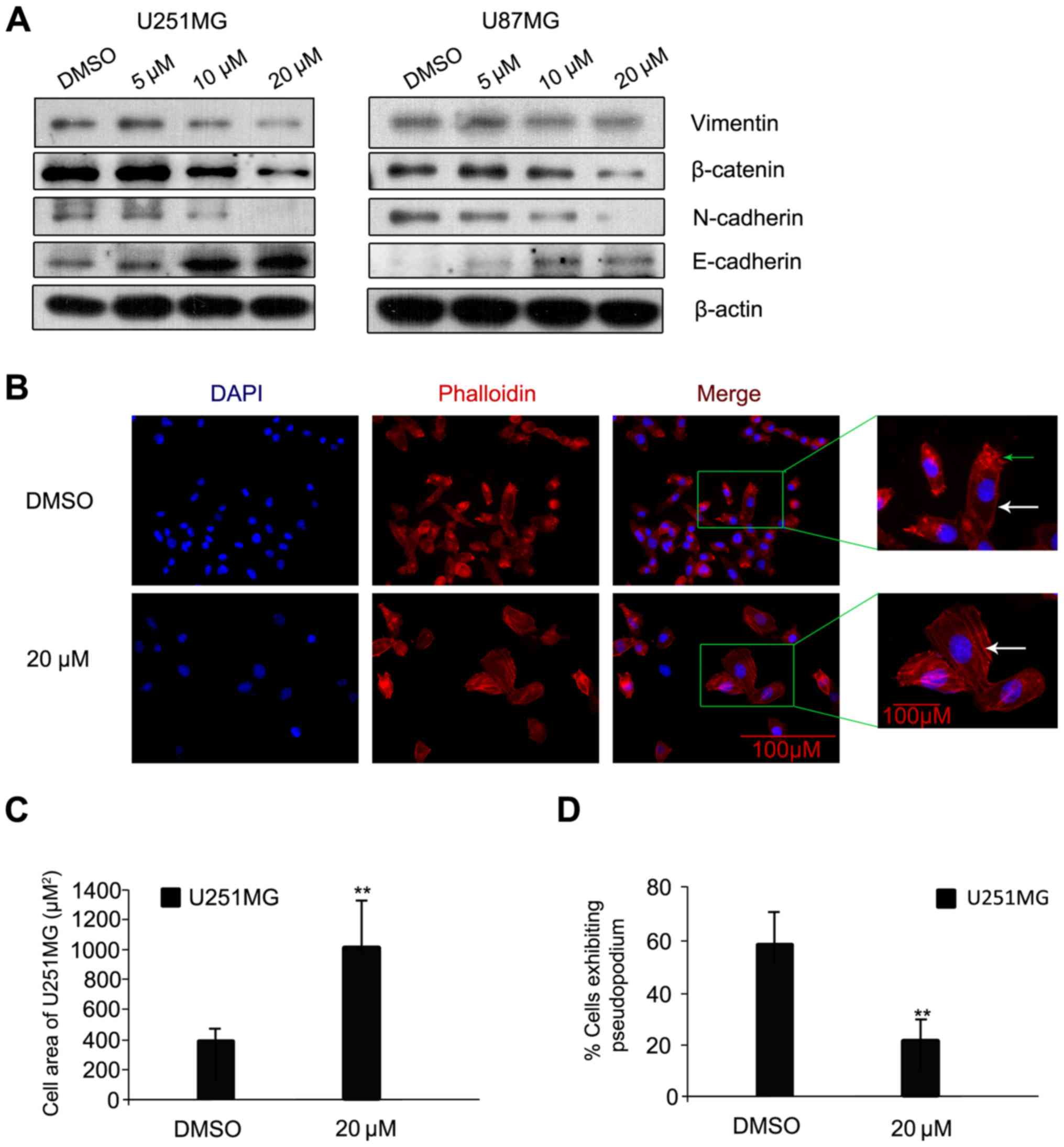
The process of cancer cell invasion is enabled by
EMT, which is the initiator of the metastatic cascade (14). Changes in protein expression levels of
E-cadherin, N-cadherin, β-catenin and Vimentin, all EMT-associated
proteins, were assessed. Following luteolin treatment for 24 h, the
protein levels of N-cadherin, β-catenin and Vimentin were visibly
decreased; while the protein level of E-cadherin increased
(Fig. 4A). A previous study indicated
that N-cadherin is upregulated in cancer cells while E-cadherin is
downregulated; known as the ‘cadherin switch’ (15). N-cadherin interacts with the
fibroblast growth factor receptor, leading to overexpression of
MMP-9 and cellular invasion (16).
The effects on luteolin on U251MG cell morphology and cytoskeleton
organization were then examined using rhodamine-labeled phalloidin,
which interacts with polymeric F-actin. The cells were treated with
luteolin (20 µM) or with DMSO as a negative control for 24 h under
the same conditions as those used in the scratch-induced migration
assay, and then imaged. Control U251MG glioblastoma cells were
characterized by small cell bodies and long extensions, while the
luteolin-treated cells exhibited flat morphology and were visibly
larger (Fig. 4B). In addition, the
glioblastoma cells had visibly fewer pseudopodia following luteolin
treatment compared with the control (Fig.
4B; as indicated by the green arrow). The majority of the
polymeric actin in the control cells appeared to be concentrated in
membrane ruffles, while in luteolin-treated cells, polymeric actin
was organized into stress fibers (Fig.
4B; as indicated by the white arrow). The cell area of U251MG
cells was significantly higher following treatment with 20 µM
luteolin (Fig. 4C) and the percentage
of U251MG cells exhibiting pseudopodia was significantly decreased
following treatment with 20 µM luteolin (Fig. 4D).
The p-IGF-1R/phosphoinositide 3-kinase
(PI3K)/AKT/mTOR signaling pathway is involved in the
luteolin-induced reduction in cell migration
The PI3K/AKT/mTOR signaling pathway is involved in
regulating the migration of cancer cells. Therefore, the effect of
luteolin on this pathway was examined. Luteolin treatment visibly
decreased the protein levels of p-AKT and p-mTOR in a
concentration-dependent manner (Fig.
5A). IGF-1R is a transmembrane heterotetramer with a
cytoplasmic tyrosine kinase domain that activates the PI3K/AKT and
RAS-rapidly accelerated fibrosarcoma-mitogen-activated protein
kinase (MAPK) signaling pathways (17). IGF-1R is overexpressed in multiple
types of cancer (18). Previous
studies have demonstrated that picropodophyllin, an inhibitor of
IGF-1R phosphorylation, inhibits the growth of human glioblastoma
cell lines and causes tumor regression not only in subcutaneous
xenografts, but also in intracerebral xenografts, along with
reduced phosphorylation of IGF-1R and AKT (19). Therefore, the protein levels of
p-IGF-1R, which was upstream of PI3K, were assessed. These also
decreased following treatment with luteolin for 24 h (Fig. 5A). To further explore the involvement
of p-IGF-1R and this pathway in the anti-migratory effect of
luteolin, IGF-1 was used to upregulate p-IGF-1R. The U251MG and
U87MG cells were serum-starved overnight and then treated for 24 h
with 20 µM luteolin prior to 1 h stimulation with 100 ng/ml IGF-1.
The results revealed that IGF-1 recovered the level of p-AKT,
p-mTOR, MMP-2, MMP-9, N-cadherin and Vimentin, which decreased
following treatment of luteolin (Fig.
5B). In addition, the protein levels of TIMP-1, TIMP-1 and
E-cadherin also decreased following treatment with IGF-1 (Fig. 5B). These results suggested that
luteolin-induced inhibition of migration in glioblastoma cells
partially occurred through the p-IGF-1R/PI3K/AKT/mTOR signaling
pathway.
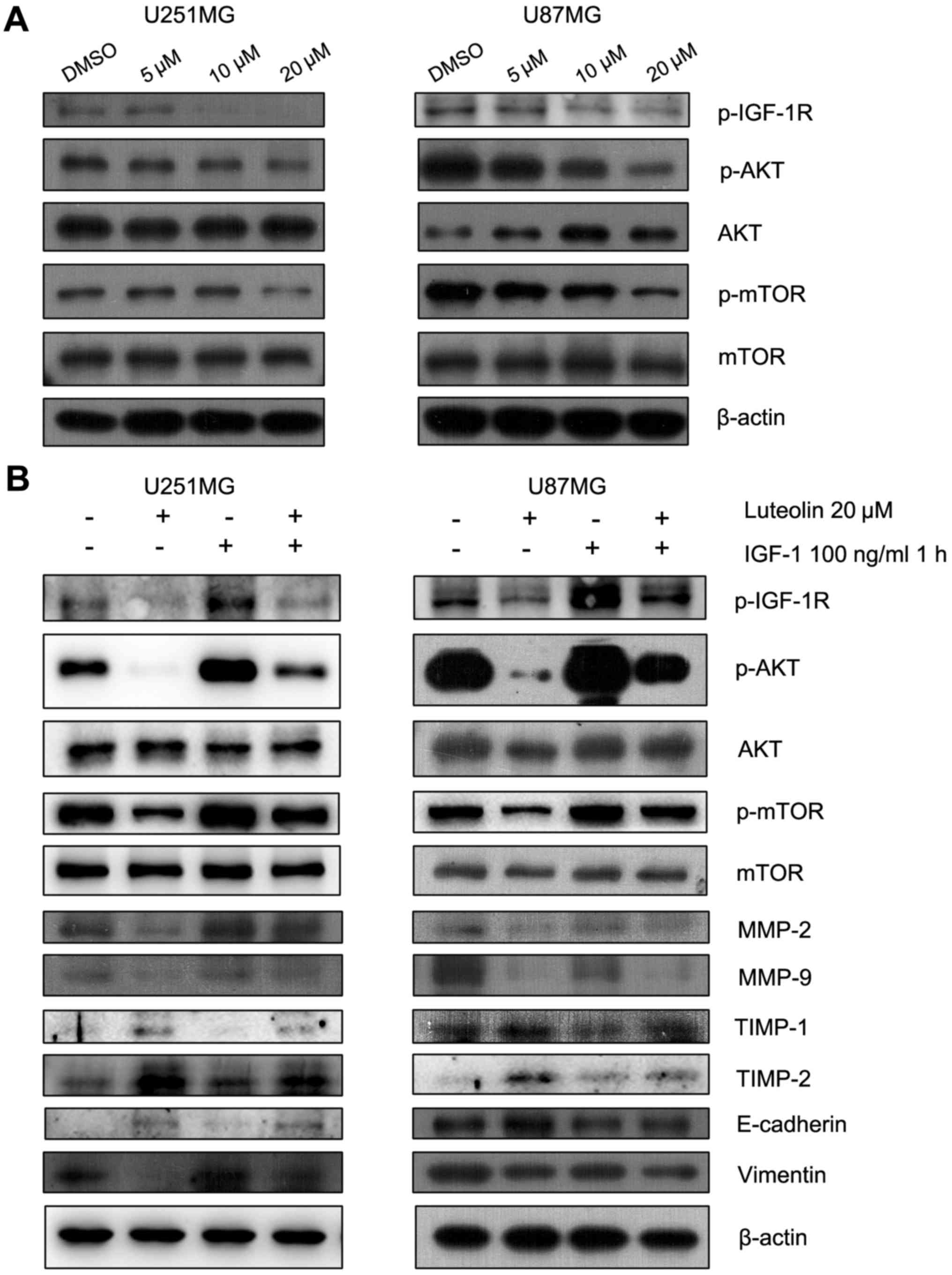 | Figure 5.The p-IGF-1R/phosphoinositide 3
kinase/AKT/mTOR signaling pathway was involved in the
anti-migration progress of luteolin. (A) Cells were exposed to
various concentrations of luteolin for 24 h and the expression of
p-IGF-1R, p-AKT, AKT, p-mTOR and mTOR was detected by western blot.
(B) U251MG and U87MG cells were serum-starved overnight and then
treated for 24 h with 20 µM luteolin prior to 1 h stimulation with
100 ng/ml IGF-1. The expression of p-IGF-1R, p-AKT, AKT, p-mTOR,
mTOR, MMP-2, MMP-9, TIMP-1, TIMP-2, E-cadherin, Vimentin and
β-actin was determined. p-, phosphorylated; IGF-1R, insulin-like
growth factor-1 receptor; AKT, protein kinase B; mTOR, mammalian
target of rapamycin; IGF-1, insulin-like growth factor-1; MMP,
matrix metalloproteinase; TIMP, tissue inhibitor of
metalloproteinase. |
Discussion
The present study explored the effect of luteolin on
the migration and EMT of human glioblastoma cells. Luteolin was
demonstrated to reduce the migration of glioblastoma cells and to
weaken the EMT process via suppression of the
p-IGF-1R/PI3K/AKT/mTOR signaling pathway. These results provided
substantial evidence for the anti-migration and anti-EMT effects of
luteolin against human glioblastoma.
The degradation of the vascular basement membrane is
required for tumor cells to invade though the basement membrane and
migrate to distant secondary sites, and is performed by type IV
collagenases known as MMPs (20,21). MMPs
are zinc-binding endopeptidases that promote cancer cell migration
and invasion via breakdown of the ECM (22). TIMPs are the endogenous inhibitors of
the MMPs (23). Therefore, the
decreased expression of MMPs and upregulation of TIMPs may provide
a potential therapeutic target to inhibit tumor migration. In the
present study, luteolin was demonstrated to decrease MMP-2 and
MMP-9 protein levels and increase TIMP-1 and TIMP-2 protein levels.
These results suggested that the anti-migratory effect of luteolin
was associated with an altered MMP/TIMP balance.
The term EMT describes a process by which stationary
epithelial cells lose their characteristic polarity, disassemble
their cell-cell junctions and become increasingly motile (24). EMT is considered to be critical event
in the process of cancer migration. Thus, the present study
analyzed the protein levels of EMT-associated factors, including
E-cadherin, N-cadherin, β-catenin and Vimentin. The protein levels
of N-cadherin, β-catenin and Vimentin decreased following treatment
with luteolin, while E-cadherin protein levels increased. The
significance of the observed morphological changes lies in the
implications for cell motility: Once the actin is organized into
stress fibers and focal adhesions are assembled, cells flatten and
become attached to the ECM (25).
Cells in motion need to assemble and disassemble actin structures
to progress, alternating between attachment to and detachment from
the ECM. Luteolin treatment induced alterations in cell morphology,
which were attributed to reorganization of the actin cytoskeleton.
These results are important when contemplating novel treatments for
human glioblastoma.
IGF-1R is a transmembrane heterotetramer with a
cytoplasmic tyrosine kinase domain that activates the PI3K/AKT and
RAS/MAPK signaling pathways. IGF-1R overexpression is a
characteristic common to multiple types of human cancer (26). IGF-1R has previously been suggested to
be involved in cerebellum tumors and in neuroblastomas (27,28).
Furthermore, the cyclolignan picropodophyllin, a specific inhibitor
of IGF-1R, inhibits the growth of human GBM cell lines along with
reduced phosphorylation of IGF-1R and AKT (29). In addition, the PI3K/AKT/mTOR
signaling pathway controls the migration of cancer cells, and
anticancer drugs inhibiting the PI3K/AKT/mTOR axis reduce cell
migration (30). The present study
demonstrated that luteolin decreased the phosphorylation of IGF-1R,
AKT and mTOR in a concentration-dependent manner. The specific
activator of IGF-1R, IGF-1, was able to recover the decreased
protein level of MMP-2, MMP-9 and Vimentin and decrease the level
of TIMP-1 and TIMP-2, suggesting that the p-IGF-1R/PI3K/AKT/mTOR
signaling pathway was associated with luteolin-mediated inhibition
of migration.
Taken together, the results of the present provided
evidence that luteolin exerted anti-migratory and anti-EMT effects
in human glioblastoma cells via inhibition of the
p-IGF-1R/PI3K/AKT/mTOR signaling pathway. These results suggest
that luteolin may be attractive therapeutic agent for the
development of future treatment protocols.
Acknowledgements
The authors would like to thank Dr Han Yanling
(Department of Neurosurgery, Jinling Hospital, School of Medicine,
Nanjing University, Nanjing, China) for the technical assistance.
The present study was supported by grants from the National Natural
Science Foundation of China (grant no. 81371357) and the China
Postdoctoral Science Foundation (grant no. 2014M562665,
2015T81136).
References
|
1
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol.
15:(Suppl 2). ii1–ii56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin Y, Shi R, Wang X and Shen HM:
Luteolin, a flavonoid with potentials for cancer prevention and
therapy. Curr Cancer Drug Targets. 8:634–646. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moon YJ, Wang X and Morris ME: Dietary
flavonoids: Effects on xenobiotic and carcinogen metabolism.
Toxicol In Vitro. 20:187–210. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chian S, Thapa R, Chi Z, Wang XJ and Tang
X: Luteolin inhibits the Nrf2 signaling pathway and tumor growth in
vivo. Biochem Biophys Res Commun. 447:602–608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chang J, Hsu Y, Kuo P, Kuo Y, Chiang L and
Lin C: Increase of Bax/Bcl-XL ratio and arrest of cell cycle by
luteolin in immortalized human hepatoma cell line. Life Sci.
76:1883–1893. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee HJ, Wang CJ, Kuo HC, Chou FP, Jean LF
and Tseng TH: Induction apoptosis of luteolin in human hepatoma
HepG2 cells involving mitochondria translocation of Bax/Bak and
activation of JNK. Toxicol Appl Pharmacol. 203:124–131. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim DY, Jeong Y, Tyner AL and Park JH:
Induction of cell cycle arrest and apoptosis in HT-29 human colon
cancer cells by the dietary compound luteolin. Am J Physiol
Gastrointest Liver Physiol. 292:G66–G75. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang X, Wang H, Fan L, Wu X, Xin A, Ren H
and Wang XJ: Luteolin inhibits Nrf2 leading to negative regulation
of the Nrf2/ARE pathway and sensitization of human lung carcinoma
A549 cells to therapeutic drugs. Free Radic Biol Med. 50:1599–1609.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Youdim KA, Qaiser MZ, Begley DJ,
Rice-Evans CA and Abbott NJ: Flavonoid permeability across an in
situ model of the blood-brain barrier. Free Radic Biol Med.
36:592–604. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jang S, Kelley KW and Johnson RW: Luteolin
reduces IL-6 production in microglia by inhibiting JNK
phosphorylation and activation of AP-1. Proc Natl Acad Sci USA.
105:7534–7539. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hendriks JJ, Alblas J, van der Pol SM, van
Tol EA, Dijkstra CD and de Vries HE: Flavonoids influence monocytic
GTPase activity and are protective in experimental allergic
encephalitis. J Exp Med. 200:1667–1672. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Etienne-Manneville S: In vitro assay of
primary astrocyte migration as a tool to study Rho GTPase function
in cell polarization. Methods Enzymol. 406:565–578. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Subramani R, Lopez-Valdez R, Arumugam A,
Nandy S, Boopalan T and Lakshmanaswamy R: Targeting insulin-like
growth factor 1 receptor inhibits pancreatic cancer growth and
metastasis. PLoS One. 9:e970162014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aigner K, Dampier B, Descovich L, Mikula
M, Sultan A, Schreiber M, Mikulits W, Brabletz T, Strand D, Obrist
P, et al: The transcription factor ZEB1 (deltaEF1) promotes tumour
cell dedifferentiation by repressing master regulators of
epithelial polarity. Oncogene. 26:6979–6988. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hazan RB, Qiao R, Keren R, Badano I and
Suyama K: Cadherin switch in tumor progression. Ann N Y Acad Sci.
1014:155–163. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baserga R, Peruzzi F and Reiss K: The
IGF-1 receptor in cancer biology. Int J Cancer. 107:873–877. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
King H, Aleksic T, Haluska P and Macaulay
VM: Can we unlock the potential of IGF-1R inhibition in cancer
therapy? Cancer Treat Rev. 40:1096–1105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yin S, Girnita A, Strömberg T, Khan Z,
Andersson S, Zheng H, Ericsson C, Axelson M, Nistér M, Larsson O,
et al: Targeting the insulin-like growth factor-1 receptor by
picropodophyllin as a treatment option for glioblastoma. Neuro
Oncol. 12:19–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim YS, Lee HA, Lim JY and Kim Y, Jung CH,
Yoo SH and Kim Y: β-Carotene inhibits neuroblastoma cell invasion
and metastasis in vitro and in vivo by decreasing level of
hypoxia-inducible factor-1α. J Nutr Biochem. 25:655–664. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hiraoka N, Allen E, Apel IJ, Gyetko MR and
Weiss SJ: Matrix metalloproteinases regulate neovascularization by
acting as pericellular fibrinolysins. Cell. 95:365–377. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu KW, Chen JC, Lai TY, Yang JS, Weng SW,
Ma YS, Lu PJ, Weng JR, Chueh FS, Wood WG and Chung JG: Gypenosides
inhibits migration and invasion of human oral cancer SAS cells
through the inhibition of matrix metalloproteinase-2 −9 and
urokinase-plasminogen by ERK1/2 and NF-kappa B signaling pathways.
Hum Exp Toxicol. 30:406–415. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brew K, Dinakarpandian D and Nagase H:
Tissue inhibitors of metalloproteinases, evolution, structure and
function. Biochim Biophys Acta. 1477:267–283. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Raftopoulou M and Hall A: Cell migration:
Rho GTPases lead the way. Dev Biol. 265:23–32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clemmons DR: Modifying IGF1 activity: An
approach to treat endocrine disorders, atherosclerosis and cancer.
Nat Rev Drug Discov. 6:821–833. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wojtalla A, Salm F, Christiansen DG,
Cremona T, Cwiek P, Shalaby T, Gross N, Grotzer MA and Arcaro A:
Novel agents targeting the IGF-1R/PI3K pathway impair cell
proliferation and survival in subsets of medulloblastoma and
neuroblastoma. PLoS One. 7:e471092012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang JY, Del Valle L, Gordon J, Rubini M,
Romano G, Croul S, Peruzzi F, Khalili K and Reiss K: Activation of
the IGF-IR system contributes to malignant growth of human and
mouse medulloblastomas. Oncogene. 20:3857–3868. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin S, Girnita A, Stromberg T, Khan Z,
Andersson S, Zheng H, Ericsson C, Axelson M, Nistér M, Larsson O,
et al: Targeting the insulin-like growth factor-1 receptor by
picropodophyllin as a treatment option for glioblastoma. Neuro
Oncol. 12:19–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng CK, Fan QW and Weiss WA: PI3K
signaling in glioma-animal models and therapeutic challenges. Brain
Pathol. 19:112–120. 2009. View Article : Google Scholar : PubMed/NCBI
|