Introduction
Renal cell carcinoma (RCC) originates from the
malignant tumor of the epithelium system of urinary tubule of renal
parenchyma. RCC has become the malignant tumor with the highest
mortality in the urinary system, accounting for 2–3% of adult
malignant tumors (1,2). The incidence of RCC shows an upward
trend each year, ranking 10 in the incidence rate of male malignant
tumors in China in 2008. The causes and pathogenesis of RCC are
still unclear.
MicroRNAs (miRs) are non-coding, small RNAs 19–25
nucleotides long. The function of miRs is mainly to degrade mRNA of
target genes through binding to the 3′ untranslated region
(3), thereby regulating the
expression and function of downstream genes. miRs participate in
the regulation of development, cell apoptosis, proliferation and
differentiation and other fundamental cell biology activities
(4,5).
In recent years it was discovered that many miRs play an important
role in the development of tumors such as proto-oncogenes or tumor
suppressor genes. Among them miR-21 is the most prominent.
Experimental results suggest that in most of the tumors, such as
malignant glioma, breast cancer, bile duct cancer, pancreatic
cancer, lung cancer, rectal cancer, bladder cancer, kidney cancer,
and esophageal cancer, the expression of miR-21 is significantly
increased (6–14) indicating that miR-21 plays an
important role in the development of tumors.
The Hippo signaling pathway is an evolutionarily
conserved regulator of cell growth inhibition. In mammals, when the
receptor receives the growth inhibitory signals, it will
phosphorylate downstream effectors YAP after the phosphorylation
cascade reaction of a series of kinase complexes. Phosphorylated
YAP interacts with cytoskeletal proteins, is retained in the
cytoplasm and cannot enter the nucleus to exercise transcriptional
activation, thus regulating organ size and volume. In addition,
recent studies have confirmed the Hippo signaling pathway is also
involved in the regulation of cancer, tissue regeneration, and the
function of stem cells. Large tumor suppressor gene 1 (LATS1) binds
and phosphorylates YAP1 in vitro and in vivo,
affecting its transcription regulation (15,16).
Studies show that low expression of LATS1 can lead to the
occurrence of astrocytoma, breast cancer, head and neck squamous
cell carcinoma, colorectal cancer, cervical cancer, gastric cancer,
skin cancer, metastatic prostate cancer, and RCC (17–25). Some
studies propose that miR-21 can resist radiation therapy by
inhibiting the expression of LATS1 in ovarian cancer cells
(26). LATS1 mRNA may be a direct
target of miR-21 and the relationship between miR-21 and LATS1 in
RCC has not been reported yet. This study used small interfering
RNA (siRNA) to silence miR-21 expression in RCC Caki-2 cells. We
also observed the role of miR-21 in renal carcinoma cell
proliferation and invasion, and tumor stem cell phenotype to help
understand miR-21 function and its targets.
Materials and methods
Cell line and main reagents
Human renal carcinoma Caki-2 cells were from
American Type Culture Collection (ATCC; Manassas, VA, USA),
Dulbecco's modified Eagle's medium (DMEM) culture and fetal bovine
serum from Gibco Life Technologies (Carlsbad, CA, USA), methyl
thiazolyl tetrazolium (MTT) kit from Sigma (St. Louis, MO, USA),
miR isolation extraction and separation kit, RPMI-1640,
Lipofectamine® 2000 and TRIzol from Invitrogen Life
Technologies (Carlsbad, CA, USA). The quantitative (qPCR) (qPCR)
Mix kits were from Takara Bio, Inc. (Otsu, Japan) and mouse
monoclonal LATS1 antibody (dilution, 1:500; cat. no. sc-398560),
mouse monoclonal Nanog antibody (dilution, 1:500; cat. no.
sc-293121), mouse monoclonal OCT3/4 (dilution, 1:500; cat. no.
sc-5279), and mouse monoclonal GAPDH antibody (dilution, 1:500;
cat. no. sc-293335) were from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA). Primers were designed and synthesized by Shanghai
Jima Pharmaceutical Technology Co., Ltd. (Shanghai, China).
si-miR-21 and si-negative control were synthesized by Shanghai Jima
Pharmaceutical Technology. si-miR-21 forward,
5′-GATCCAUCUTCGAAGUGACTT-3′ and reverse,
5′-UGCUCUTUGACGUAUGGAGTT-3′; si-negative control forward,
5′-UUCACCGUACGUCUCACCUGT-3′ and reverse,
5′-ACUGGAACCUCUCGCGGAATT-3′; LATS1 forward,
5′-AAATGCCCACATCCGGGAAA-3′ and reverse,
5′-ACCTGGCTCTCCCCTTAACA-3′.
si-miR-21 and si-negative control
transfection of Caki-2 cells
The cells in the logarithmic growth phase were
divided into three groups based on treatment: miR-21, si-miR-21,
and si-negative control. When the cell fusion reached 80–90%,
transfection was conducted with Lipofectamine® 2000
according to instructions. At 48 h after transfection, miR was
extracted following the manufacturer's instructions. We used light
photometer to detect RNA absorbance and calculate RNA concentration
and purity. The samples with a rate from 1.8 to 2.1 were selected
for further experiments. qPCR was used to conduct comparative
analysis of miR-21 mRNA in Caki-2 in the three groups after
transfection.
qPCR
Total RNA was extracted from the three groups and
purified according to the TRIzol kit instructions; 1 µg RNA
template was converted into cDNA by reverse transcription according
to reverse transcription kit. The reaction system was 10 µl, at
37°C for 15 min and heated at 85°C for 30 sec. The amplification
condition was: 95°C for 5 min, then 95°C for 30 sec, 58°C for 30
sec, 72°C for 30 sec, a total of 30 cycles, and finally 72°C for 5
min. Analysis of dissolution curve was conducted after the
reaction.
Western blotting
Total protein from cells of the three groups was
extracted by RIPA lysis solution plus 1% PMSF, and the protein
concentration was detected by BCA (all from Biosharp, Hefei,
China). A 1/4 volume of the protein sample buffer was added,
denatured at 100°C for 10 min. SDS-polyacrylamide separating gel
(Keygen, Nanjing, China) and stacking gel were prepared, Tris
glycine electrophoretic buffer solution and equal amount of
denatured protein of ~50 µg per well were added, proteins were
transferred to PVDF membrane after electrophoresis and blocked in
3% BSA for 2 h. Primary antibodies (anti-LATS1 1:1,000; anti-GAPDH
1:1,000) were added at 4°C overnight, TBST membrane was washed 3
times/10 min, then horseradish peroxidase-labeled secondary
antibodies (1:5,000) were added at 37°C for 2 h, TBST membrane was
washed 3 times/10 min, ECL development, and development in vilber
lourmat in the dark.
Proliferation assay by MTT
Cells from each group were inoculated in 96-well
culture plate at a density of 2×103 cells/well. Three
multiple pores were set and 2 ml of 5 mg/ml MTT was added to each
pore after 24, 48, 72, 96 and 120 h. They were incubated for 4 h,
then the supernatant was discarded, 150 µl of DMSO was added,
mixed, dissolved and crystallized. The light density value [D
(490)] in each pore was determined at 490 nm wavelength, and cell
proliferation inhibitory rate (IR): was calculated IR =
[1-experimental group D (490)/the control group D (490)] × 100.
Invasion ability of Caki-2 cells by
Transwell
Experiments were conducted according to the BioCoat
Matrigel invasion chamber kit instructions. Eight different visual
fields (magnification, ×100) were counted under the microscope,
repeating 3 times, and the percentage of the invasion cells was
calculated.
Tumor sphere formation
Serum-free medium (SFM) was prepared with Ham's
DMEM-F12 (1:1), B27 (1:50), EGF (20 mg/ml), and bFGF (20 ng/ml).
The cells of the three groups were inoculated in DMEM with 10%
fetal bovine serum until the cells were in the logarithmic growth
phase, and they were washed with PBS, digested with 0.25% trypsin,
and washed in PBS twice after digestion. Then the cells were placed
in the low adhesion culture in suspension for 48 h and changes in
cell morphology were observed under an inverted microscope. The
formation of tumor spheres was observed and the numbers of tumor
spheres were counted after 14 days.
Statistical analysis
SPSS 19.0 software (SPSS, Inc., Chicago, IL, USA)
was used for plotting and statistical analysis, measurement data
are expressed by mean ± standard deviation, comparison of rate was
tested by χ2, GraphPad Prism 5.0 (GraphPad Software,
Inc., La Jolla, CA, USA) was used to carry out one-way ANOVA
analysis and plotting for qPCR results, P<0.05 was considered to
indicate a statistically significant difference.
Results
Efficiency of siRNA transfection by
qPCR
According to qPCR, transfection of Caki-2 cells with
si-miR-21 resulted in significant reduction of miR-21 compared to
transfection with si-negative control (F=71.90, P<0.05). The
silencing efficiency was 55–60% (Fig.
1).
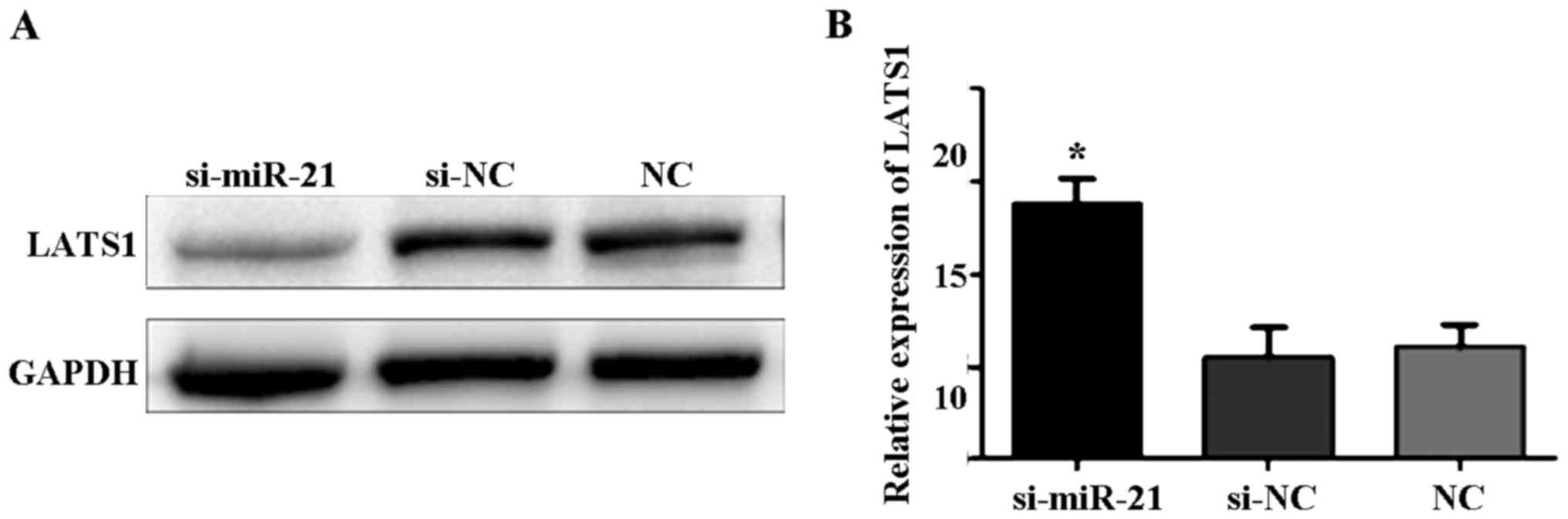
LATS1 expression after transfection of
miR-21 knockdown
Western blot analysis showed that the expression
level of LATS1 protein was significantly higher in cells
transfected with si-miR-21 compared to control cells and
si-negative cells (Fig. 2A). qPCR
showed that LATS1 mRNA expression was significantly higher in cells
transfected with si-miR-21 compared to control and si-negative
cells (F=108.5, P<0.05; Fig.
2B).
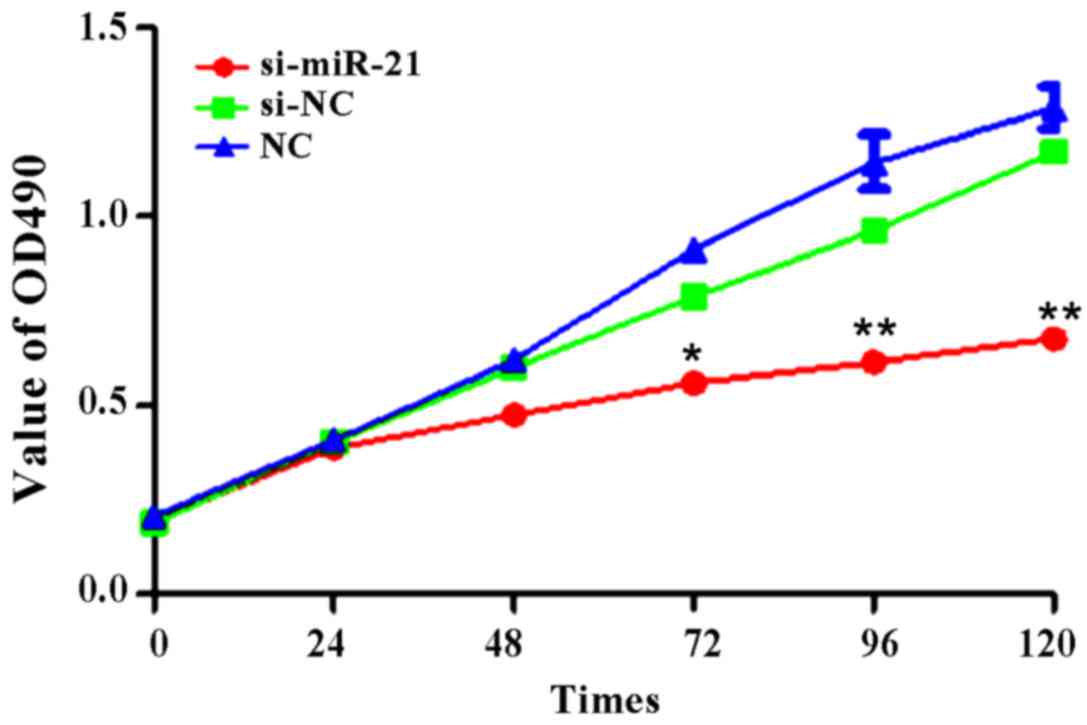
Proliferation of Caki-2 cells after
miR-21 silencing
Following cell transfection, we collecting cell
number data at 1, 24, 48, 72, 96 and 120 h. We found no significant
differences at 24 and 48 h, but si-miR-21 showed lower
proliferation after 72 h (Fig. 3).
The proliferation rate at 72 h was: 40.5±11.6% for si-miR-21,
57.4±5.9% for si-negative control, and 58.3±4.3% for
non-transfected control (t=2.375, P<0.05). Cell proliferation at
96 h was: 43.7±12.5% for si-miR-21, 75.6±7.5% for si-negative
control, and 78.3±6.9% for non-transfected control (t=4.587,
P<0.01). Cell proliferation at 120 h was: 48.9±12.3% for
si-miR-21, 81.9±4.6% for si-negative control, and 85.2±3.8% for
non-transfected control (t=5.698, P<0.01). Thus, si-miR-21
intervention inhibited the proliferation capacity of Caki-2
cells.
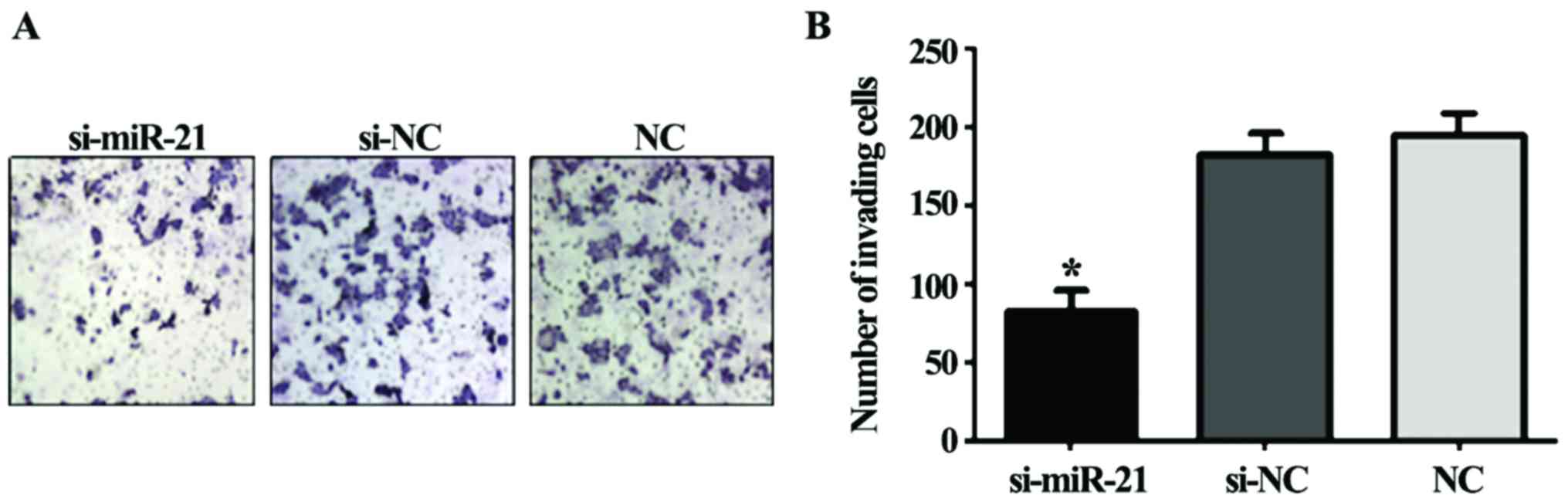
Caki-2 cell invasion after si-miR-21
transfection
The invasion ability of Caki-2 cells transfected
with si-miR-21 significantly decreased compared to the si-negative
control cells and the non-transfected cells (F=135.1, P<0.01;
Fig. 4). Thus, miR-21 knockdown
reduces the invasivity of Caki-2 cells.
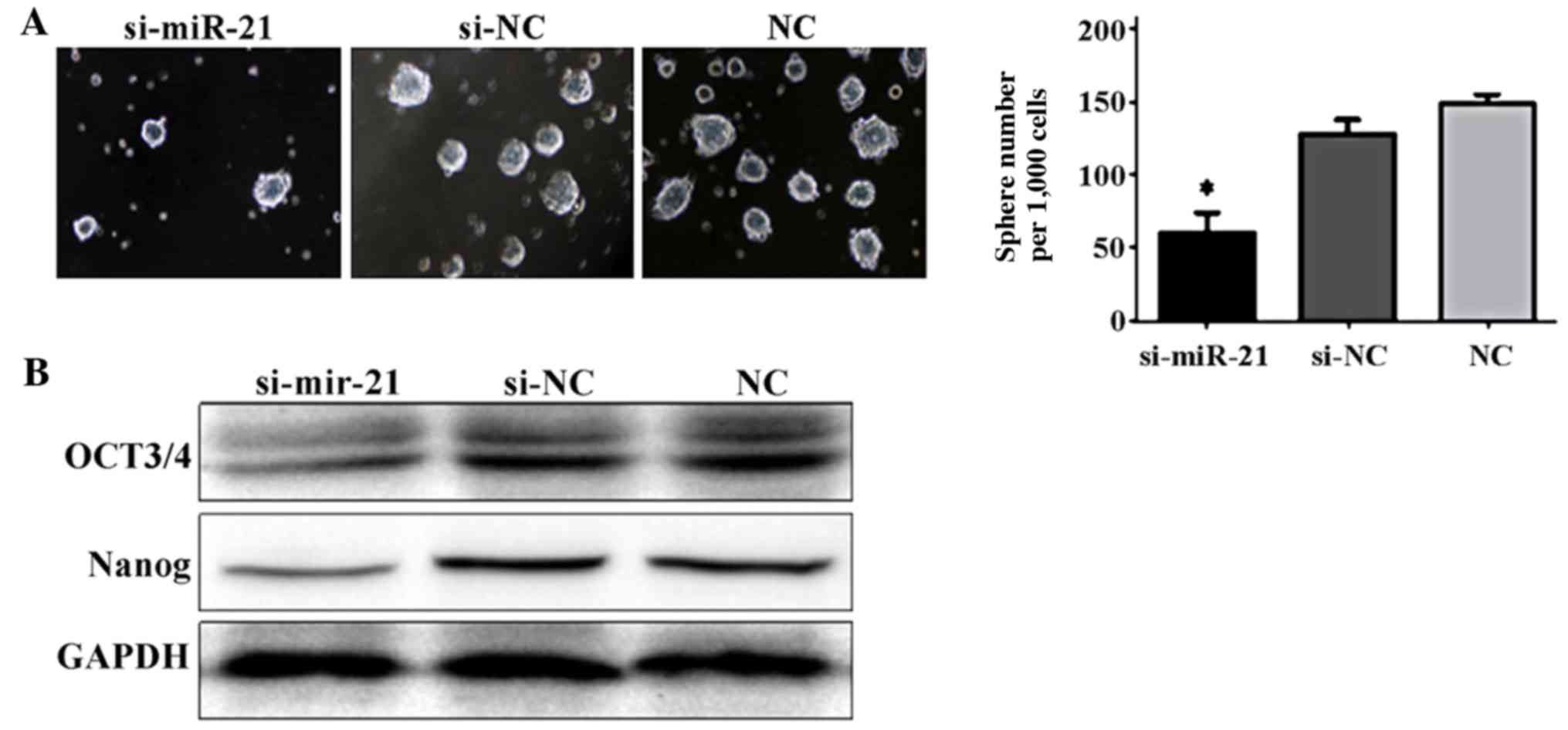
Tumor sphere formation
Compared to the si-negative control cells and the
non-transfected cells, the number of Caki-2 cells forming tumor
spheres significantly decreased in cells transfected with si-miR-21
(F=46.17, P<0.05; Fig. 4).
Phenotype of cancer stem cells
Last, we examined the ability of miR-21 to regulate
the stem cell characteristics of Caki-2 cells. For this, we
determined the expression of the stem cell markers, OCT3/4 and
Nanog. Caki-2 cells transfected with si-miR-21 showed significantly
decreased levels of OCT3/4 and Nanog compared to the si-negative
control cells and the non-transfected cells (Fig. 5). These results suggest a role for
miR-21 in the regulation of the stem cell state and the
transformation into tumorous state.
Discussion
miRs are endogenous regulatory molecules that play
roles as oncogenes or tumor suppressor genes in the development of
tumors. As one of the most studied miRs in recent years, miR-21 is
highly expressed in gastric cancer, breast cancer, colon cancer,
and other tumors, and can be used as a marker to determine the
prognosis of various tumors. Some studies suggest that miR-21
overexpression downregulate LATS1 expression. A large number of
studies have found that as one of the key factors in the hippo-Yap
signaling pathway, LATS1 expression is reduced in a variety of
tumors, which may be related to the high expression of miR-21
(27–29).
Here we showed that silencing miR-21 in Caki-2 cells
decreased the proliferation ability after 72 h and continued to
decrease until 120 h. Downregulation of miR-21 also resulted in
elevated LATS1 expression, indicating that miR-21 has a regulatory
effect on the expression of LATS1. miR-21 is known to regulate cell
differentiation, proliferation, development, apoptosis, metabolism
and cancerous transformation (30).
Tumor stem cells have self-renewal capacity and can
produce heterogeneous tumor cells. They can sustain the vitality of
the tumor cell population through self-renewal, unlimited
proliferation, movement, and migration ability to promote the
transfer of tumor cells. Cancer stem cells can be in a dormant
state for a long time and are insensitive to a variety of
chemotherapeutic drugs and external physicochemical factors killing
tumors, resulting in antitumor drug resistance and reducing the
treatment effect. To clarify the mechanism regulating tumor stem
cell state is very important for the treatment of tumors. This
study found that downregulation of miR-21 inhibited the phenotype
of tumor stem cells.
Our study results show that silencing miR-21
expression inhibited the malignant activities of renal carcinoma
cells. This inhibition may be induced by downregulation of LATS1
expression and miR-21 may be seen as a marker for diagnosis and
prognostic analysis of RCC and LATS1 may become a new target for
treatment.
References
|
1
|
Creighton CJ, Morgan M, Gunaratne PH,
Wheeler DA, Gibbs RA, Robertson Gordon A, Chu A, Beroukhim R,
Cibulskis K, Signoretti S, et al: Cancer Genome Atlas Research
Network: Comprehensive molecular characterization of clear cell
renal cell carcinoma. Nature. 499:43–49. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Junker K, Ficarra V, Kwon ED, Leibovich
BC, Thompson RH and Oosterwijk E: Potential role of genetic markers
in the management of kidney cancer. Eur Urol. 63:333–340. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Silahtaroglu A and Stenvang J: MicroRNAs,
epigenetics and disease. Essays Biochem. 48:165–185. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ross SA and Davis CD: MicroRNA, nutrition,
and cancer prevention. Adv Nutr. 2:472–485. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nimmo RA and Slack FJ: An elegant miRror:
microRNAs in stem cells, developmental timing and cancer.
Chromosoma. 118:405–418. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Selaru FM, Olaru AV, Kan T, David S, Cheng
Y, Mori Y, Yang J, Paun B, Jin Z, Agarwal R, et al: MicroRNA-21 is
overexpressed in human cholangiocarcinoma and regulates programmed
cell death 4 and tissue inhibitor of metalloproteinase 3.
Hepatology. 49:1595–1601. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dillhoff M, Liu J, Frankel W, Croce C and
Bloomston M: MicroRNA-21 is overexpressed in pancreatic cancer and
a potential predictor of survival. J Gastrointest Surg.
12:2171–2176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xie Y, Todd NW, Liu Z, Zhan M, Fang H,
Peng H, Alattar M, Deepak J, Stass SA and Jiang F: Altered miRNA
expression in sputum for diagnosis of non-small cell lung cancer.
Lung Cancer. 67:170–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rossi L, Bonmassar E and Faraoni I:
Modification of miR gene expression pattern in human colon cancer
cells following exposure to 5-fluorouracil in vitro. Pharmacol Res.
56:248–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Neely LA, Rieger-Christ KM, Neto BS,
Eroshkin A, Garver J, Patel S, Phung NA, McLaughlin S, Libertino
JA, Whitney D, et al: A microRNA expression ratio defining the
invasive phenotype in bladder tumors. Urol Oncol. 28:39–48. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lv L, Huang F, Mao H, Li M, Li X, Yang M
and Yu X: MicroRNA-21 is overexpressed in renal cell carcinoma. Int
J Biol Markers. 28:201–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feber A, Xi L, Luketich JD, Pennathur A,
Landreneau RJ, Wu M, Swanson SJ, Godfrey TE and Litle VR: MicroRNA
expression profiles of esophageal cancer. J Thorac Cardiovasc Surg.
135:255–260; discussion 260. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mo JS, Park HW and Guan KL: The Hippo
signaling pathway in stem cell biology and cancer. EMBO Rep.
15:642–656. 2014.PubMed/NCBI
|
|
16
|
Harvey K and Tapon N: The
Salvador-Warts-Hippo pathway - an emerging tumour-suppressor
network. Nat Rev Cancer. 7:182–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang Z, Li X, Hu J, Zhou W, Jiang Y, Li G
and Lu D: Promoter hypermethylation-mediated down-regulation of
LATS1 and LATS2 in human astrocytoma. Neurosci Res. 56:450–458.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Takahashi Y, Miyoshi Y, Takahata C,
Irahara N, Taguchi T, Tamaki Y and Noguchi S: Down-regulation of
LATS1 and LATS2 mRNA expression by promoter hypermethylation and
its association with biologically aggressive phenotype in human
breast cancers. Clin Cancer Res. 11:1380–1385. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Steinmann K, Sandner A, Schagdarsurengin U
and Dammann RH: Frequent promoter hypermethylation of tumor-related
genes in head and neck squamous cell carcinoma. Oncol Rep.
22:1519–1526. 2009.PubMed/NCBI
|
|
20
|
Wierzbicki PM, Adrych K, Kartanowicz D,
Stanislawowski M, Kowalczyk A, Godlewski J, Skwierz-Bogdanska I,
Celinski K, Gach T, Kulig J, et al: Underexpression of LATS1 TSG in
colorectal cancer is associated with promoter hypermethylation.
World J Gastroenterol. 19:4363–4373. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Visser S and Yang X: Identification of
LATS transcriptional targets in HeLa cells using whole human genome
oligonucleotide microarray. Gene. 449:22–29. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou GX, Li XY, Zhang Q, Zhao K, Zhang CP,
Xue CH, Yang K and Tian ZB: Effects of the hippo signaling pathway
in human gastric cancer. Asian Pac J Cancer Prev. 14:5199–5205.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nishio M, Hamada K, Kawahara K, Sasaki M,
Noguchi F, Chiba S, Mizuno K, Suzuki SO, Dong Y, Tokuda M, et al:
Cancer susceptibility and embryonic lethality in Mob1a/1b
double-mutant mice. J Clin Invest. 122:4505–4518. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao B, Li L, Wang L, Wang CY, Yu J and
Guan KL: Cell detachment activates the Hippo pathway via
cytoskeleton reorganization to induce anoikis. Genes Dev. 26:54–68.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen KH, He J, Wang DL, Cao JJ, Li MC,
Zhao XM, Sheng X, Li WB and Liu WJ: Effects of LATS1 gene
methylation on biological function in human renal cell carcinoma
and Hippo-YAP signaling pathway. J Third Mil Med Univ.
36:1249–1254. 2014.
|
|
26
|
Liu S, Song L, Zhang L, Zeng S and Gao F:
miR-21 modulates resistance of HR-HPV positive cervical cancer
cells to radiation through targeting LATS1. Biochem Biophys Res
Commun. 459:679–685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Teteloshvili N, Smigielska-Czepiel K, Yuan
Y, Seitz A, de Jong D, Rutgers B, Jellema P, van der Lei RJ,
Slezak-Prochazka I, Brouwer E, et al: Argonaute 2
immunoprecipitation revealed large tumor suppressor kinase 1 as a
novel proapoptotic target of miR-21 in T cells. FEBS J.
284:555–567. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu S, Song L, Zhang L, Zeng S and Gao F:
miR-21 modulates resistance of HR-HPV positive cervical cancer
cells to radiation through targeting LATS1. Biochem Biophys Res
Commun. 459:679–685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xia H, Qi H, Li Y, Pei J, Barton J,
Blackstad M, Xu T and Tao W: LATS1 tumor suppressor regulates G2/M
transition and apoptosis. Oncogene. 21:1233–1241. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen J and Wang X: MicroRNA-21 in breast
cancer: Diagnostic and prognostic potential. Clin Transl Oncol.
16:225–233. 2014. View Article : Google Scholar : PubMed/NCBI
|