Introduction
Betulin is a frequently occurring natural compound,
abundant in trees and grasses, making up ~33%, in mass, of the
extracts obtained from the bark of birch trees (1). It is a versatile compound that may be
used in conjunction with others to form a variety of chemicals for
a number of pharmacological usages, including in poison oak, as a
skin irritant (1). Betulinic acid,
which is derived from betulin, has been used as a cytotoxic drug to
treat tumors due to its antitumorigenic abilities (2–4). The
cytotoxicity of betulin has been the focus of previous studies;
however, the exact mechanism of action has yet to be elucidated
(5,6).
The inhibition of cellular apoptosis is a key
indicator that marks the initiation and progression of cancer
(7). Therefore, current cancer
treatments, including chemotherapy and immunotherapy, function
primarily through promoting the apoptosis of tumor cells (8,9).
Programmed cell death may be induced via either a receptor or
mitochondrial-signaling pathway. Receptor-induced apoptosis
functions through intracellular signaling transmissions via death
receptors, whereas the latter occurs through the destruction of
mitochondria (10).
The primary regulators of mitochondrion-dependent
apoptosis are B-cell lymphoma 2 (Bcl-2) proteins (11). These signaling molecules belong to the
Bcl-2 homology 3 (BH3) family, a subset of the Bcl-2 family that
contain a single BH3 domain, with >10 members, including NOXA
and Bcl-2-interacting mediator of cell death (Bim), which respond
well to single and mixed pro-apoptotic signals (12,13). They
are able to trigger apoptosis by inducing the apoptosis regulator
Bcl-2-associted X protein (Bax), while counteracting the effect of
the anti-apoptotic gene Bcl-2, in addition to stimulating caspases
resulting in mitochondrial dysfunction (14,15).
The aim of the present study was to investigate the
antitumor properties of betulin against human colon cancer cells
and elucidate its underlying mechanisms of action at a molecular
level.
Materials and methods
Cell culture
The colon cancer cell lines HCT116 and HT29, and the
normal human colon epithelial cell line CCD 841 CoN were purchased
from the American Type Culture Collection (Manassas, VA, USA), and
cultured in Dulbecco's modified Eagle's medium containing 100 U/ml
penicillin, 10% bovine serum and 100 µg/ml streptomycin in a
humidified incubator with 5% CO2 at 37°C. Betulin was
obtained from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany).
Analysis of cell viability and colony
formation
An MTT assay was used to determine cell viability.
Briefly, 1,000 cells were seeded into each well of 96-well plate
and the formazan grains formed by viable cells were solubilized
with 200 ml dimethyl sulfoxide. The color intensity was measured at
550 nm via a 96-well plate reader (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). In the crystal violet staining assay, a total of
5,000 cells were placed in in 12-well plates for 14 days at 37°C
and then stained with 1% crystal violet for 30 sec at room
temperature (Sigma-Aldrich; Merck KGaA).
Nuclear staining using Hoechst
33258
Colon cancer cells were fixed using 4%
paraformaldehyde following betulin treatment (10 mg/mL). Hoechst
33342 (10 µg/ml; Sigma-Aldrich; Merck KGaA) was used to stain cells
for 10 min at 37°C. Following this, cells were washed with PBS 2
times and a fluorescent microscope at ×200 magnification (DM1000
LED; Leica Microsystems, Ltd., Milton Keynes, UK) was used to
observe the apoptotic cells. Cells which exhibited nuclear
fragmentation and chromatin condensation were considered to be
apoptotic cells, and cells that exhibited round and regular nuclei
were considered to be normal cells. Twenty fields were randomly
selected in each slide to count apoptotic cells.
Flow cytometric analysis
Flow cytometric analysis was performed as previously
outlined by He et al (16).
Briefly, cells were re-suspended in cold binding buffer from the
Alexa Flouor® 488 Annexvin V Cell Apoptosis kit
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) (200
µl), followed by the addition of propidium iodide (5 µl) and
Annexin V-fluorescein isothiocyanate (10 µl) (Invitrogen; Thermo
Fisher Scientific, Inc.). Cells were incubated at room temperature
for 15 min in darkness, and 300 µl binding buffer from the Alexa
Flouor® 488 Annexvin V Cell Apoptosis kit was added to
block the reaction. A flow cytometer was used (BD Biosciences, San
Jose, CA, USA). Cells which were positive for Annexin-V were
considered to be apoptotic cells. Data was analyzed using BD
AccuriTM C6 Software (version 1.0.264.21; BD Biosciences).
Reverse-transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from betulin-treated cells
using a Mini RNA Isolation II kit (Zymo Research Corp., Irvine, CA,
USA), according to the manufacturer's protocol. Superscript II
reverse transcriptase was used to create cDNA from the total RNA.
Real-time PCR with SYBR Green (Invitrogen; Thermo Fisher
Scientific, Inc.) was conducted as previously described (17). The thermocycling conditions were as
follows; 94°C 5 min, 94°C 30 second, 58°C 1 min, 72°C 2 min, 72°C
10 min. The following primers were used: p53-upregulated modulator
of apoptosis (PUMA), 5′-CGACCTCAACGCACAGTACGA-3′ (forward) and
5′-AGGCACCTAATTGGGCTCCAT-3′ (reverse); Bim,
5′-GGAGACGAGTTTAACGCTTAC-3′ (forward) and
5′-CAAGCAAAATGTCTGCATGG-3′ (reverse); NOXA,
5′-GCTGGAAGTCGAGTGTGCTA-3′ (forward) and 5′-CCTGAGCAGAAGAGTTTGGA-3′
(reverse); Bcl-2-associated death promoter (BAD),
5′-CGGAGGATGAGTGACGAGTT-3′ (forward) and 5′-GATGTGGAGCGAAGGTCACT-3′
(reverse); β-actin, 5′-GACCTCACAGACTACCTCAT-3′ (forward) and
5′-AGACAGCACTGTGTTGGCTA-3′ (reverse). The mRNA levels were
quantified using the 2−ΔΔCq method (17). β-actin was used as the internal
control. Experiments for each gene were conducted in
triplicate.
Small-interfering RNA (siRNA) and
plasmid transfection
NOXA siRNA duplexes 200 pmol
(5′-GUAAUUAUUGACACAUUUCUU-3′) and control scrambled siRNA 200 pmol
(Guangzhou RiboBio Co., Ltd., Guangzhou, China) or pCDNA and
pcDNA-NOXA plasmid (Guangzhou RiboBio Co., Ltd.) were introduced
into HCT116 cells with Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) for 1 day and were treated with 10
µg/ml betulin for 24 h following transfection.
Caspase evaluation
Cells were treated with betulin (10 mg/ml) for 0, 4
8, 12 and 24 h. Cell lysates (50 µg) were incubated with 200 nM
fluorigenic substrates for caspase-3, −8 and −9 (N-acet
yl-Asp-Glu-Val-Asp-7-amido-4-trifluoromethylcoumarin,
N-acetyl-Ile-Glu-Thr-Asp-7-amido-4-trifl uoromethylcoumarin and
N-acetyl-Leu-Glu-His-Asp-7-amido-4-trifluoromethylcoumarin,
respectively) (Sigma-Aldrich; Merck KGaA) in a buffer solution
comprising 20 mM 4-(2-hydroxyethyl)-1-piperazine-ethanesulfonic
acid (pH 7.4), 100 mM NaCl, 0.1%
3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate,
dithiothreitol (10 mM) and 10% sucrose at 37°C for 1 h. Following
incubation, fluorescence was determined using a Biotek microplate
reader (BioTek Instruments, Inc., Winooski, VT, USA) at emission
and excitation at 535 nm and 405 nm, respectively.
Cytochrome c analysis
A 10 µg/ml concentration of betulin was administered
to HCT116 cells for 24 h. The isolated cytoplasm components and
mitochondrial particles were analyzed using the Mitochondrial
Fractionation kit (Active Motif, Inc., Carlsbad, CA, USA),
according to the manufacturer's protocol. The total protein
concentrations was calculated using a BCA assay (Thermo Fisher
Scientific, Inc.). Western blotting was performed to evaluate
cytochrome c expression in the cytoplasm and mitochondria.
Briefly, protein was extracted using radioimmunoprecipitation assay
buffer (Abcam, Cambridge, UK). In total, 50 µg protein was loaded
into each well (12.5% Bis-Tris gel). Proteins were then transferred
to a polyvinylidene difluoride membrane. The enhanced
chemiluminescence western blotting detection system (Tanon 4200;
Tanon Science and Technology Co., Ltd., Shanghai, China) and each
band was analyzed following western blotting, using Quantity One
software (version 1.03; Tanon Science and Technology Co., Ltd.).
Following treatment with betulin, cells were fixed with 4%
paraformaldehyde solution for 10–20 min at room temperature and
incubated in PBS containing 0.1% Tween-20 and 5% normal goat serum
for 30 min at room temperature, followed by antibody incubation in
a diluted solution in the ratio of cytochrome c (dilution
1:200) at room temperature for 3 h. Primary antibodies included
cleaved caspase-9 (catalog no. 20750, dilution 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA), cleaved caspase-3
(catalog no. 9661, dilution 1:1,000; Cell Signaling Technology,
Inc.), cleaved caspase-8 (catalog no. 9496, dilution 1:1,000; Cell
Signaling, Danvers, MA, USA), cytochrome c (catalog no.
ab13575, dilution 1:1,000; Abcam, Shanghai, China), and NOXA
(catalog no. ab23563, dilution 1:1,000; Abcam, Shanghai, China).
Horseradish peroxidase-conjugated secondary antibodies included
goat anti-rabbit IgG (catalog no. ab6702, dilution 1:5,000; Abcam,
Shanghai, China) and goat anti-mouse IgG (catalog no. ab6708,
dilution 1:5,000; Abcam, Shanghai, China) were incubated for 1 h at
room temperature.
Immunofluorescence staining was performed using
anti-mouse antibodies (cat. no. F0111; dilution 1:500; Dako;
Agilent Technologies, Inc., Santa Clara, CA, USA) conjugated with
fluorescein isothiocyanate. Subsequently, cells were washed with
PBS (containing 0.1% Tween) and nuclei were treated with DAPI
staining (dilution 1:5,000) for 10 min. Immunofluorescence staining
was visualized using an Olympus microscope (magnification, ×40).
All experiments were performed in triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation
and analyzed with Student's t-test or one-way analysis of variance
(ANOVA), Tukey's multiple comparison was applied as a post hoc test
following ANOVA. Statistical differences were determined with SPSS
22.0 (IBM Corp., Armonk, NY, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Cytotoxicity of betulin in colon
cancer cells
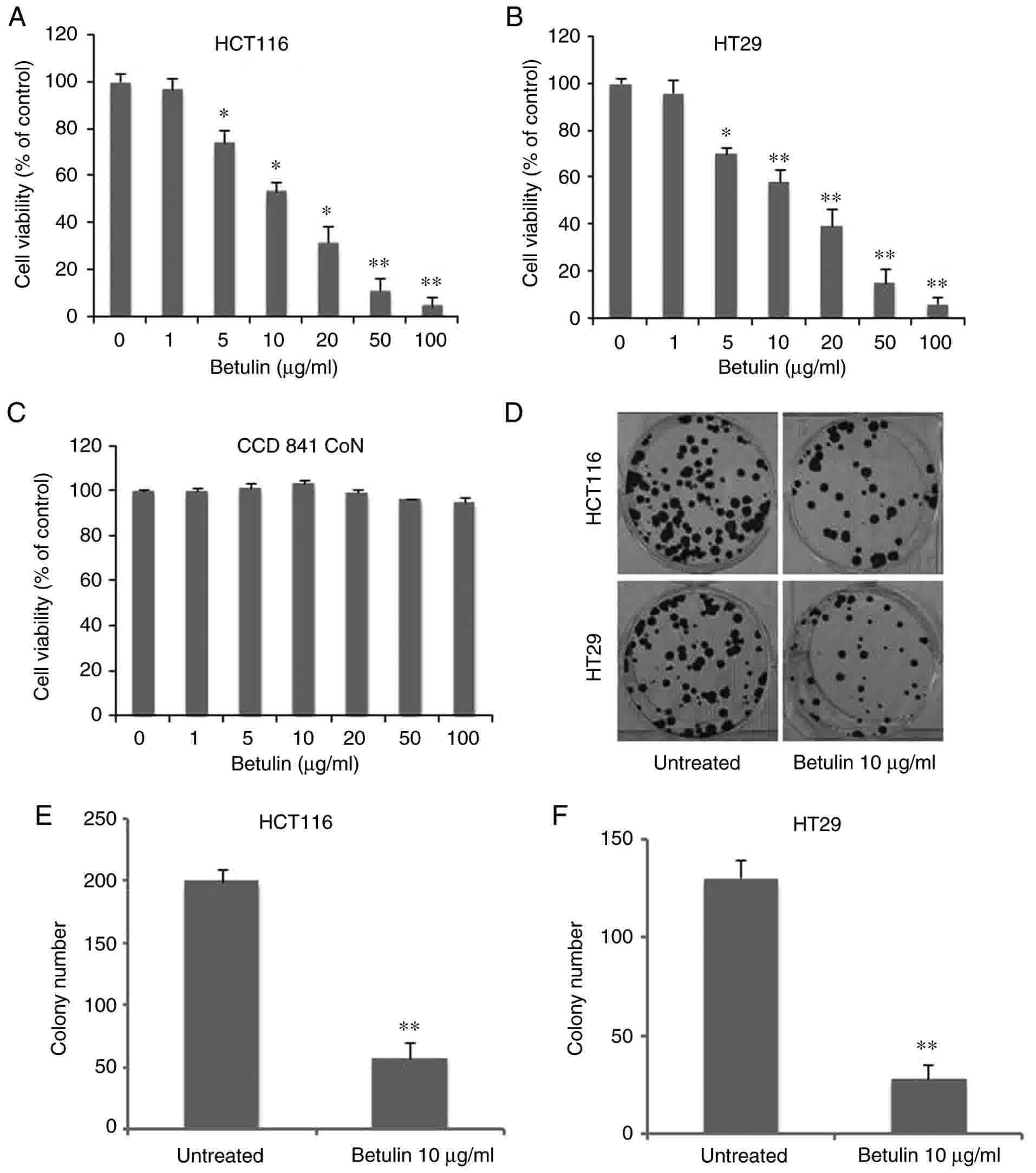
In order to determine the antitumor capabilities of
betulin against colon cancer cell lines, an MTT assay was conducted
to examine the cytotoxic effects of betulin in HCT116 and HT29
cells. As Fig. 1A demonstrates,
treatment with betulin significantly decreased the viability of
HCT116 cells at concentrations >5 µg/ml, reaching >90% at 100
µg/ml following 72 h of incubation. Betulin treatment of the other
cell line examined, HT29, also exhibited a similar dose-dependent
effect (Fig. 1B). The median lethal
dose value of betulin (~10 µg/ml) was similar for HCT116 and HT29
cells. Consequently, the standard concentration of betulin for
subsequent experiments was set at 10 µg/ml. However, this dosage
had limited effect on the normal human colon epithelial cells CCD
841 CoN (Fig. 1C). Additionally, the
long-term colony-formation capacities of the two colon cancer cell
lines following betulin treatment were determined using crystal
violet staining. Compared with the untreated group, betulin-treated
cells produced >60±5.3% fewer colonies in HCT116 and HT29 cells
(Fig. 1D). However, HT29 cells were
more sensitive to the effects of betulin (Fig. 1E-F). These results demonstrated that
betulin is cytotoxic towards colon cancer cells.
Betulin induces apoptosis in colon
cancer cells
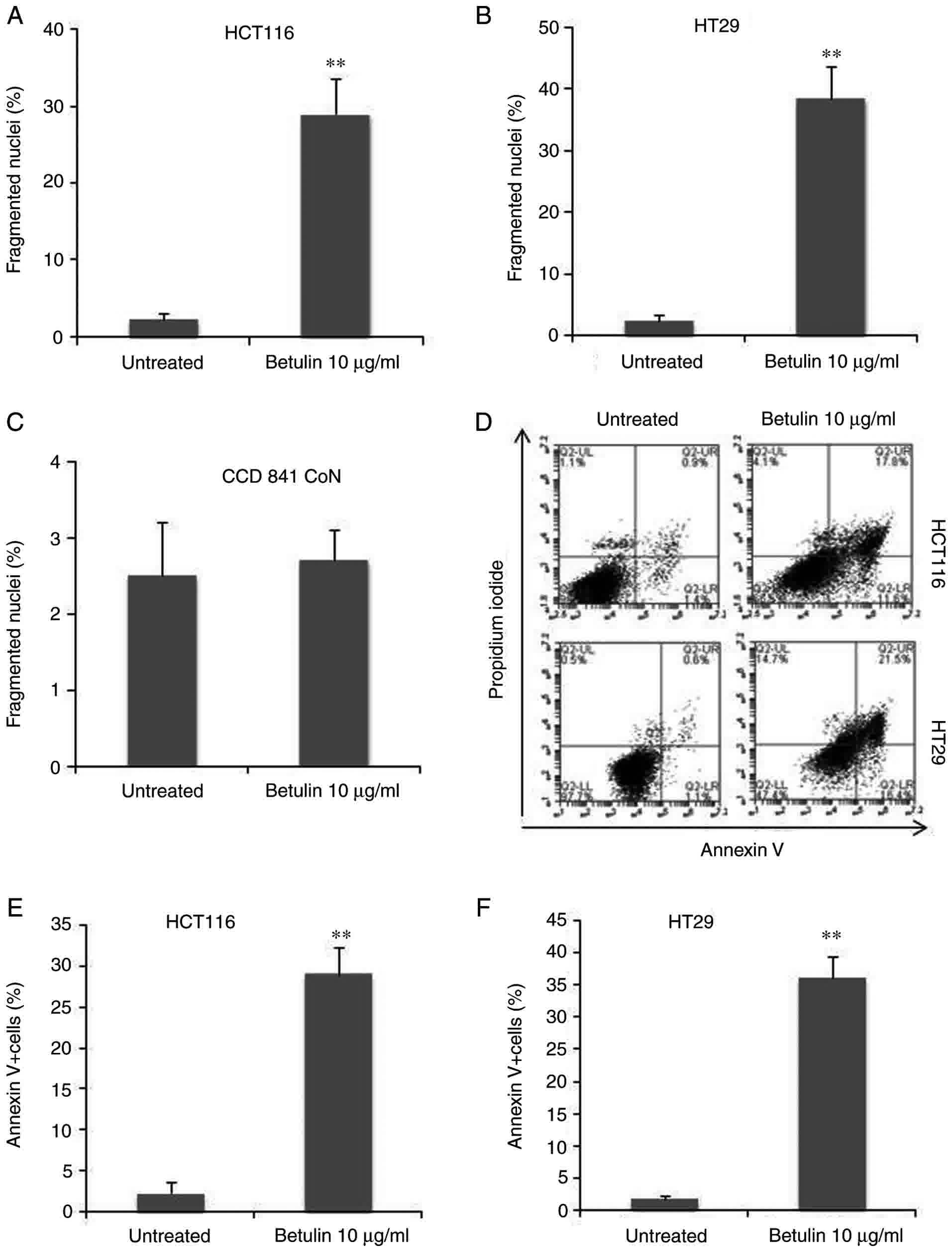
In order to examine the association between
decreasing cell viability and programmed cell death, the preset
study examined and compared the apoptotic attributes of HCT116 and
HT29 cells following betulin treatment. First, the results of the
present study identified an increase in the number of floating
cells following betulin treatment compared with the untreated
groups, which was suggestive of apoptosis due to HCT116 cells being
adherent. This was also accompanied by several other typical
characteristics of apoptotic cell death, including the rounding of
cells and the emergence of irregular bulges in cell membranes (data
not shown). Secondly, HCT116 and HT29 cells with condensed
chromatin and micronucleation following nuclear staining with
Hoechst 33258 were counted with and without treatment with 10 µg/ml
betulin. Betulin treatment induced between 30±3.4% and 40±4%
fragmented nuclei (+) cells after 48 h in HCT116 and HT29 cells,
respectively (Fig. 2A and B).
However, this dosage had limited effect on the normal human colon
epithelial cells CCD 841 CoN (Fig.
2C). Furthermore, apoptosis was assessed using Annexin V
analysis; the results demonstrated that >30% of HCT116 and HT29
cells were Annexin V (+) 48 h after being treated with betulin
compared with their respective control groups (Fig. 2D-F). These findings indicated that
programmed cell death may be induced by betulin in human colon
cancer cells.
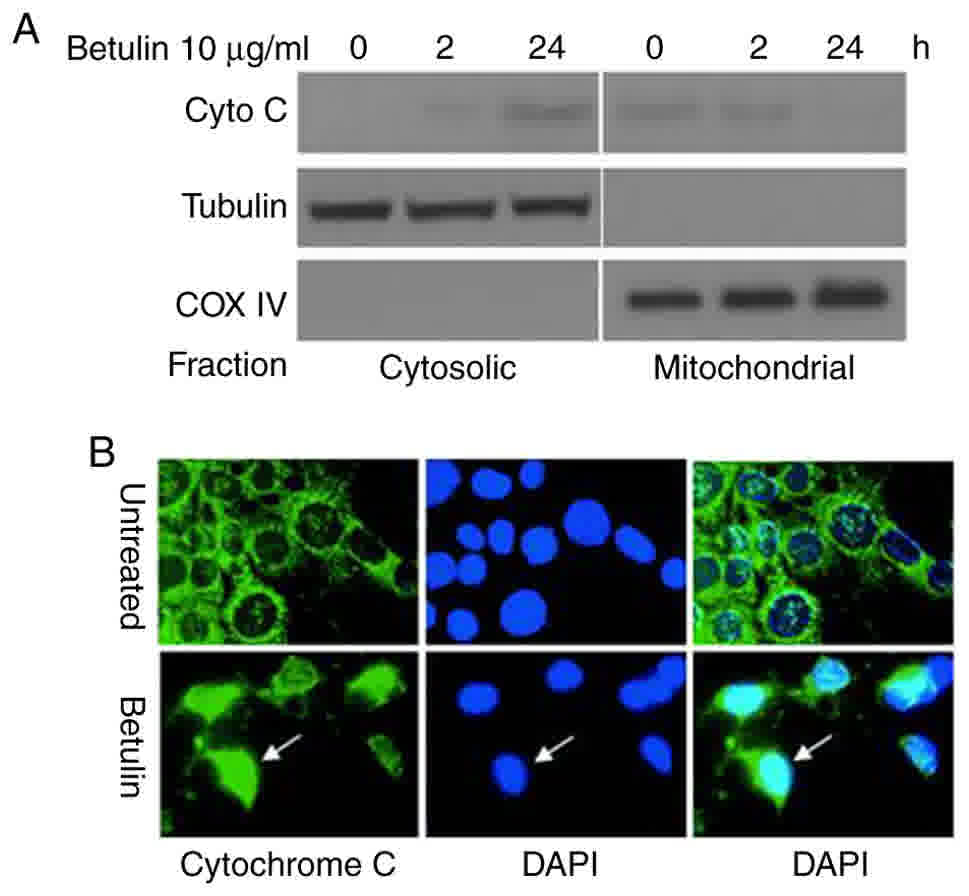
Betulin induces the release of
caspase-3 and −9, and cytochrome c
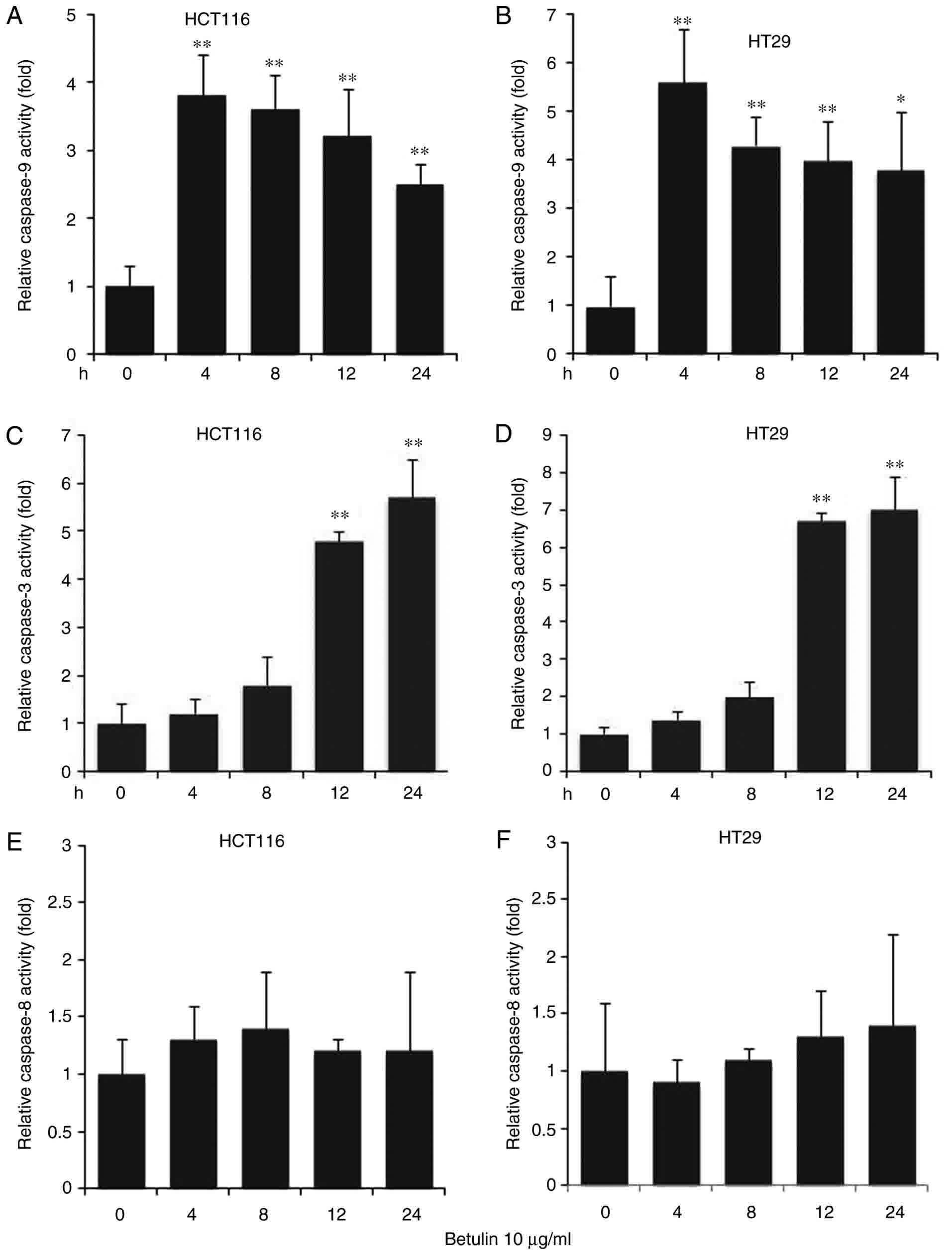
The effect of betulin on activating the upstream
effectors caspase-8 and −9, and the downstream effector caspase-3
was investigated in colon cancer cells. A significantly increased
level of caspase-9 was observed in betulin-treated cells after 4 h
in comparison with the respective negative control groups;
thereafter, the levels of capspase-9 gradually decreased (Fig. 3A and B). A significant increase in the
activity of caspase-3 was not detected until after 8 h of betulin
treatment (Fig. 3C and D); however,
the increase in caspase-3 activity was observed from 4 to 24 h of
betulin treatment. However, the level of relative caspase-8
activity did not alter significantly throughout the experiment in
the cancer cell lines examined (Fig. 3E
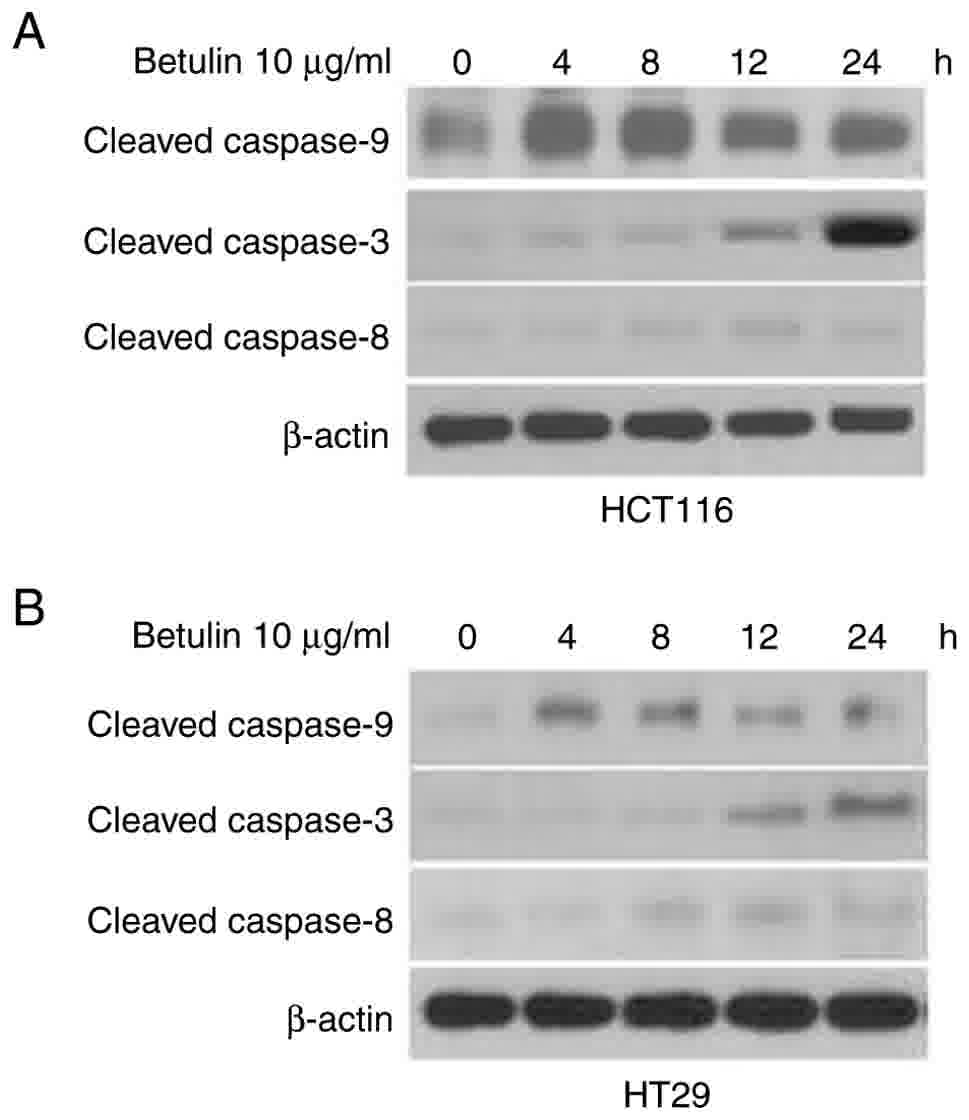
and F). Western blot analysis confirmed the increased level of
cleaved caspase-9, −3 and −8 following betulin treatment in HCT116
and HT29 cells (Fig. 4). As betulin
was demonstrated to stimulate caspase-9 activation, the present
study proceeded to examine the releasing level of cytochrome
c in HCT116 cells following betulin treatment. Using
immunoblotting and immunofluorescence techniques, the results of
the present study demonstrated that cytochrome c was
released in the cytosolic area of cells following treatment with
betulin; furthermore, its level decreased in the mitochondrial
fraction following 24 h treatment with betulin in contrast with
that in of the control group (Fig. 5A and
B). These results indicated that betulin was able to activate
caspase-9 and −3, in addition to releasing cytochrome c in
colon cancer cells.
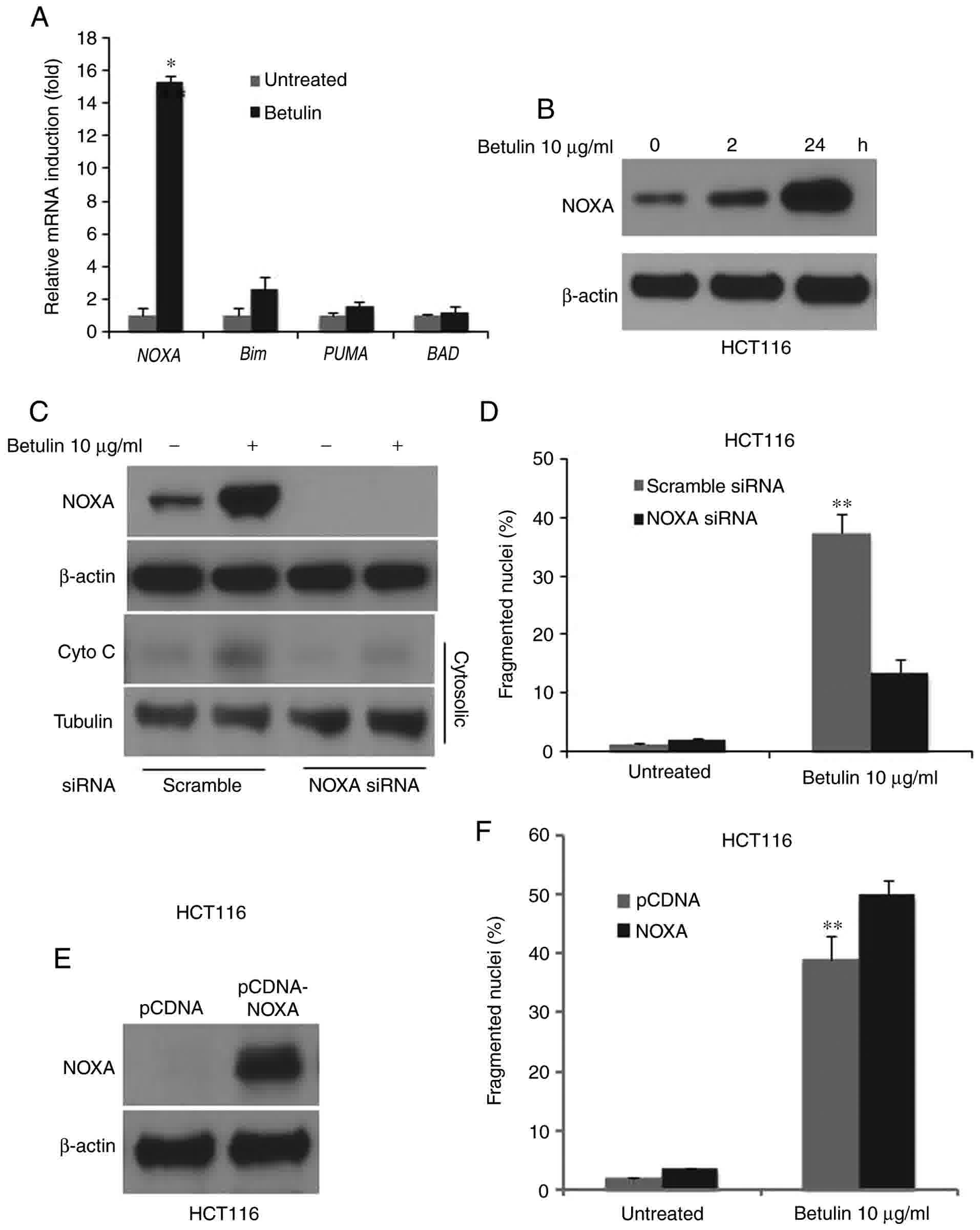
NOXA mediates apoptosis in
betulin-treated colon cancer cells
In order to determine the possible effect of the
BH3-only proteins in betulin-induced apoptosis, the relative mRNA
levels of its major members in betulin-treated HCT116 colon cancer
cells were determined using RT-qPCR. Results demonstrated a
markedly increased mRNA level of NOXA and Bim, whereas PUMA and BAD
remained unchanged following betulin treatment (Fig. 6A). Notably, NOXA was induced almost
15-fold at the mRNA level and significantly increased at the
protein level as demonstrated using western blot analysis (Fig. 6B). siRNA against NOXA was employed to
investigate whether NOXA serves a role in betulin-induced apoptosis
in colon cancer cells. The fragmented nuclei (+) cell percentage
and cytochrome c release following betulin treatment was
significantly abrogated in the NOXA siRNA group compared with the
scrambled siRNA group (Fig. 6C and
D). Furthermore, overexpression of NOXA further enhanced the
betulin-induced apoptosis (Fig. 6E and
F). These results suggested that NOXA mediated apoptosis in
betulin-treated colon cancer cells.
Discussion
Vinca alkaloids and taxanes are two
categories of naturally occurring compounds that are used as
chemotherapeutic agents. Vinblastine and vincristine, derived from
Catharanthus roseus, were the first to be applied in a
clinical setting in 1958, which were followed by various
derivatives including vindesine and vinflunine (18). Betulin is in same class as taxanes
known as isoprenoids (19). Betulin
is recognized as a positive agent against human immunodeficiency
virus due to its ability to suppress the replication process in
human immunodeficiency virus (20).
Consequently, an increasing number of studies are now focusing on
its antitumor potential. In the present study, it was demonstrated
that the naturally obtainable compound betulin was able to activate
apoptosis in human colon cancer cells via the caspase-3 and −9
activation pathways, additionally releasing cytochrome c.
Furthermore, NOXA was markedly induced in comparison with several
BH3 proteins following betulin treatment. Furthermore, knockdown of
NOXA also significantly abrogated the betulin-induced apoptosis in
colon cancer cells. Therefore, the results of the present study
contribute to the overall understanding of the mechanism of
programmed cell death stimulated by betulin in human colon cancer
cells.
A number of studies have previously demonstrated
that programmed cell death is a key procedure in the mechanism of
how chemotherapeutic agents act to destroy tumor cells (21). Protein cleavage may be induced via
activation of caspases, which leads to apoptosis. Caspase
activation occurs one of two ways: i) Through the activation of
death receptors on the cell surface, which induces the upstream
caspase-8 and therefore results in the activation of the downstream
caspase-3 and −7; or ii) through the mitochondria, by the release
of cytochrome c, which activates the apoptotic protease
apoptotic protease-activating factor 1 (Apaf-1), leading to the
activation of caspase-9 (22–24). Through examination of the
caspase-mediated process, the present study demonstrated that the
apoptosis induced by betulin in human colon cancer cells is not, to
the best of our knowledge, associated with other external apoptotic
pathways, which is consistent with the effect of betulin in other
types of tumor cell (25).
The Bcl-2 family protein controls the
permeabilization procedures of the outer membrane of mitochondria
and also regulates the release of cytochrome c. Previous
studies have identified that that the function of and interactions
between different proteins within the Bcl-2 family are restricted,
which also provides insight into the activation of Bax and Bcl-2
homologous antagonist killer (26,27). At
present, only Bid and Bim have been identified as the BH3 proteins
capable of interacting directly with Bax, whereas other family
members, including BAD, Puma and NOXA, exhibit their functions
indirectly, i.e., via ‘sensitization’ or ‘derepression’ and
exclusive interactions with protective Bcl-2-like members.
Consequently, various members of the BH3 domain-only group exhibit
differences in their response to stimuli and in the pathways with
which they are capable of regulating (28). The results of the present study
demonstrated that the induction of NOXA is necessary for
betulin-induced apoptotic cell death and the release of cytochrome
c in colon cancer cells. On the basis of the effect of NOXA
on mediating the permeabilization process of the outer
mitochondrial membrane (29,30), our hypothesis is that the
NOXA-dependent action of betulin-induced apoptosis is common in
colon cancer cells. As there was a small increase in cytochrome
c in the cytosolic fraction following NOXA knockdown, other
mechanisms may also be involved. The limitation of the present
study is the focus on the short-term effect of betulin on colon
cancer cells; however, the long-term effect in vivo and
in vitro require further study.
In conclusion, the results of the present study
demonstrated the antitumor abilities of betulin by triggering
apoptosis in human colon cancer cells. Cytochrome c was also
released and, together with the subsequent activation of caspases,
apoptosis was also induced via the mitochondrial apoptotic pathway.
The induction of NOXA also served a role in the apoptotic response
of colon cancer cells following betulin treatment. The results of
the present study have provided a novel mechanistic insight into
this type of naturally occurring chemical compound as a cancer
therapeutic and therefore has the potential to contribute to
overall understanding and the future development of betulin as a
novel cancer therapeutic.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Project of the Shanghai Municipal Commission of Health and Family
Planning (grant no. 201440355).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author, on reasonable
request.
Authors' contributions
Conception and design, XQ; development of
methodology: ZZ, CZ, ZC, FZ, LH and XL; analysis and interpretation
of data, ZZ, CZ, ZC; writing, review, and/or revision of the
manuscript, ZZ and XQ.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Alakurtti S, Makela T, Koskimies S and
Yli-Kauhaluoma J: Pharmacological properties of the ubiquitous
natural product betulin. Eur J Pharm Sci. 29:1–13. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gauthier C, Legault J, Lebrun M, Dufour P
and Pichette A: Glycosidation of lupane-type triterpenoids as
potent in vitro cytotoxic agents. Bioorg Med Chem. 14:6713–6725.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kvasnica M, Sarek J, Klinotova E, Dzubak P
and Hajduch M: Synthesis of phthalates of betulinic acid and
betulin with cytotoxic activity. Bioorg Med Chem. 13:3447–3454.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rabi T, Shukla S and Gupta S: Betulinic
acid suppresses constitutive and TNFalpha-induced NF-kappaB
activation and induces apoptosis in human prostate carcinoma PC-3
cells. Mol Carcinog. 47:964–973. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pyo JS, Roh SH, Kim DK, Lee JG, Lee YY,
Hong SS, Kwon SW and Park JH: Anti-cancer effect of betulin on a
human lung cancer cell line: A pharmacoproteomic approach using 2 D
SDS PAGE coupled with nano-HPLC tandem mass spectrometry. Planta
Med. 75:127–131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rzeski W, Stepulak A, Szymanski M,
Juszczak M, Grabarska A, Sifringer M, Kaczor J and
Kandefer-Szerszen M: Betulin elicits anti-cancer effects in tumour
primary cultures and cell lines in vitro. Basic Clin Pharmacol
Toxicol. 105:425–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brown JM and Attardi LD: The role of
apoptosis in cancer development and treatment response. Nat Rev
Cancer. 5:231–237. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Herr I and Debatin KM: Cellular stress
response and apoptosis in cancer therapy. Blood. 98:2603–2614.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Whiteside TL: Apoptosis of immune cells in
the tumor microenvironment and peripheral circulation of patients
with cancer: Implications for immunotherapy. Vaccine. 20 Suppl
4:A46–A51. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thomenius MJ, Wang NS, Reineks EZ, Wang Z
and Distelhorst CW: Bcl-2 on the endoplasmic reticulum regulates
Bax activity by binding to BH3-only proteins. J Biol Chem.
278:6243–6250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
van Delft MF, Wei AH, Mason KD, Vandenberg
CJ, Chen L, Czabotar PE, Willis SN, Scott CL, Day CL, Cory S, et
al: The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and
efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized.
Cancer Cell. 10:389–399. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alves NL, Derks IA, Berk E, Spijker R, van
Lier RA and Eldering E: The Noxa/Mcl-1 axis regulates
susceptibility to apoptosis under glucose limitation in dividing T
cells. Immunity. 24:703–716. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lei K and Davis RJ: JNK phosphorylation of
Bim-related members of the Bcl2 family induces Bax-dependent
apoptosis. Proc Natl Acad Sci USA. 100:2432–2437. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He K, Zheng X, Li M, Zhang L and Yu J:
mTOR inhibitors induce apoptosis in colon cancer cells via
CHOP-dependent DR5 induction on 4E-BP1 dephosphorylation. Oncogene.
35:148–157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(Delta Delta C(T) method. Methods. 25:402–408.. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He K, Zheng X, Zhang L and Yu J: Hsp90
inhibitors promote p53-dependent apoptosis through PUMA and Bax.
Mol Cancer Ther. 12:2559–2568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Risinger AL, Giles FJ and Mooberry SL:
Microtubule dynamics as a target in oncology. Cancer Treat Rev.
35:255–261. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gershenzon J and Dudareva N: The function
of terpene natural products in the natural world. Nat Chem Biol.
3:408–414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fujioka T, Kashiwada Y, Kilkuskie RE,
Cosentino LM, Ballas LM, Jiang JB, Janzen WP, Chen IS and Lee KH:
Anti-AIDS agents, 11. Betulinic acid and platanic acid as anti-HIV
principles from syzigium claviflorum, and the anti-HIV activity of
structurally related triterpenoids. J Nat Prod. 57:243–247. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi Y: Mechanisms of caspase activation
and inhibition during apoptosis. Mol Cell. 9:459–470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Riedl SJ and Salvesen GS: The apoptosome:
Signalling platform of cell death. Nat Rev Mol Cell Biol.
8:405–413. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y, He K, Huang Y, Zheng D, Gao C, Cui L
and Jin YH: Betulin induces mitochondrial cytochrome c release
associated apoptosis in human cancer cells. Mol Carcinog.
49:630–640. 2010.PubMed/NCBI
|
|
27
|
Chen L, Willis SN, Wei A, Smith BJ,
Fletcher JI, Hinds MG, Colman PM, Day CL, Adams JM and Huang DC:
Differential targeting of prosurvival Bcl-2 proteins by their
BH3-only ligands allows complementary apoptotic function. Mol Cell.
17:393–403. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kuwana T, Bouchier-Hayes L, Chipuk JE,
Bonzon C, Sullivan BA, Green DR and Newmeyer DD: BH3 domains of
BH3-only proteins differentially regulate Bax-mediated
mitochondrial membrane permeabilization both directly and
indirectly. Mol Cell. 17:525–535. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Strasser A: The role of BH3-only proteins
in the immune system. Nat Rev Immunol. 5:189–200. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chipuk JE, Bouchier-Hayes L and Green DR:
Mitochondrial outer membrane permeabilization during apoptosis: The
innocent bystander scenario. Cell Death Differ. 13:1396–1402. 2006.
View Article : Google Scholar : PubMed/NCBI
|