Introduction
Esophageal cancer (EC) accounts for between 1 and 3%
of all incidences of cancer occurring in the USA, but exhibits a
higher incidence in Asia (1). In
China, esophageal squamous cell carcinoma (ESCC) is the predominant
histological subtype accounting for ~90% of all EC cases (2). Despite progress in the application of
combined chemotherapy and radiotherapy, ESCC remains an intractable
cancer. Thus, the development of novel antitumor agents to improve
the prognosis of patients with ESCC is urgently required.
Histone acetylation status is controlled by histone
acetyltransferases (HATs) and histone deacetylases (HDACs), which
are associated with chromatin structure and influence gene
transcription (3–5). HATs direct the addition of acetyl groups
to lysine residues on histones and regulate gene expression. HDACs
remove acetyl groups from histones, silencing the transcription of
several oncogenes and tumor suppressor genes. HDAC inhibitors have
been revealed to serve a therapeutic role in numerous distinct
human cancer cell lines (6). FK228, a
novel HDAC inhibitor, has been demonstrated to exhibit antitumor
activity in several cancer cells, alone or in combination with
other drugs or therapeutic methods (7,8) and was
approved by the US Food and Drug Administration in 2009 (9) for use in patients with cutaneous T-cell
lymphoma with a Phase II study currently in progress in Italy
(10). The molecular mechanisms
underlying the antitumor effects of FK228 including apoptosis,
growth arrest and angiogenesis inhibition have been reported, and
gene expression profiling has been performed to evaluate the
effects of FK228 (11–14). However, the effects of FK228 on the
global levels of histone acetylation and the proteome of EC109
cells remain poorly understood.
The present study aimed at employing integrated
approaches including the stable isotope labeling by amino acids in
cell culture (SILAC) technique and mass spectrometry-based
quantitative proteomics to quantify the dynamic changes in histone
lysine acetylation and in the proteome in one pair of EC109
cells.
Materials and methods
Cell culture
The ESCC cell line EC109 was acquired from Shanghai
Cell Bank (Shanghai, China) and was maintained in Dulbecco's
modified Eagle's medium (DMEM; Pierce; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified atmosphere containing 5% CO2.
(13C)Lysine labeling and
FK228 treatment of EC109 cells
EC109 cells were grown to 80% confluence in high
glucose (4.5 g/l) DMEM (with glutamine and sodium pyruvate)
containing 10% FBS and 1% penicillin-streptomycin at 37°C with 95%
air and 5%CO2. The cells were subsequently labeled with
either ‘heavy isotopic lysine’ [(13C)lysine,
(13C6,15N4)arginine] or
‘light isotopic lysine’ [(12C)lysine] for >6
generations prior to being harvested to achieve >97% labeling
efficiency by using a SILAC Protein Quantitation kit (Pierce;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Cells were further cultured in SILAC medium to obtain the
desired cell number (~5×108) in 15,150-cm2
flasks.
The ‘light’ labeled cells were treated with 43.5
ng/ml FK228, and the ‘heavy’ labeled cells were treated with the
equivalent measure of dimethyl sulfoxide. Following treatment, the
cells were maintained in SILAC medium for 24 h before being
harvested and washed twice with ice-cold PBS supplemented with 2 µM
trichostatin A and 30 mM nicotinamide. Following snap-freezing in
liquid nitrogen, the cell pellets were obtained and stored in a
−80°C freezer for future use.
Histone extraction and trypsin
digestion
The harvested ‘heavy’ and ‘light’ labeled cells were
lysed using 2X NETN buffer (200 mM NaCl, 2 mM EDTA, 100 mM Tris-HCl
and 1.0% NP-40; pH7.2) supplemented with 0.5% Triton X-100 on ice
for 30 min. The supernatants were retained following centrifugation
at 20,000 g for 10 min at 4°C. Next, equal amounts of crude
proteins were mixed in the supernatants labeled ‘heavy’ or ‘light’,
and the crude proteins were then precipitated by adding
trifluoroacetic acid (TFA) at a final concentration of 15% (v/v)
(soluble fraction). The protein pellets were washed twice with 20°C
acetone prior to being dissolved in 100 mM
NH4HCO3 (pH 8.0) for trypsin digestion. The
remaining cell pellets were dissolved in 8 M urea to extract the
chromatin-binding proteins. The protein concentration was measured
using a bicinchoninic acid assay kit (Thermo Fisher Scientific,
Inc.) and equal amounts of chromatin-binding proteins were added to
the urea solution and the proteins were precipitated by adding TFA
at a final concentration of 15% (v/v) (nuclear pellet fraction).
The protein pellets were washed twice with −20°C acetone prior to
being dissolved in 100 mM NH4HCO3 for trypsin
digestion.
Trypsin (Promega Corporation, Madison, WI, USA) was
added to the protein solution at a trypsin to protein ratio of 1:50
(w/w) for digestion at 37°C for 16 h. DTT was added to obtain a
final concentration of 5 mM prior to incubation at 50°C for 30 min.
Iodoacetic acid was subsequently added to alkylate the proteins and
achieve a final concentration of 15 mM; this was then followed by
incubation at room temperature in the dark for 30 min. The
alkylation reaction was quenched by adding 30 mM cysteine (final
concentration) at room temperature for another 30 min. Trypsin was
then added again at a trypsin to protein ratio of 1:100 (w/w) for
digestion at 37°C for 3 h to complete the digestion cycle.
Western blot
Total proteins (30 µg) were isolated from FK228
treated EC109 cells and were separated using SDS-PAGE (15% gel),
and then transferred to nitrocellulose membranes. After being
blocked for 1 h with the Tris/NaCl containing 5% milk powder, the
membranes were probed using an acetylated lysine antibody in a
1:1,000 dilution overnight at 4°C (cat. no., ab80178; PTM Biolabs,
Hangzhou, China). Secondary horseradish peroxidase-labeled
anti-goat/rabbit immunoglobulin G (cat. no., 111-035-006; Zhongshan
Corp., Beijing, China) was used in a 1:10,000 dilution for 2 h at
room temperature. The protein expression of each sample was
normalized by that of GAPDH with Quantity One software (version
4.6.9; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Affinity enrichment of histone
lysine-acetylated peptides
Affinity enrichment was performed for the histone
peptides. To enrich the lysine-acetylated peptides, the tryptic
peptides were divided into three groups and dissolved in NETN
buffer (100 mM NaCl, 1 mM EDTA, 50 mM Tris-HCl and 0.5% NP-40, pH
8.0) separately. The peptides were then incubated with pre-washed
antibody beads (PTM Biolabs, Hangzhou, China) at 4°C overnight with
gentle shaking. Next, the beads were washed four times with NETN
buffer and twice with double-distilled water. The bound peptides
were eluted from the beads using 0.1% TFA and the eluted fractions
were combined and vacuum-dried. The resulting peptides were cleaned
using C18ZipTips (EMD Millipore, Billerica, MA, USA),
according to the manufacturer's protocol, prior to liquid
chromatography tandem-mass spectrometry (LC-MS/MS) analysis.
LC-MS/MS analysis
The peptides were dissolved in 0.1% formic acid and
loaded directly onto a reversed-phase pre-column (Acclaim PepMap
100; Thermo Fisher Scientific, Inc.). Peptide separation was
performed using a reversed-phase analytical column (Acclaim PepMap
RSLC; Thermo Fisher Scientific, Inc.) with a linear gradient of
5–35% solvent B (0.1% TFA in 98% acetonitrile) for 30 min and
35–80% solvent B for 10 min at a constant flow rate of 300 nl/min
on an EASY-nLC 1000 UPLC system (Thermo Fisher Scientific, Inc.).
The resulting peptides were analyzed using a Q Exactive™ Plus mass
spectrometer (Thermo Fisher Scientific, Inc.). Subsequently, the
peptides were subjected to a nanospray ionization source, followed
by tandem mass spectrometry (MS/MS) in Q Exactive (Thermo Fisher
Scientific, Inc.) coupled online to the UPLC. Intact peptides were
detected in the Orbitrap at a resolution of 70,000. The peptides
were selected for MS/MS using 27% normalized collision energy (NCE)
with 12% stepped NCE; ion fragments were detected in the Orbitrap
at a resolution of 17,500. A data-dependent procedure that
alternated between one MS scan followed by 20 MS/MS scans was
applied for the top 20 precursor ions above a threshold ion count
of 3×104 in the MS survey scan with 15.0 sec dynamic
exclusion. The electrospray voltage applied was 1.8 kV. Automatic
gain control was used to prevent overfilling of the ion trap;
1×105 ions were accumulated for generation of the MS/MS
spectra. For the MS scans, the m/z scan range was
between 350 and 1,600 Da.
Database search
The resulting MS/MS data were processed using
MaxQuant with integrated Andromeda search engine (version 1.4.1.2;
www.maxquant.org). Tandem mass spectra were
searched against the UniProt/Swiss-Protdatabase concatenated with a
reverse decoy database. Trypsin/P was specified as the cleavage
enzyme, allowing up to two missing cleavages, four modifications
per peptide and five charges. The mass error was set to 10 ppm for
precursor ions and 0.02 Da for fragment ions. Carbamidomethylation
on cysteine residues was specified as a fixed modification, and
oxidation on methionine residues and acetylation on the N-termini
of the proteins were specified as variable modifications. False
discovery rate thresholds for the protein, peptide and modification
site were specified at 1%. The minimum peptide length was set at 6.
All the other parameters in MaxQuant were set to default
values.
Functional enrichment analysis
To further explore the effect of the differentially
expressed proteins on the physiological processes in the cells and
to identify the internal associations between the differentially
expressed proteins, the functions of the differentially expressed
proteins were classified and the significance of functional
enrichment of Gene Ontology (GO) terms (biological process,
cellular component, molecular function), domains and the Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway were analyzed.
Protein annotation, classification and
subcellular location prediction
The GO annotation proteome was derived from the
UniProt-GOA database (www.ebi.ac.uk/GOA). First, the identifier (ID) of the
identified protein was converted into a UniProt ID and then mapped
to GO IDs by the protein ID. If identified proteins were not
annotated by the UniProt-GOA database, InterProScansoft would be
used to annotate the protein's GO function using the protein
sequence alignment method. The proteins were then classified by
their GO annotation based on three categories: Biological process,
cellular component and molecular function. The KEGG database was
used to annotate the protein pathways. First, the KEGG online
service tool, KAAS, was used to annotate the protein's KEGG
database description. The annotation result was then mapped on the
KEGG pathway database using the KEGG online service tool, KEGG
mapper. Next, WoLFPSORT (https://wolfpsort.hgc.jp/), a subcellular localization
predication software, was used to predict the subcellular
localization of the proteins. A two-tailed Fisher's exact test was
used to statistically analyze the difference among members of
protein clusters in specific annotation terms. P<0.05 was
considered to indicate a statistically significant difference.
Results and Discussion
FK228 increased global histone lysine
acetylation in the EC109 cells
The viability of the EC109 cells decreased upon
treatment with increasing doses of FK228. The cells were maintained
in SILAC medium for 24 h for the lysine acetylation experiments.
The global lysine acetylome was detected using western blotting
(data not shown).
Next, site-specific identification of histone lysine
acetylation in the EC109 cells was carried out. SILAC-based
quantitative proteomics was carried out to profile histone lysine
acetylation with or without FK228 treatment. A total of 87 lysine
acetylation (Kac) sites from the histones and HATs were identified
in the present study, of which 25 sites were quantifiable and 19
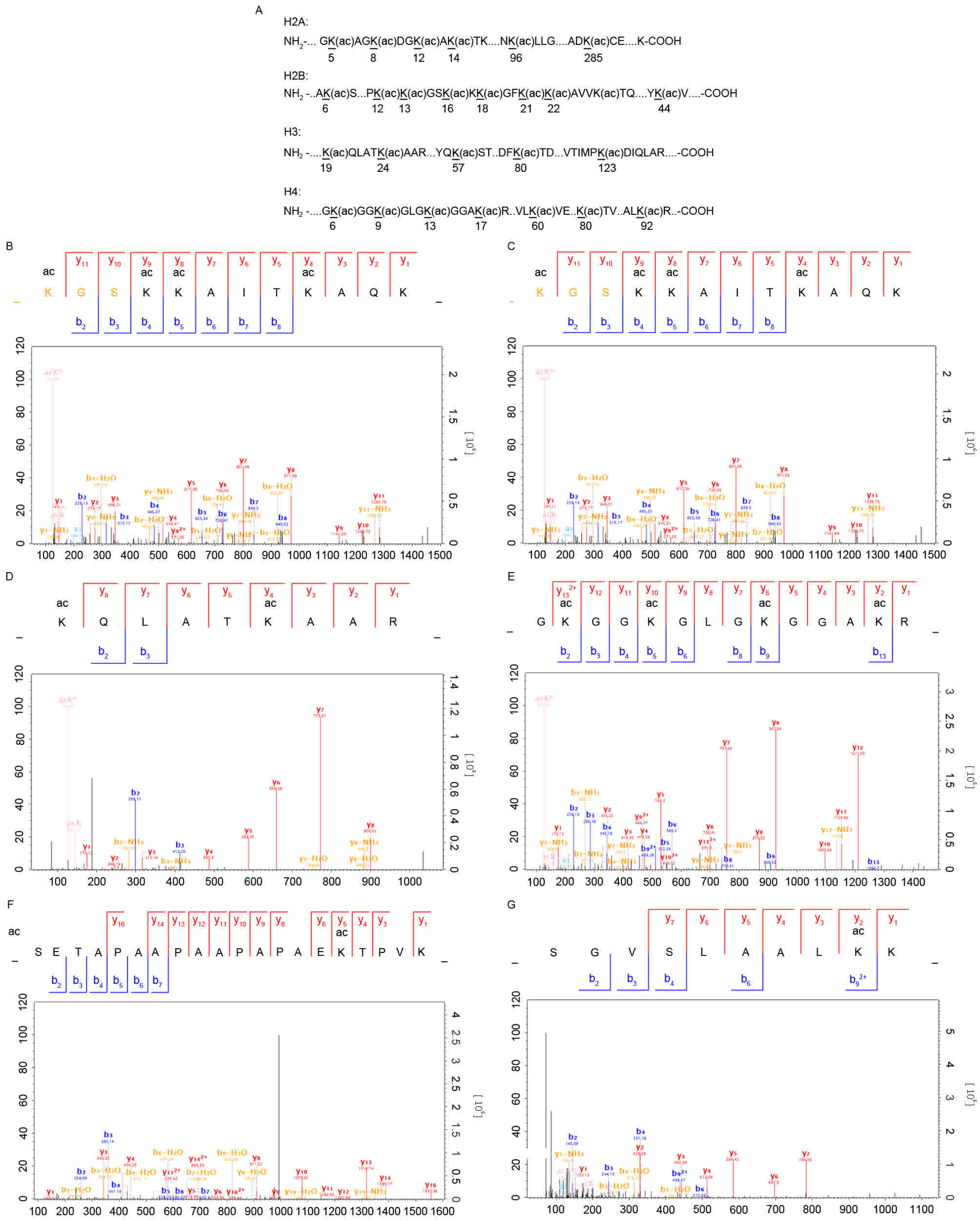
were quantified with a ratio of >1.3 (Fig. 1A). The representative spectra of the
histone lysine-acetylated peptides are shown in Fig. 1B-F, including the spectra for the
H3K24ac, H2K14ac, H2K17ac and H2K22ac peptides. Notably, no
downregulated Kac sites were quantified in the present study.
Furthermore, the 62 unquantified Kac sites were only identified in
the FK228-treated cells. The distinct profiles of histone lysine
acetylation following FK228 treatment revealed the effects of FK228
on the EC109 cells. The sequences of the identified
lysine-acetylated peptides in the core histones and the
corresponding quantitative changes in the Kac profiles in response
to FK228 treatment are summarized in Table I.
 | Table I.Summary of the identified lysine
acetylation sites and the quantifiable changes in the EC109 cells
treated with FK228. |
Table I.
Summary of the identified lysine
acetylation sites and the quantifiable changes in the EC109 cells
treated with FK228.
| Modified site | Modified sequence of
peptide | Quantifiable fold
change |
|---|
| H1K17ac | _APAEK(ac)TP_ | 2.92 |
| H2AK96ac | _EELNK(ac)LLG_ | 1.36 |
| H2AK285ac | _WGADK(ac)CEE_ | 1.39 |
| H2BK12ac |
_PAPK(ac)K(ac)GS_ | 130.67 |
| H2BK13ac |
_PAPK(ac)K(ac)GS_ | 130.67 |
| H2BK16ac |
_GSK(ac)K(ac)AVTK_ | 130.67 |
| H2BK17ac |
_GSK(ac)K(ac)AVTK_ | 130.67 |
| H2BK18ac |
_K(ac)GSK(ac)K(ac)AV_ | 130.67 |
| H2BK21ac | _ K(ac)K(ac)AVTK(ac)
_ | 130.67 |
| H2BK22ac | _K(ac)K(ac)AVTK(ac)
_ | 130.67 |
| H2BK44ac | _YSVYVYK(ac)VLK_ | 1.51 |
| H2BK109ac | _GELAK(ac)HAVS_ | 1.39 |
| H3K24ac | _QLATK(ac)AAR_ | 5.94 |
| H3K57ac | _QK(ac)STELLIR_ | 1.49 |
| H3K80ac | _AQDFK(ac)TDR_ | 1.14 |
| H3K123ac | _PK(ac)DIQLAR_ | 1.4 |
| H4K32ac | _IQGITK(ac)PAIR_ | 1.23 |
| H4K60ac | _GVLK(ac)VFLEN_ | 1.19 |
| H4K80ac | _K(ac)TVTAMD_ | 1.59 |
| H4K92ac | _MDVVYALK(ac)R_ | 1.18 |
The abundance of the identified Kac sites in the
EC109 cells was compared. The relative abundance was calculated
based on the ratio of the peak areas of the same parent ions
labeled by the light and heavy isotopes in the MS spectra. Among
these Kac sites, H2BK12ac, H2BK13ac, H2BK16ac, H2BK17ac, H2BK18ac
H2BK21ac and H2BK22ac exhibited significant differential abundance
sites, with a 130-fold difference between the FK228-treated and
-untreated cells. To the best of our knowledge, the present study
is the first to identify these Kac sites in EC109 cells and their
functions require further investigation in future studies. In the
acetylation site of H3K24, its abundance in the EC109 cells exposed
to FK228 was increased 5.9-fold compared with that observed in the
EC109 cells that were not treated with FK228. The acetylation level
in H1K17ac was also increased 2.92-fold in the EC109 cells
following FK228 treatment. These results provide a comprehensive
identification of sites of histone lysine acetylation and reveal
the potential epigenetic mechanism underlying the effects of FK228
in the EC109 cells. The results of the present study suggest that
FK228-based therapy may have potential for use in treating
ESCC.
FK228, one of the powerful HDAC inhibitors, inhibits
class I HDAC enzymes more than class II HDAC enzymes. It inhibits
the removal of acetyl groups from the lysine residues of N-terminal
histone tails (15) and also alters
the acetylation of other nuclear and cytoplasmic proteins including
heat-shock protein 90 chaperone proteins (11). Histone acetylation states are crucial
to a number of cellular processes; for example, the cycles of
acetylation and deacetylation may prepare the genes including
oncogenes and tumor suppressor genes for future activation. The
present study observed the distinct effects of FK228 on histone
lysine acetylation in the EC109 cells. Almost all the quantifiable
lysine-acetylated sites were in the N-termini of core histones,
suggesting the profound epigenetic modulation mechanism underlying
the effect FK228 in the EC109 cells. Among the quantifiable
lysine-acetylated sites, 22 sites were novel acetylated sites and
seven sites significantly increased their acetylation level in
response to FK228 exposure. Further studies should focus on the
lysine-acetylated sites that demonstrated significant alterations
in histones to explore the epigenetic mechanism underlying the
effect of FK228 in the EC109 cells.
Proteome profile of the EC109 cells
treated with FK228
HATs, which acetylate the lysine residues of histone
proteins, facilitate the access of numerous transcriptional factors
to the DNA and consequently activate the expression of the target
genes. Therefore, the increased or decreased level of histone
lysine acetylation may correspondingly alter the proteome of the
EC109 cells. To confirm this, quantitative proteomics was performed
to profile the proteome of the EC109 cells with or without FK228
exposure. A total of 5,279 proteins were identified in the EC109
cells, of which 3,515 proteins were quantified. Of these 3,515
quantifiable proteins, 675 proteins demonstrated increased histone
lysine acetylation levels and 186 proteins demonstrated decreased
levels, which suggested that the distinct profiles of histone
lysine acetylation induced by FK228 treatment may lead to distinct
proteomic profiles.
Biological functions of the quantified
proteins in the EC109 cells treated with FK228
To gain a better understanding of the function and
characteristics of the quantified proteins, the functions or
features of the proteins from several distinct categories,
including GO terms, domains, pathways and subcellular localization
were noted. The proteins were classified by GO annotation on the
basis of three categories: Biological process, cellular component
and molecular function. In the biological process category
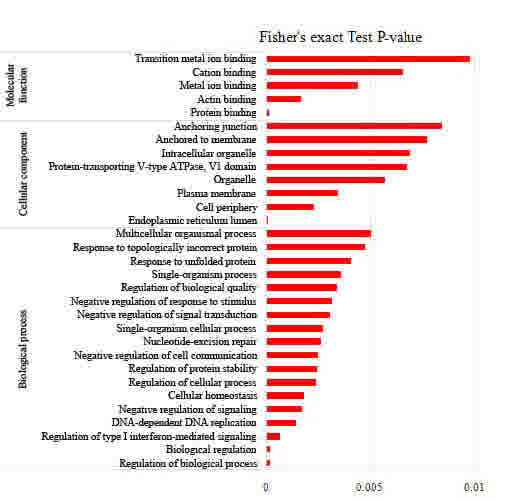
(Fig. 2), the proteins associated
with the regulation of biological processes, biological regulation,
regulation of the type I interferon-mediated signaling pathway,
regulation of protein stability, regulation of cellular processes
and negative regulation of signal transduction were upregulated,
whereas those associated with cellular hormone metabolism, hormone
metabolism, carboxylic acid metabolism, lipid metabolism, oxoacid
metabolism, steroid metabolism, hormone biosynthesis and organic
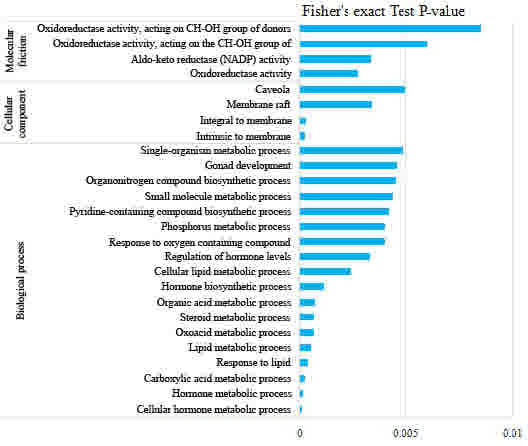
acid metabolism were downregulated (Fig.
3). These results implied that FK228 treatment affected
biological regulation and hormone metabolism in the EC109
cells.
Enrichment analysis of the molecular functions
revealed that proteins associated with protein binding, actin
binding, metal ion binding and cation binding were upregulated
(Fig. 2), whereas those associated
with oxidoreductase activity and aldo-ketoreductase
(nicotinamide-adenine dinucleotide phosphate) activity were
downregulated (Fig. 3). These results
suggested that the proteins involved in binding functions, which
may regulate enzyme activity, were enriched following FK228
exposure. Analysis by cellular component revealed that the
endoplasmic reticulum lumen was the major cellular component in the
EC109 cells that responded to the acetylation changes. Therefore,
the GO analysis suggested that the enriched biological functions
were associated with lysine acetylation, implying an
HDAC-inhibiting role of FK228 in the EC109 cells.
Domain structure is a major functional
element in proteins
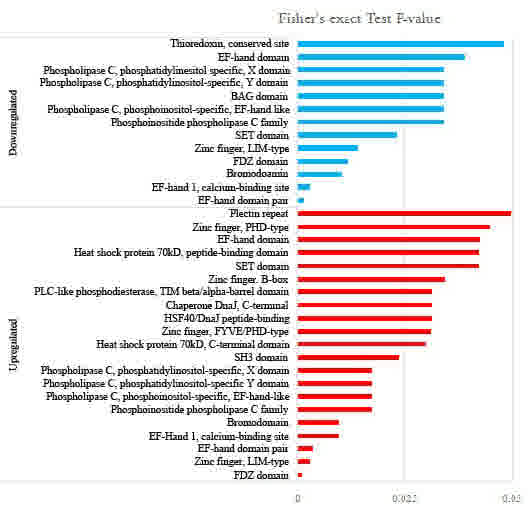
To investigate the domain features of the enriched
proteins, the proteins that were enriched upon FK228 treatment were
analyzed using Interpro domain enrichment analysis. The results
revealed that protein domains including the PDZ domain, zinc finger
domain and EF hand domain were enriched following FK228 treatment
(Fig. 4).
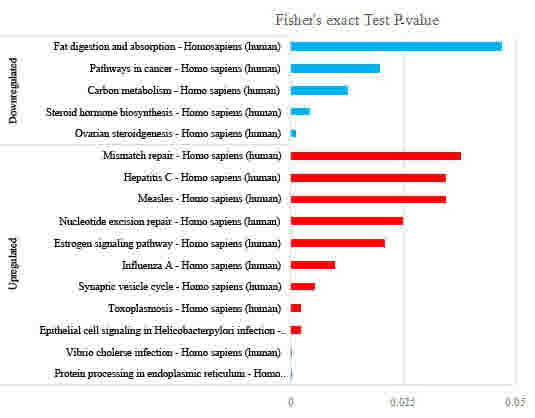
Next, pathway clustering analysis of the
FK228-responsive proteome from the KEGG database was performed to
examine the cellular pathways affected by FK228 treatment (Fig. 5). The results revealed that protein
processing in the endoplasmic reticulum, Vibrio cholerae
infection, epithelial cell signaling in Helicobacter pylori
infection and toxoplasmosis were the most distinct signaling
pathways exhibiting increased protein levels in the FK228-treated
cells. This suggested a role for FK228 in these signaling pathways.
However, the protein level in the signaling pathway of ovarian
steroidogenesis and steroid hormone biosynthesis was decreased in
the FK228-treated cells. These results were in agreement with those
observed in biological process regulation, where the hormone
metabolism signaling pathway and others were downregulated.
In the present study, quantitative proteomics and
bioinformatic analysis were used to characterize the global profile
of histone lysine acetylation and the quantitative proteomic
changes in EC109 cells treated with FK228. A number of Kac sites
including novel acetylation sites were identified in the EC109
cells. As histone lysine acetylation regulates the activation of
gene transcription by regulating the binding of histone proteins to
DNA, the different levels of histone lysine acetylation observed in
the present study may affect the expression profile of the
proteome. Bioinformatic analysis further revealed that FK228
regulates protein expression by affecting multiple biological
signaling pathways and protein complexes in the EC109 cells. The
results of the present study improve current knowledge of the
therapeutic properties and the molecular mechanism underlying the
effects of FK228 as an HDAC inhibitor in EC109 cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Zhejiang Cancer Hospital (grant no. 2013Y012) and the Medical and
the Health Science Project of Zhejiang Province (grant no.
2015KYA033).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZP analyzed and interpreted the data, and was a
major contributor in writing the manuscript. MW performed
experiments and contributed to data analysis. ZY performed
experiments and contributed materials and analysis tools. SZ
contributed significantly to data analysis and manuscript
preparation. XX was responsible for conception of the study, and
manuscript revision. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of
interest.
Glossary
Abbreviations
Abbreviations:
|
ESCC
|
esophageal squamous cell carcinoma
|
|
HATs
|
histone acetyltransferases
|
|
HDACs
|
histone deacetylases
|
|
FBS
|
fetal bovine serum
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
GO
|
Gene Ontology
|
References
|
1
|
Layke JC and Lopez PP: Esophageal cancer:
A review and update. Am Fam Physician. 73:2187–2194.
2006.PubMed/NCBI
|
|
2
|
Qi YJ, Chao WX and Chiu JF: An overview of
esophageal squamous cell carcinoma proteomics. J proteomics.
75:3129–3137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Emanuele S, Lauricella M and Tesoriere G:
Histone deacetylase inhibitors: Apoptotic effects and clinical
implications (Review). Int J Oncol. 33:637–646. 2008.PubMed/NCBI
|
|
4
|
Glaser KB: HDAC inhibitors: Clinical
update and mechanism-based potential. Biochem Pharmacol.
74:659–671. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fukuda H, Sano N, Muto S and Horikoshi M:
Simple histone acetylation plays a complex role in the regulation
of gene expression. Brief Funct Genomic Proteomic. 5:190–208. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marks PA, Richon VM, Breslow R and Rifkind
RA: Histone deacetylase inhibitors as new cancer drugs. Curr Opin
Oncol. 13:477–483. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jain S, Jirau-Serrano X, Zullo KM, Scotto
L, Palermo CF, Sastra SA, Olive KP, Cremers S, Thomas T, Wei Y, et
al: Preclinical pharmacologic evaluation of pralatrexate and
romidepsin confirms potent synergy of the combination in a murine
model of human T-cell lymphoma. Clin Cancer Res. 21:2096–2106.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stühmer T, Arts J, Chatterjee M, Borawski
J, Wolff A, King P, Einsele H, Leo E and Bargou RC: Preclinical
anti-myeloma activity of the novel HDAC-inhibitor JNJ-26481585. Br
J Haematol. 149:529–536. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
VanderMolen KM, McCulloch W, Pearce CJ and
Oberlies NH: Romidepsin (Istodax, NSC 630176, FR901228, FK228,
depsipeptide): A natural product recently approved for cutaneous
T-cell lymphoma. J Antibiot (Tokyo). 64:525–531. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pellegrini C, Dodero A, Chiappella A,
Monaco F, Degl'Innocenti D, Salvi F, Vitolo U, Argnani L, Corradini
P and Zinzani PL: Italian Lymphoma Foundation (Fondazione Italiana
Linfomi Onlus, FIL): A phase II study on the role of gemcitabine
plus romidepsin (GEMRO regimen) in the treatment of
relapsed/refractory peripheral T-cell lymphoma patients. J Hematol
Oncol. 9:382016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu X, Guo ZS, Marcu MG, Neckers L, Nguyen
DM, Chen GA and Schrump DS: Modulation of p53, ErbB1, ErbB2, and
Raf-1 expression in lung cancer cells by depsipeptide FR901228. J
Natl Cancer Inst. 94:504–513. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Archer SY, Meng S, Shei A and Hodin RA:
p21(WAF1) is required for butyrate-mediated growth inhibition of
human colon cancer cells. Proc Natl Acad Sci USA. 95:6791–6796.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kwon HJ, Kim MS, Kim MJ, Nakajima H and
Kim KW: Histone deacetylase inhibitor FK228 inhibits tumor
angiogenesis. Int J Cancer. 97:290–296. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Insinga A, Monestiroli S, Ronzoni S,
Gelmetti V, Marchesi F, Viale A, Altucci L, Nervi C, Minucci S and
Pelicci PG: Inhibitors of histone deacetylases induce
tumor-selective apoptosis through activation of the death receptor
pathway. Nat Med. 11:71–76. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuo MH and Allis CD: Roles of histone
acetyltransferases and deacetylases in gene regulation. Bioessays.
20:615–626. 1998. View Article : Google Scholar : PubMed/NCBI
|