Introduction
Dendritic cells (DCs) are the most important
antigen-presenting cells in the body; DC-based cancer immunotherapy
has been investigated in previous years (1–4).
Additionally, the approval of the first DC vaccine, Provenge
(generic name, sipuleucel-T) (5), by
the US Food and Drug Administration for the treatment of prostate
cancer in 2010 was a milestone in the immunotherapeutic application
of DC vaccines. Whole-tumor cell lysates (TCLs) have been used as
the source of tumor antigens for the development of DC vaccines;
however, the clinical outcomes of TCL-based DC vaccines have been
unsatisfactory owing to the weak immunogenicity of tumor cells
(6). To improve the efficacy of
TCL-based DC vaccines, viruses have been used to enhance the
immunity of TCLs, to further enhance the antigen delivery and to
increase the antigen-presenting ability of DCs (7).
The Newcastle Disease Virus (NDV) is a bird RNA
virus of the Paramyxovirus family. Specifically, NDV belongs
to the genus Avulavirus, which does not include any known
natural pathogens of humans (8). Over
the last 6 decades, NDV has been tested as an anticancer agent
because of its oncolytic properties. Owing to the oncolytic and
immunostimulatory properties of NDV, necrotic tumor cells destroyed
by the virus are phagocytosed and perceived as dangerous by
antigen-presenting cells. These professional antigen-presenting
cells then process tumor-associated antigens (TAA), become
activated and present the processed TAA peptides to T cells for
cognate interaction and the induction of an immune response. Owing
to NDV's properties of tumor-selective replication, oncolytic
capacity and immune stimulation (9),
it is considered a promising candidate for use in enhancing the
efficacy of DC vaccines (10–12). Virus-induced augmentation of the
antigenicity of tumor antigens had been observed in several model
systems (13,14).
The aim of the present study was to determine
whether DCs loaded with NDV-infected TCL (NDV-TCL) possessed a
stronger antitumor ability than DCs loaded only with TCL. The
results of the present study indicate the presence of a potential
novel method for increasing the efficacy of DC vaccines and for the
development of novel DC vaccines for immunotherapy in the
future.
Materials and methods
Patient information
In the present study, 12 lung cancer patients were
recruited from The First Hospital of Jilin University (Changchun,
China) from August 2015 to December 2015. The age range was 45–68
years old and the mean age was 56 years old. There were eight
female and four male patients. The inclusion criteria were as
follows: No radiation and chemotherapy for more than 1 month prior
to blood collection. Other clinical data of the patients have been
listed in Table I. The present study
was reviewed and approved by the ethics committee of The First
Hospital of Jilin University, with written informed consent
provided by all participants.
 | Table I.Characteristics of the 12 lung cancer
patients. |
Table I.
Characteristics of the 12 lung cancer
patients.
| No. | Sex | Age, years | Diagnosis | Stage |
|---|
| 1 | F | 50 | Lung cancer
(adenocarcinoma) | T2N3M0, IIIB |
| 2 | F | 60 | Lung cancer
(adenocarcinoma) | T3N3M1, IV |
| 3 | F | 49 | Lung cancer
(adenocarcinoma) | T4N2M1, IV |
| 4 | F | 62 | Lung cancer
(adenocarcinoma) | T1N3M0, IIIB |
| 5 | F | 46 | Lung cancer
(squamous carcinoma) | T4N2M0, IIA |
| 6 | M | 62 | Lung cancer
(squamous carcinoma) | T2N1M0, IIIA |
| 7 | M | 58 | Lung cancer (small
cell) | Limited stage |
| 8 | M | 45 | Lung cancer (small
cell) | Limited stage |
| 9 | M | 60 | Lung cancer (small
cell) | Limited stage |
| 10 | F | 64 | Lung cancer (small
cell) | Extensive
stage |
| 11 | F | 48 | Lung cancer (small
cell) | Extensive
stage |
| 12 | F | 68 | Lung cancer (small
cell) | Extensive
stage |
Generation of DCs
Peripheral blood mononuclear cells (PBMCs) were
isolated by density gradient centrifugation (400 × g at room
temperature for 30 min) using Ficoll medium (Takeda Pharmaceuticals
International GmbH, Zurich, Switzerland) from whole blood samples
(50 ml) of the 12 lung cancer patients. DCs were induced from
adherent cells derived from PBMCs following 2 h of incubation at
37°C in a CO2 incubator. Non-adherent cells were
collected and used as T cells for the cytokine secretion studies.
The adherent cells were cultured in CellGenix GMP DC (CellGenix,
Freiburg, Germany) with 50 ng/ml interleukin-4 (IL-4) and 50 ng/ml
recombinant human granulocyte macrophage-colony stimulating factor
(GM-CSF) both (Miltenyi Biotec, GmbH Bergisch Gladbach, Germany)
for 5 days, and the same concentration of cytokines (IL-4 and
GM-CSF) was added on day 2. On day 5, 1 µg/ml lipopolysaccharide
(LPS; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added into
the culture medium. On day 7, the cultured cells were harvested and
stained with cluster of differentiation (CD)14-FITC (cat. no.
347493), human leukocyte antigen-antigen D related (HLA-DR)-PerCP
(cat. no. 347364) (both from BD Biosciences, San Jose, CA, USA),
CD1a-PE (cat. no. 555807), CD80-PE (cat. no. 340294), CD86-FITC
(cat. no. 555657) and CD83-APC (cat. no. 551073) (all from BD
Pharmingen; BD Biosciences) at room temperature for 15 min. At the
same time, the cells were stained with mouse immunoglobulin G
(IgG)2b κ-FITC (cat. no. 555742), mouse IgG1 κ-PE (cat. no.
555749), mouse IgG2a κ-PerCP (553933) and mouse IgG1κ-APC (cat. no.
555751) (all from BD Biosciences) at room temperature for 15 min.
These antibodies were used as control isotype-matched antibodies.
All antibodies used above were ready for use and did not require
dilution. The surface phenotype of the DCs was determined using
FACSCalibur flow cytometer (BD Biosciences). The data were analyzed
by FlowJo V7.6 software (Tree Star, Inc., Ashland, OR, USA).
Giemsa staining of the DCs
To evaluate the morphology of DCs, DCs were
collected, washed with PBS once and seeded (2.4×105
cells/ml) onto a clean microscope slide. The cells were then
air-dried and stained with Giemsa for morphological examination by
light microscopy (magnification, ×1,000). Giemsa staining was
performed using the Diff-Quik kit (Nanjing Jiancheng Bioengineering
Institute, Nanjing, China), according to the manufacturer's
protocol.
NDV source
NDV J76 was kindly supplied by Yuanguo Li (Military
Veterinary Institute, AMMS, Key laboratory of Jilin Province for
Zoonosis Prevention and Control, Changchun, China). The working
stock of the virus was sterilized by passing it through a 0.22-µm
filter prior to using it to infect the tumor cells.
Production of TCL and NDV-TCL
For the preparation of the TCL, human lung
adenocarcinoma A549 cells were obtained from Beijing Beina
Chuanglian Biotechnology Institute (Beijing, China) and cultured in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.). The tumor cells
(1×106 cells/ml) were collected in a tube, washed with
PBS twice, resuspended in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) and then used to produce the TCL through five
cycles of freezing in liquid nitrogen and thawing at 37°C in a
water bath, followed by low-speed centrifugation (460 × g at room
temperature for 10 min) to remove subcellular particles. For
preparing the NDV-TCL, the tumor cells were first incubated for 1 h
at 37 in a CO2 incubator in the presence of NDV J76 (16
hemagglutinating units per 1×106 cells). The
NDV-infected tumor cells were then washed with PBS to remove the
unbound virus and further cultured for 48 h. The same approach as
that used for preparing the TCL was then adopted for the
preparation of the NDV-TCL. The protein concentrations of the TCL
and NDV-TCL were then determined using the Bicinchoninic Protein
Assay kit (Wuhan Boster Biological Technology, Ltd., Wuhan,
China).
Loading DCs with TCL and NDV-TCL
For preparation of the DCs loaded with TCL and
NDV-TCL, on day 7 following chemokine treatment, the cultured DCs
(1×106 cells/ml) were loaded with TCL (100 µg/ml) or
NDV-TCL (100 µg/ml); 1 µg/ml LPS was then added after 4 h. On day
8, the DCs were collected for the subsequent experiments. The
surface phenotype of the different DCs (DCs, TCL-DCs, NDV-TCL-DCs)
was then determined using a FACSCalibur flow cytometer (BD
Biosciences) following staining with HLA-DR-PerCP, CD80-PE,
CD86-FITC and CD83-APC (ready-to-use dilution, as aforementioned)
at room temperature for 15 min. Appropriate isotype-matched
antibodies were used as controls. The data were analyzed by FlowJo
V7.6 software.
Expression of programmed death ligand
1 (PD-L1) on the different DCs
PD-L1 expression on the different DCs was also
determined using a FACSCalibur flow cytometer with PD-L1-APC
(dilution: ready for use; cat. no. 329708; BioLegend, Inc., San
Diego, CA, USA) at room temperature for 15 min on day 8. Data were
analyzed using FlowJo 7.6 software.
T-cell proliferation assays
Allogenic T cells were obtained from one healthy
volunteer (male, 56 years old) and stained with 5 µM carboxy
fluorescein succinimidyl ester (Molecular Probes; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
labeled cells were then washed, suspended (1×106
cells/ml) and co-cultured with the allogenic DCs, TCL-DCs, or
NDV-TCL-DCs at a ratio of 5:1 in 200 µl RPMI-1640 medium with 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.), 1X non-essential amino
acid (Gibco; Thermo Fisher Scientific, Inc.), 2 mM L-glutamine
(Sigma-Aldrich; Merck KGaA), 55 nM β-mercaptoethanol
(Sigma-Aldrich; Merck KGaA), and 100 nM sodium pyruvate
(Sigma-Aldrich; Merck KGaA) in 96-well rounded bottom plates. After
5 days, the cells were collected, washed with PBS and incubated
with CD3-APC (dilution: ready for use; cat. no. 340440; BD
Biosciences) for 15 min at room temperature, and the proliferating
T cells were then characterized by flow cytometry, as
aforementioned.
Determination of intracellular
interferon (IFN)-γ and IL-2 levels
The T cells were co-cultured with autologous DCs,
TCL-DCs, or NDV-TCL-DCs at a ratio of 5:1 for 24 h. The secretion
of IFN-γ and IL-2 from the T cells was then determined using the BD
Cytofix/Cytoperm™ kit (BD Biosciences) according to the
manufacturer's protocol. In brief, the co-cultured cells were
harvested and adjusted to a concentration of 1×106
cells/ml in RPMI-1640 medium containing 10% FBS and incubated with
0.1% GolgiStop (BD Biosciences) for 4 h. Next, the cells were
stained with CD3-PerCP (cat. no. 347344), CD4-FITC (cat. no.
340133) or CD8-FITC (cat. no. 555366) (all BD Biosciences) at room
temperature for 15 min, followed by intracellular staining with
IFN-γ-APC (cat. no. 554702) or IL-2–PE (cat. no. 554428) or rat
IgG2b κ-PE (556925) or mouse IgG1 κ-APC (cat. no. 555751) (all from
BD Biosciences) at room temperature for 30 min to determine
intracellular cytokine levels. All the antibodies used above were
ready for use and did not require dilution.
Determination of soluble IFN-γ
levels
The T cells (1×106 cells/ml) were
co-cultured with autologous DCs, TCL-DCs, or NDV-TCL-DCs at a ratio
of 5:1 at 37°C for 24 h in a humidified atmosphere of 5%
CO2. The level of IFN-γ in the cell culture supernatants
was then quantified using the cytometric bead array human IFN-γ
Flex set kit (BD Biosciences) on the FACSCalibur flow cytometer,
according to the manufacturer's protocol. The data were analyzed by
FlowJo V7.6 software
Statistical analysis
The data were analyzed using non-parametric
Friedman's 2-way analysis of variance by ranks test, followed by
pairwise comparisons, the statistical comparison method in SPSS 23
software, and the P-values were adjusted. All statistical analyses
were performed using IBM SPSS 23 software (IBM Corp., Armonk, NY,
USA). P<0.05 was considered to indicate a statistically
significant difference. Each experiment was repeated in
triplicate.
Results
Characteristics of the DCs
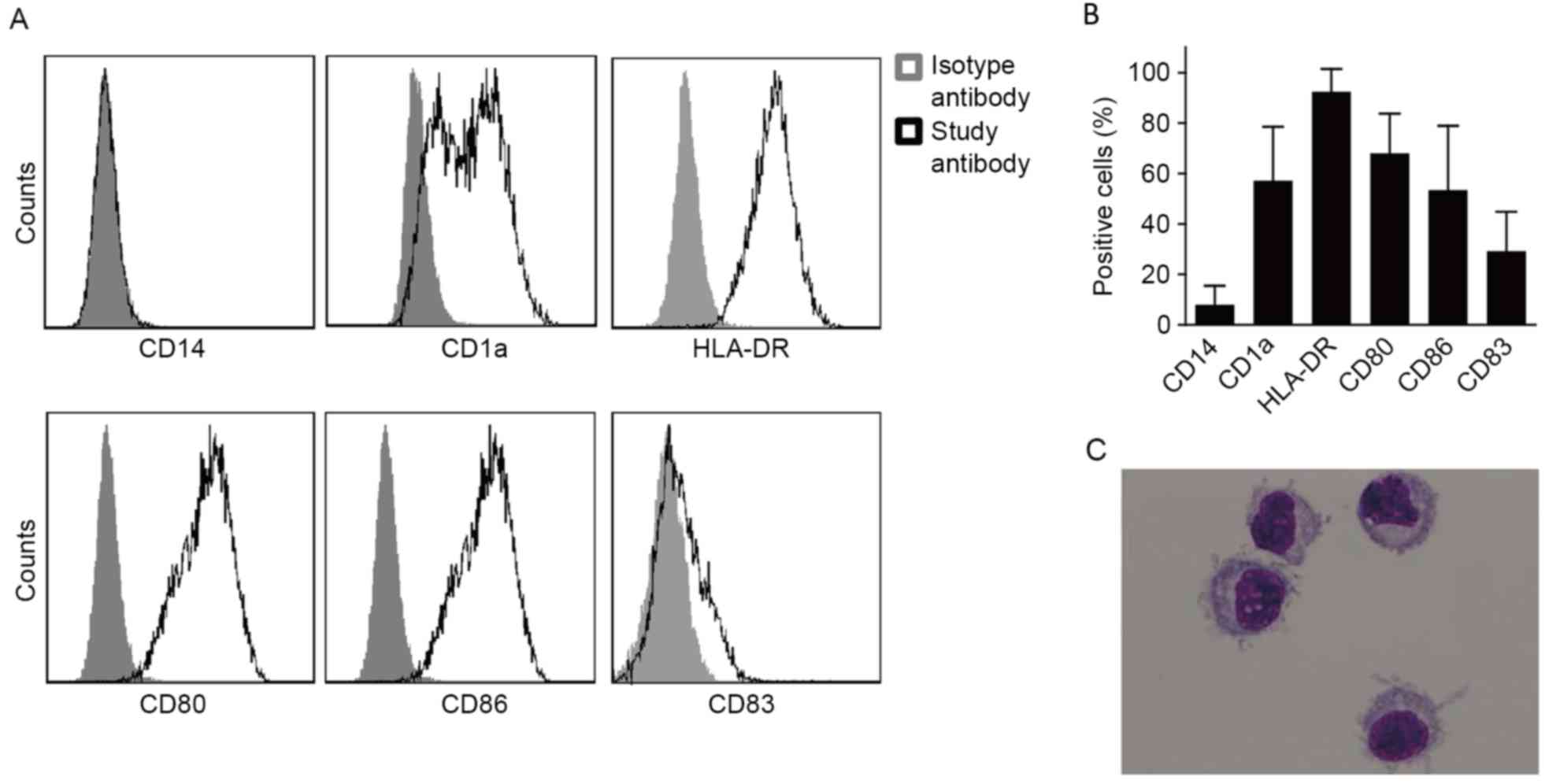
DCs induced by human PBMCs exhibit a high level of
HLA-DR expression, a moderate level of CD1a, CD80 and CD86
expression, and a low level of CD83 expression. However, a low
level of CD14 expression was also exhibited by the DCs (Fig. 1A and B).
Mature DCs exhibited a laterally positioned nucleus
and less cytoplasm. A number of dendritic protuberances were also
observed on the cell membrane surface, which is the typical
morphological characteristic of DCs (Fig.
1C).
NDV-TCL-DCs exhibit increased levels
of co-stimulatory molecule expression
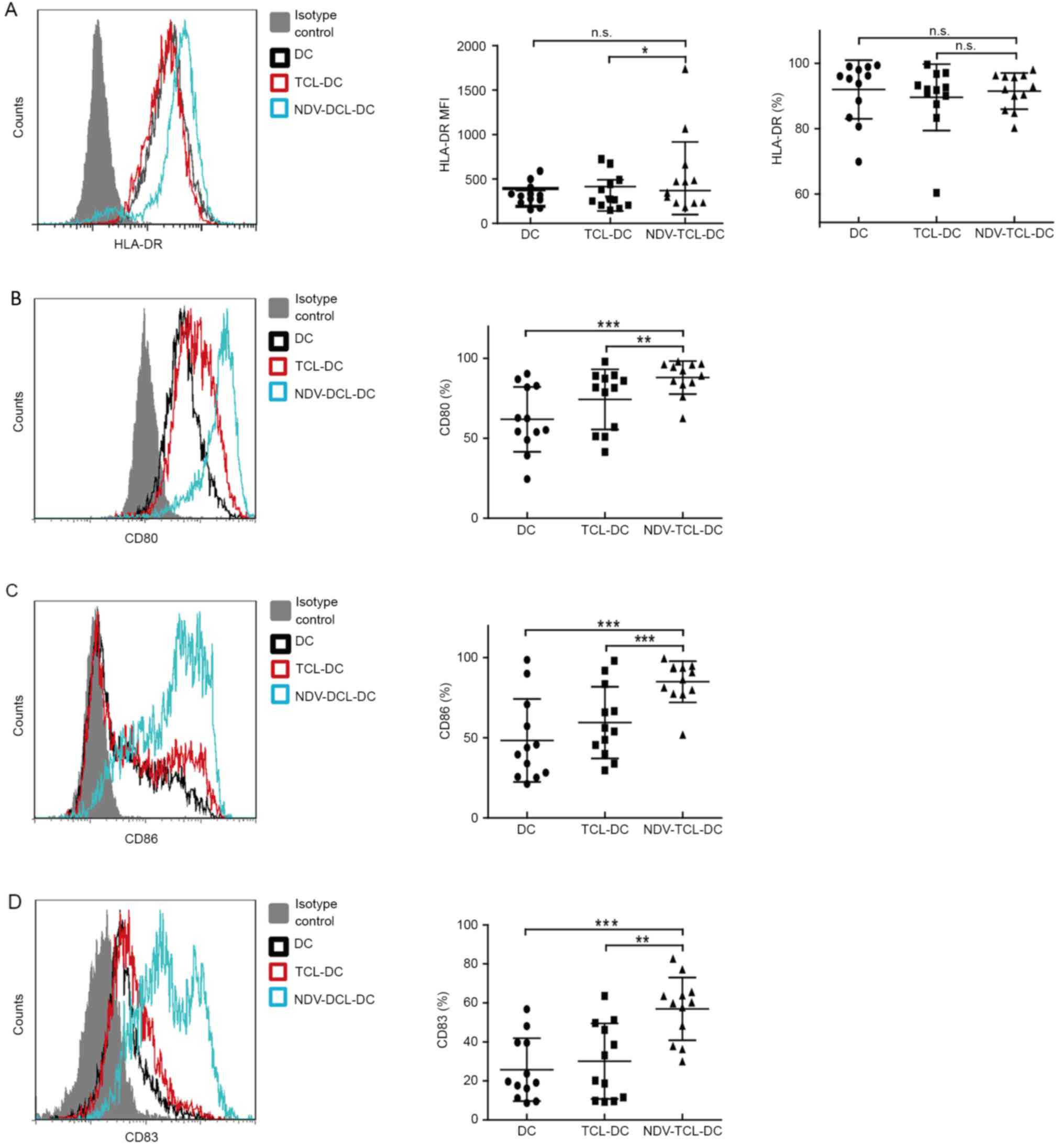
DC activation of naive T cells requires two signals:
One is initiated by the T-cell receptor (TCR) recognition of the
antigen peptide presented by the major histocompatibility complex
(MHC), including HLA-DR; the other is mediated by the
co-stimulatory molecule CD28 on T cells, with its ligands CD80 and
CD86, which are expressed on mature DCs. Nearly all of the
NDV-TCL-DCs (median, 91%; range, 80–98%) expressed HLA-DR (Fig. 2A) and the expression level was
increased compared with the TCL-DCs. CD80, CD83 and CD86 are
important markers of mature DCs. During the process of DC
maturation, the expression of CD83, CD80 and CD86 on DCs is
upregulated. In the present study, the expression of CD80, CD83 and
CD86 on the NDV-TCL-DCs was increased compared with that on the
TCL-DCs (P<0.01, P<0.001 and P<0.01, respectively) and
unloaded DCs (P<0.001; Fig. 2B-D).
These data indicated that the NDV-TCL-DCs were more mature than the
TCL-DCs and that they possessed a stronger potential to promote T
cells to exhibit antitumor ability.
NDV-TCL-DCs demonstrate an increased
PD-L1 expression compared with TCL-DCs
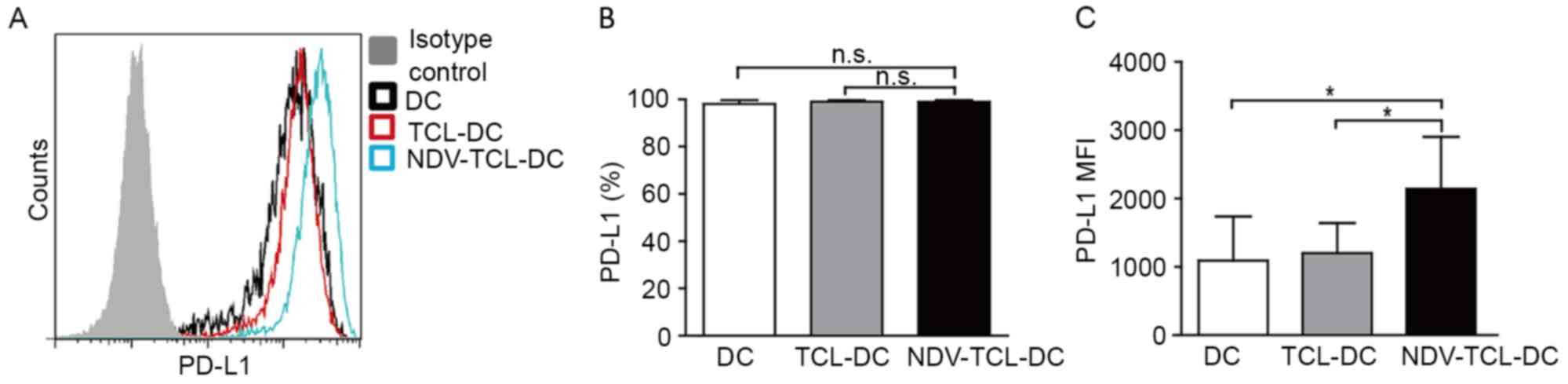
A number of studies have previously reported that
PD-L1 is expressed at high levels on mature DCs, along with
co-stimulatory molecules (15–18). In
the present study, it was demonstrated that although the expression
of PD-L1 on DCs did not differ significantly between the three
groups, the expression of PD-L1 on the NDV-TCL-DCs tended to
increase in comparison with the other two groups (Fig. 3A). These data further confirmed that
the NDV-TCL-DCs may be more mature than the TCL-DCs (19).
NDV-TCL-DCs promote allogeneic T-cell
proliferation
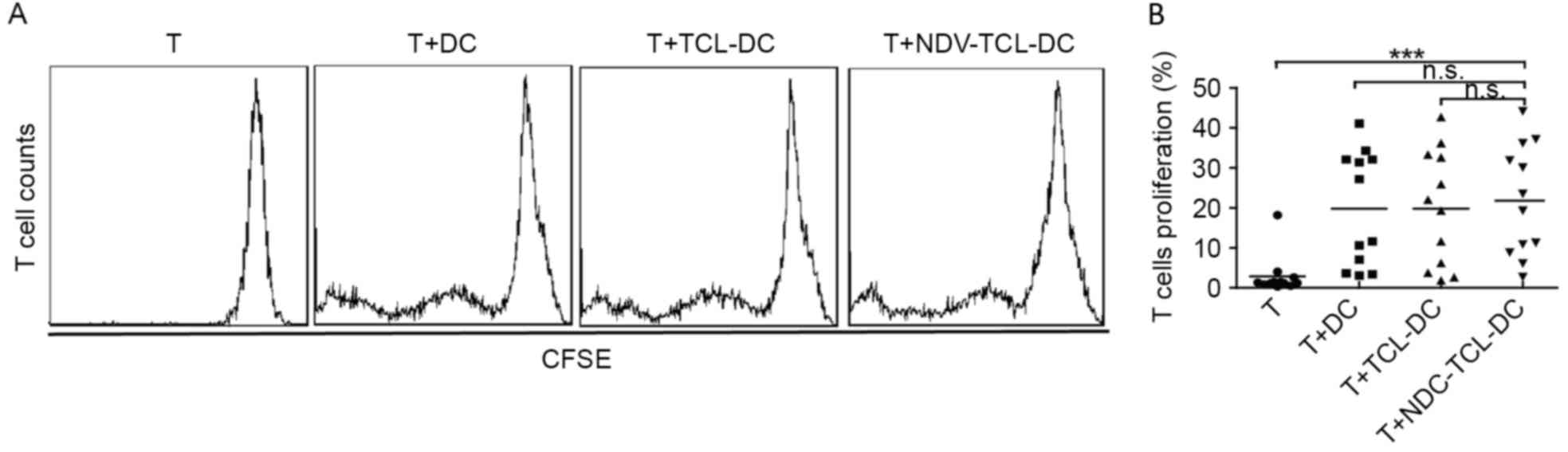
A mixed lymphocyte reaction is an ex vivo cellular
immune reaction that occurs between two allogeneic lymphocyte
populations. This assay was used to evaluate the ability of the
NDV-TCL-DCs to promote T-cell proliferation in vitro. As
depicted in Fig. 4, incubation with
the NDV-TCL-DCs significantly increased the proliferation of T
cells compared with the T cells alone (P<0.001). However, there
were no significant differences between the increase in T-cell
proliferation caused by the NDV-TCL-DCs and that caused by TCL-DCs,
which indicated that NDV-TCL-DCs had the same potential to
stimulate allogeneic T-cell proliferation as TCL-DCs.
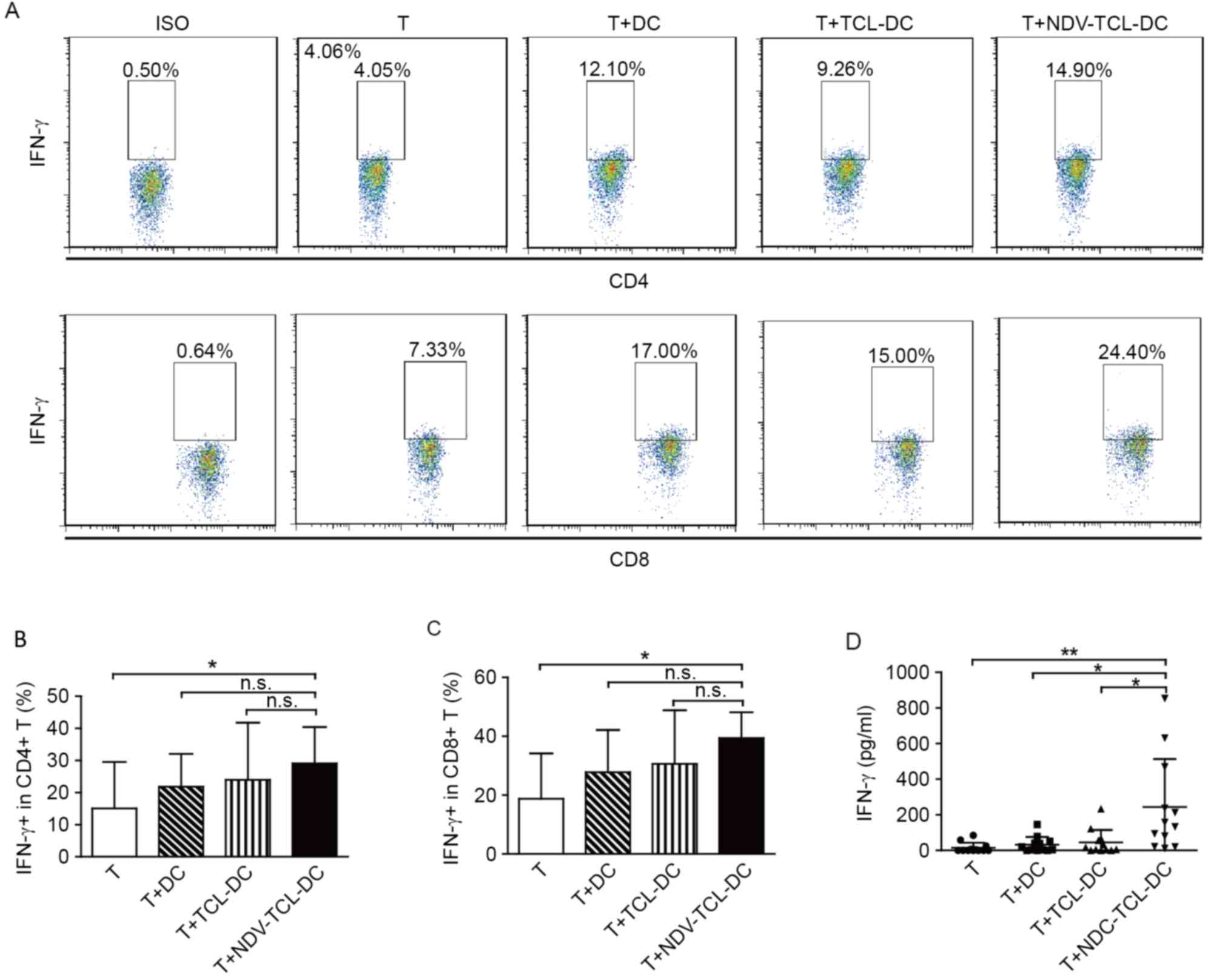
NDV-TCL-DCs enhance the IFN-γ
secretion of autologous CD4+ and CD8+ T
cells
The secretion of IFN-γ is regarded as an index of
T-cell activation. The results of the present study demonstrated
that incubation with NDV-TCL-DCs significantly increased the
intracellular IFN-γ production in both CD4+ and
CD8+ T cells compared with T cells alone (P<0.05;
Fig. 5A-C). The increase in IFN-γ
production by the T cells indicates that stimulation with
NDV-TCL-DCs can effectively activate CD4+ and
CD8+ T cells. Simultaneously, IFN-γ release in the
supernatants of the T cells co-cultured with NDV-TCL-DCs was
measured using the cytometric bead array technique. As depicted in
Fig. 5D, there was a negligible level
of IFN-γ in the supernatants of the T cells cultured alone and
those co-cultured with TCL-DCs. However, the level of IFN-γ in the
supernatant of T cells co-cultured with NDV-TCL-DCs was
significantly increased (P<0.01; Fig.
5D). These experiments revealed significant increases in the
activation of CD4+ T cells and CD8+ cytotoxic
T lymphocytes in response to co-culture with NDV-TCL-DCs.
Additionally, the results support our hypothesis that immunization
with NDV-TCL-DCs may activate specific CD4+ and
CD8+ T cells in cancer patients.
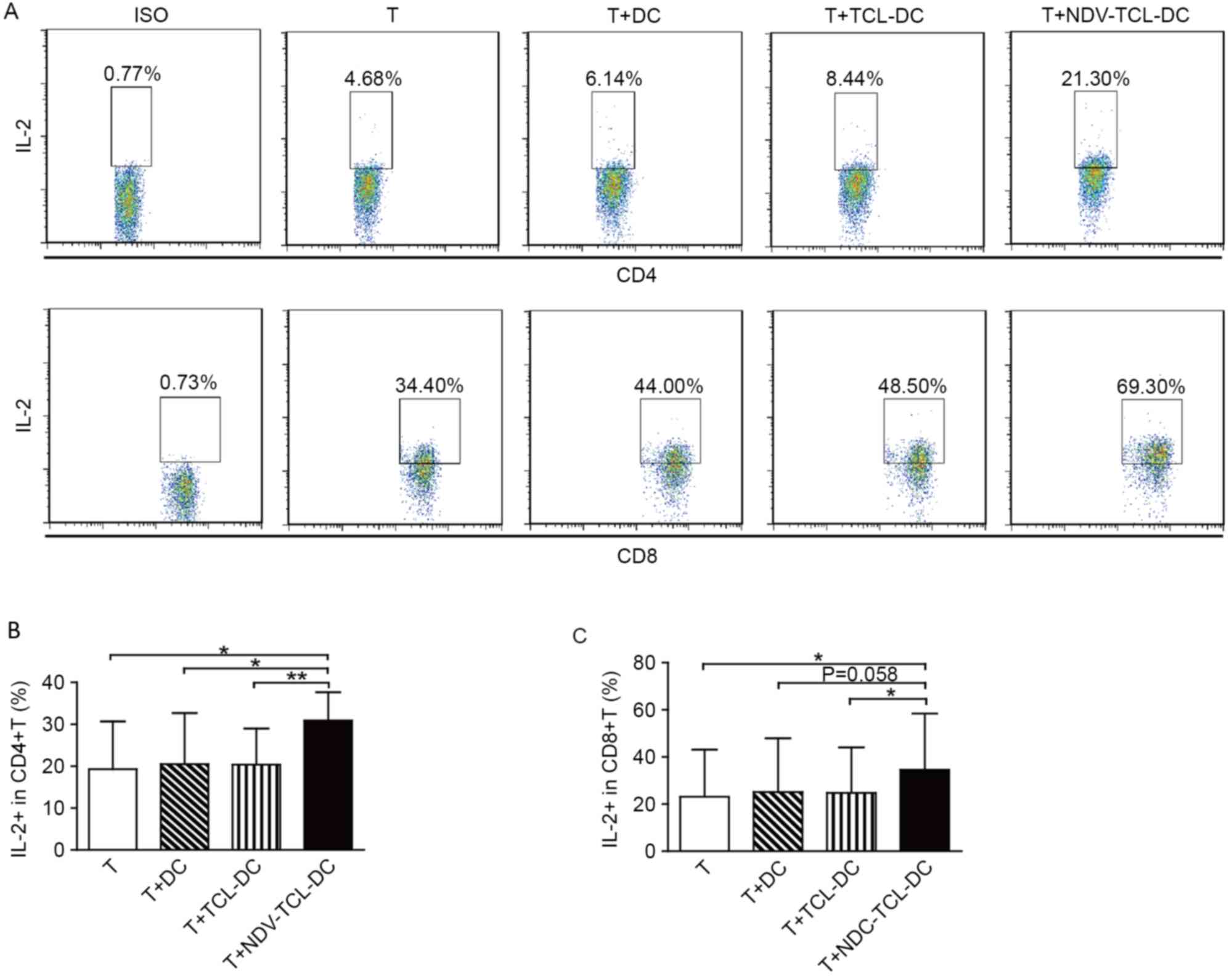
NDV-TCL-DCs enhances IL-2 secretion by
autologous CD4+ and CD8+ T cells
IL-2 serves a critical role in effector T-cell
development and antitumor functions. Therefore, the secretion of
intracellular IL-2 from CD4+ and CD8+ T cells
co-cultured with the NDV-TCL-DCs was also measured; it was
demonstrated that co-culture with the NDV-TCL-DCs significantly
enhanced the percentage of IL-2-positive CD4+ and
CD8+ T cells (P<0.05; Fig.
6). However, this effect was not observed upon co-culture with
TCL-DCs or DCs alone. These results indicated that the NDV-TCL-DCs
had the ability to enhance the percentage of IL-2-positive T
cells.
Discussion
The present study attempted to develop a novel DC
vaccine using DCs loaded with NDV-infected TCL. NDV-TCL-DCs
promoted T-cell proliferation and antitumor cytokine secretion. The
results of the present study indicated that loading DCs with
NDV-TCL could enhance the antigen-presenting ability of DCs. We
hypothesize that this novel DC-loading strategy could aid the
development of novel DC vaccines for tumor immunotherapy in the
future.
DC vaccines emerged in an effort to avoid possible
interference with therapeutic efficacy due to the dysfunction of
endogenous DCs, which were commonly observed in cancer patients
(20,21). However, it is important that the DCs
used in these vaccines are presented in a ‘mature’ state to
activate an antigen-specific immune response upon T-cell encounter.
This differentiated state is characterized by the expression of
several co-stimulatory molecules, which are necessary to activate
secondary signals in the immunological synapse (22). These molecules include CD80, CD86,
CD40, CD70 and inducible T-cell co-stimulator ligand (ICOS-L),
which interact with their counterparts CD28, CD40L, CD27, and ICOS,
respectively, which are expressed by T cells. In addition, mature
DCs should exhibit elevated levels of antigen-presenting molecules,
including MHC class I, MHC class II and CD1 (20,23). The
present study demonstrated that co-culture with the NDV-TCL-DCs
upregulated the expression of HLA-DR, CD80, CD86 and CD83, which
further facilitated the process of T-cell proliferation, and
activation. Without such co-stimulation, T-cell anergy or immune
tolerance could occur.
In addition to the aforementioned molecules, PD-L1
is constitutively expressed at low levels on hematopoietic cells,
including resting T cells, B cells, myeloid cells and immature DCs,
as well as on non-hematopoietic cells, including those in the
lungs, heart and other organs (24–26).
Previous studies have reported that PD-L1 is also expressed at high
levels on mature DCs, along with other co-stimulatory molecules
(17,27). PD-L1 expression on DCs is necessary
for maintaining immune tolerance in humans (28,29). PD-L1
expression on antigen presenting cells may inhibit the induction of
cytotoxic T lymphocytes by transducing negative signals onto T
cells; however, it has also been reported that the expression of
PD-L1 on antigen presenting cells, along with CD80 and CD86,
enhances the induction of antigen-specific cytotoxic T lymphocytes,
an effect probably depends on the fine-tuning of the excessive
stimulation of CD80 and CD86 (27).
In the present study, the NDV-TCL-DCs demonstrated an increasing
trend of PD-L1 expression, which was accompanied by a higher level
of CD80 and CD86 expression; therefore, NDV-TCL could increase the
PD-L1, CD80 and CD86 expression in DCs. This may be the reason why
the NDV-TCL-DCs were demonstrated to have the strongest ability to
induce T-cell proliferation and cytokine secretion.
The ability to secrete cytokines is critical in
determining whether T cells are activated when they recognize an
alloantigen (30). Monitoring the
secretion of major cytokines, including IFN-γ and IL-2 by
CD4+ and CD8+ T cells, could be useful in
determining the immune response to stimulation with DCs. The
experiments were conducted accordingly and it was demonstrated that
the secretion of IFN-γ and IL-2 by the CD4+ and
CD8+ T cells was significantly higher when they were
co-cultured with the NDV-TCL-DCs compared with that when
co-cultured with the TCL-DCs. Activated CD4+ and
CD8+ T cells are necessary for the antitumor activity of
the immune system (31). Therefore,
the results of the present study indicated that the NDV-TCL-DCs had
the ability to promote the secretion of antitumor cytokines from T
cells and that these DCs may therefore be a potential candidate for
use in tumor vaccines.
In conclusion, NDV-TCL-DCs were successfully
produced and it was demonstrated that they possess the potential
ability to promote T-cell proliferation and antitumor cytokine
secretion. However, the underlying mechanisms were not elucidated.
Future work should focus on investigation of these mechanisms.
Nevertheless, the results of the present study will be helpful in
developing novel DC-based tumor vaccines in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 31470798,
31700764 and 81402436), the Provincial Science Fund of Jilin
Provincial Department of Finance (grant nos. 2014N147, 2017C022,
20150204027YY, 20170622011JC, 20140414014GH and 20150520155JH), the
Norman Bethune Program of Jilin University (grant no. 2012202) and
the Youth Fund of the First Hospital of Jilin University (grant no.
JDYY72016003).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HJ and WL conceived and designed the research. LZ,
CN and ML performed the experiments and analyzed the data. DX, JX,
XS and JC contributed to sample collection. CN, LZ, and XS wrote
the manuscript. All authors reviewed the manuscript.
Ethics approval and consent to
participate
A total of 12 lung cancer patient were recruited
from the First Hospital of Jilin University (Changchun, China). All
of them provided written informed consent for the use of
biospecimens for research purposes according to the Declaration of
Helsinki. The present study was permitted by the Ethics Committee
of the First Hospital of Jilin University and carried out in
accordance with the approved guideline ‘Use of experimental animals
and human subjects.’
Consent for publication
The patients provided written informed consent for
the publication of all the associated data in this article. The
consent forms are available on reasonable request and will be
treated confidentially.
Competing interests
The authors declare that there was no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DCs
|
dendritic cells
|
|
NDV
|
Newcastle Disease Virus
|
|
TCL
|
tumor cell lysate
|
|
NDV-TCL
|
lysate of tumor cells infected with
NDV
|
|
TCL-DCs
|
DCs loaded with the TCL
|
|
NDV-TCL-DCs
|
DCs loaded with NDV-TCL
|
|
TAA
|
tumor associate antigens
|
|
PBMCs
|
Peripheral blood mononuclear cells
|
|
FBS
|
fetal bovine serum
|
|
MHC
|
major histocompatibility complex
|
|
ICOS-L
|
inducible T-cell co-stimulator
ligand
|
References
|
1
|
Sehgal K, Dhodapkar KM and Dhodapkar MV:
Targeting human dendritic cells in situ to improve vaccines.
Immunol Lett. 162:59–67. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steinman RM: Some interfaces of dendritic
cell biology. APMIS. 111:675–697. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Andrews DM, Scalzo AA, Yokoyama WM, Smyth
MJ and Degli-Esposti MA: Functional interactions between dendritic
cells and NK cells during viral infection. Nat Immunol. 4:175–181.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schirrmacher V, Ahlert T, Pröbstle T and
Steiner HH, Herold-Mende C, Gerhards R, Hagmüller E and Steiner HH:
Immunization with virus-modified tumor cells. Semin Oncol.
25:677–696. 1998.PubMed/NCBI
|
|
5
|
Thara E, Dorff TB, Pinski JK and Quinn DI:
Vaccine therapy with sipuleucel-T (Provenge) for prostate cancer.
Maturitas. 69:296–303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cohn L and Delamarre L: Dendritic
cell-targeted vaccines. Front Immunol. 5:2552014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guillerme JB, Boisgerault N, Roulois D,
Ménager J, Combredet C, Tangy F, Fonteneau JF and Gregoire M:
Measles virus vaccine-infected tumor cells induce tumor antigen
cross-presentation by human plasmacytoid dendritic cells. Clin
Cancer Res. 19:1147–1158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Maamary J, Array F, Gao Q, García-Sastre
A, Steinman RM, Palese P and Nchinda G: Newcastle disease virus
expressing a dendritic cell-targeted HIV gag protein induces a
potent gag-specific immune response in mice. J Virol. 85:2235–2246.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fournier P and Schirrmacher V: Oncolytic
Newcastle Disease Virus as cutting edge between tumor and host.
Biology (Basel). 2:936–975. 2013.PubMed/NCBI
|
|
10
|
Melchjorsen J, Jensen SB, Malmgaard L,
Rasmussen SB, Weber F, Bowie AG, Matikainen S and Paludan SR:
Activation of innate defense against a paramyxovirus is mediated by
RIG-I and TLR7 and TLR8 in a cell-type-specific manner. J Virol.
79:12944–12951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kato H, Sato S, Yoneyama M, Yamamoto M,
Uematsu S, Matsui K, Tsujimura T, Takeda K, Fujita T, Takeuchi O
and Akira S: Cell type-specific involvement of RIG-I in antiviral
response. Immunity. 23:19–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fournier P, Arnold A and Schirrmacher V:
Polarization of human monocyte-derived dendritic cells to DC1 by in
vitro stimulation with Newcastle Disease Virus. J BUON. 14(Suppl
1): S111–S122. 2009.PubMed/NCBI
|
|
13
|
Austin FC and Boone CW: Virus augmentation
of the antigenicity of tumor cell extracts. Adv Cancer Res.
30:301–345. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sinkovics JG and Horvath JC: Virus therapy
of human cancers. Melanoma Res. 13:431–432. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Freeman GJ, Long AJ, Iwai Y, Bourque K,
Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne
MC, et al: Engagement of the PD-1 immunoinhibitory receptor by a
novel B7 family member leads to negative regulation of lymphocyte
activation. J Exp Med. 192:1027–1034. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamazaki T, Akiba H, Iwai H, Matsuda H,
Aoki M, Tanno Y, Shin T, Tsuchiya H, Pardoll DM, Okumura K, et al:
Expression of programmed death 1 ligands by murine T cells and APC.
J Immunol. 169:5538–5545. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brown JA, Dorfman DM, Ma FR, Sullivan EL,
Munoz O, Wood CR, Greenfield EA and Freeman GJ: Blockade of
programmed death-1 ligands on dendritic cells enhances T cell
activation and cytokine production. J Immunol. 170:1257–1266. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nguyen LT and Ohashi PS: Clinical blockade
of PD1 and LAG3 - potential mechanisms of action. Nat Rev Immunol.
15:45–56. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Williams EH, Williams CA and McLeod JD:
Identification of PDL-1 as a novel biomarker of sensitizer exposure
in dendritic-like cells. Toxicol In Vitro. 24:1727–1735. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kalinski P, Edington H, Zeh HJ, Okada H,
Butterfield LH, Kirkwood JM and Bartlett DL: Dendritic cells in
cancer immunotherapy: Vaccines or autologous transplants? Immunol
Res. 50:235–247. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao F, Liu C, Guo J, Sun W, Xian L, Bai D,
Liu H, Cheng Y, Li B, Cui J, et al: Radiation-driven lipid
accumulation and dendritic cell dysfunction in cancer. Sci Rep.
5:96132015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Reichardt P, Dornbach B and Gunzer M: APC,
T cells, and the immune synapse. Curr Top Microbiol Immunol.
340:229–249. 2010.PubMed/NCBI
|
|
23
|
Linette GP and Carreno BM: Dendritic
cell-based vaccines: Shining the spotlight on signal 3.
Oncoimmunology. 2:e265122013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Talay O, Shen CH, Chen L and Chen J: B7-H1
(PD-L1) on T cells is required for T-cell-mediated conditioning of
dendritic cell maturation. Proc Natl Acad Sci USA. 106:2741–2746.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang SC, Latchman YE, Buhlmann JE,
Tomczak MF, Horwitz BH, Freeman GJ and Sharpe AH: Regulation of
PD-1, PD-L1, and PD-L2 expression during normal and autoimmune
responses. Eur J Immunol. 33:2706–2716. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rodig N, Ryan T, Allen JA, Pang H, Grabie
N, Chernova T, Greenfield EA, Liang SC, Sharpe AH, Lichtman AH and
Freeman GJ: Endothelial expression of PD-L1 and PD-L2
down-regulates CD8+ T cell activation and cytolysis. Eur J Immunol.
33:3117–3126. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Goto T, Nishida T, Takagi E, Miyao K,
Koyama D, Sakemura R, Hanajiri R, Watanabe K, Imahashi N, Terakura
S, et al: Programmed death-ligand 1 on antigen-presenting cells
facilitates the induction of antigen-specific cytotoxic T
lymphocytes: Application to adoptive T-cell immunotherapy. J
Immunother. 39:306–315. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martin-Orozco N, Wang YH, Yagita H and
Dong C: Cutting Edge: Programmed death (PD) ligand-1/PD-1
interaction is required for CD8+ T cell tolerance to tissue
antigens. J Immunol. 177:8291–8295. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Okazaki T and Honjo T: The PD-1-PD-L
pathway in immunological tolerance. Trends Immunol. 27:195–201.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Millán O, Rafael-Valdivia L, Torrademé E,
López A, Fortuna V, Sánchez-Cabus S, López-Púa Y, Rimola A and
Brunet M: Intracellular IFN-gamma and IL-2 expression monitoring as
surrogate markers of the risk of acute rejection and personal drug
response in de novo liver transplant recipients. Cytokine.
61:556–564. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shimoda M, Tomimaru Y, Charpentier KP,
Safran H, Carlson RI and Wands J: Tumor progression-related
transmembrane protein aspartate-β-hydroxylase is a target for
immunotherapy of hepatocellular carcinoma. J Hepatol. 56:1129–1135.
2012. View Article : Google Scholar : PubMed/NCBI
|