Introduction
Cholangiocarcinoma (CCA) is a malignant tumor of the
bile duct epithelium. The incidence of CCA is highest in northeast
Thailand, where the liver fluke (Opisthorchis viverrini,
hereafter Ov) infection rate is high (1). The association between Ov infection and
CCA is well established in this region (2,3) and Ov
infection is defined as a major risk factor for CCA (4).
The majority of patients with CCA have a poor
prognosis as early stage CCA being difficult to diagnose and the
onset of symptoms only occurring in advanced stages, causing
patients to seek treatment in later stages of disease (5). Surgical resection is curative only for
patients that present at an early stage, not for those at late or
advanced stages (6,7). Therefore, novel approaches, particularly
those involving targeted therapy, are urgently required.
Multiple protein kinases are activated in CCA
tissues and cell lines; these include protein kinases of the
phosphatidylinositol 3-kinase (PI3K)/RAC serine/threonine-protein
kinase (Akt) pathway (8). The
PI3K/Akt signaling pathway serves an important role in a number of
cellular processes, including cell growth, the cell cycle and
programmed cell death (9,10). The activation of this pathway is thus
involved in the progression of various types of cancer, including
CCA (9–13). Therefore, targeting the PI3K/Akt
pathway may be beneficial for CCA therapy.
Buparlisib or NVP-BKM120 is a highly selective
pan-class I PI3K inhibitor (14). The
ability of NVP-BKM120 to induce apoptosis or exert
anti-proliferative effects via the PI3K/Akt/mechanistic target of
rapamycin (mTOR) pathway has been demonstrated in various types of
cancer (15–18). In acute myeloid leukemia, NVP-BKM120
has been demonstrated to induce cell growth arrest and apoptosis by
inhibiting this signaling pathway (15). In addition, the antitumor activity of
NVP-BKM120 has also been demonstrated in bone and soft tissue
sarcoma, multiple myeloma and glioma (16–18).
Increased PI3K/Akt/mTOR activation has been revealed in multiple
types of cancer, including CCA (8,10,19,20);
NVP-BKM120 may thus possess the potential to inhibit the
progression of cancer. Therefore, the present study aimed to
evaluate the inhibitory effect of NVP-BKM120 on CCA progression and
to assess the molecular mechanism by which NVP-BKM120 suppresses
the development of CCA.
Materials and methods
Cell lines and cell culture
The human CCA M213, KKU100, M055, M139 and OCA17
cell lines, were established by Dr Banchop Sripa at the
Cholangiocarcinoma Research Institute (CARI), Khon Kaen University
(Khon Kaen, Thailand) (21). Cell
lines were cultured in Ham's F-12 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 2 mg/ml
sodium bicarbonate, 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
in a humidified atmosphere at 37°C which contained 5%
CO2.
Antibodies and reagents
The antibodies used in the present study were as
follows: Rabbit polyclonal anti-Ki67 antibodies (1:300, cat. no.
ab15580; Abcam, Cambridge, UK), rabbit monoclonal anti-Akt
antibodies (1:500, cat. no. ab32505; Abcam), rabbit polyclonal
anti-phospho-phosphatase and tensin homolog (PTEN; Ser380)
antibodies (1:500, cat. no. ab47332; Abcam), rabbit polyclonal
anti-phospho-Akt (Ser473) antibodies (1:1,000, cat. no. 9271; Cell
Signaling Technology, Inc., Danvers, MA, USA), rabbit monoclonal
anti-mTOR antibodies (1:1,000, cat. no. 2983; Cell Signaling
Technology, Inc.), rabbit polyclonal anti-phospho-mTOR (Ser2448)
antibodies (1:1,000, cat. no. 2971; Cell Signaling Technology,
Inc.), rabbit polyclonal anti-B-cell lymphoma-2 (Bcl-2) antibodies
(1:1,000, cat. no. 2876; Cell Signaling Technology, Inc.), mouse
monoclonal anti-Bcl-2 associated protein X (Bax) antibodies
(1:1,000, cat. no. 610983; BD Biosciences, Franklin Lakes, NJ, USA)
and mouse monoclonal anti-β-actin antibodies (1:10,000, cat. no.
5541; Sigma-Aldrich, Merck KGaA, Darmstadt, Germany). NVP-BKM120
was purchased from Active Biochem Ltd. (Hong Kong, China).
Growth inhibition assay
The growth inhibitory effect of NVP-BKM120 was
determined using a sulforhodamine B (SRB) assay. CCA cells
(5×103 cells in 100 µl Ham's F-12 medium) (Gibco; Thermo
Fisher Scientific, Inc.) were seeded in 96-well plates and
incubated overnight at 37°C and 5% CO2. The cells were
then treated with NVP-BKM120 in various concentrations, including
1, 10, 100, 1,000, 10,000 and 100,000 nM, and incubated for 48 h.
Following this, cells were fixed with 10% cold trichloroacetic acid
for 1 h at 4°C and stained with 0.4% w/v SRB in 1% v/v acetic acid
for 30 min at room temperature. Excess dye was washed with 1%
acetic acid and stained cells were solubilized with 200 µl of 10 mM
unbuffered Tris-base. The absorbance was measured using a
microplate reader (Sunrise; Tecan Group Ltd., Maanedorf,
Switzerland) at 540 nm. The percentage of growth inhibition (% GI)
in three independent experiments was calculated using the formula,
% GI=1-(Nt/Nc) ×100. Nt and Nc refer to the absorbance of the
treated and control groups, respectively. The half-maximal
inhibitory concentration (IC50) was evaluated using
interpolation from dose-response curves, as described previously
(22).
Animal study
The six-week-old, female athymic BALB/c nude mice (3
mice/group) weight range 18–20 g were purchased from the National
Animal Laboratory (Mahidol University, Nakhon Pathom, Thailand).
Mice were housed under pathogen-free conditions and given ad
libitum access to food and water in a temperature-controlled
room of 23±2°C, with a 12/12 h light/dark cycle, 10–15 air
changes/h, room humidity of 30–60% and light levels 350–400 lux at
the Animal Center, Faculty of Medicine, Khon Kaen University. Mice
were injected subcutaneously with 3×106 cells of the
M213 CCA cell line. When a tumor became visible, mice were divided
into three groups. The control group was orally administrated with
the vehicle (NMP-PEG3000; Sigma-Aldrich, Merck KGaA), whilst the
treatment groups received 10 or 30 mg/kg of NVP-BKM120 orally for
14 days. Body weight and tumor volume (tumor volume=0.5 ×
width2 × length) were determined twice a week. Relative
tumor volume was calculated by using the formula: Relative tumor
volume=tumor volume day x/tumor volume day 0). After three weeks of
the experiment, the mice were anesthetized with 2% isoflurane and
cervical dislocation was used for euthanasia. The tumor and organs
collected for further experimentation. All experiments were
approved by the Animal Ethics Committee of the Khon Kaen University
(NELAC22/2557).
Immunohistochemical assay for
Ki67
Nude mice tumor tissues were fixed in 10% buffered
formaldehyde at room temperature for a week, embedded in paraffin
blocks and then sectioned at a thickness of 4 µm. Ki67
Immunostaining was performed to determine cell proliferation using
tissue sections. Tissue sections were deparaffinized and rehydrated
with xylene and an ethanol series. The antigen was then retrieved
with Tris-EDTA buffer (pH 9) using a pressure cooker at 120°C for 3
min. Endogenous enzymes and non-specific bonding were blocked using
0.3% hydrogen peroxide and 10% skimmed milk with 30 min agitation
at room temperature, respectively. Anti-Ki67 antibodies were
incubated at room temperature for 1 h followed by a further
incubation at 4°C overnight in a moisture chamber. The sections
were then incubated with secondary antibodies conjugated to
horseradish peroxidase (Envision; Dako; Agilent Technologies, Inc.,
Santa Clara, CA, USA) at room temperature for 1 h and the signal
was developed using 0.1% diaminobenzidine tetrahydrochloride for 5
min. Sections were counterstained with hematoxylin at room
temperature for 2 min and dehydrated using an ethanol series
followed by xylene prior to mounting. Sections were observed using
a light microscope (Nikon Eclipse Ni-U; Nikon, Tokyo, Japan). Ki67
positive cells in each tumor section were counted in at least five
fields of view at a magnification of ×400 (NIS-Elements version
4.0; Nikon, Tokyo, Japan) and the percentage of Ki67 positive cells
were calculated.
In situ terminal deoxynucleotidyl
transferase dUTP nick-end labeling (TUNEL) assay for apoptosis
A TUNEL assay was performed to detected DNA
fragments in apoptotic cells. The TUNEL assay was performed using
paraffin-embedded tumor tissue from nude mice using the in
situ Cell Death Detection kit, POD (cat. no. 11 684 817 910;
Roche, Mannheim, Germany). Briefly, nude mice tumor tissues were
fixed in 10% buffered formaldehyde at room temperature for a week,
embedded in paraffin blocks and then sectioned at a thickness of 4
µm. Tissue sections were deparaffinized and rehydrated with xylene
and an ethanol series (100, 90, 80 and 70% ethanol) and treated
with 20 µg/ml proteinase K at 37°C for 30 min. TUNEL reaction
mixture was added 50 µl/section, incubate at 37°C for 1 h.
Converter-POD (anti-fluorescein antibody conjugated with
horseradish peroxidase) was added 50 µl/section. Tissue sections
were incubated at 37°C for 30 min and the signal was developed
using 0.1% diaminobenzidine tetrahydrochloride for 5 min. Tissue
sections were dehydrated prior to mounting (Bio Optica Milano SpA,
Milan, Italy). The percentage of TUNEL-positive cells was
determined using the light microscope from at least five fields of
view at a magnification of ×400 (NIS-Elements version 4.0;
Nikon).
Western blot analysis
Total protein was extracted from 50 mg of frozen
nude mouse tumor tissue, stored at −80°C. Tumor tissues were lysed
in 200 µl radioimmunoprecipitation assay lysis buffer (150 mM NaCl,
0.5 M Tris-HCl pH 7.4, 1% Tween-20, 1% sodium deoxycholate and 0.1%
SDS) for 10 min on ice. then homogenized using a grinder. Whole
lysates were then centrifuged at a speed of 14,000 g at 4°C for 5
min and the supernatant was collected and stored at −80°C until
further use. Protein concentration was determined using a
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.). Protein extract were solubilized in 4X SDS buffer (1 M
Tris-HCL pH 6.8, SDS, glycerol, β-mercaptoethanol, bromophenol
blue) containing dithiothreitol and boiled at 95°C for 5 min.
Protein was loaded (30 µg/well) and separated by SDS-PAGE (10% gel)
and then transferred to a polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). The membrane was blocked using 10%
skimmed milk in TBS overnight at 4°C, then incubated with
antibodies against Bax, Bcl-2, p-Akt, Akt, p-mTOR, mTOR, p-PTEN and
β-actin at room temperature for 1 h. The membrane was then rinsed
using TBS containing 0.1% polyoxyethylenesorbitan monolaurate
(TTBS) and incubated with secondary antibodies conjugated to
horseradish peroxidase (Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) at room temperature for 1 h prior to rinsing with TTBS
followed by TBS. The signal was developed using an ECL Prime
Western Blotting Detection system using ImageQuant™ LAS
4000 Control Software (GE Healthcare, Chicago, IL, USA). Human
β-actin was used as a loading control.
Statistical analysis
The results of the growth inhibition assay, Ki67
staining, apoptosis assay and relative tumor volume of nude mice
are represented as the mean ± standard deviation. Statistical
comparisons between two groups of Ki67 staining and apoptosis assay
was determined using unpaired Student's t-test. The relative tumor
volume of mice from different groups was analyzed using two-way
analysis of variance followed with the Bonferroni method.
Statistical analysis was performed using GraphPad Prism 5 (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant result.
Results
Growth inhibitory effect of NVP-BKM120
on CCA cell lines and CCA-inoculated nude mice
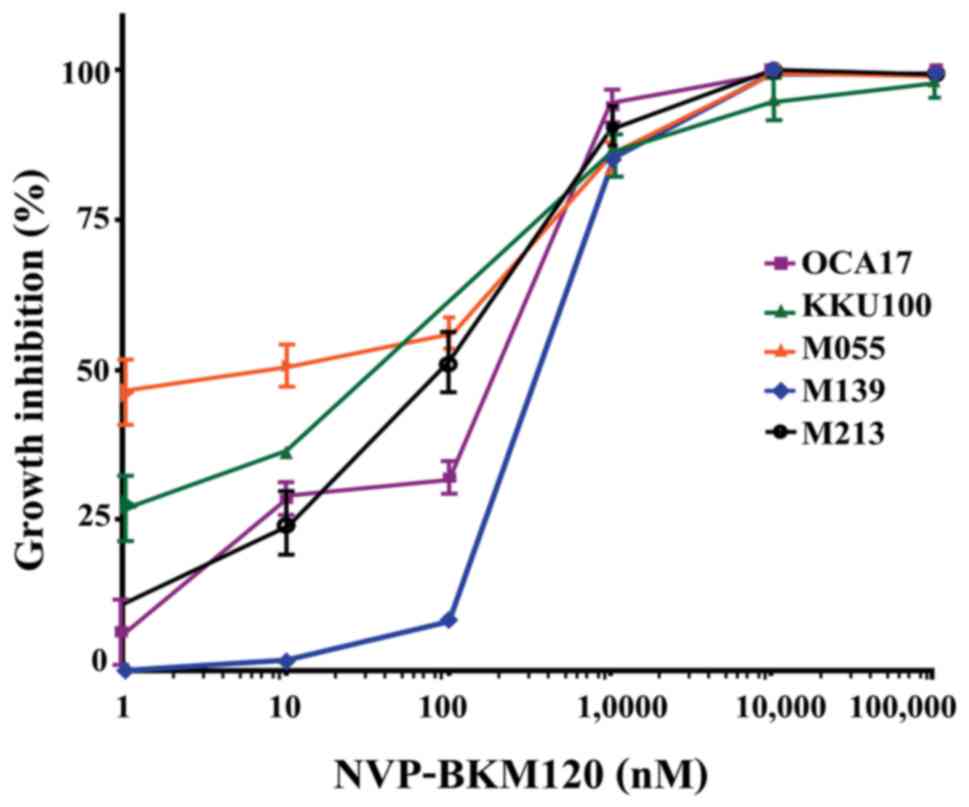
NVP-BKM120 inhibits CCA cell growth in a
dose-dependent manner (Fig. 1), with
the following IC50 values: M213, 85±38 nM; KKU100, 33±15
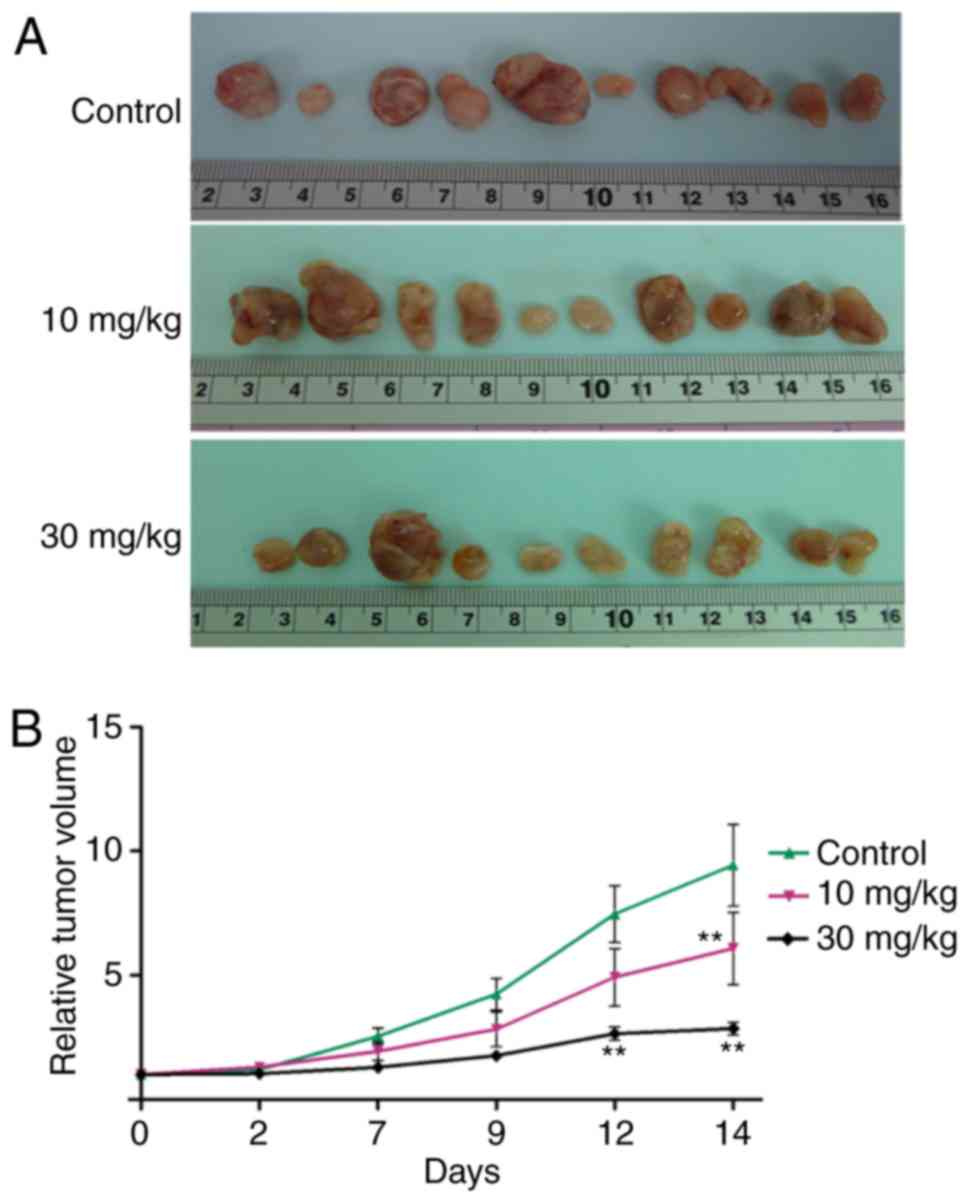
nM; M055, 4±4 nM; M139, 318±158 nM; and OCA17, 253±129 nM (Table I). Furthermore, NVP-BKM120 at
concentrations of 10 and 30 mg/kg significantly suppressed tumor
growth when compared with the control on days 14 and 12,
respectively (P<0.01; Fig. 2).
 | Table I.Growth inhibitory effect of
NVP-BKM120 on CCA cell lines. |
Table I.
Growth inhibitory effect of
NVP-BKM120 on CCA cell lines.
| CCA cell line | IC50 of
NVP-BKM120, nM |
|---|
| M213 | 85±38 |
| KKU100 | 33±15 |
| M055 | 4±4 |
| M139 | 318±158 |
| OCA17 | 253±129 |
NVP-BKM120 inhibits cancer cell
proliferation and induces apoptosis in the CCA mouse model
As demonstrated in vitro, NVP-BKM120 inhibits
CCA cell growth. Therefore, the effect of NVP-BKM120 on the mouse
model was assessed. An immunohistochemical assay was performed to
detect the Ki67 proliferation marker in the paraffin-embedded tumor
tissues of nude mice. The percentage of Ki67 nuclear staining in
the tumor cells of 30 mg/kg NVP-BKM120 treated mice was
significantly decreased when compared with the control mice
(P<0.05; Fig. 3A and B). In
addition, NVP-BKM120 treatment caused a significant increase in the
percentage of apoptotic cells in treatment groups when compared
with those in the control groups (10 mg/kg, P<0.001; 30 mg/kg,
P<0.01; Fig. 3C and D). Western
blot analysis demonstrated that NVP-BKM120 induced the expression
of the pro-apoptotic protein Bax, whereas the expression of Bcl-2,
which is anti-apoptotic, was decreased (Fig. 3E and F).
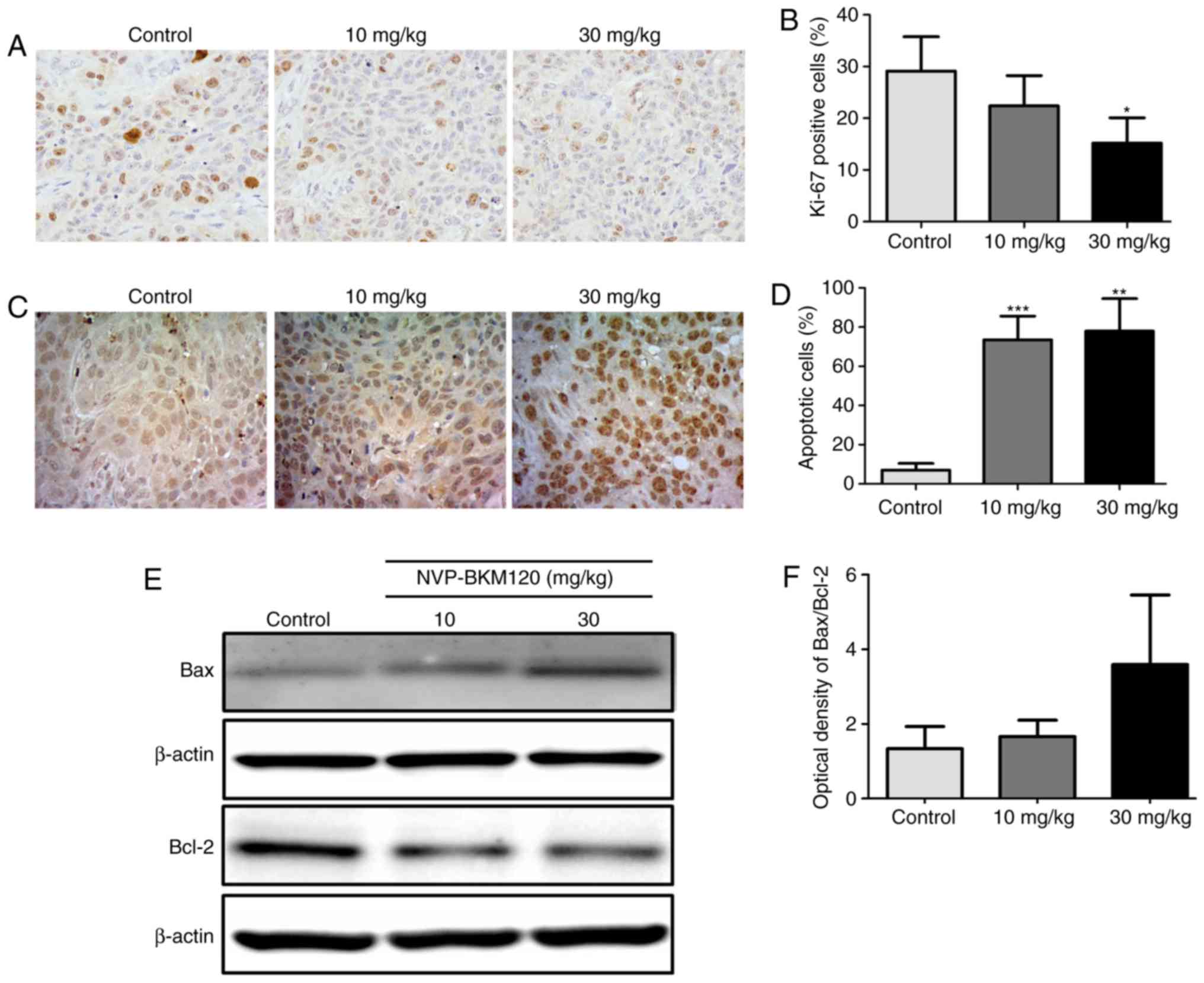 | Figure 3.NVP-BKM120 inhibits cancer cell
proliferation and induces apoptosis in a CCA mouse model. (A)
Proliferative cells in nude mouse tumor tissue sections were
determined using Ki67 immunostaining (magnification, ×400). (B) The
percentage of proliferative cells were significantly reduced in
mice treated with 30 mg/kg NVP-BKM120 when compared with controls.
(C) Apoptotic cells were detected using a in situ terminal
deoxynucleotidyl transferase dUTP nick end labeling assay
(magnification, ×400). (D) NVP-BKM120 treatment significantly
induced cell death in a dose-dependent manner when compared with
the control group. (E) Western blot analysis demonstrated an
increase in the expression of the pro-apoptotic protein, Bax,
whereas that of the anti-apoptotic protein Bcl-2, was decreased.
(F) Protein expression ratio of Bax/Bcl-2. Data in (B) and (D) are
presented as the mean ± standard deviation (3 mice in each group)
analyzed using a Student's t-test for independent samples. Data in
(F) are presented as the mean ± standard deviation of protein band
intensity, which was normalized with β-actin. *P<0.05,
**P<0.01, ***P<0.001. CCA, cholangiocarcinoma; Bax, Bcl-2
associated protein X; Bcl-2, B-cell lymphoma-2. |
Molecular mechanisms by which
NVP-BKM120 suppresses CCA cell growth
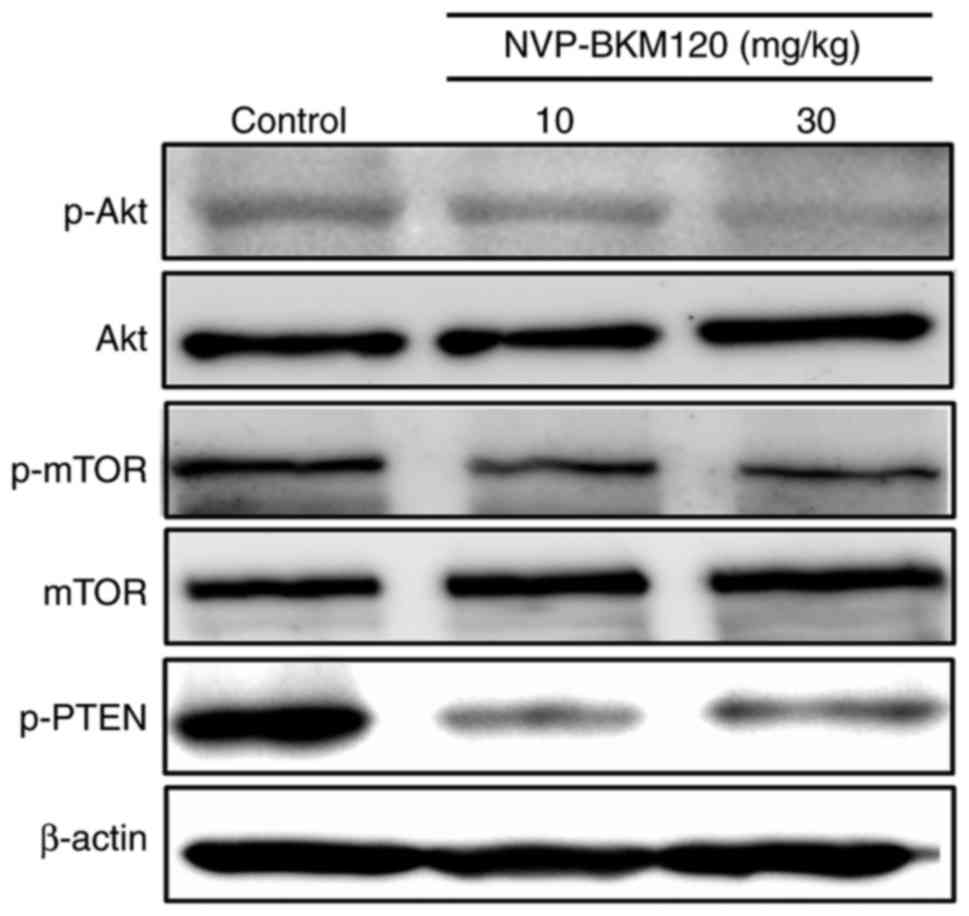
The present study also assessed the molecular
mechanism by which NVP-BKM120 suppressed CCA cell growth via the
PI3K/Akt pathway using a western blot assay. The results indicated
that NVP-BKM120 inhibits Akt and mTOR phosphorylation in a
dose-dependent manner. In addition, the drug also inhibits the
phosphorylation of PTEN, which is a negative regulator of the
PI3K/Akt/mTOR pathway (Fig. 4).
Discussion
PI3K/Akt/mTOR is a signaling pathway that serves a
notable role in various cellular processes, including growth, the
cell cycle and cell survival (23–25).
Previous studies have implicated this pathway in the progression of
certain types of cancer that are associated with poor patient
outcome (20,26–28). A
previous study observed an increase in PI3K/Akt/mTOR pathway
activation in CCA tissue and cell lines (8). Additionally, Yothaisong et al
(10) demonstrated that the increased
activation of this pathway was associated with CCA metastasis.
Therefore, targeting the PI3K/Akt/mTOR pathway in the treatment of
patients with CCA may be a beneficial approach.
Buparlisib or NVP-BKM120 is a highly selective
pan-class I PI3K inhibitor. It specifically blocks PI3K with a
specificity at least 50-fold higher than for other protein kinases
(14). The effect of NVP-BKM120 on
the inhibition of tumor growth and apoptosis induction has been
demonstrated in various types of cancer. It has been revealed to
inhibit cell growth, induce apoptosis and reduce the number and
size of colonies in bone and soft tissue sarcomas (16). Koul et al (29) revealed that NVP-BKM120 treatment
inhibits glioma cell proliferation. In addition, Martin et
al (17) demonstrated that
NVP-BKM120 treatment inhibited the growth of multiple myeloma
cells, and the antitumor effect of NVP-BKM120 was also detected in
breast cancer (30). Therefore, the
current study investigated the inhibitory effect of NVP-BKM120 on
CCA cells and evaluated its potential use in CCA therapy.
The present study demonstrated that NVP-BKM120
treatment inhibited CCA cell growth in a dose-dependent manner at
nanomolar concentrations. However, the IC50 levels of
NVP-BKM120 in glioma, breast cancer and lung adenocarcinoma
exhibited the same effects at micromolar ranges (29–31).
The present study assessed the effect of NVP-BKM120
on CCA development using the CCA-inoculated nude mouse model. The
oral administration of NVP-BKM120 at 10 and 30 mg/kg concentrations
significantly inhibited CCA cell growth when compared with
controls. This is consistent with the results of previous studies,
which demonstrated the growth inhibitory effect of NVP-BKM120 in
breast cancer and glioblastoma (32,33).
NVP-BKM120 also exhibited minimal or no toxic effects in normal
cells (34). The growth inhibitory
effect of NVP-BKM120 is indicated by immunostaining for the
proliferation marker Ki67. The results of the present study
indicated that the percentage of Ki67-positive nuclear-stained
cells in tumor tissues of the 30 mg/kg NVP-BKM120-treated group was
significantly lower than that of control group, similar to a
previous result reported in colorectal cancer (35). The effect of NVP-BKM120 treatment on
apoptosis using a TUNEL assay was then assessed. The number of
apoptotic cells was significantly increased in the tumor tissue of
mice treated with 10 or 30 mg/kg NVP-BKM120 when compared with
tissue from control animals. Furthermore, there was an increase in
expression of the pro-apoptotic protein Bax and a reduction in that
of the anti-apoptotic protein Bcl-2 in NVP-BKM120-treated mice.
These results indicated that NVP-BKM120 treatment inhibited CCA
progression via the induction of apoptosis, which reflect the
results of a previous study into acute lymphoblastic leukemia
(36).
There is considerable evidence to indicate that
increased activation of the PI3K/Akt signaling pathway can be
induced by various mechanisms, including mutation of PI3K or Akt,
constitutive activation of an upstream regulator, the loss of PTEN
expression and an increased expression of phospho-PTEN, which is
the inactive form (10,37). In addition, the results of the present
study revealed that increased activation of the PI3K/Akt signaling
pathway in CCA is primarily caused by, loss of function of the
negative regulator PTEN function via loss of expression, increased
phosphorylation and increased expression of the pathway components
(10). Thus, the molecular mechanism
by which NVP-BKM120 inhibits CCA development was assessed. The
current study analyzed the expression and phosphorylation of Akt,
mTOR and PTEN, which are over expressed in CCA and involved in
tumor cell proliferation. The results demonstrated that 30 mg/kg
NVP-BKM120 treatment markedly reduced Akt phosphorylation. These
are congruent to the results of a previous study of follicular
lymphoma, which revealed that 30 mg/kg NVP-BKM120 treatment also
reduced the phosphorylation of Akt (38). Furthermore, these results are similar
to a study concerning acute lymphoblastic leukemia, which
demonstrated that NVP-BKM120 (at 1, 10 and 50 µM) inhibits the
phosphorylation of Akt and mTOR in a dose-dependent manner
(36). In addition, the present study
also revealed a decrease in phospho-PTEN in the tumor tissues of
NVP-BKM120-treated mice. These results revealed that NVP-BKM120
inhibits CCA cell growth via the inhibition of Akt, mTOR and PTEN
phosphorylation.
In conclusion, the present study revealed that
NVP-BKM120 exhibited antitumor activity against CCA, inhibiting of
CCA cell growth at nanomolar concentrations. NVP-BKM120 suppressed
CCA growth and induce apoptosis in CCA-inoculated mice without
toxicity (data not shown). The current study also revealed that
NVP-BKM120 exerted an anticancer effect by blocking Akt, mTOR and
PTEN activation. These results indicated that targeting the
PI3K/Akt/mTOR signaling pathway with NVP-BKM120 led to the
suppression of CCA cell growth and the induction of cell death. The
present study thus provides data necessary for the development of
NVP-BKM120 treatment for CCA alone or in combination with
conventional chemotherapeutic drugs.
Acknowledgements
The authors would like to thank Professor Trevor N.
Petney for help with editing the manuscript.
Funding
The present study was supported by the Thailand
Research Fund a Research Assistantship Grant of the Faculty of
Medicine, Khon Kaen University (grant no. AS59204), Khon Kaen
University grant (grant no. KKU61) and grants from the Higher
Education Research Promotion and National Research University
Project of Thailand, Office of the Higher Education Commission,
through the Center of Excellence in Specific Health Problems in
Greater Mekong Sub-region cluster (SHeP-GMS), Khon Kaen University
allocated to WL.
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SP designed the experiments, performed the
experiments, analyzed the data, written the manuscript, prepared
figures and tables and reviewed drafts of the manuscript. HD and SY
designed the experiments, performed the experiments, analyzed the
data and prepared figures and tables. ATe, NN and PY designed the
experiments and provided reagents/materials/analysis tools. NK, ATi
and SS reviewed the drafts of the manuscript, designed experiments
and provided reagents/materials. WL designed the experiments,
analysed the data, provided reagents/materials/analysis tools and
reviewed the drafts of the manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Animal Ethics
Committee of the Khon Kaen University (NELAC22/2557; Khon Kaen,
Thailand).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vatanasapt V, Sriamporn S and Vatanasapt
P: Cancer control in Thailand. Jpn J Clin Oncol. 32:(Suppl):.
S82–S91. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Ohshima H, Srivatanakul P and
Vatanasapt V: Cholangiocarcinoma: Epidemiology, mechanisms of
carcinogenesis and prevention. Cancer Epidemiol Biomarkers Prev.
2:537–544. 1993.PubMed/NCBI
|
|
3
|
Thamavit W, Bhamarapravati N, Sahaphong S,
Vajrasthira S and Angsubhakorn S: Effects of dimethylnitrosamine on
induction of cholangiocarcinoma in Opisthorchis viverrini-infected
Syrian golden hamsters. Cancer Res. 38:4634–4639. 1978.PubMed/NCBI
|
|
4
|
Sripa B and Pairojkul C:
Cholangiocarcinoma: Lessons from Thailand. Curr Opin Gastroenterol.
24:349–356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Banales JM, Cardinale V, Carpino G,
Marzioni M, Andersen JB, Invernizzi P, Lind GE, Folseraas T, Forbes
SJ, Fouassier L, et al: Expert consensus document:
Cholangiocarcinoma: Current knowledge and future perspectives
consensus statement from the European Network for the Study of
Cholangiocarcinoma) ENS-CCA). Nat Rev Gastroenterol Hepatol.
13:261–280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Blechacz B and Gores GJ:
Cholangiocarcinoma: Advances in pathogenesis, diagnosis, and
treatment. Hepatology. 48:308–321. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Khan SA, Thomas HC, Davidson BR and
Taylor-Robinson SD: Cholangiocarcinoma. Lancet. 366:1303–1314.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dokduang H, Juntana S, Techasen A, Namwat
N, Yongvanit P, Khuntikeo N, Riggins GJ and Loilome W: Survey of
activated kinase proteins reveals potential targets for
cholangiocarcinoma treatment. Tumour Biol. 34:3519–3528. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fresno Vara JA, Casado E, de Castro J,
Cejas P, Belda-Iniesta C and González-Barón M: PI3K/Akt signalling
pathway and cancer. Cancer Treat Rev. 30:193–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yothaisong S, Dokduang H, Techasen A,
Namwat N, Yongvanit P, Bhudhisawasdi V, Puapairoj A, Riggins GJ and
Loilome W: Increased activation of PI3K/AKT signaling pathway is
associated with cholangiocarcinoma metastasis and PI3K/mTOR
inhibition presents a possible therapeutic strategy. Tumour Biol.
34:3637–3648. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
DeGraffenried LA, Fulcher L, Friedrichs
WE, Grünwald V, Ray RB and Hidalgo M: Reduced PTEN expression in
breast cancer cells confers susceptibility to inhibitors of the PI3
kinase/Akt pathway. Ann Oncol. 15:1510–1516. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shaw RJ and Cantley LC: Ras, PI(3)K and
mTOR signalling controls tumour cell growth. Nature. 441:424–430.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shukla S, Maclennan GT, Hartman DJ, Fu P,
Resnick MI and Gupta S: Activation of PI3K-Akt signaling pathway
promotes prostate cancer cell invasion. Int J Cancer.
121:1424–1432. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Maira SM, Pecchi S, Huang A, Burger M,
Knapp M, Sterker D, Schnell C, Guthy D, Nagel T, Wiesmann M, et al:
Identification and characterization of NVP-BKM120, an orally
available pan-class I PI3-kinase inhibitor. Mol Cancer Ther.
11:317–328. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Allegretti M, Ricciardi MR, Licchetta R,
Mirabilii S, Orecchioni S, Reggiani F, Talarico G, Foà R, Bertolini
F, Amadori S, et al: The pan-class I phosphatidyl-inositol-3 kinase
inhibitor NVP-BKM120 demonstrates anti-leukemic activity in acute
myeloid leukemia. Sci Rep. 5:181372015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Anderson JL, Park A, Akiyama R, Tap WD,
Denny CT and Federman N: Evaluation of in vitro activity of the
class I PI3K inhibitor Buparlisib (BKM120) in pediatric bone and
soft tissue sarcomas. PLoS One. 10:e01336102015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Martin SK, Gan ZY, Fitter S, To LB and
Zannettino AC: The effect of the PI3K inhibitor BKM120 on tumour
growth and osteolytic bone disease in multiple myeloma. Leuk Res.
39:380–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Speranza MC, Nowicki MO, Behera P, Cho CF,
Chiocca EA and Lawler SE: BKM-120 (Buparlisib): A
phosphatidyl-inositol-3 kinase inhibitor with anti-invasive
properties in glioblastoma. Sci Rep. 6:201892016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chang KY, Tsai SY, Chen SH, Tsou HH, Yen
CJ, Liu KJ, Fang HL, Wu HC, Chuang BF, Chou SW, et al: Dissecting
the EGFR-PI3K-AKT pathway in oral cancer highlights the role of the
EGFR variant III and its clinical relevance. J Biomed Sci.
20:432013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Malinowsky K, Nitsche U, Janssen KP, Bader
FG, Späth C, Drecoll E, Keller G, Höfler H, Slotta-Huspenina J and
Becker KF: Activation of the PI3K/AKT pathway correlates with
prognosis in stage II colon cancer. Br J Cancer. 110:2081–2089.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sripa B, Leungwattanawanit S, Nitta T,
Wongkham C, Bhudhisawasdi V, Puapairoj A, Sripa C and Miwa M:
Establishment and characterization of an opisthorchiasis-associated
cholangiocarcinoma cell line (KKU-100). World J Gastroenterol.
11:3392–3397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun SY, Yue P, Dawson MI, Shroot B, Michel
S, Lamph WW, Heyman RA, Teng M, Chandraratna RA, Shudo K, et al:
Differential effects of synthetic nuclear retinoid
receptor-selective retinoids on the growth of human non-small cell
lung carcinoma cells. Cancer Res. 57:4931–4939. 1997.PubMed/NCBI
|
|
23
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chalhoub N and Baker SJ: PTEN and the
PI3-kinase pathway in cancer. Annu Rev Pathol. 4:127–150. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Deng L, Chen J, Zhong XR, Luo T, Wang YP,
Huang HF, Yin LJ, Qiu Y, Bu H, Lv Q and Zheng H: Correlation
between activation of PI3K/AKT/mTOR pathway and prognosis of breast
cancer in Chinese women. PLoS One. 10:e01205112015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ocana A, Vera-Badillo F, Al-Mubarak M,
Templeton AJ, Corrales-Sanchez V, Diez-Gonzalez L, Cuenca-Lopez MD,
Seruga B, Pandiella A and Amir E: Activation of the PI3K/mTOR/AKT
pathway and survival in solid tumors: Systematic review and
meta-analysis. PLoS One. 9:e952192014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi J, Yao D, Liu W, Wang N, Lv H, Zhang
G, Ji M, Xu L, He N, Shi B and Hou P: Highly frequent PIK3CA
amplification is associated with poor prognosis in gastric cancer.
BMC Cancer. 12:502012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Koul D, Fu J, Shen R, LaFortune TA, Wang
S, Tiao N, Kim YW, Liu JL, Ramnarian D, Yuan Y, et al: Antitumor
activity of NVP-BKM120-a selective pan class I PI3 kinase inhibitor
showed differential forms of cell death based on p53 status of
glioma cells. Clin Cancer Res. 18:184–195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu Y, Guo R, Wei J, Zhou Y, Ji W, Liu J,
Zhi X and Zhang J: Effects of PI3K inhibitor NVP-BKM120 on
overcoming drug resistance and eliminating cancer stem cells in
human breast cancer cells. Cell Death Dis. 6:e20202015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liang YC, Wu HG, Xue HJ, Liu Q, Shi LL,
Liu T and Wu G: Effects of PI3K inhibitor NVP-BKM120 on acquired
resistance to gefitinib of human lung adenocarcinoma H1975 cells. J
Huazhong Univ Sci Technolog Med Sci. 33:845–851. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ayub A, Yip WK and Seow HF: Dual
treatments targeting IGF-1R, PI3K, mTORC or MEK synergize to
inhibit cell growth, induce apoptosis, and arrest cell cycle at G1
phase in MDA-MB-231 cell line. Biomed Pharmacother. 75:40–50. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Filbin MG, Dabral SK, Pazyra-Murphy MF,
Ramkissoon S, Kung AL, Pak E, Chung J, Theisen MA, Sun Y,
Franchetti Y, et al: Coordinate activation of Shh and PI3K
signaling in PTEN-deficient glioblastoma: New therapeutic
opportunities. Nat Med. 19:1518–1523. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Civallero M, Cosenza M, Pozzi S, Bari A,
Ferri P and Sacchi S: Activity of BKM120 and BEZ235 against
Lymphoma Cells. Biomed Res Int. 2015:8709182015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roper J, Sinnamon MJ, Coffee EM, Belmont
P, Keung L, Georgeon-Richard L, Wang WV, Faber AC, Yun J, Yilmaz
ÖH, et al: Combination PI3K/MEK inhibition promotes tumor apoptosis
and regression in PIK3CA wild-type, KRAS mutant colorectal cancer.
Cancer Lett. 347:204–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pereira JK, Machado-Neto JA, Lopes MR,
Morini BC, Traina F, Costa FF, Saad ST and Favaro P: Molecular
effects of the phosphatidylinositol-3-kinase inhibitor NVP-BKM120
on T and B-cell acute lymphoblastic leukaemia. Eur J Cancer.
51:2076–2085. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Park E, Park J, Han SW, Im SA, Kim TY, Oh
DY and Bang YJ: NVP-BKM120, a novel PI3K inhibitor, shows synergism
with a STAT3 inhibitor in human gastric cancer cells harboring KRAS
mutations. Int J Oncol. 40:1259–1266. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Matas-Céspedes A, Rodriguez V, Kalko SG,
Vidal-Crespo A, Rosich L, Casserras T, Balsas P, Villamor N, Giné
E, Campo E, et al: Disruption of follicular dendritic
cells-follicular lymphoma cross-talk by the pan-PI3K inhibitor
BKM120)Buparlisib). Clin Cancer Res. 20:3458–3471. 2014. View Article : Google Scholar : PubMed/NCBI
|