Introduction
Lichen sclerosus (LS) is a chronic relapsing disease
that predominantly affects the female anogenital region. It may
occur at any age, but the majority of cases are reported in
postmenopausal women (1). It evokes
multiple symptoms, including intense itching, pain, burning,
stenosis and dysuria, affecting the quality of normal life and
sexual activity (1). Currently, there
is no cure for LS; instead treatment is primarily aimed at the
inhibition of itching, and prevention of scar formation and vulvar
anatomical deformities. Previous studies have indicated that
numerous factors are involved in LS, including autoimmune
disturbance (2), abnormal hormone
secretion (3), genetic factors and
inflammation (4), although there is
no consensus regarding their relative importance. Since the
malignant potential of invasive vulvar carcinoma for LS is ~4%,
~60% of invasive squamous cell carcinoma are identified in the
adjacent site of the LS (5), and thus
it is considered as a precancerous lesion. The typical pathological
changes of LS include skin epidermis atrophy over keratosis, a
dermal layer of connective tissue fibrosis forming a homogeneous
zone and lymphocyte infiltration. Genes associated with LS have
been identified; these have primarily focused on the dysregulated
epidermis, for the hyalinization of collagen fibers in fibroblast
hinders further investigation in the superior dermis. The
keratinocyte protein, galectin-7, is an apoptosis-associated
protein that serves multiple roles in epidermal differentiation,
maturation and regeneration (6).
Although galectin-7 is not a canonical secreted protein that is
transported through the vesicular biosynthetic pathway, it has the
ability to reach the cell surface in a fully folded form via an
‘unconventional protein secretion’ mechanism (7). Thus, we hypothesized that galectin-7 may
be secreted by keratinocytes and infiltrate the superior dermis,
influencing the cellular activity of the dermal fibroblasts.
Patients and methods
Patient selection
The present study was approved by the Ethics
Committee of China Medical University (Shenyang, China) and written
informed consent as obtained from all patients. The present study
involved 5 outpatients, 10 in-patients and 10 healthy biopsy
tissues obtained from sexual plastic surgery between September 2011
and December 2013. The mean age of the 15 patients with VLS was
55.7±4.3 years (range, 34–65 years). Patients were diagnosed via
the evident VLS morphology and samples were further confirmed by
histopathological analysis. All patients had no comorbidities or
other medical history and were comparable.
Immunohistochemistry
A total of 15 VLS and 10 normal vulva skin 4%
formalin-fixed (Boster Biological Technology, Pleasanton, CA, USA)
at room temperature overnight. The paraffin-embedded 4 µm sections
of biopsies were examined using an optical microscope
(magnification, ×200) for galectin-7 expression. Rabbit monoclonal
anti-galectin-7 antibody (1:400; cat. no. AB108623; Abcam,
Cambridge, UK) was used as the primary antibody at 4°C overnight.
Alkaline phosphatase-conjugated goat monoclonal anti-rabbit
antibody (1:625; cat. no. ZB-2301; OriGene Technologies, Inc.,
Beijing, China) was used as the secondary antibody at room
temperature for 1 h. All immunohistochemical staining were also
performed on control biopsy samples. The specificity of the primary
antibody was confirmed with the negative signal obtained by
substituting normal rabbit serum (ZLI-9023; OriGene Technologies,
Inc.) for the primary antibody in the protocol. The pixel of
integrated density from 6 sections/sample was measured using ImageJ
1.46 (National Institutes of Health, Bethesda, MD, USA), and a
median pixel was calculated.
Western blot analysis
The protein from 10 paired VLS and healthy vulvar
tissue samples were extracted using radioimmunoprecipitation assay
buffer (P0013B; Beyotime Institute of Biotechnology, Shanghai,
China) and quantified using a bicinchoninic acid assay. Loading
samples were diluted with protein lysis buffer to produce a final
concentration of 5 µg/µl. SDS-PAGE (15%) was performed to separate
the protein samples (14 µl) by size. Following membrane transfer to
a polyvinylidene fluoride membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA), was blocked with blocking buffer containing 5%
non-fat milk in PBS with Tween-20 at room temperature for 2 h.
Blocked membranes were incubated overnight at 4°C with a rabbit
monoclonal antibody against galectin-7 (1:1,000; cat. no. AB108623;
Abcam) and rabbit monoclonal β-actin (1:1,000; cat. no. SC-47778;
Santa Cruz Biotechnology Inc., Dallas, TX, USA) at a dilution of
1:800. Signals were detected using
horseradish-peroxidase-anti-rabbit secondary antibodies (1:1,000;
cat. no. A0277) and an electrochemiluminescence detection kit (both
Beyotime Institute of Biotechnology). Images were captured using a
Quantity One v.4.62 software (Bio-Rad Laboratories, Inc.).
Isolation and cultivation of primary
dermal fibroblasts
Samples were placed in 70% ethanol for 10 sec and
washed five times with PBS. Then, samples were placed in Dulbecco's
modified Eagle medium (DMEM; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) with antibiotics. Additional fat tissues were
discarded. Subsequently, the samples were cut into 2–3 mm pieces,
placed in a petri dish and incubated at 37°C in a humidified
atmosphere with 5% CO2 and the medium was replaced every
3 days. Fibroblast outgrowths were harvested by trypsinization and
re-seeded (100–110 cells/ml) in a T10 flask in DMEM. Cells were
allowed to reach 90% confluence prior to freezing (−80°C) or
splitting for use in the galectin-7 experiments.
Cell viability assay
After three passages, fibroblasts were seeded into
96-well plates at a density of 5×103 cells/well. After
24 h, different concentrations of galectin-7 were added (0.2, 0.5,
1, 2.0 and 5.0 µg/ml) to the DMEM. Cells were cultivated for
another 48 h, then 20 µl MTS was added to 100 µl medium and the
plate was incubated at 37°C avoiding exposure to light for 4 h.
Optical density values were obtained with a plate reader at 490 nm,
data were corrected with blank controls.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
qPCR primers were designed as follows: Collagen I
forward, 5′-CCCCCTCCCCAGCCACAAAG-3′ and reverse,
5′-TCTTGGTCGGTGGGTGACTCT-3′ (product size, 360 bp); collagen III
forward, 5′-CCAAACTCTATCTGAA-3′ and reverse, 5′-GGACTCATAGAATACA-3′
(product size, 449 bp); β-actin forward,
5′-ATCTGGCACCACACTTCTACA-3′ and reverse,
5′-GTTTCGTGGATGCCACAGGCT-3′ (product size, 577 bp).
Briefly, cells were harvested following incubation
with different galectin-7 concentrations. According to the results
of cell viability assay, the cells were assigned as normal human
fibroblasts group (N-HF), low concentration group (≤1 µg/ml,
L-galectin) and high concentration group (>1 µg/ml, H-galectin).
The RNA was isolated with TRIzol (Life Technologies; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol from
duplicate or triplicate cell samples. cDNA synthesis was performed
on 1 µg RNA with SuperScript™ III First-Strand system
(Invitrogen; Thermo Fisher Scientific, Inc.) and oligo(dT) primers,
according to the manufacturers protocols. The cDNA was diluted to a
final volume of 200 and 1 µl was added to 500 nM of the forward and
reverse primers in a final volume of 10 µl per PCR reaction. qPCR
was performed with SYBR® Green (Takara Biotechnology
Co., Ltd., Dalian, China) under the following conditions:
Pre-denature 95°C for 1 min; then denature at 95°C for 30 sec;
annealing 44°C for 30 sec; elongation 72°C for 1 min (35 cycles).
Melting curve data was collected following amplification. The
negative control was loaded with ddH2O. Data were
analyzed using the ΔΔCq method (8).
Charts were plotted using Microsoft Excel 2007 (Microsoft
Corporation, Redmond, WA, USA).
Statistical analysis
Statistical analyses were conducted using SPSS
v.17.0 software (SPSS Inc., Chicago, IL, USA). Independent samples
t-tests were used to compare the expression difference of
Gelectin-7 between vulvar normal skin and VLS. Statistical
comparisons among groups in the cell viability assay and qPCR were
analyzed using one-way analysis of variance with the least
significant difference post hoc test. All data are presented as the
mean ± standard error of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
High galectin-7 level in VLS
tissues
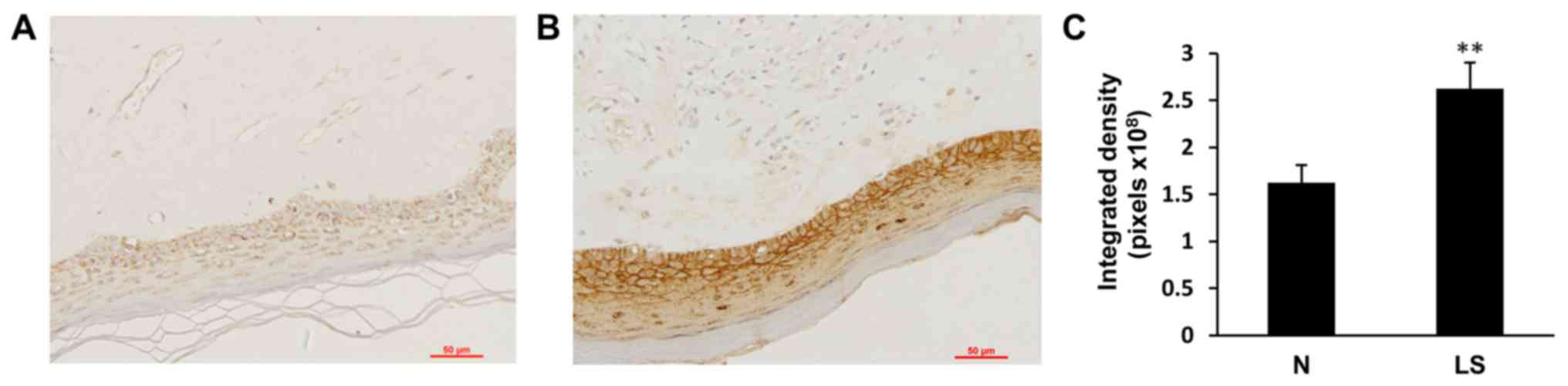
To determine the expression of galectin-7 in the
epidermis, immunohistochemical staining of galectin-7 was performed
in healthy control biopsy and VLS samples. In the healthy control
skin, moderate expression of galectin-7 was detected in the inner
part of epidermis, with an even distribution in the nucleus and
cytoplasm. In contrast, VLS tissue exhibited significantly elevated
immune expression of galectin-7 in the superficial epidermis
(Fig. 1; P<0.01). To further
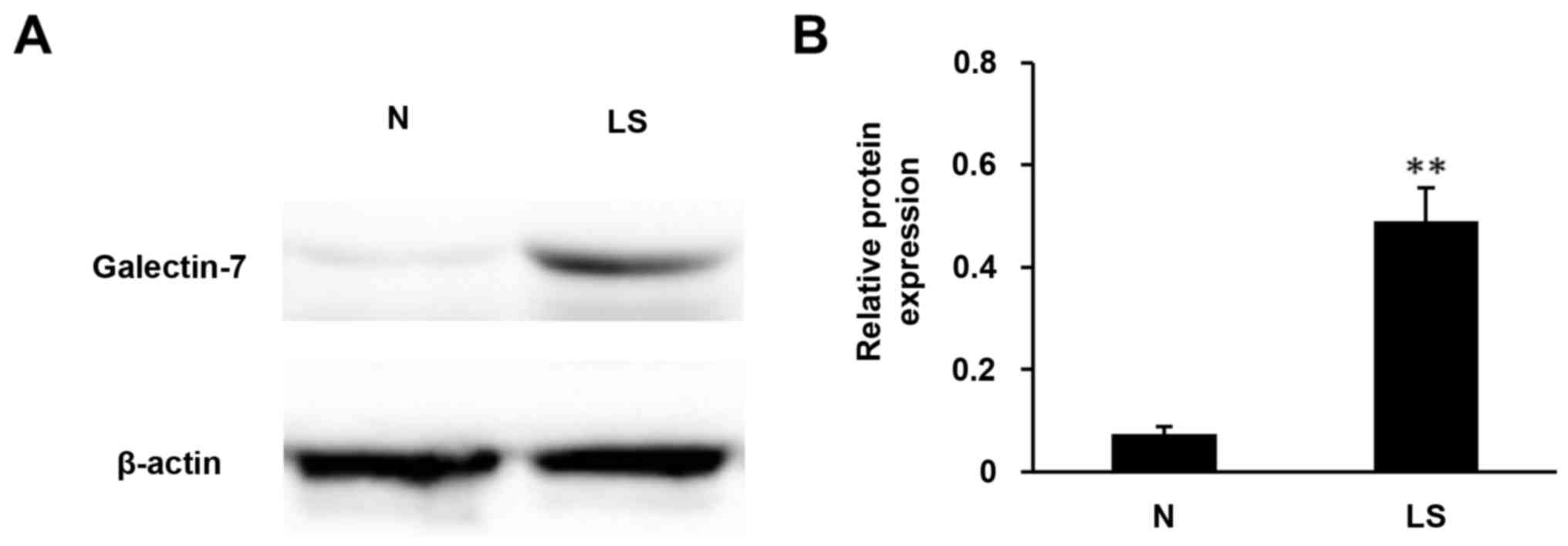
confirm the immunohistochemical staining results, the protein
content was extracted from tissues and western blotting was
performed with the galectin-7 antibody. The results demonstrated
>5-fold upregulation in galectin-7 expression in VLS tissue
compared with healthy tissue (Fig. 2;
P<0.01). These data indicated a positive association between
galectin-7 expression and VLS.
Galectin-7 inhibits the viability of
primary dermal fibroblast cells
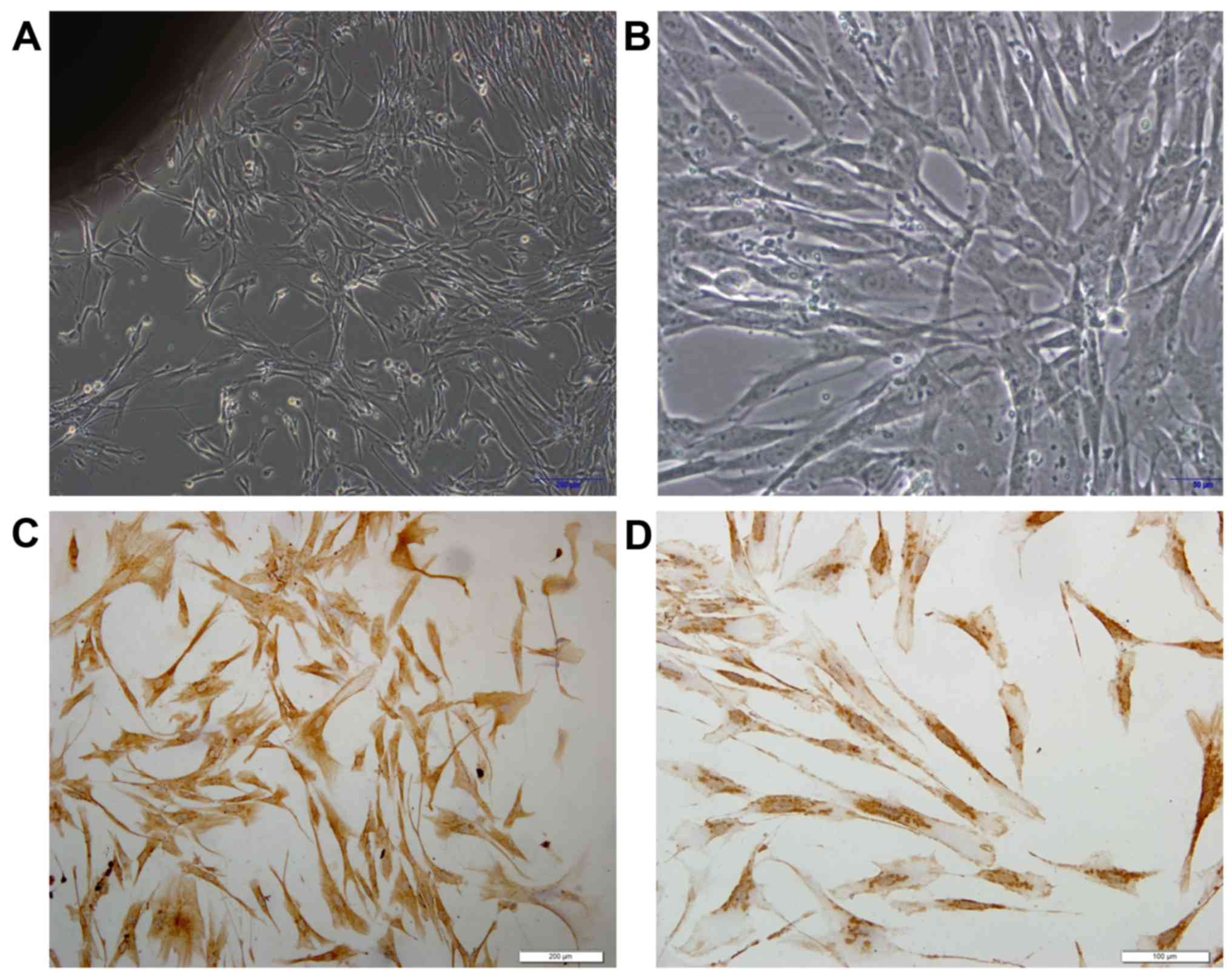
To determine whether galectin-7 influences vulvar
dermis fibroblasts, primary dermal fibroblast cells were isolated
from healthy control biopsy tissue. Phase contrast microscopy
demonstrated spindle shaped fibroblasts, whereby the cells were
bipolar and refractile, with enlarged nuclei (Fig. 3A and B). To confirm that the cells
were fibroblasts, the typical mesoderm marker Vimentin, which is
expressed in fibroblasts was used for staining (9). It was observed that all the isolated
fibroblast cells exhibited Vimentin expression (Fig. 3C and D). Whether galectin-7 serves a
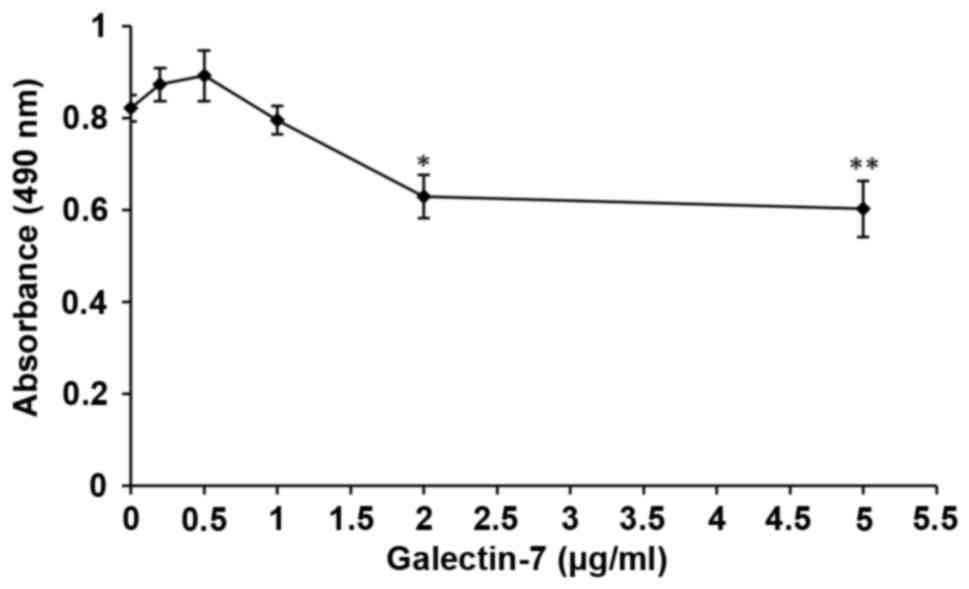
role in the growth rate of the isolated primary dermal fibroblasts
was then investigated. There were significant overall group effects
in the result of the MTS cell viability assay [F(5,12)=7.631; P=0.002]. It revealed that
galectin-7 exerted a significantly inhibitory effect on fibroblasts
at concentrations >1 µg/ml. Compared with that of untreated
fibroblasts, fibroblast viability was significantly impaired at the
concentrations of 2 µg/ml (P<0.05) and 5 µg/ml (P<0.01)
(0.82±0.03 vs. 0.63±0.81 and 0.60±0.11, respectively), indicating
that galectin-7 serves a negative role on the growth of primary
dermal fibroblasts (Fig. 4).
Galectin-7 regulates Collagen I and
collagen III at transcriptional level
A typical feature of VLS is the accumulation and
dis-organization of collagen I and collagen III in the dermal
fibroblasts (10). We hypothesized
that galectin-7 may contribute to VLS through regulating the dermal
fibroblast function and influencing collagen levels. Isolated
primary dermal fibroblasts were seeded in the presence of
galectin-7 and a qPCR analysis was performed to determine the
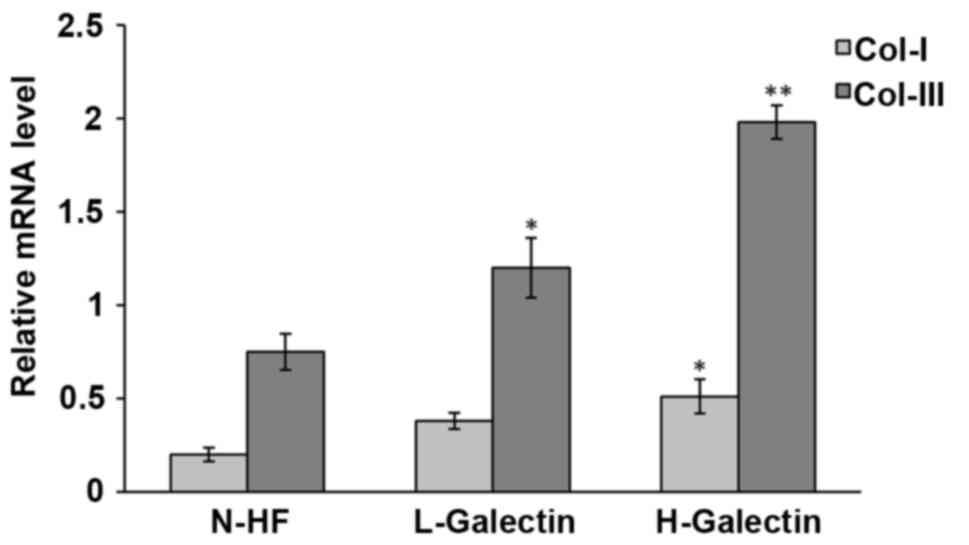
transcriptional level of collagen I and collagen III. The results
demonstrated that the levels of collagen I
[F(2.6)=6.267; P=0.034] and collagen III
[F(2,6)=26.669; P=0.001] were significantly
elevated following the administration of galectin-7 in a
dose-dependent manner (Fig. 5). It
would be interesting to explore in the future whether the
accumulated collagens are irregularly distributed in the cytoplasm
of fibroblasts, and whether they contribute to the hyalinization
encountered in VLS.
Discussion
In the current study, it was observed that
galectin-7 serves important roles in primary dermal fibroblasts,
including an inhibitory role on their growth rate, as well as a
promoter role on the transcription of collagen I and collagen III.
The increased expression of galectin-7 in patients with VLS
significantly influenced the viability of fibroblasts.
Members of the galectin family are recognized for
their ability to bind β-galactosides via a highly conserved
carbohydrate recognition domain. They serve an important role in
several physiological processes, including embryonic development,
intercellular adhesion, host-pathogen interactions, cell migration
and the immune response (11).
Galectin-7 belongs to the galectin family and was initially
described as a marker of epithelial differentiation, due to its
expression in the stratified epithelium of various tissues. It is
induced by p53, thus it is also known as p53-induced gene 1
(12). The induction of galectin-7
triggers the release of cytochrome c and increases the
activity of JUN N-terminal kinase (JNK), triggering programmed
apoptosis (13), which provides
evidence of the inhibitory effect on cell viability. The results of
the present study demonstrated that galectin-7 was significantly
upregulated in VLS tissue from patients compared with healthy
tissue, indicating that a high level of galectin-7 may increase the
apoptosis rate of epidermal cells and inhibit their viability,
leading to the shrinking and loss of epidermal tissue. There is
evidence that the epidermis of extracellular superoxide dismutase
(EC-SOD) transgenic mice produces more galectin-7 compared with
wild type animals, and furthermore the epidermis of EC-SOD
transgenic mice is thinner compared with that in their wild type
counterparts (14). Transgenic mice
were developed specifically overexpressing galectin-7 to aid in
research into the basal epidermal keratinocytes. It was reported
that an excess of galectin-7 causes a destabilization of the
adherens junctions associated with defects in epidermal repair
(15). Another study demonstrated
that galectin-7 is highly expressed in the epidermis of patients
with actinic keratosis, and that galectin-7 upregulates the
apoptosis of the epidermis via T lymphocytes (16). The galectin-7 gene was reported to be
upregulated by 8-fold in bronchial epithelial cells from asthmatic
children following RSV infection in vitro, compared with
tissues from the healthy control patients, indicating that this
protein may be associated with bronchial epithelial cell apoptosis
in asthma (17). These findings
suggest that the overexpression of galectin-7 is associated with
VLS epidermal atrophy.
In our previous study, it was demonstrated that
galectin-7 was downregulated in the early stage of vulvar squamous
cell carcinoma (18). Since it is a
pro-apoptosis protein, this may be associated with the progression
of malignancy. Other reports have revealed a high expression of
galectin-7 in certain cancer cell lines, including in breast cancer
(19), cervical cancer (20), and late and poorly differentiated
vulvar squamous cell carcinoma (18).
Galectin-7 has also been associated with a poor prognosis in
ovarian cancer (21). In summary, the
role of galectin-7 in apoptosis may be organ- or tissue-dependent,
and may be associated with the methylation pattern of galectin-7
itself (22,23).
Due to the multifunctional of the galectin family,
its sub-cellular localization has been extensively studied. Based
on the fact that galectins are able to be detected in intracellular
and extracellular locations, as well as in the plasma of patients
with tumors, recombinant galectins have been used widely in various
in vitro models (11). All
lectins, whether dimers or oligomers, bind to cell surface
receptors, including growth factor receptors and integrins
(24,25). The results of the immunohistochemistry
assay in the present study revealed that galectin-7 was expressed
exclusively in the epidermis. Galectin-7 is specifically
synthesized and secreted by keratinized epithelial cells.
Non-keratinizing cells can only be obtained through membrane
surface receptors (12). In the
immunohistochemistry experiments in the present study, galectin-7
was not detected in the dermis. It was hypothesized that galectin-7
formed in the epidermis can reach the dermis via paracrine and bind
to the surface receptor of dermal fibroblast cell membrane, which
affects cell function. Therefore, in subsequent cell experiments,
recombinant galectin-7 was added to the culture medium to detect
changes in collagen fibers. A previous study reported that
recombinant galectin-3 binds to the glycol-receptor and stimulates
the expression of matrix metalloproteinase 9 (26). We hypothesized that the elevated
galectin-7 expression in the VLS epidermis may be released to the
dermal part of the skin and bind to fibroblasts, and thus,
galectin-7 may modulate the function of dermal fibroblasts.
In the dermis layer, dermal fibroblasts generate
connective tissue allowing the skin to recover from injury. One of
the primary functions of dermal fibroblasts is to produce proteins
which form the extracellular matrix (27); among these proteins, collagen I and
collagen III are the main collagens generated (28). To the best of our knowledge, no
suitable animal models for Lichen sclerosus have been produced
previously (29). Therefore, primary
dermal fibroblasts isolated from the healthy control tissues
through biopsy were cultured to mimic the in vivo
progression of VLS. As revealed by the results of the present
study, a high concentration of galectin-7 (>1 µg/ml)
significantly inhibited the viability of fibroblasts. Notably,
exposure to a low dose (<1 µg/ml) slightly promoted the
viability of fibroblasts, although this was not significant. It is
not known whether this is associated with different levels of
galectin-7 expression in the epidermis. Due to the characteristics
of pathological tissues, it is difficult to perform further
research, and there have been no literature regarding any change in
the number of dermal cells in VLS.
In the present study, the expression of collagens
and its association with galectin-7 in primary dermal fibroblasts
were also analyzed. Collagen I and collagen III, the primary
collagens generated by dermal fibroblasts, were upregulated by
galectin-7 in a dose-dependent manner, suggesting that galectin-7
may be an essential regulator for the hyalinization mediated by
fibroblasts in VLS. Thus, galectin-7 has the potential of being a
target in VLS therapy. Galectin-7 might positively regulate the
collagen expression through the JNK pathway since the apoptotic
role of Galcetin-7 is mediated by accelerating the release of
cytochrome C and the upregulation of JNK. Hashimoto et al
(30) reported that tumor growth
factor-β promoted the transdifferentiation of fibroblasts into
myofibroblasts through JNK and that also led to the production of
extracellular matrix proteins, leading to pulmonary fibrosis
(30). In VLS skin, the hyalinized
collagen is found below the epithelium. Whether galectin-7 has a
role in the irregular distribution of collagen fibers still needs
to be clarified. Certain data have indicated that galectin-7
induces the viability of T cells, primarily the cluster of
differentiation 8+ sub-population (31), and possibly function as an accelerant
of T-cell responses (32). A previous
study described the autoimmune phenotype in VLS, including high
levels of Th1-specific cytokines as well as a dense infiltration of
T cells (33). All these data suggest
that galectin-7 may serve a role in the elevated immune response
encountered in VLS.
In conclusion, the present study identified that the
expression of galectin-7 was significantly upregulated in VLS
tissues. In addition, galectin-7 significantly inhibited the
viability of dermal fibroblasts and stimulated the production of
collagen, thus galectin-7 may be a potential drug target for the
treatment of patients with VLS.
Acknowledgements
This manuscript has been edited by the professional
English writing editor, Dr. Ewen MacDonald from the University of
Eastern Finland.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30973190 and
81400202).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ drafted the manuscript and performed the
immunohistochemical and western blot analysis. SZ conducted the
statistical analyses. HL performed the PCR. XQ performed the cell
culturing. XW has provided conceptualization and approved the final
version to be published. All authors made substantial contributions
to this manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of China Medical University (Shenyang, China) and written
informed consent as obtained from all patients.
Consent for publication
Written informed consents from all patients were
obtained. There is no identifying information in the article.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Val I and Almeida G: An overview of lichen
sclerosus. Clin Obstet Gynecol. 48:808–817. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Regauer S, Reich O and Beham-Schmid C:
Monoclonal gamma-T-cell receptor rearrangement in vulvar lichen
sclerosus and squamous cell carcinomas. Am J Pathol. 160:1035–1045.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Birenbaum DL and Young RC: High prevalence
of thyroid disease in patients with lichen sclerosus. J Reprod Med.
52:28–30. 2007.PubMed/NCBI
|
|
4
|
Hantschmann P, Sterzer S, Jeschke U and
Friese K: P53 expression in vulvar carcinoma, vulvar
intraepithelial neoplasia, squamous cell hyperplasia and lichen
sclerosus. Anticancer Res. 25:1739–1745. 2005.PubMed/NCBI
|
|
5
|
Pugliese JM, Morey AF and Peterson AC:
Lichen sclerosus: Review of the literature and current
recommendations for management. J Urol. 178:2268–2276. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fujimoto N, Asano C, Ono K and Tajima S:
Verruciform Xanthoma results from epidermal apoptosis with
galectin-7 overexpression. J Eur Acad Dermatol Venereol.
27:922–923. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cummings RD and Liu FT: Galectins. Cold
Spring Harbor Laboratory Press. 33:2009.
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nishioka M, Venkatesan N, Dessalle K,
Mogas A, Kyoh S, Lin TY, Nair P, Baglole CJ, Eidelman DH, Ludwig MS
and Hamid Q: Fibroblast-epithelial cell interactions drive
epithelial-mesenchymal transition differently in cells from normal
and COPD patients. Respir Res. 16:722015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Godoy CA, Teodoro WR, Velosa AP, Garippo
AL, Eher EM, Parra ER, Sotto MN and Capelozzi VL: Unusual
remodeling of the hyalinization band in vulval lichen sclerosus by
type V collagen and ECM 1 protein. Clinics (Sao Paulo). 70:356–362.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Di Lella S, Sundblad V, Cerliani JP,
Guardia CM, Estrin DA, Vasta GR and Rabinovich GA: When galectins
recognize glycans: From biochemistry to physiology and back again.
Biochemistry. 50:7842–7857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Polyak K, Xia Y, Zweier JL, Kinzler KW and
Vogelstein B: A model for p53-induced apoptosis. Nature.
389:300–305. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barkan B, Cox AD and Kloog Y: Ras
inhibition boosts galectin-7 at the expense of galectin-1 to
sensitize cells to apoptosis. Oncotarget. 4:256–268. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee JS, Lee Y, Jeon B, Jeon Y, Yoo H and
Kim TY: EC-SOD induces apoptosis through COX-2 and galectin-7 in
the epidermis. J Dermatol Sci. 65:126–133. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gendronneau G, Sanii S, Dang T, Deshayes
F, Delacour D, Pichard E, Advedissian T, Sidhu SS, Viguier M,
Magnaldo T and Poirier F: Overexpression of galectin-7 in mouse
epidermis leads to loss of cell junctions and defective skin
repair. PLoS One. 10:e01190312015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamaguchi T, Hiromasa K, Kabashima-Kubo R,
Yoshioka M and Nakamura M: Galectin-7, induced by cis-urocanic acid
and ultraviolet B irradiation, down-modulates cytokine production
by T lymphocytes. Exp Dermatol. 22:840–842. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yin GQ, Zhao SY, Guo SP, Zhao YH, Liu XC
and Jiang ZF: Galectin-7 is associated with bronchial epithelial
cell apoptosis in asthmatic children. Zhonghua Er Ke Za Zhi.
44:523–526. 2006.(In Chinese). PubMed/NCBI
|
|
18
|
Jiang Y, Tian R, Yu S, Zhao YI, Chen Y, Li
H, Qiao Y and Wu X: Clinical significance of galectin-7 in vulvar
squamous cell carcinoma. Oncol Lett. 10:3826–3831. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Grosset AA, Poirier F, Gaboury L and
St-Pierre Y: Galectin-7 expression potentiates her-2-positive
phenotype in breast cancer. PLoS One. 11:e01667312016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Higareda-Almaraz JC, Ruiz-Moreno JS,
Klimentova J, Barbieri D, Salvador-Gallego R, Ly R,
Valtierra-Gutierrez IA, Dinsart C, Rabinovich GA, Stulik J, et al:
Systems-level effects of ectopic galectin-7 reconstitution in
cervical cancer and its microenvironment. BMC Cancer. 16:6802016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim HJ, Jeon HK, Lee JK, Sung CO, Do IG,
Choi CH, Kim TJ, Kim BG, Bae DS and Lee JW: Clinical significance
of galectin-7 in epithelial ovarian cancer. Anticancer Res.
33:1555–1561. 2013.PubMed/NCBI
|
|
22
|
Demers M, Couillard J, Giglia-Mari G,
Magnaldo T and St-Pierre Y: Increased galectin-7 gene expression in
lymphoma cells is under the control of DNA methylation. Biochem
Biophys Res Commun. 387:425–429. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim SJ, Hwang JA, Ro JY, Lee YS and Chun
KH: Galectin-7 is epigenetically-regulated tumor suppressor in
gastric cancer. Oncotarget. 4:1461–1471. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rabinovich GA, Toscano MA, Jackson SS and
Vasta GR: Functions of cell surface galectin-glycoprotein lattices.
Curr Opin Struct Biol. 17:513–520. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Garner OB and Baum LG: Galectin-glycan
lattices regulate cell-surface glycoprotein organization and
signalling. Biochem Soc Trans. 36:1472–1477. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dange MC, Agarwal AK and Kalraiya RD:
Extracellular galectin-3 induces MMP9 expression by activating p38
MAPK pathway via lysosome-associated membrane protein-1 (LAMP1).
Mol Cell Biochem. 404:79–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quan T, Little E, Quan H, Qin Z, Voorhees
JJ and Fisher GJ: Elevated matrix metalloproteinases and collagen
fragmentation in photodamaged human skin: Impact of altered
extracellular matrix microenvironment on dermal fibroblast
function. J Invest Dermatol. 133:1362–1366. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lochner K, Gaemlich A, Sudel KM, Südel KM,
Venzke K, Moll I, Knott A, Stäb F, Wenck H, Döring O, et al:
Expression of decorin and collagens I and III in different layers
of human skin in vivo: A laser capture microdissection study.
Biogerontology. 8:269–282. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Canady J, Karrer S, Fleck M and Bosserhoff
AK: Fibrosing connective tissue disorders of the skin: Molecular
similarities and distinctions. J Dermatol Sci. 70:151–158. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hashimoto S, Gon Y, Takeshita I, Matsumoto
K, Maruoka S and Horie T: Transforming growth Factor-beta1 induces
phenotypic modulation of human lung fibroblasts to myofibroblast
through a c-Jun-NH2-terminal kinase-dependent pathway. Am J Respir
Crit Care Med. 163:152–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rossi NE, Reiné J, Pineda-Lezamit M,
Pulgar M, Meza NW, Swamy M, Risueno R, Schamel WW, Bonay P,
Fernández-Malavé E and Regueiro JR: Differential antibody binding
to the surface alphabetaTCR.CD3 complex of CD4+ and CD8+ T
lymphocytes is conserved in mammals and associated with
differential glycosylation. Int Immunol. 20:1247–1258. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Luo Z, Ji Y, Zhou H, Huang X, Fang J, Guo
H, Pan T and Chen ZK: Galectin-7 in cardiac allografts in mice:
Increased expression compared with isografts and localization in
infiltrating lymphocytes and vascular endothelial cells. Transplant
Proc. 45:630–634. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Terlou A, Santegoets LA, van der Meijden
WI, Heijmans-Antonissen C, Swagemakers SM, van der Spek PJ, Ewing
PC, van Beurden M, Helmerhorst TJ and Blok LJ: An autoimmune
phenotype in vulvar lichen sclerosus and lichen planus: A Th1
response and high levels of microRNA-155. J Invest Dermatol.
132:658–666. 2012. View Article : Google Scholar : PubMed/NCBI
|