Introduction
Gastric cancer is a highly invasive and aggressive
malignancy that ranks the fourth most common cancer and the leading
cause of cancer-associated mortality in China (1,2). The
incidence of gastric cancer is approximately two times higher in
men compared with women. Studies have shown that individuals
infected with Helicobacter pylori have an increased risk of
gastric cancer (3,4) and a poor prognosis in most countries,
with a 5-year relative survival rate <30% (5).
The advancement of modern medicine and technology
has been accompanied with a series of intensive studies on cancer
tumorigenesis. The application of naturally occurring compounds
with anticancer properties can be regulated by a variety of
mechanisms, including crosslinking of DNA strands and immune
responses, induction of cell cycle arrest, which consequently lead
to cell death (6,7). A number of studies indicated that cancer
cell apoptosis could be considered a point of convergence for all
anticancer therapies, and active phytochemicals have a direct role
in promoting apoptosis (8,9).
Pachymic acid (PA) is a lanostane-type triterpenoid
from Poriacocos, which is an important component in
traditional Chinese medicine. Previous studies have shown that PA
possesses anti-emetic, anti-inflammatory and anticancer properties.
It has been demonstrated that PA is able to inhibit the
proliferation and invasion of pancreatic cancer cells by decreasing
MMP-7 expression (10). PA was able
to activate reactive oxygen species (ROS)-dependent JUN N-terminal
kinase mitochondrial and endoplasmic reticulum stress pathways
following cell cycle arrest at G2/M phase and cell apoptosis
occurred in lung cancer cells (11).
PA reduced proliferation and induced apoptosis through inactivation
of AKT signaling and downregulation of AKT downstream protein
expression in prostate cancer cells (12). Another study demonstrated that PA is
able to stimulate glucose uptake by enhancing the expression and
translocation of glucose transporter type 4 (13). However, the therapeutic effects and
potential mechanisms of PA on gastric cancer are poorly understood
and therefore remain to be evaluated.
The present study aimed to examine the hypothesis
that PA is able to have an effect on the viability, cell cycle
progression and apoptosis of SGC-7901 human gastric cancer cells
in vitro. Herein, the effects of PA on DNA synthesis,
mitochondrial function and ROS production, as well as the
expression levels of Bax (BCL2 associated X protein)/B-cell
lymphoma 2 (Bcl-2) ratio, cytochrome c (cyt-c) and caspase-3 and
the activation of Janus kinase 2 (JAK2)/signal transducer and
activator of transcription 3 (STAT3) in SGC-7901 were also
investigated.
Materials and methods
Materials
PA was purchased from Nanjing Zelang Medical
Technological Co. Ltd., (Nanjing, China) and dissolved in dimethyl
sulfoxide (DMSO) at 10 mg/ml and stored at −20°C. Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit was supplied by Biouniquer Technology Co. Ltd.
(Nanjing, Jiangsu, China). Hoechst 33258 was purchased from
Beyotime Institute of Biotechnology (Shanghai, China).
Tetramethylrhodamine, methyl ester (TMRM) was purchased from Santa
Cruz Biotechnology, Inc., (Dallas, TX, USA). The fluorescent probe
dihydroethidium (DHE) was purchased from Vigorous Biotechnology
Beijing Co., Ltd (Beijing, China).
Cell culture
The SGC-7901 human gastric cancer cell line was
obtained from American Type Culture Collection, (Manassas, VA,
USA). SGC-7901 cells were cultured in DMEM medium (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin G and 100 µg/ml streptomycin in a humidified incubator
at 37°C and 5% CO2.
Cell proliferation assay
The Cell Count kit-8 (CCK-8; Signalway Antibody LLC,
College Park, MD, USA) was performed to examine the effects of PA
and AG490 (Selleck Chemicals, Houston, TX, USA) on cell
proliferation. Briefly, SGC-7901 cells were cultured in 96-well
plates to 80% confluence. Different concentrations of PA (20, 40
and 80 µM) and AG490 (100 µM) were added into each well at 12, 24,
48 and 72 h, and then incubated with CCK-8 solution at 37°C as the
manufacturer's instructions followed by further 1 h incubation
under the same incubator conditions. The absorbance was read for
the supernatant of each well at 450 nm using a microplate reader
(Utrao Medical Instrument Co. Ltd., Shanghai, China).
Flow cytometric analysis of cell
cycle
SGC-7901 cells (3×105 cells/well) were
dispensed into 6-well plates and treated with different
concentrations of PA (20, 40 and 80 µM) for 24 h. The cells were
harvested and fixed in 70% ethanol and then stored at −20°C for 24
h. The cells were washed twice with PBS and resuspended in 100 µl
RNase A at 37°C, followed by staining with 50 µg/ml propidium
iodide (PI) for 10 min at room temperature in the dark. The stained
cells were analyzed using a flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). The DNA content at
G0/G1, S and G2/M was analyzed using BD
Accuri C6 software (version, 1.0.264.21; BD Biosciences).
Flow cytometric analysis of
apoptosis
SGC-7901 cells (3×105 cells/well) were
dispensed into 6-well plates and treated with different
concentrations of PA (20, 40 and 80 µM) and AG490 (100 µM) for 24
h. Double staining with Annexin V fluorescein isothiocyanate
(FITC)/PI was performed following the manufacturer's protocol. A
total of 5×104 SGC-7901 cells were collected by
centrifugation at 1,000 × g for 5 min at 25°C and then resuspended
in a binding buffer containing 195 µl Annexin V FITC and 5 µl PI
for 10 min in an ice bath in the dark prior to flow cytometric
analysis.
DNA fragmentation assay
SGC-7901 cells were dispensed into 6-well plates and
treated with a series of concentrations of PA (20, 40 and 80 µM)
for 24 h. Then, the cells were harvested and fixed with 3%
paraformaldehyde for 5 min at room temperature. After dying, the
cells were stained with Hoechst 33258 (10 ml) for 10 min, following
mounting with 20 mM citric acid and 50 mM orthophosphate in 50%
glycerol. The cells were then stored at −20°C prior to analysis.
Following treatment, the cells were washed with PBS, and features
of apoptosis (including condensed and fragmented nuclei) were
evaluated using a fluorescence microscope (DMI3000B; Leica
Microsystems GmbH, Wetzlar, Germany).
Flow cytometric analysis of Dψm and
ROS generation
SGC-7901 cells (3×105 cells/well) were
dispensed into 6-well plates and treated with different
concentrations of PA (20, 40 and 80 µM) for 24 h at 37°C. Dψm was
detected using TMRM. A total of 1×106/ml SGC-7901 cells
were resuspended with 100 nM TMRM, and the cells were incubated in
the dark at 37°C for 15–20 min and analyzed using a flow cytometer.
The intracellular levels of ROS were assessed using the fluorescent
probe DHE. The SGC-7901 cells (1×106/ml) were
resuspended with 50 µM DHE at 37°C for 30 min, and the intensity of
fluorescence was measured using a flow cytometer.
Western blot analysis
The cells (1×106 cell/well) were
dispensed into 6-well plates followed by treatment with PA and
AG490 for the indicated times and lysed using
radioimmunoprecipitation buffer containing protease inhibitor
Beyotime Institute of Biotechnology, Inc.). The protein
concentration was quantified using a bicinchoninic acid protein
assay kit (Thermo Fisher Scientific, Inc.). The proteins (30 µg)
were separated by 10% SDS-PAGE and further transferred onto a
nitrocellulose membrane (EMD Millipore, Billerica, MA, USA).
Following blocking with 5% non-fat milk overnight at 4°C, the
membranes were probed with specific primary antibodies: Polyclonal
rabbit anti-Bax (1:1,000; cat. no. sc493; Santa Cruz Biotechnology,
Inc.), polyclonal rabbit anti-Bcl-2 (1:1,000; cat. no. sc-492;
Santa Cruz Biotechnology, Inc.), polyclonal mouse anti-cyt-c
(1:1,000; cat. no. ab13575; Abcam, Cambridge, MA, USA) and
monoclonal rabbit anti-caspase-3 (1:500; cat. no. ab44976; Abcam)
overnight at 4°C. GAPDH was used for the normalization of each
protein to ensure the loading of equal quantities of protein. After
washing three times with TBST for 15 min, the blots were incubated
with goat anti-rabbit or goat anti-mouse horseradish
peroxidase-conjugated secondary antibodies (1:1,000; cat. no. A0208
and A0216; Beyotime Institute of Biotechnology) in TBST for 1 h at
room temperature. Following another round of washing, the signals
were detected by enhanced chemiluminescence method (Pierce; Thermo
Fisher Scientific, Inc.) and quantified by densitometry (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Tumor xenograft model
In order to clarify the role of PA in vivo,
5-week-old male athymic nude mice (n=24) were used in the present
study. A total of 2×106 SGC-7901 cells were injected
subcutaneously into the left armpit of these nude mice to establish
the gastric cancer xenograft model. A total of 24 male athymic nude
mice were randomly assigned to four different groups with six mice
per group: Vehicle control (0.1% DMSO in physiological saline) and
PA (10, 30 and 60 mg/kg). Following 1–2 weeks of tumor formation,
the tumor size was determined every 4 days as previously described
(14). The mice were sacrificed, and
the tumors were weighed on a digital balance following
intraperitoneal injection of PA every day for 4 weeks. All the
experiments were approved by the Animal Ethics Committee at Gongli
Hospital of Shanghai Pu Dong New District (Shanghai, China).
Statistical analysis
The data was presented as the mean ± standard
deviation, and significant differences between two groups were
analyzed with the unpaired, two-tailed Student t-test. One-way
analysis of variance, followed by Tukey's post hoc test, was used
to analyze the significance of differences between more than two
groups. Statistical analyses were performed using GraphPad Prism
software (version 5.0; GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
PA inhibits the viability of SGC-7901
gastric cancer cells in vitro
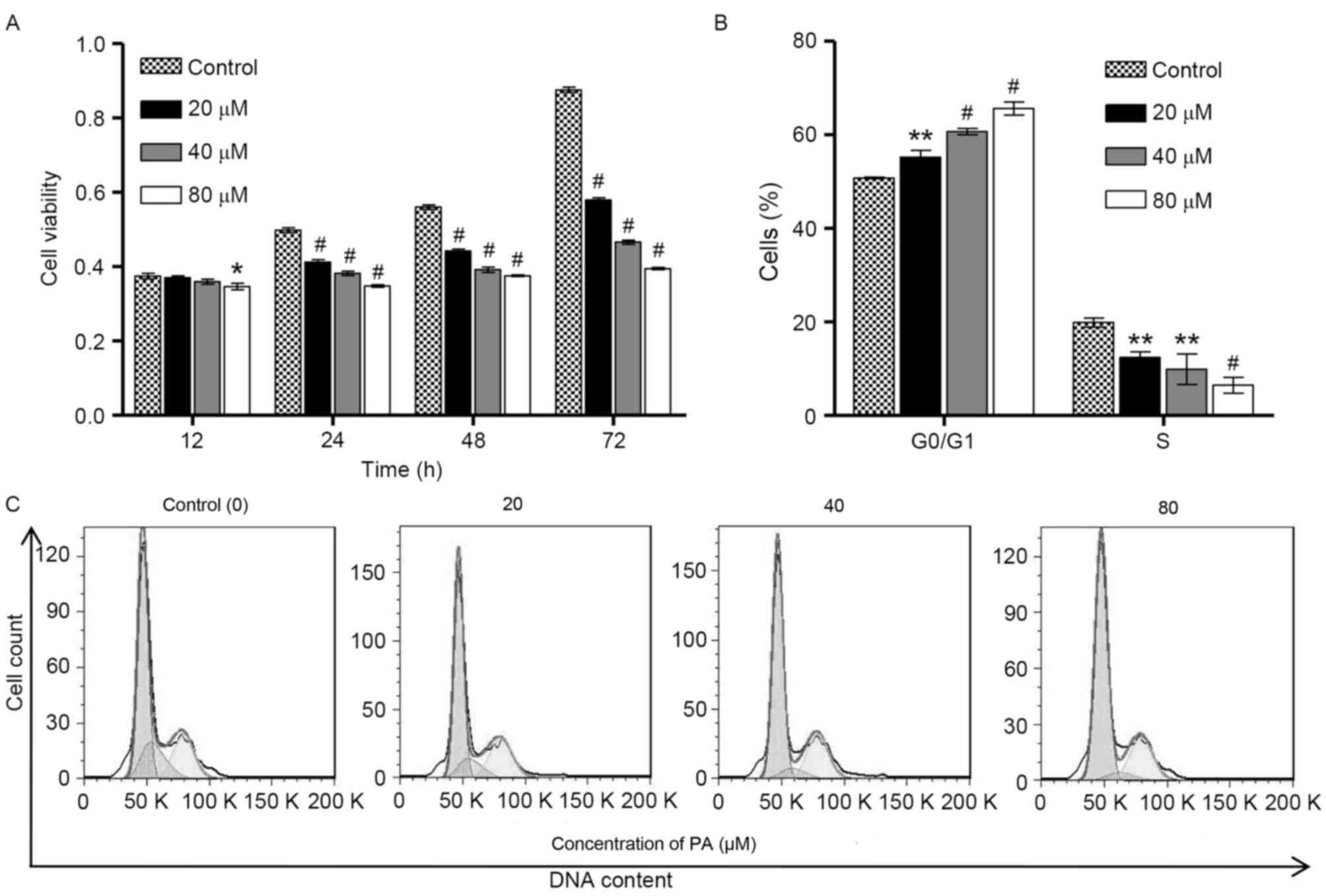
To examine the biological function of PA in gastric
cancer, CCK-8 assay was performed to investigate the viability of
SGC-7901 cells following exposure to different concentrations of PA
(0, 20, 40 and 80 µM) for 12, 24, 48 and 72 h. As shown in Fig. 1A, PA induced significant decreases in
the viability of SGC-7901 cells in a time- and
concentration-dependent manner. To examine whether the inhibitory
effect of PA on the viability of SGC-7901 cells was associated with
the induction of cell cycle arrest, flow cytometry was performed on
the SGC-7901 cells that were treated with different concentrations
of PA (0, 20, 40 and 80 µM) for 24 h. As shown in Fig. 1B and C, the cell population in
G0/G1 phase of SGC-7901 cells was
significantly increased by PA treatment in a
concentration-dependent manner, accompanied with a decrease in the
cell population in S phase, suggesting that PA is able to induce
cell cycle arrest at G0/G1 phase. However, PA
treatment did not affect the cell population at G2/M phase of
SGC-7901 cells. These observations indicate that the inhibition of
cell growth by PA is implicated with the induction of
G0/G1 phase arrest.
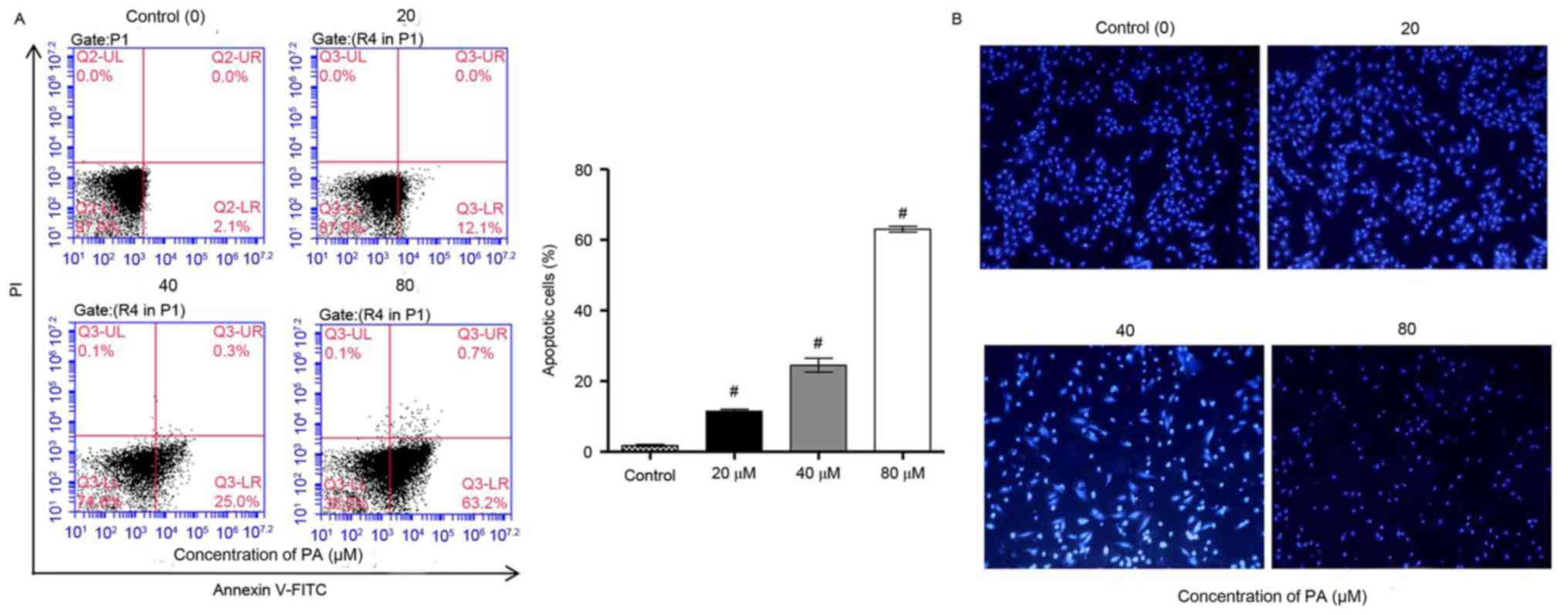
PA induces the apoptosis of SGC-7901
gastric cancer cells in vitro
It was further examined whether the inhibitory
effect of PA on the viability of SGC-7901 cells was associated with
induction of apoptosis. Flow cytometry using Annexin V-PI staining
analysis were performed on the SGC-7901 cells that were treated
with various concentrations of PA (0, 20, 40 and 80 µM) for 24 h.
The data indicated that PA treatment significantly increased the
apoptosis of SGC-7901 cells in a concentration-dependent manner
(Fig. 2A). Furthermore, the effects
of PA on apoptosis were examined by Hoechst 33258 staining, which
is able to differentiate between normal and apoptotic cells. Data
from fluorescence microscopy indicated that PA treatment markedly
increased the apoptosis of SGC-7901 cells in a
concentration-dependent manner, where chromosome condensation and
nuclear fragmentation were observed in PA-treated SGC-7901 cells
(Fig. 2B). Taken together, these
results indicated that the inhibition of cell growth and the
induction of cell cycle arrest by PA are implicated with the
induction of apoptosis.
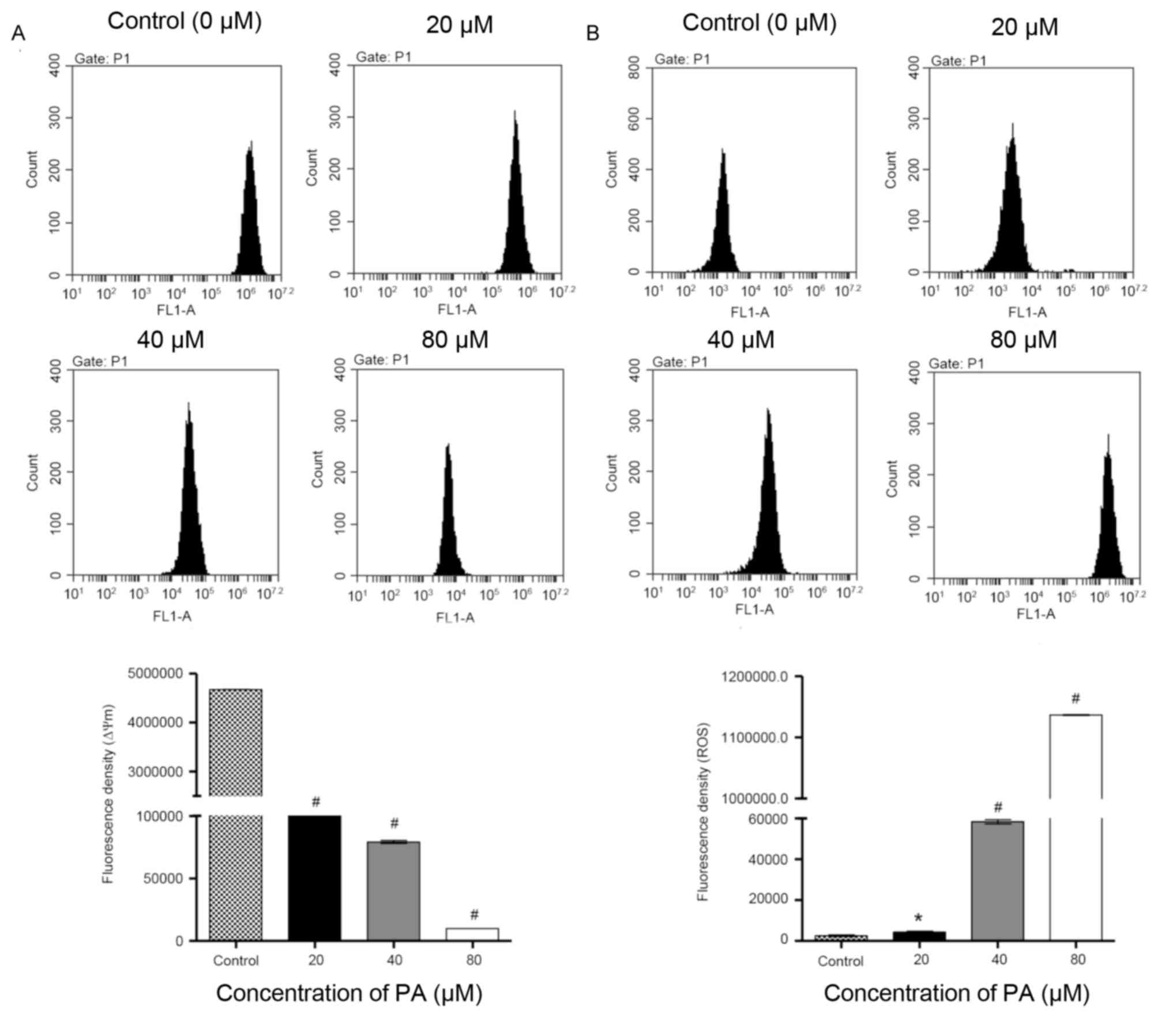
Effects of PA on mitochondrial
membrane potential (Δψm) and ROS generation
Mitochondria have an essential role in death signal
transduction during the apoptotic process, where the mitochondrial
membrane pores are opened and Δψm is disrupted (15). As presented in Fig. 3A, data from flow cytometry indicated
that PA-treated SGC-7901 cells exhibited a significant decrease in
Δψm in a concentration-dependent manner in comparison with the
control cells. Previous studies have reported that ROS generation
is able to trigger cell apoptosis by activating the mitochondrial
pathway (16). Fluorescence probe DHE
was therefore used in SGC-7901 cells that were treated with
different concentrations of PA (0, 20, 40 and 80 µM) for 24 h. As
indicated in Fig. 3B, the treatment
with PA significantly increased the intracellular accumulation of
ROS in a concentration-dependent manner. Taken together, these data
demonstrate that induction of cell apoptosis by PA is implicated
with the mitochondrial pathway.
Effects of PA on protein expression
and JAK2/STAT3 activation in SGC-7901 gastric cancer cells
To elucidate the mechanism of PA-induced apoptosis
in SGC-7901 cells, the expression of apoptosis-associated proteins
was detected by western blotting. As indicated in Fig. 4A, the treatment with PA (20, 40 and 80
µM) for 48 h increased the levels of Bax, cyt-c and
caspase-3proteins in a concentration-dependent manner in comparison
with the control cells, but decreased the expression level of
Bcl-2. Moreover, the inactivation of JAK2 and STAT3 was also
detected in SGC-7901 cells that were treated with different
concentrations of PA (Fig. 4B). Taken
together, these data indicate that induction of cell apoptosis by
PA is implicated with modulation of Bcl-2, Bax, cyt-c and caspase-3
expression as well as the inactivation of JAK2/STAT3 signaling.
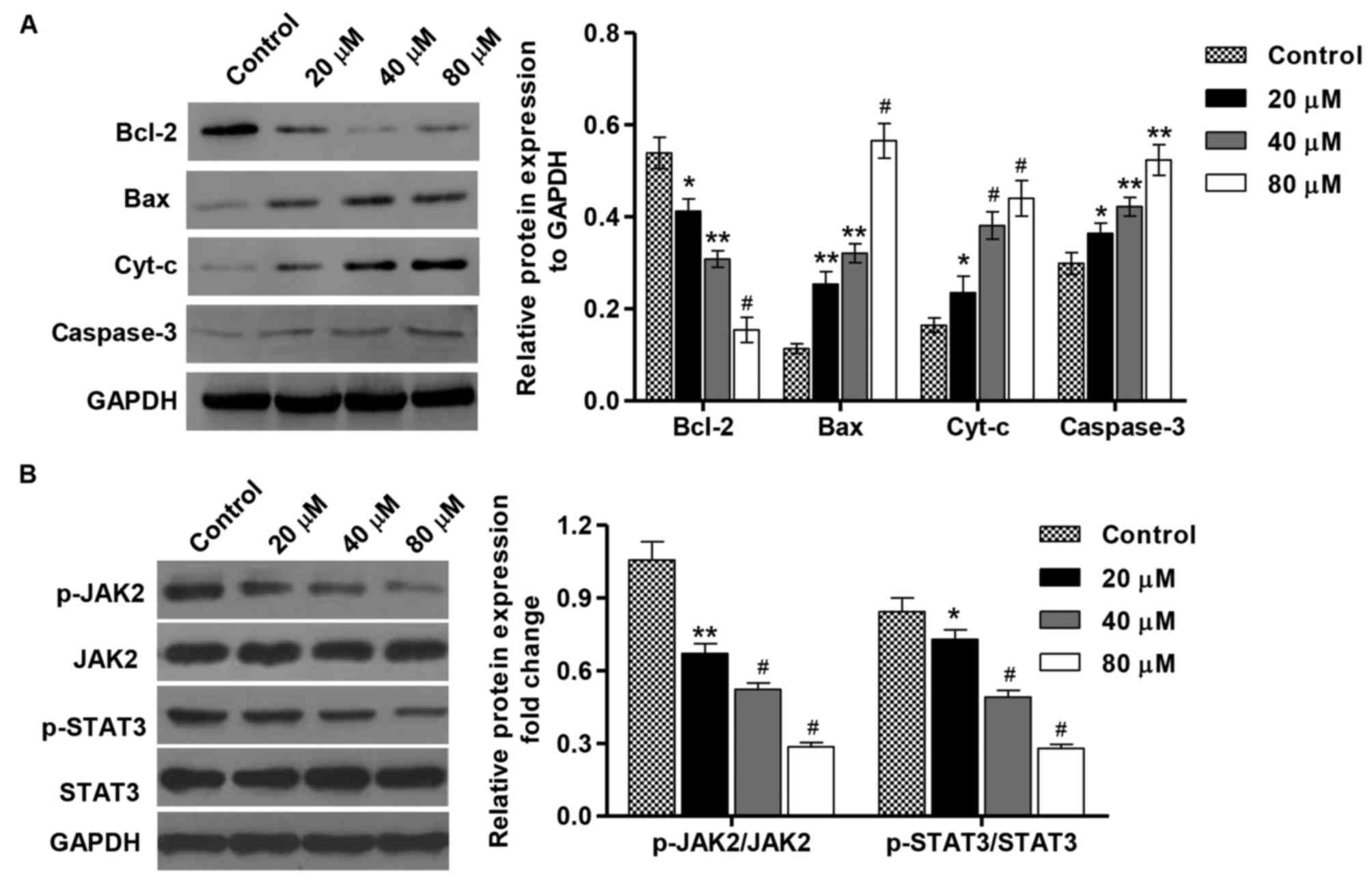 | Figure 4.Effect of PA on protein expression and
JAK2/STAT3 activation in SGC-7901 gastric cancer cells. SGC-7901
cells were treated with different concentrations of PA (20, 40 and
80 µM), and protein expression of (A) Bcl-2, Bax, cyt-c and
caspase-3. (B) The activation of JAK2 and STAT3 was assessed by
western blotting. These experiments were performed on the same
membrane. *P<0.05, **P<0.01, #P<0.001 vs. the
control group. Bax, BCL2 associated X protein; Bcl-2, B-cell
lymphoma 2; cyt-c, cytochrome c; JAK2, Janus kinase 2; p-,
phosphorylated; STAT3, signal transducer and activator of
transcription 3; PA, pachymic acid. |
Inhibition of the JAK2/STAT3 signaling
pathway inhibits cell viability and induces apoptosis in SGC-7901
gastric cancer cells
To confirm that the PA-induced tumor inhibitory
effects are dependent on the downstream activation of JAK2/STAT3,
the effects of a specific JAK2 inhibitor (AG490) on the viability
and apoptosis of cells that were treated with AG490 (100 µM) and in
the absence or presence of PA (80 µM) were determined. The
treatment of SGC-7901 cells with AG490 (100 µM) for 1 h resulted in
the inhibition of JAK2 and STAT3 activation (Fig. 5A). Furthermore, the treatment with
AG490 alone or in combination with PA significantly inhibited the
viability and induced the apoptosis of SGC-7901 cell compared with
the control (Fig. 5B-D). Consistent
with these findings, AG490 treatment in SGC-7901 cells resulted in
increases in the levels of Bax, cyt-c and caspase-3 proteins in
comparison with the control cells, but the expression level of
Bcl-2 was decreased (Fig. 5E). These
data suggest that PA is able to inhibit cell viability and induce
apoptosis via inactivation of the JAK2/STAT3 signaling pathway in
SGC-7901 cells.
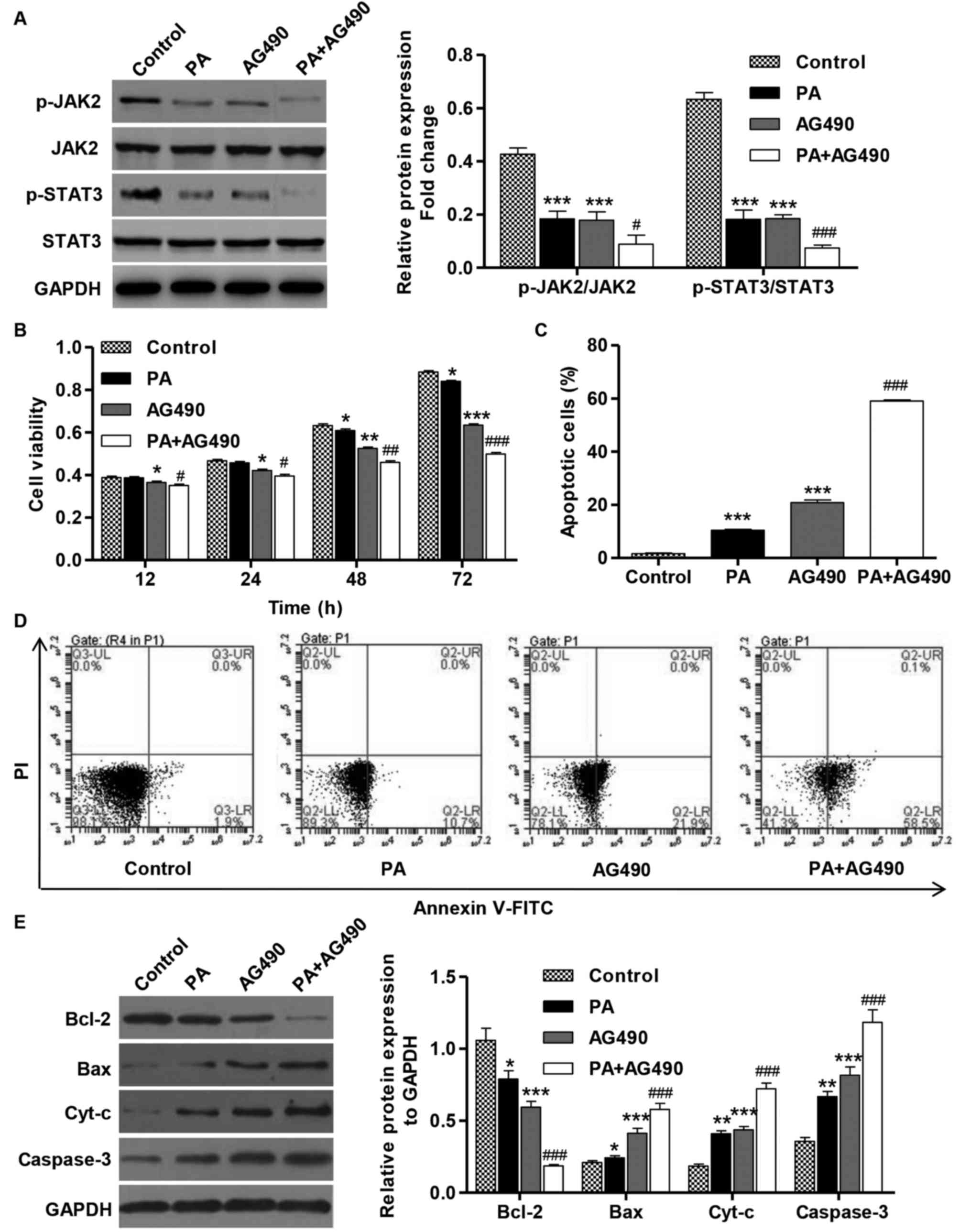 | Figure 5.AG490 inhibits cell viability and
induces apoptosis of SGC-7901 gastric cancer cells. SGC-7901 cells
were treated with PA (80 µM) in the absence or presence of AG490
(100 µM). (A) The activation of JAK2 and STAT3 was analyzed by
western blotting. (B) CCK-8 assay was performed to examine cell
viability. (C) Annexin V-FITC/PI staining was used to detect cell
apoptosis. (D) Representative cell apoptosis profiles of SGC-7901
cells after PA and/or AG490 treatment. (E) The expression of Bcl-2,
Bax, cyt-c and caspase-3 proteins was analyzed by western blotting.
These experiments were performed on the same membrane. *P<0.05,
**P<0.01 and ***P<0.001 vs. the control group.
#P<0.05, ##P<0.01,
###P<0.001 vs. PA or AG490 groups. Bax, BCL2
associated X protein; Bcl-2, B-cell lymphoma 2; cyt-c, cytochrome
c; FITC, fluorescein isothiocyanate; JAK2, Janus kinase 2; PA,
pachymicacid; p-, phosphorylated; PI, propidium iodide; STAT3,
signal transducer and activator of transcription 3. |
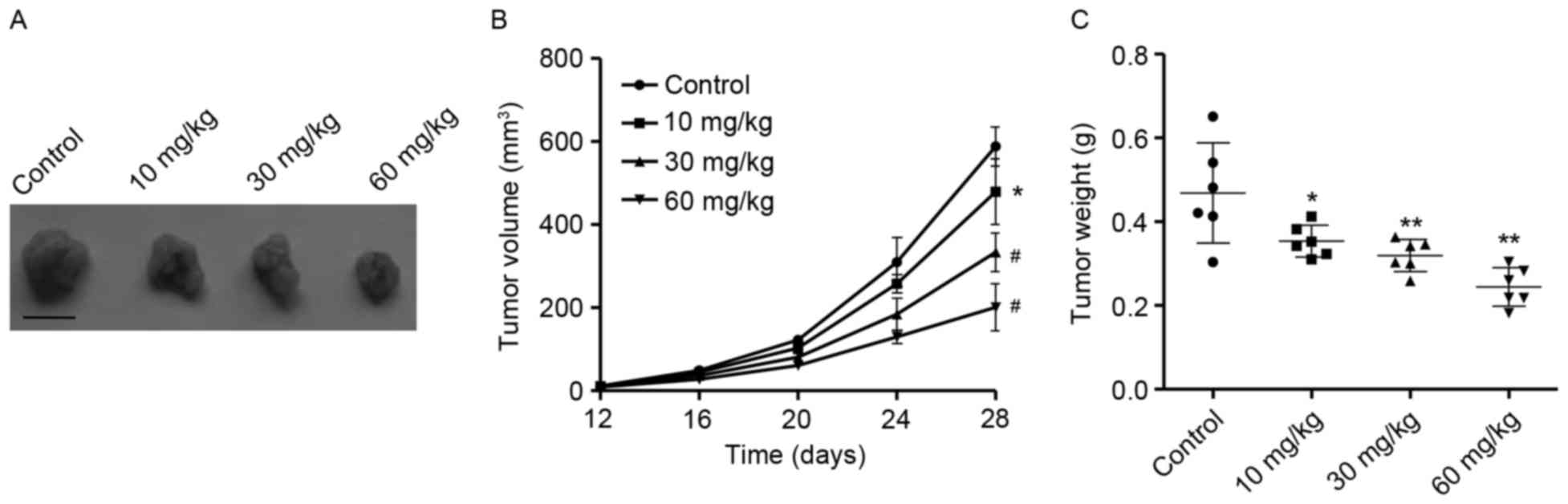
Antitumor activity of PA in vivo
In order to elucidate the antitumor activity of PA
in vivo, tumor xenograft models with SGC-7901 cells were
employed. As presented in Fig. 6A and
B, PA treatment at concentrations of 10, 30 and 60 mg/kg
significantly suppressed tumor growth following 28 days in a
concentration-dependent manner in comparison with the control
group. Moreover, 28 days following treatment with various
concentrations of PA, tumor weight was significantly decreased in a
concentration-dependent manner in comparison with the control group
(Fig. 6C). These results suggest that
PA exhibited potent antitumor activity in vivo.
Discussion
Although chemotherapy is the primary method for the
treatment of gastric cancer, the overall survival rate for patients
with advanced gastric cancer remains low (5). Natural products have a critical role as
an effective source of antitumor agents. Although PA is a promising
bioactive molecule, which possesses an anti-carcinogenic activity
in numerous types of cancer (12,17,18), the
biological activity of PA in gastric cancer is poorly understood.
In present study, the data indicated that PA was able to inhibit
cell viability in a concentration- and time-dependent manner.
Previous studies have shown that cell cycle arrest and apoptosis
are the mechanisms that induce cell death (19). In the present study, it was observed
that treatment with PA was able to induce
G0/G1 phase arrest and apoptosis in SGC-7901
gastric cancer cells. In agreement with our results reported that
PA induced cell cycle arrest and apoptosis in lung cancer cells
(11) and gallbladder cancer cells
(20).
ROS are metabolites with high activity produced
during normal cellular metabolism. However, the increase in
intracellular ROS in cells may be sufficient to induce apoptosis
(21). Furthermore, it has been
reported that cell apoptosis is preceded by the following: ROS
generation, loss of Δψm, release of cyt-c and activation of
caspase-3 (22). Referring to the
results of a previous study and the present study, ROS may be
implicated in PA-induced activation of the mitochondrial pathway
(11). A decrease in Δψm and
increased intracellular accumulation of ROS were associated with
PA-induced activation of the Bax/Bcl-2 signaling pathway.
The expression level of Bax is an important
indicator of cell apoptosis, which indicated the involvement of the
mitochondrial signaling pathway PA-induced apoptosis of SGC-7901
cells. Once activated, Bax is inserted into the mitochondrial
membrane, which leads to mitochondrial dysfunction (23). The data of the present study indicated
that the expression of Bcl-2 decreased gradually and the expression
of Bax in the PA treatment group was increased in a
concentration-dependent manner. Bcl-2 regulates apoptosis mainly by
preventing the release of cyt-c from the mitochondrion to
cytoplasm, activating downstream caspases and ultimately caspase-3
(24). In the present study,
caspase-3 activation was coincident with cyt-c release, which
suggests that the activation of caspase-3 is involved
inthemitochondrial apoptotic pathway, which was observed in
SGC-7901 cells that were treated with PA.
The pathogenic role of JAK/STAT signaling pathway
has been documented in a number of cancer types, including breast,
ovarian, lung, colorectal and gastric cancer (25–29). In
the present study, treatment with PA also resulted in the
inactivation of JAK2 and STAT3 in SGC-7901 cells in a
dose-dependent manner, suggesting an important role of the
JAK2/STAT3 signaling pathway in mediating the inhibitory effects of
PA on tumorgrowth. These observations are consistent with previous
findings by the present authors where OPB-31121, a novel small
molecular inhibitor, was demonstrated to disrupt the JAK2/STAT3
signaling pathway and to exhibit antitumor activity in gastric
cancer cells (29).
A xenograft model with SGC-7901 cells in nude mice
was used to validate the chemotherapeutic potential of PA on the
growth of gastric cancer cells in vivo. To the best of our
knowledge, this is the first time that the in vivo
anti-proliferative activity of PA has been demonstrated, which
further supports the concept that the antitumor effects of PA are
strongly dependent on its ability to inhibit tumor growth. This is
in line with previous findings that 25 mg/kg PA was able to
significantly suppress the growth of pancreatic cancer tumor in
vivo without toxicity, which was also associated with the
inhibition of cell proliferation and apoptosis (30).
In conclusion, the present study demonstrated that
treatment with PA was able to result in marked anticancer activity
in gastric cancer cells by inhibiting cell proliferation as well as
inducing cell cycle arrest and apoptosis via the inactivation of
the JAK2/STAT3 signaling pathway. Therefore, PA appears to be a
potentially attractive bioactive phytochemical for the treatment of
gastric cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author contributions
KS and HX conceived and designed the experiments,
and wrote the paper. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All the experiments were approved by the Animal
Ethics Committee at Gongli Hospital of Shanghai Pu Dong New
District (Shanghai, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Aggarwal A, Guo DL, Hoshida Y, Yuen ST,
Chu KM, So S, Boussioutas A, Chen X, Bowtell D, Aburatani H, et al:
Topological and functional discovery in a gene coexpression
meta-network of gastric cancer. Cancer Res. 66:232–241. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Giordano A and Cito L: Advances in gastric
cancer prevention. World J Clin Oncol. 3:128–136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Islami F and Kamangar F: Helicobacter
pylori and esophageal cancer risk: A meta-analysis. Cancer Prev Res
(Phila). 1:329–338. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kamangar F, Qiao YL, Blaser MJ, Sun XD,
Katki H, Fan JH, Perez-Perez GI, Abnet CC, Zhao P, Mark SD, et al:
Helicobacter pylori and oesophageal and gastric cancers in a
prospective study in China. Br J Cancer. 96:172–176. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zheng L, Wu C, Xi P, Zhu M, Zhang L, Chen
S, Li X, Gu J and Zheng Y: The survival and the long-term trends of
patients with gastric cancer in Shanghai, China. BMC Cancer.
14:3002014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ko JK, Leung WC, Ho WK and Chiu P: Herbal
diterpenoidsinduce growth arrest and apoptosis in colon cancer
cells with increased expression of the nonsteroidal
anti-inflammatory drug-activated gene. Eur J Pharmacol. 559:1–13.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yen CY, Chiu CC, Chang FR, Chen JY, Hwang
CC, Hseu YC, Yang HL, Lee AY, Tsai MT, Guo ZL, et al:
4beta-Hydroxywithanolide E from Physalisperuviana (golden berry)
inhibits growth of human lung cancer cells through DNA damage,
apoptosis and G2/M arrest. BMC Cancer. 10:462010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shiezadeh F, Mousavi SH, Amiri MS,
Iranshahi M, Tayarani-Najaran Z and Karimi G: Cytotoxic and
apoptotic potential of Rheum turkestanicumJanisch root extract on
human cancer and normal cells. Iran J Pharm Res. 12:811–819.
2013.PubMed/NCBI
|
|
9
|
Rajput S, Kumar BP, Dey KK, Pal I, Parekh
A and Mandal M: Molecular targeting of Akt by thymoquinone promotes
G 1 arrest through translation inhibition of cyclin D1 and induces
apoptosis in breast cancer cells. Life Sci. 93:783–790. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng S, Eliaz I, Lin J, Thyagarajan-Sahu
A and Sliva D: Triterpenes from Poria cocos suppress growth and
invasiveness of pancreatic cancer cells through the downregulation
of MMP-7. Int J Oncol. 42:1869–1874. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma J, Liu J, Lu C and Cai D: Pachymic acid
induces apoptosis via activating ROS-dependent JNK and ER stress
pathways in lung cancer cells. Cancer Cell Int. 15:782015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gapter L, Wang Z, Glinski J and Ng KY:
Induction of apoptosis in prostate cancer cells by pachymic acid
from Poriacocos. Biochem Biophys Res Commun. 332:1153–1161. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang YC, Chang WL, Huang SF, Lin CY, Lin
HC and Chang TC: Pachymic acid stimulates glucose uptake through
enhanced GLUT4 expression and translocation. Eur J Pharmacol.
648:39–49. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tomayko MM and Reynolds CP: Determination
of subcutaneous tumor size in athymic (nude) mice. Cancer Chemother
Pharmacol. 24:148–154. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ly JD, Grubb D and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qu K, Shen NY, Xu XS, Su HB, Wei JC, Tai
MH, Meng FD, Zhou L, Zhang YL and Liu C: Emodin induces human T
cell apoptosis in vitro by ROS-mediated endoplasmic reticulum
stress and mitochondrial dysfunction. Acta Pharmacol Sin.
34:1217–1228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hong R, Shen MH, Xie XH and Ruan SM:
Inhibition of breast cancer metastasis via PITPNM3 by pachymic
acid. Asian Pac J Cancer Prev. 13:1877–1880. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ling H, Zhang Y, Ng KY and Chew EH:
Pachymic acid impairs breast cancer cell invasion by suppressing
nuclear factor-κB-dependent matrix metalloproteinase-9 expression.
Breast Cancer Res Treat. 126:609–620. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
King K and Cidlowski J: Cell cycle
regulation and apoptosis. Ann Rev Physiol. 60:601–617. 1998.
View Article : Google Scholar
|
|
20
|
Chen Y, Lian P, Liu Y and Xu K: Pachymic
acid inhibits tumorigenesis in gallbladder carcinoma cells. Int J
Clin Exp Med. 8:17781–17788. 2015.PubMed/NCBI
|
|
21
|
Curtin JF, Donovan M and Cotter TG:
Regulation and measurement of oxidative stress in apoptosis. J
Immunol Methods. 265:49–72. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Herrera B, Fernández M, Alvarez AM,
Roncero C, Benito M, Gil J and Fabregat I: Activation of caspases
occurs downstream from radical oxygen species production, Bcl-xL
down-regulation, and early cytochrome C release in apoptosis
induced by transforming growth factor beta in rat fetal
hepatocytes. Hepatology. 34:548–556. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Green DR and Chipuk JE: Apoptosis: Stabbed
in the BAX. Nature. 455:1047–1049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu X, Zhang K, Wang Q, Chen S, Gou Y, Cui
Y and Li Q: Cisplatin-mediated c-myc overexpression and cytochrome
c (cyt c) release result in the up-regulation of the death
receptors DR4 and DR5 and the activation of caspase 3 and caspase
9, likely responsible for the TRAIL-sensitizing effect of
cisplatin. Med Oncol. 32:1332015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marotta LL, Almendro V, Marusyk A,
Shipitsin M, Schemme J, Walker SR, Bloushtain-Qimron N, Kim JJ,
Choudhury SA, Maruyama R, et al: The JAK2/STAT3 signaling pathway
is required for growth of CD44(+)CD24(−) stem cell-like breast
cancer cells in human tumors. J Clin Invest. 121:2723–2735. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Colomiere M, Ward AC, Riley C, Trenerry
MK, Cameron-Smith D, Findlay J, Ackland L and Ahmed N: Cross talk
of signals between EGFR and IL-6R through JAK2/STAT3 mediate
epithelial-mesenchymal transition in ovarian carcinomas. Br J
Cancer. 100:134–144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao M, Gao FH, Wang JY, Liu F, Yuan HH,
Zhang WY and Jiang B: JAK2/STAT3 signaling pathway activation
mediates tumor angiogenesis by upregulation of VEGF and bFGF in
non-small-cell lung cancer. Lung Cancer. 73:366–374. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiong H, Du W, Zhang YJ, Hong J, Su WY,
Tang JT, Wang YC, Lu R and Fang JY: Trichostatin A, a histone
deacetylase inhibitor, suppresses JAK2/STAT3 signaling via inducing
the promoter-associated histone acetylation of SOCS1 and SOCS3 in
human colorectal cancer cells. Mol Carcinog. 51:174–184. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim MJ, Nam HJ, Kim HP, Han SW, Im SA, Kim
TY, Oh DY and Bang YJ: OPB-31121, a novel small molecular
inhibitor, disrupts the JAK2/STAT3 pathway and exhibits an
antitumor activity in gastric cancer cells. Cancer Lett.
335:145–152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng S, Swanson K, Eliaz I, McClintick
JN, Sandusky GE and Sliva D: Pachymic acid inhibits growth and
induces apoptosis of pancreatic cancer in vitro and in vivo by
targeting ER stress. PloS One. 10:e01222702015. View Article : Google Scholar : PubMed/NCBI
|