Introduction
Renal cell carcinoma (RCC) is the most common tumor
arising in the kidney and accounts for 2–3% cases among all types
of cancer. In western countries, RCC has risen by ~2% during the
last two decades (1). An epidemiology
study indicated that RCC (particularly clear cell RCC) exhibits a
moderate male predilection (2).
Previous studies demonstrated that the occurrence, development and
prognosis of RCC are associated with specific genes, including von
Hippel-Lindau tumor suppressor (3)
and polybromo-1 (4).
MicroRNAs (miRNAs/miRs) are a class of conservative
and small non-coding RNAs, which negatively regulate the expression
level of target genes by binding to sequences in 3′-untranslated
regions (5). Previous studies
indicated that miRNAs are implicated in the recurrence, development
and prognosis of various types of cancer. Several miRNAs may be
deregulated in RCC. For example, miR-15a was significantly
downregulated in RCC tissues compared with adjacent normal tissues
(6). Furthermore, Li et al
(6) demonstrated that the
downregulation of miR-15a may suppress cell proliferation and
invasion by directly targeting eukaryotic initiation factor 4E
during RCC progression. Additionally, miR-149-5p may act as a tumor
suppressor in cellular migration, invasion and apoptosis in RCC,
whereas miR-142-3p may serve an oncogenic function (7,8).
miR-425-5p is located on human chromosome 3 and is
aberrantly expressed in various types of cancer, including gastric
(9), cervical (10) and hepatocellular carcinoma (11). Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) analysis indicated an
overexpression of miR-425-5p in RCC tissues, thus suggesting its
potential use as a biomarker for the diagnosis of RCC (12,13).
However, the molecular mechanisms underlying the effects miR-425-5p
in RCC remain to be elucidated. In the present study, not only the
expression of miR-425-5p was detected in RCC tissues and cell lines
but also the function of miR-425-5p was investigated in RCC cell
lines in vitro.
Materials and methods
Specimens
A total of 24 paired RCC and adjacent normal tissue
specimens (5 cm distance from the tumor margin) were obtained from
the Department of Urology, Peking University Shenzhen Hospital
(Shenzen, China) between January 2013 and December 2013. The
patients that were recruited to the study met the following main
conditions: i) Men or women with age between 18–75 and diagnosis of
renal cell carcinoma by pathology; ii) normal function of the main
organs; iii) subjects volunteered to join the study, signed the
informed consent form, had good compliance and cooperated with the
follow-up; iv) no history of other malignant tumors, and serious,
uncontrolled concomitant diseases that may affect the compliance to
the study or interfere with the interpretation of the results of in
the past 5 years. Patients were excluded when participation in the
study was likely to be associated with a greater risk to the
patient, and when other severe illnesses or laboratory
abnormalities that could interfere with the interpretation of the
results of the study were apparent.
After surgery, all specimens were immersed in
RNAlater® RNA Stabilization Reagent (Qiagen GmbH,
Hilden, Germany) and stored at −80°C. All patients signed written
informed consent. The study was approved by the Ethics Committee of
Peking University Shenzhen Hospital. The clinicopathological
features of patients are presented in Table I (14,15).
 | Table I.Clinicopathological characteristics
of patients with renal cell carcinoma. |
Table I.
Clinicopathological characteristics
of patients with renal cell carcinoma.
|
Characteristics | Patients, n |
|---|
| Age, years |
|
|
Mean | 51 |
|
Range | 27–72 |
| Sex |
|
|
Male | 18 |
|
Female | 6 |
| Histological
type |
|
| Clear
cell | 20 |
|
Papillary | 4 |
| Fuhrman grade
(14) |
|
| I | 15 |
| II | 7 |
|
III | 1 |
| IV | 1 |
| AJCC clinical stage
(15) |
|
| I | 15 |
| II | 8 |
|
III+IV | 1 |
RT-qPCR
Total RNA was isolated from tissues using TRIzol
reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA),
according to the manufacturer's instructions. RNA was purified
using RNeasy Maxi kit (Qiagen GmbH). The concentration of RNA was
measured using a NanoDrop 2000c spectrophotometer (Thermo Fisher
Scientific, Inc.). RNA was reverse-transcribed into cDNA using a
miScriptII RT kit (Qiagen GmbH), according to the manufacturer's
instructions. qPCR was performed using miScript SYBR-Green PCR kit
(Qiagen GmbH) and the Roche LightCycler 480 Real-Time PCR System
(Roche Diagnostics GmbH, Mannheim, Germany), according to the
manufacturer's instructions. The thermocycling conditions were as
follows: Initial activation step at 95°C for 15 min, followed by 40
cycles, denaturation at 94°C for 15 sec, annealing at 55°C for 30
sec and extension at 72°C for 30 sec. U6 was used as an internal
control. Relative expression values were calculated using the
2−ΔΔCq method (16).
Cell culture
HK2 and RCC (ACHN, 786O, Caki-1 and 769P) cell lines
were obtained from the Guangdong and Shenzhen Key Laboratory of
Male Reproductive Medicine and Genetics (Shenzhen, China). ACHN
cells were cultured in Dulbecco's modified Eagle's medium (DMEM;
Thermo Fisher Scientific, Inc.). 786-O and 769P cells were cultured
in RPMI-1640 medium (Thermo Fisher Scientific, Inc.). Caki-1 cells
were cultured in McCoy's 5A modified medium (Thermo Fisher
Scientific, Inc.). Cells were grown in media supplemented with 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 1%
glutamine and 1% antibiotics (100 U/ml penicillin and 100 mg/ml
streptomycin; Gibco; Thermo Fisher Scientific, Inc.) at 37° in a
humidified atmosphere containing 5% CO2.
Cell transfection
miRNA mimics may enhance the regulation of
endogenous miRNAs, whereas miRNA inhibitors inhibit the function of
miRNA (17). The number of cells
(5×103-1×106 cells/well) depended on the
volume of the plate. Transfection using miR-425-5p mimic and
miR-425-5p inhibitor may upregulate and downregulate the expression
level of miR-425-5p, respectively, according to the manufacturer's
protocol (Table II), ACHN and 786-O
cells were cultured in 96, 48, 24 or 6-well plates and treated with
5, 20, 40 or 100 pmol small interfering RNA (siRNA) using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.) at
37° in a humidified atmosphere containing 5% CO2,
respectively. Prior to adding DMEM, cells were incubated with
Opti-MEM medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented without 10% FBS and 1% glutamine for 6 h. The
transfection efficiency was analyzed using RT-qPCR. The sequences
of miR-425-5p mimic, inhibitor, negative control (NC) mimic, NC
inhibitor (Shanghai GenePharma Co., Ltd., Shanghai, China) are
presented in Table III.
| Table II.Concentration of the miRNA
mimic/inhibitors used for transfection. |
Table II.
Concentration of the miRNA
mimic/inhibitors used for transfection.
| Plate | siRNA, µl | Final volume,
µl |
Lipofectamine® 2000, µl |
|---|
| 96-well | 0.5 (5 pmol) | 100 | 0.25 |
| 24-well | 1 (20 pmol) | 500 | 1 |
| 12-well | 2 (40 pmol) | 1,000 | 2 |
| 6-well | 5 (100 pmol) | 2,000 | 5 |
| Table III.Sequences used in the study. |
Table III.
Sequences used in the study.
| Name | Sequence |
|---|
| miR-425-5p | Forward:
5′-AATGACACGATCACTCCCGTTGA-3′ |
|
| Reverse: Universal
primers (miScript SYBR-Green PCR kit) |
| U6 | Forward:
5′-CTCGCTTCGGCAGCACA-3′ |
|
| Reverse:
5′-ACGCTTCACGAATTTGCGT-3′ |
| miR-425-5p
mimic | Forward:
5′-AAUGACACGAUCACUCCCGUUGA-3′ |
|
| Reverse:
5′-AACGGGAGUGAUCGUGUCAUUUU-3′ |
| miR-425-5p
inhibitor |
5′-AGGCGAAGGAUGACAAAGGGAA-3′ |
| NC | Forward:
5′-UUCUCCGAACGUGUCACGUTT-3′ |
|
| Reverse:
5′-ACGUGACACGUUCGGAGAATT-3′ |
| Inhibitor NC |
5′-CAGUACUUUUGUGUAGUACAA-3′ |
Cell counting kit-8 (CCK-8)
A CCK-8 assay (Beyotime Institute of Biotechnology,
Haimen, China) was used to assess the proliferative ability of
786-O and ACHN cells. Cells (5,000 cells/well) were seeded in
96-well plates and then transfected with miR-425-5p mimic,
inhibitor, NC mimic or NC inhibitor (Shanghai GenePharma Co.,
Ltd.). In brief, a total of 10 µl CCK-8 solution was added into
each well and cells were incubated for 30 min at 37°C in a
humidified atmosphere containing 5% CO2. After cells
were incubated for 0, 24, 48 and 72 h, cell proliferation was
determined by measuring the absorbance at 450 nm (with 620 nm as
the reference wavelength) using a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The experiment was
performed in triplicate.
MTT assay
MTT assay (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was used to assess the viability of 786-O and ACHN cells.
A total of 5,000 cells/well were seeded in 96-well plates. A total
of 3 days after transfection, 20 µl MTT (5 mg/ml; Sigma-Aldrich;
Merck KGaA) was added into each well plate for 4 h at 37°C. After
removing supernatant, 100 µl dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA) was added into each well. The cells were
then agitated using a shaker (TSB-108; Qilinbeier, Jiangsu, China)
at 0.16 g/min for 10 min in dark at room temperature. The viability
was evaluated at a wavelength of 595 nm (with 620 nm as the
reference wavelength) using a microplate reader (Bio-Rad
Laboratories, Inc.). The experiment was performed in
triplicate.
Wound healing assay
The wound healing assay was used to assess the
migratory ability of the ACHN and 786-O cells. A total of
2×104 cells were seeded in each well of a 96-well plate.
Prior to transfection, cells were starved for 24 h. Then, cells
were transfected with miR-425-5p mimic, inhibitor, NC mimic or NC
inhibitor as aforementioned. After transfection, cells were
cultured in serum and antibiotics-free media at 37°C for 24 h.
Next, wounds were inflicted by dragging a sterile pipette tip.
Images were captured using a digital camera system at 0, 12 and 24
h at 37°C. The experiment was performed in triplicate.
Transwell assay
The migratory and invasive abilities of the ACHN and
786-O cells were assessed using 24-well Transwell chambers (BD
Biosciences, Franklin Lakes, NJ, USA) with 8 µm pore size inserts.
At 24 h post-transfection, a total of ~ 3×104 cells were
plated in each upper chamber. The lower chamber of the Transwell
contained 600 µl medium (DMEM or RPMI-1640) supplemented with 10%
FBS and 1% glutamine. For the invasion assay, Matrigel-coated
Transwell chambers, according to the manufacturer's instructions.
At 48 h, cells were fixed using 0.1% paraformaldehyde for 25 min
and stained using 4% crystal violet stain for 25 min at room
temperature. Images were captured using a digital camera. The
experiment was performed in triplicate. Cells were counted in 4
randomly selected field of view using a light microscope at
magnification, ×100.
Flow cytometry
Flow cytometry was used to assess the apoptotic rate
in ACHN and 786-O cells. A total of 1×106 cells were
plated in a 6-well plate. At 24 h, cells were transfected with
miR-425-5p mimic, inhibitor, NC mimic or NC inhibitor as
aforementioned. At 48 h post-transfection, cells were harvested and
washed using ice-cold PBS. Next, cells were stained with 5 µl
Annexin V-fluorescein isothiocyanate (FITC; Invitrogen; Thermo
Fisher Scientific, Inc.) and propidium iodide (PI; Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. The apoptotic rate was evaluated using a flow
cytometer (EPICS, Xl-4, Beckman Coulter, Inc., Brea, CA, USA).
FlowJo software version 7.6.1 (FlowJo LLC, Ashland, OR, USA) was
used for analysis of the data. The experiment was performed in
triplicate.
Statistical analysis
Data were analyzed using SPSS software version 23.0
(IBM Corp., Armonk, NY, USA). Data are expressed as the mean ±
standard deviation. The expression level of miR-425-5p in tissues
was analyzed using a paired Student's t-test. The expression level
of miR-425-5p in RCC cell lines was evaluated using one-way
analysis of variance followed by Bonferroni's post hoc test.
Student's t-test was used to analyze the results of between two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
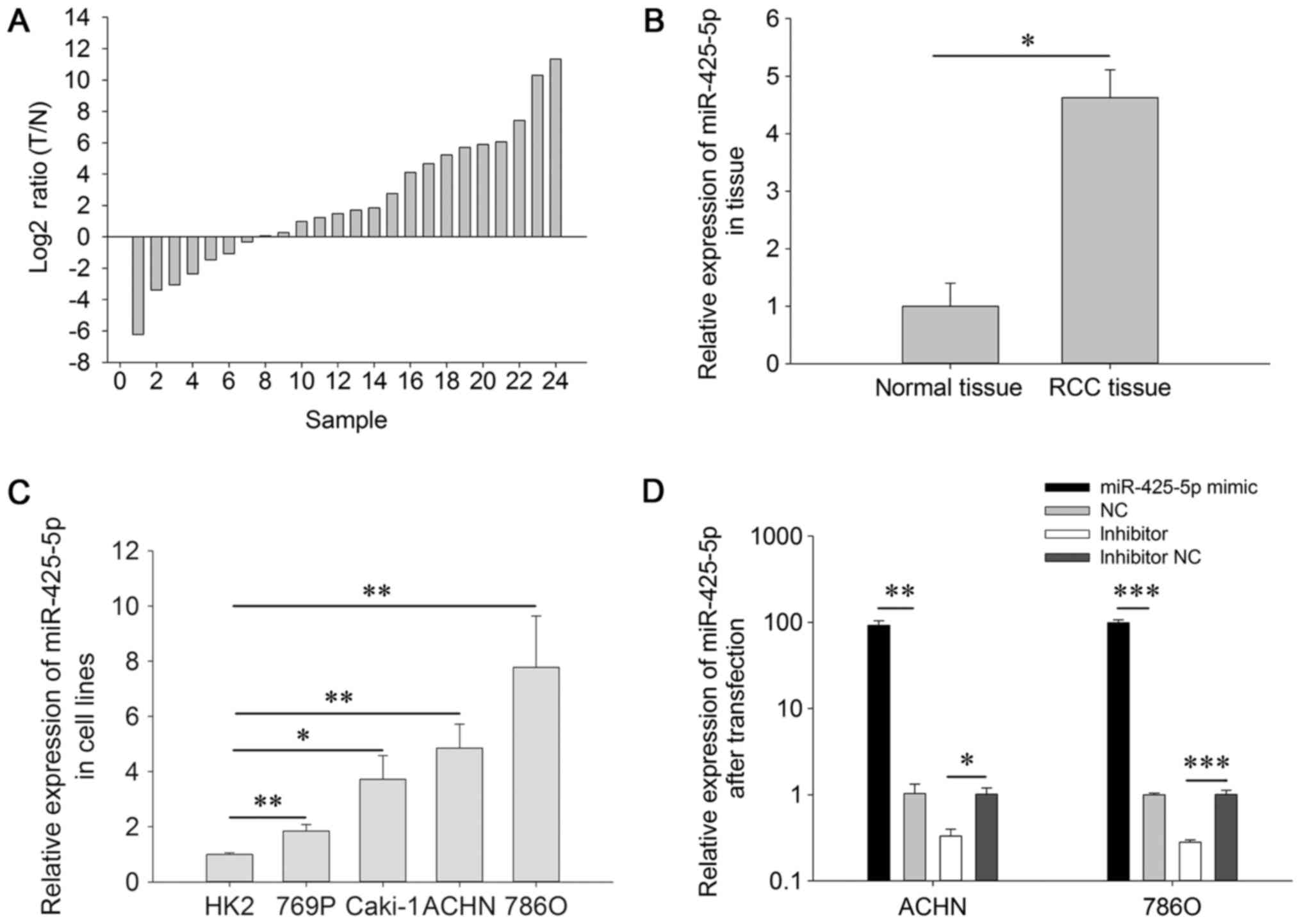
The expression level of miR-425-5p is
upregulated in RCC tissues and cell lines
RT-qPCR was performed to detect the expression of
miR-425-5p in RCC tissues and cell lines. Fig. 1A presents the ratio of miR-425-5p
[log2ratio (Tumor tissue/Normal tissue)] in 24 paired
RCC and adjacent normal tissue specimens. As demonstrated in
Fig. 1B, the expression level of
miR-425-5p was significantly increased in RCC tissues (4.628±0.483)
compared with that in adjacent normal tissues (1.000±0.399;
P=0.020). The expression levels of miR-425-5p in RCC cell lines
were significantly increased compared with those in normal kidney
cells (P<0.01). Therefore, these results suggest that miR-425-5p
may act as an oncogene in RCC.
Cell transfection efficiency
RT-qPCR was performed to detect the expression level
of miR-425-5p after transfection. At 24 h post-transfection, the
expression levels of miR-425-5p were increased by 99.008-fold in
786O cells (P<0.001; Fig. 1D) and
92.137-fold in ACHN cells (P=0.003; Fig.
1D) in response to miR-425-5p mimic compared with NC mimic. At
24 h post-transfection, the expression levels of miR-425-5p were
increased by 0.281 (786O cells; P<0.001) and 0.330-fold (ACHN
cells; P=0.037) in response to miR-425-5p inhibitor compared with
NC inhibitor.
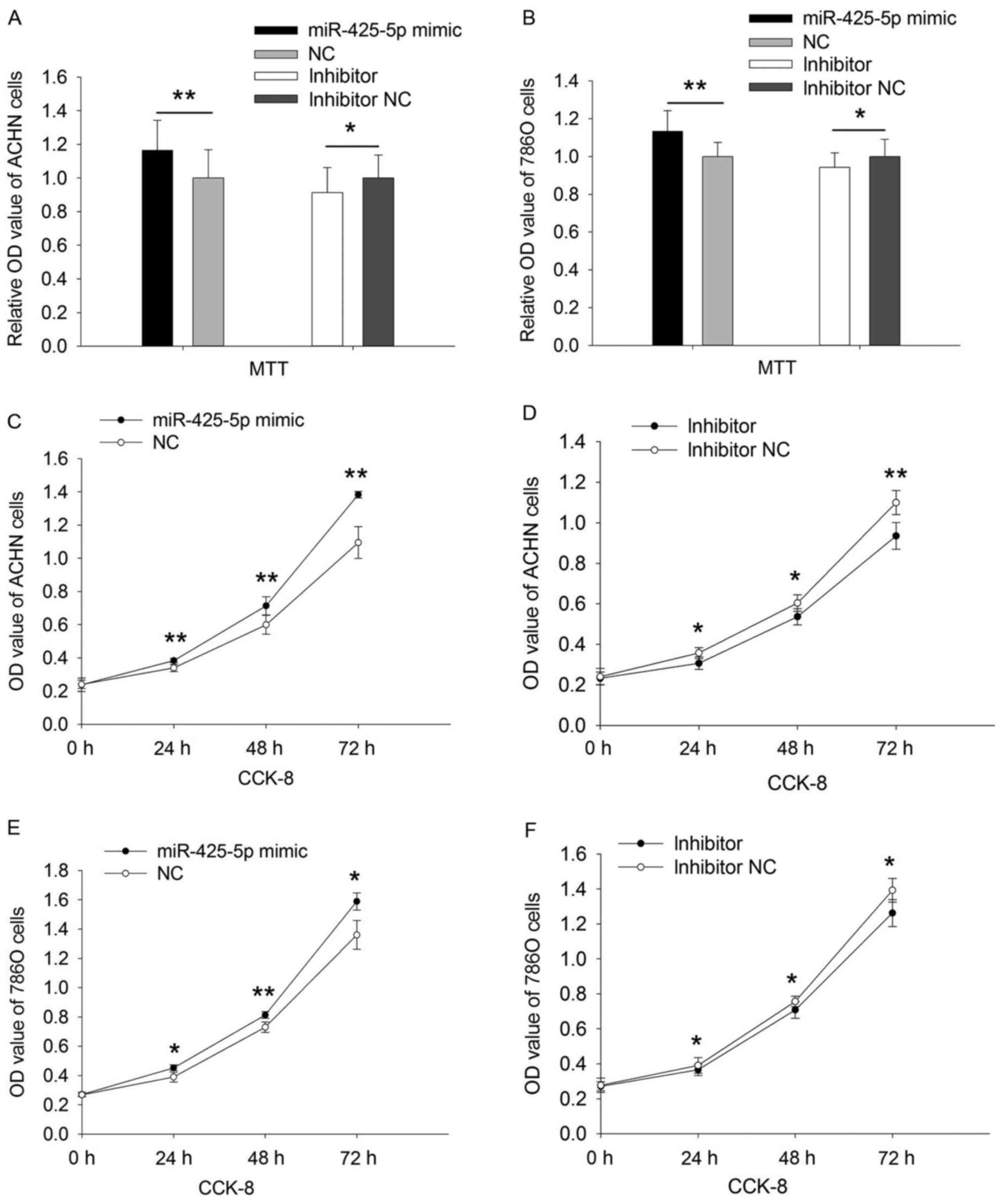
Upregulation or downregulation of
miR-425-5p regulates the viability of ACHN and 786O cells
MTT assay was used to evaluate cell viability. As
presented in Fig. 2, the relative
viability of ACHN cells transfected with the miR-425-5p mimic was
increased compared with that in NC mimic group (P=0.002). The
relative viability of 786O cells transfected with miR-425-5p mimic
was increased compared with that in NC mimic group (P=0.002).
Additionally, the relative viability of ACHN cells transfected with
the miR-425-5p inhibitor was decreased compared with that in the NC
inhibitor group (P=0.048; Fig. 2A).
These results were consistent in 786O cells (P=0.038; Fig. 2B).
Upregulation or downregulation of
miR-425-5p regulates the proliferation of ACHN and 786O cells
CCK-8 assay was used to assess the proliferative
ability of ACHN and 786O cells. At 24, 48 and 72 h
post-transfection, the viability of ACHN cells was upregulated by
12.877 (P=0.009), 19.068 (P=0.003) and 26.441% (P=0.002) in
response to miR-425-5p mimic compared with that in NC mimic group
(Fig. 2C), whereas the viability of
ACHN cells was downregulated by 14.183 (P=0.014), 11.483 (P=0.040)
and 14.948% (P=0.009; Fig. 2D) in
response to miR-425-5p inhibitor compared with that in NC inhibitor
group. Additionally, the viability of 786O cells was upregulated by
15.991 (P=0.034), 11.485 (P=0.006) and 16.863% (P=0.013) in
response to miR-425-5p mimic compared with that in NC mimic group
(Fig. 2E) during those time points
whereas the viability of 786O cells was decreased by 6.299
(P=0.050), 4.443 (P=0.013), 9.344% (P=0.013; Fig. 2F) in response to miR-425-5p inhibitor
compared with that in NC inhibitor group. Therefore, upregulation
or downregulation of miR-425-5p may promote or inhibit,
respectively, the proliferative ability of ACHN and 786O cells.
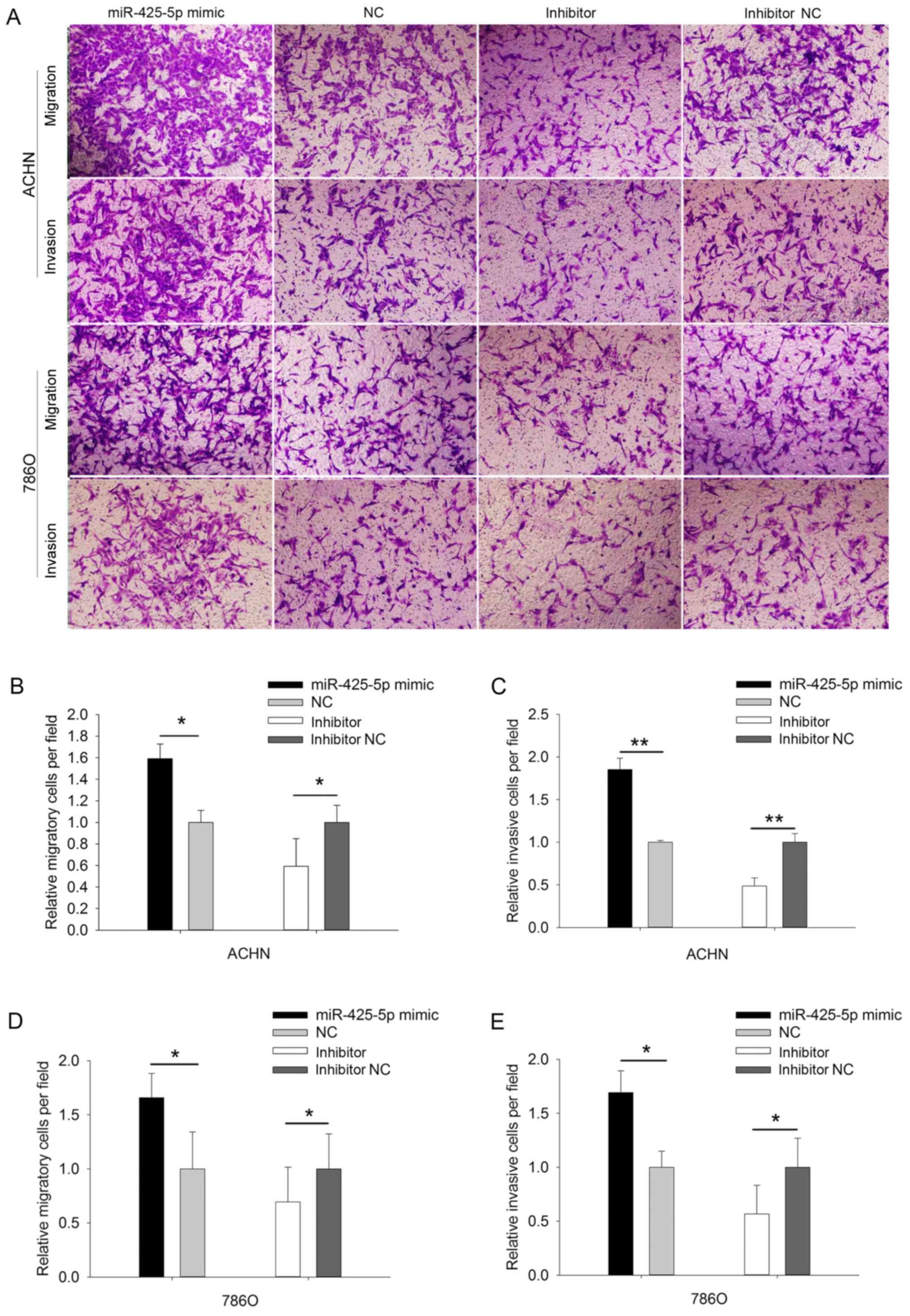
miR-425-5p regulates migration and
invasion in ACHN and 786O cells
The migratory and invasive abilities of ACHN and
786O cells were evaluated using Transwell (Fig. 3A-E) and wound scratch assays (Fig. 4A-C). The results demonstrated that the
migratory ability of the ACHN cells was upregulated (P=0.039;
Fig. 3B) in the miR-425-5p mimic
group compared with that in the NC mimic group. However, their
migratory ability was by 40.738% (P=0.048) in response to treatment
with miR-425-5p inhibitor (Fig. 3B).
Additionally, the invasive ability of the ACHN cells was
upregulated (P=0.001) in the miR-425-5p mimic group, whereas the
invasive ability of the ACHN cells was downregulated (P=0.008) in
miR-425-5p inhibitor group (Fig. 3C).
For the 786O cells, those results were consistent with the above
results of ACHN cells (P<0.05).
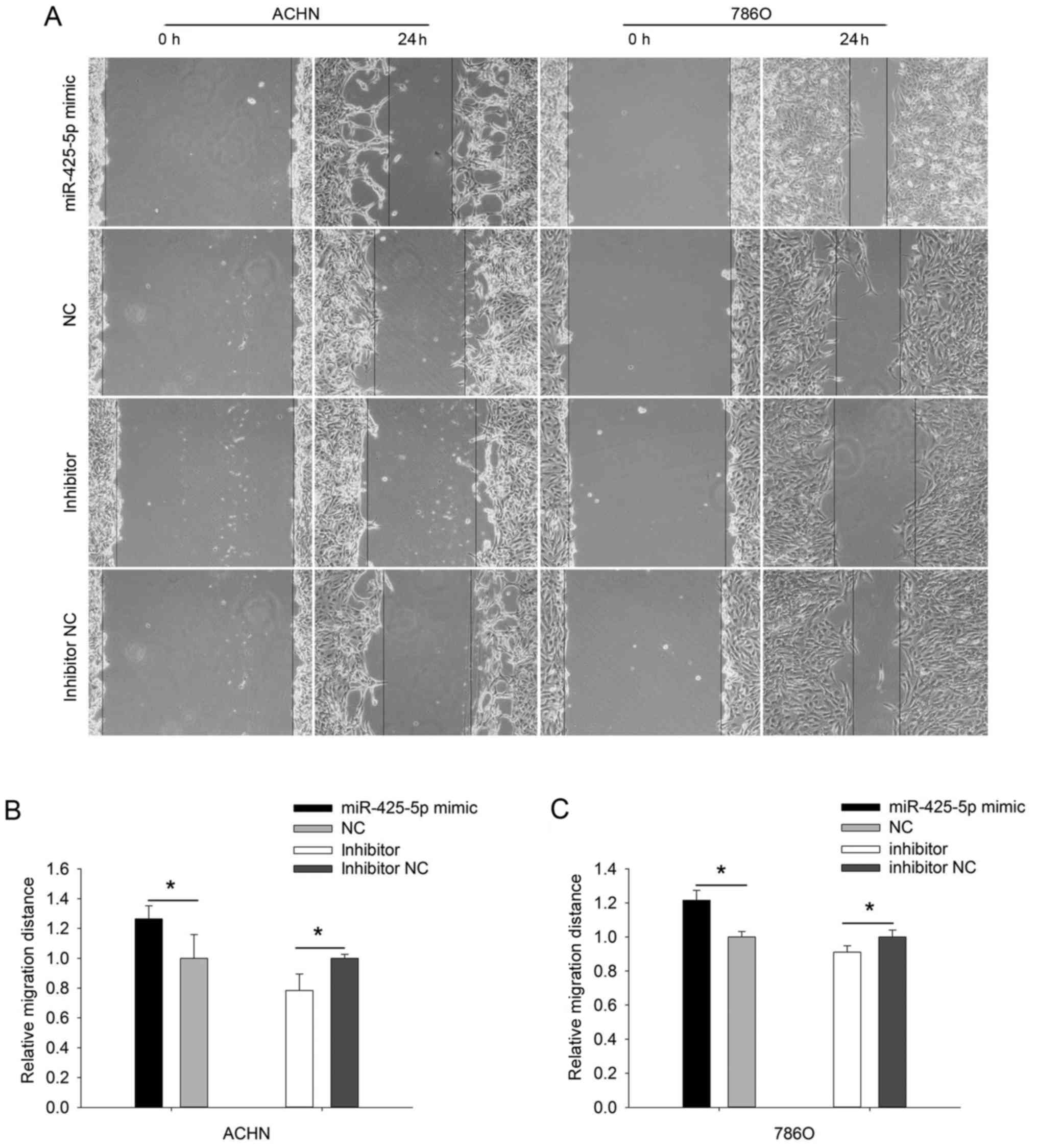
The results of the wound healing assay (Fig. 4A-C) demonstrated that the migratory
ability of the ACHN cells was upregulated (P=0.025) in the
miR-425-5p mimic group, whereas their migratory ability was
downregulated (P=0.048) in the miR-425-5p inhibitor group (Fig. 4B). Those results in 786O cells were
consistent with the above results in ACHN cells (P<0.05).
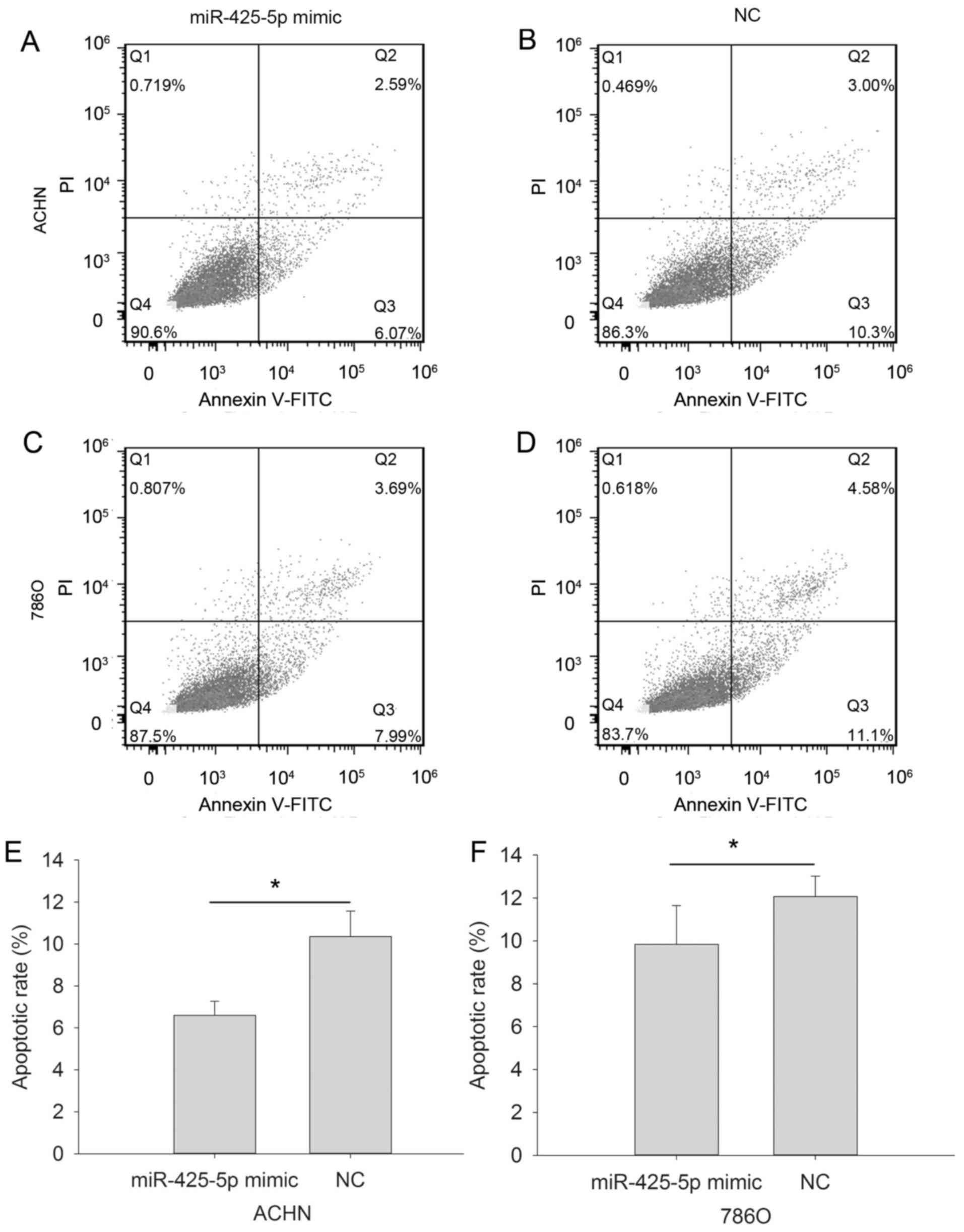
Upregulation of miR-425-5p inhibits
apoptosis in ACHN and 786O cells
Early apoptotic rate was determined using flow
cytometry (Fig. 5A-F). The results
demonstrated that the early apoptotic rate of ACHN cells
transfected with the miR-425-5p mimic was decreased compared with
that of the NC mimic group (6.593±0.671% vs. 10.360±1.211%;
P=0.015; Fig. 5E. The apoptotic rate
of the 786O cells in response to miR-425-5p mimic and NC was
observed to be 9.843±1.807% vs. 12.067±0.950%, respectively
(P=0.046; Fig. 5F). However, no
differences in early apoptotic rate were observed between the
inhibitor and NC inhibitor groups for 786O and ACHN cells were
observed (data not shown).
Discussion
miRNAs are a class of endogenous non-coding RNAs
that serve important functions in tumorigenesis. However, the
underlying molecular mechanism remains unclear (18). Numerous studies have demonstrated that
miRNAs may function as oncogenes or tumor suppressors and may serve
an important biological role in various tumor types (19).
Sun et al (10)
demonstrated that miR-425-5p is upregulated in cervical cancer
tissues compared with matched non-cancerous tissues. Additionally,
miR-425-5p is upregulated in the serum of patients with cervical
cancer, which suggests that it may act as a potential biomarker in
cervical cancer. Zhang et al (9) demonstrated that miR-425-5p is
upregulated in gastric cancer cell lines and may regulate cell
migration and invasion in vitro. These results were also
confirmed by Peng et al (20).
Wang et al (21) revealed that
miR-425 may drive tumor formation and growth, and promote the
progression of lung cancer. Furthermore, Di Leva et al
(22) demonstrated that miR-425 may
promote the expression of epithelial markers by targeting SATB
homeobox 1, cyclin 2 (CCND2) and Fascin actin-bundling protein 1 in
aggressive breast cancer cells. Recently, it was reported that
miR-425-5p may regulate chemoresistance via programmed cell death
10 in colorectal cancer cells lines (23). A study by Fang et al (24) indicated an upregulation of miR-425-5p
in hepatocellular carcinoma tissues, which was associated with poor
clinicopathological characteristics and prognosis. Furthermore,
miR-425-5p may promote metastasis via inhibiting suppressor of
cancer cell invasion (SCAI)-mediated dysregulation of the
transcriptional regulation of integrin β1-Focal Adhesion Kinase 1
(ITGB1-Fak), SRC proto-oncogene-Ras homolog gene family member A
(Src-RhoA)/cell division cycle 42 (CDC42), phosphatase and tensin
homolog (PTEN/AKT)/Murine thymoma viral (v-Akt) oncogene homolog
and TIMP metallopeptidase inhibitor 2-zinc-dependent matrix
metalloproteinases 2/zinc-dependent matrix metalloproteinases 9
(TIMP2-MMP2/MMP9) signaling pathways (24).
miR-425-5p serves an important role in various
diseases. Di Pietro et al (25) demonstrated that miR-425-5p was
significantly downregulated in traumatic brain injury at early
timepoints and may be used as a diagnostic biomarker. A study on
Alzheimer's disease (AD) demonstrated that miR-339 and miR-425 may
be used as potential diagnostic biomarkers for AD. Additionally,
miRNAs may inhibit the β-secretase 1 (BACE1) protein expression
level and are involved in the pathogenesis of AD (26). Gao et al (27) found that the over-expression of
miR-425-5p may reverse the NaAsO2-induced anti-angiogenesis by
directly targeting cerebral cavernous malformation 3.
However, there are several limitations. Firstly, due
to the long storage of specimens and the absence of some clinical
data, the present study was unable to analyze the association
between the expression of mir-425-5p and clinical data. Secondly,
further experiments are required to confirm the results of the
present study. For example, it is recommended to block cell
proliferation using cell cycle inhibitors prior to analyzing the
effect on cell migration instead of hunger, which may strengthen
the results of the present study. Additionally, although miR-425-5p
expression was increased by ~100-fold in RCC cells in response to
miR-425-5p mimic in vitro, the increase was lower (5-fold)
in RCC tissues compared to controls. The regulation of miR-425-5p
expression may be more complex in vivo than in vitro.
Although CCK-8 assay was used to evaluate the proliferative ability
of RCC cells, further assays including 5-Bromo-2-deoxyUridine
(BrdU) and 5-Ethynyl-2′-deoxyuridine (EdU) assays (28), are required to confirm the results of
the present study.
The results of the present study demonstrated that
the expression levels of miR-425-5p were upregulated in RCC cell
lines and tissues compared with controls. Furthermore, the results
of functional assays suggested that miR-425-5p may function as an
oncogene in RCC. Therefore, the molecular mechanisms underlying the
function of miR-425-5p in RCC require further investigation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81101922), the
Science and Technology Development Fund Project of Shenzhen (grant
nos. JCYJ20150403091443329 and JCYJ20170307111334308), the
‘San-ming’ Project of Medicine in Shenzhen and the fund of
Guangdong Key Medical Subject.
Availability of data and materials
The data sets generated during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
YQL conceived and designed the experiments. JQ, YWL,
XP, YLL, TH, CL, LiaZ and LiwZ performed the experiments, analyzed
the data and drafted the paper. SS, YD, LT, YH, XW, ZC, FZ, JY and
LN assisted in collecting tissue specimens and writing the
manuscript. All authors have read and approved this manuscript.
Ethics approval and consent to
participate
All patients signed informed consent forms prior to
the initiation of the present study. The present study was approved
by the Ethical Review Committee of the Peking University Shenzhen
Hospital and complied with the Declaration of Helsinki.
Consent for publication
All patients provided their consent for data sharing
and the publication of any associated images.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ljungberg B, Bensalah K, Canfield S,
Dabestani S, Hofmann F, Hora M, Kuczyk MA, Lam T, Marconi L,
Merseburger AS, et al: EAU guidelines on renal cell carcinoma: 2014
update. Eur Urol. 67:913–924. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Udager AM and Mehra R: Morphologic,
molecular and taxonomic evolution of renal cell carcinoma: A
conceptual perspective with emphasis on updates to the 2016 world
health organization classification. Arch Pathol Lab Med.
140:1026–1037. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gnarra JR, Tory K, Weng Y, Schmidt L, Wei
MH, Li H, Latif F, Liu S, Chen F, Duh FM, et al: Mutations of the
VHL tumour suppressor gene in renal carcinoma. Nat Genet. 7:85–90.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Varela I, Tarpey P, Raine K, Huang D, Ong
CK, Stephens P, Davies H, Jones D, Lin ML, Teague J, et al: Exome
sequencing identifies frequent mutation of the SWI/SNF complex gene
PBRM1 in renal carcinoma. Nature. 469:539–542. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li G, Chong T, Xiang X, Yang J and Li H:
Downregulation of microRNA-15a suppresses the proliferation and
invasion of renal cell carcinoma via direct targeting of eIF4E.
Oncol Rep. 38:1995–2002. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jin L, Li Y, Liu J, Yang S, Gui Y, Mao X,
Nie G and Lai Y: Tumor suppressor miR-149-5p is associated with
cellular migration, proliferation and apoptosis in renal cell
carcinoma. Mol Med Rep. 13:5386–5392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Y, Chen D, Jin LU, Liu J, Li Y, Su Z,
Qi Z, Shi M, Jiang Z and Yang S: Oncogenic microRNA-142-3p is
associated with cellular migration, proliferation and apoptosis in
renal cell carcinoma. Oncol Lett. 11:1235–1241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Z, Li Y, Fan L, Zhao Q, Tan B, Li Z
and Zang A: microRNA-425-5p is upregulated in human gastric cancer
and contributes to invasion and metastasis in vitro and
in vivo. Exp Ther Med. 9:1617–1622. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun L, Jiang R, Li J, Wang B, Ma C, Lv Y
and Mu N: MicoRNA-425-5p is a potential prognostic biomarker for
cervical cancer. Ann Clin Biochem. 54:127–133. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vaira V, Roncalli M, Carnaghi C, Faversani
A, Maggioni M, Augello C, Rimassa L, Pressiani T, Spagnuolo G, Di
Tommaso L, et al: MicroRNA-425-3p predicts response to sorafenib
therapy in patients with hepatocellular carcinoma. Liver Int.
35:1077–1086. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hao JF, Ren KM, Bai JX, Wang SN, Shao B,
Cao N and Li X: Identification of potential biomarkers for clear
cell renal cell carcinoma based on microRNA-mRNA pathway
relationships. J Cancer Res Ther. 10:Suppl. C167–C177. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ge YZ, Xin H, Lu TZ, Xu Z, Yu P, Zhao YC,
Li MH, Zhao Y, Zhong B, Xu X, et al: MicroRNA expression profiles
predict clinical phenotypes and prognosis in chromophobe renal cell
carcinoma. Sci Rep. 5:103282015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang X, Liu Y, Cao G, Zhang X, Xu H, Xu H
and Wang J: Expression of the EphA1 protein is associated with
Fuhrman nuclear grade in clear cell renal cell carcinomas. Int J
Clin Exp Pathol. 8:6821–6827. 2015.PubMed/NCBI
|
|
15
|
Moch H, Artibani W, Delahunt B, Ficarra V,
Knuechel R, Montorsi F, Patard JJ, Stief CG, Sulser T and Wild PJ:
Reassessing the current UICC/AJCC TNM staging for renal cell
carcinoma. Eur Urol. 56:636–643. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qian H, Deng X, Huang ZW, Wei J, Ding CH,
Feng RX, Zeng X, Chen YX, Ding J, Qiu L, et al: An HNF1α-regulated
feedback circuit modulates hepatic fibrogenesis via the crosstalk
between hepatocytes and hepatic stellate cells. Cell Res.
25:930–945. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Inui M, Martello G and Piccolo S: MicroRNA
control of signal transduction. Nat Rev Mol Cell Biol. 11:252–263.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng WZ, Ma R, Wang F, Yu J and Liu ZB:
Role of miR-191/425 cluster in tumorigenesis and diagnosis of
gastric cancer. Int J Mol Sci. 15:4031–4048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Li Z, Ge Q, Wu W, Zhu Q, Luo J and
Chen L: Characterization of microRNA transcriptome in tumor,
adjacent and normal tissues of lung squamous cell carcinoma. J
Thorac Cardiovasc Surg. 149:1404–1414, e1404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Di Leva G, Piovan C, Gasparini P, Ngankeu
A, Taccioli C, Briskin D, Cheung DG, Bolon B, Anderlucci L, Alder
H, et al: Estrogen mediated-activation of miR-191/425 cluster
modulates tumorigenicity of breast cancer cells depending on
estrogen receptor status. PLoS Genet. 9:e10033112013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Hu X, Miao X, Zhu K, Cui S, Meng
Q, Sun J and Wang T: MicroRNA-425-5p regulates chemoresistance in
colorectal cancer cells via regulation of Programmed Cell Death 10.
J Cell Mol Med. 20:360–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang F, Song T, Zhang T, Cui Y, Zhang G
and Xiong Q: MiR-425-5p promotes invasion and metastasis of
hepatocellular carcinoma cells through SCAI-mediated dysregulation
of multiple signaling pathways. Oncotarget. 8:31745–31757.
2017.PubMed/NCBI
|
|
25
|
Di Pietro V, Ragusa M, Davies D, Su Z,
Hazeldine J, Lazzarino G, Hill LJ, Crombie N, Foster M and Purrello
M: MicroRNAs as novel biomarkers for the diagnosis and prognosis of
mild and severe traumatic brain injury. J Neurotrauma.
34:1948–1956. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ren RJ, Zhang YF, Dammer EB, Zhou Y, Wang
LL, Liu XH, Feng BL, Jiang GX, Chen SD, Wang G and Cheng Q:
Peripheral blood microrna expression profiles in alzheimer's
disease: screening, validation, association with clinical phenotype
and implications for molecular mechanism. Mol Neurobiol.
53:5772–5781. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gao Y, Yin Y, Xing X, Zhao Z, Lu Y, Sun Y,
Zhuang Z, Wang M, Ji W and He Y: Arsenic-induced anti-angiogenesis
via miR-425-5p-regulated CCM3. Toxicol Lett. 254:22–31. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Buck SB, Bradford J, Gee KR, Agnew BJ,
Clarke ST and Salic A: Detection of S-phase cell cycle progression
using 5-ethynyl-2′-deoxyuridine incorporation with click chemistry,
an alternative to using 5-bromo-2′-deoxyuridine antibodies.
Biotechniques. 44:927–929. 2008. View Article : Google Scholar : PubMed/NCBI
|