Introduction
Breast cancer is an aggressive malignancy, and is a
major threat to the health of women worldwide. According to the
2018 Global Cancer Statistics, breast cancer is the second leading
cause of cancer-associated mortality worldwide, following lung
cancer. Furthermore, there were ~2.1 million new cases in 2018
worldwide, and these accounted for approximately a quarter of the
total number of female patients with cancer (1). In China, breast cancer is the fifth
leading cause of cancer-associated mortality, according to
statistics collected by the China National Cancer Center in 2018
(2). On the therapy options for
breast cancer, molecular typing is a critical basis. Based on the
expression of estrogen receptor (ER), progesterone receptor (PR),
human epidermal growth factor receptor 2 (HER2) and Ki-67, there
are five primary molecular subtypes of breast cancer: Luminal A,
luminal B, triple-negative/basal like, HER2-enriched and
normal-like (3). Currently,
molecular typing-based integrative treatment regimens consist of
surgery, radiotherapy, chemotherapy, endocrine therapy and targeted
therapy, which have improved treatment efficacy, including
improving overall survival (OS) and progression-free survival
(4,5). However, most patients with cancer still
experience drug resistance, recurrence and metastasis (6). In recent years, CXC chemokine receptor
2 (CXCR2) has emerged as a critical functional receptor. CXCR2
serves an important role in various aspects of breast cancer,
including the diverse range of pathological processes associated
with tumor progression (7). Combined
analysis demonstrated that patients with solid tumors and elevated
CXCR2 expression had poorer prognosis, including OS,
recurrence-free survival and disease-free survival (8). The present review summarizes the
biological roles of CXCR2 in breast cancer and systematically
examines the pathways and mechanisms of CXCR2 associated with the
initiation and development of breast cancer, as well as the
potential therapeutic value of an anti-CXCR2 treatment.
Structure and interactions of the chemokine
CXC and its receptor CXCR2
Chemokines are small (6–14 kDa), secreted peptides
that mediate the migration of leukocytes to inflammation and
secondary lymphoid organs. Chemokines are also essential for other
pathophysiological processes, including infectious diseases,
asthma, and atherosclerosis (9–11). Based
on the position of the four cysteine (Cys) residues at the
N-terminus, the chemokines are divided into four subtypes: CXC (α),
CC (β), XC (γ) and CX3C (δ) (12).
To date, 50 chemokines and 20 chemokine receptors have been
identified; the majority of chemokines belong to the CC and CXC
subgroups (13). CXC is further
classified into ELR+ and ELR− CXC chemokines,
based on the glutamate-leucine-arginine (Glu-Leu-Arg, ELR) sequence
that occurs before the first Cys at the N-terminus (14).
The coding sequence of the CXCR2 gene is located at
2q34-35 and contains three exons and two introns (15). CXCR2 is a G protein-coupled receptor
that contains seven transmembrane regions, an extracellular
N-terminus and an intracellular C-terminus (16). The N-terminus, the fourth
transmembrane domain and the second extracellular loop are
prerequisites for ligand binding and specificity, and determine the
rate of receptor internalization (17). The C-terminus region is involved in
receptor phosphorylation, internalization and G protein coupling.
Only ligand monomers activate CXCR2, which interact via a two-site,
two-step model. This model involves the binding of the N-terminal
domain of CXCR2 with the N-loop and core domain of ligands at site
1. At site 2, CXCR2 activation is conferred by the insertion of the
N-terminus signal domain of the ligands into the orthosteric pocket
of the receptor (18). Interleukin-8
(IL-8), also termed CXCL8, is a CXCR2 ligand and was the first CXC
chemokine derived from the medium of lipopolysaccharide and
polyhydroxyalkanoate-stimulated human monocytes (19). In humans, CXCR2 is also known as IL-8
receptor B and interacts with ELR+CXC chemokines with
high affinity. These chemokines include GRO-α/CXCL1, GRO-β/CXCL2,
GRO-γ/CXCL3, ENA-78/CXCL5, GCP-2/CXCL6, NAP-2/CXCL7 and IL-8/CXCL8,
which mediate angiogenesis (20).
Signaling pathways of CXCR2 activation
CXCR2 possesses no kinase activity. In addition to
being coupled to G proteins, it binds to other proteins, such as G
protein coupled receptor kinase 2/6, β-arrestin1/2, adaptor
protein-2, protein phosphatase 2A and vasodilator-stimulated
phosphoprotein. This enables CXCR2 to mediate different signaling
cascades in breast cancer (21). G
proteins are heterotrimeric protein complexes that are comprised of
three subunits, known as α, β and γ, which are inactive in their
resting state. Upon binding of the ligands to CXCR2, CXCR2
physically couples to the G protein (22). CXCR2 becomes subsequently activated
and the guanosine disphosphate linked to the Gα subunit of the G
protein complex is converted to GTP. This transformation causes Gα
to dissociate from the receptor and Gβγ, leading to the activation
of several downstream signaling pathways.
As presented in Fig.
1, the three main pathways activated by CXCR2 are the
phosphatidylinositol-3 kinase (PI3K)/Akt pathway, the phospholipase
C (PLC)/protein kinase C (PKC) pathway and the
Ras/Raf/extracellular signal related kinases (ERK1/2) pathway
(23–25). The PI3K/Akt pathway is the primary
downstream signaling cascade mediated by PI3K, and protein kinase B
(PKB) plays an essential role in this pathway. PKB is the oncogene
product of the retrovirus Aktδ, which is also known as Akt. Akt
activates IκB kinase (IKK), which phosphorylates IκB to expose the
nuclear localization signals of NF-κB. This allows NF-κB subunits
to translocate to the nucleus (26).
The PI3K/Akt pathway is one of the most commonly altered pathways
in human malignant tumors, and it is critical for cell survival,
motility and angiogenesis (27). The
phosphorylation of CXCR2 also results in the activation of
mitogen-activated protein kinases (MAPK) signaling, which includes
ERK. Ras activates ERK through Raf and MEK, and phosphorylated ERK
translocates from the cytoplasm to the nucleus. This mediates the
transcriptional activation of c-Jun, c-Fos, Elk-1, AP-1 and ATF,
which participate in various biological functions, such as cell
proliferation and differentiation, morphology maintenance,
cytoskeleton construction, apoptosis and tumorigenesis (28,29). The
activation of PLC induces another signaling transduction pathway
that generates two secondary messengers, inositol triphosphate and
diacylglycerol. This leads to calcium mobilization from the
endoplasmic reticulum and activates PKC. Subsequently, c-Jun
N-terminal kinase influences cell apoptosis by mediating the
activity of p53 and Bcl-2 (30,31).
Additionally, the release of the Gα subunit from the G protein
trimer inhibits the conversion of ATP into cyclic AMP by adenylate
cyclase, and this decreases the intracellular levels of cyclic AMP
(21). Furthermore, CXCR2 signaling
could trigger the pathways of focal adhesion kinase (FAK), Rho,
Rac, the Janus kinase/signal transducer and the activator of the
transcription pathways (32–34).
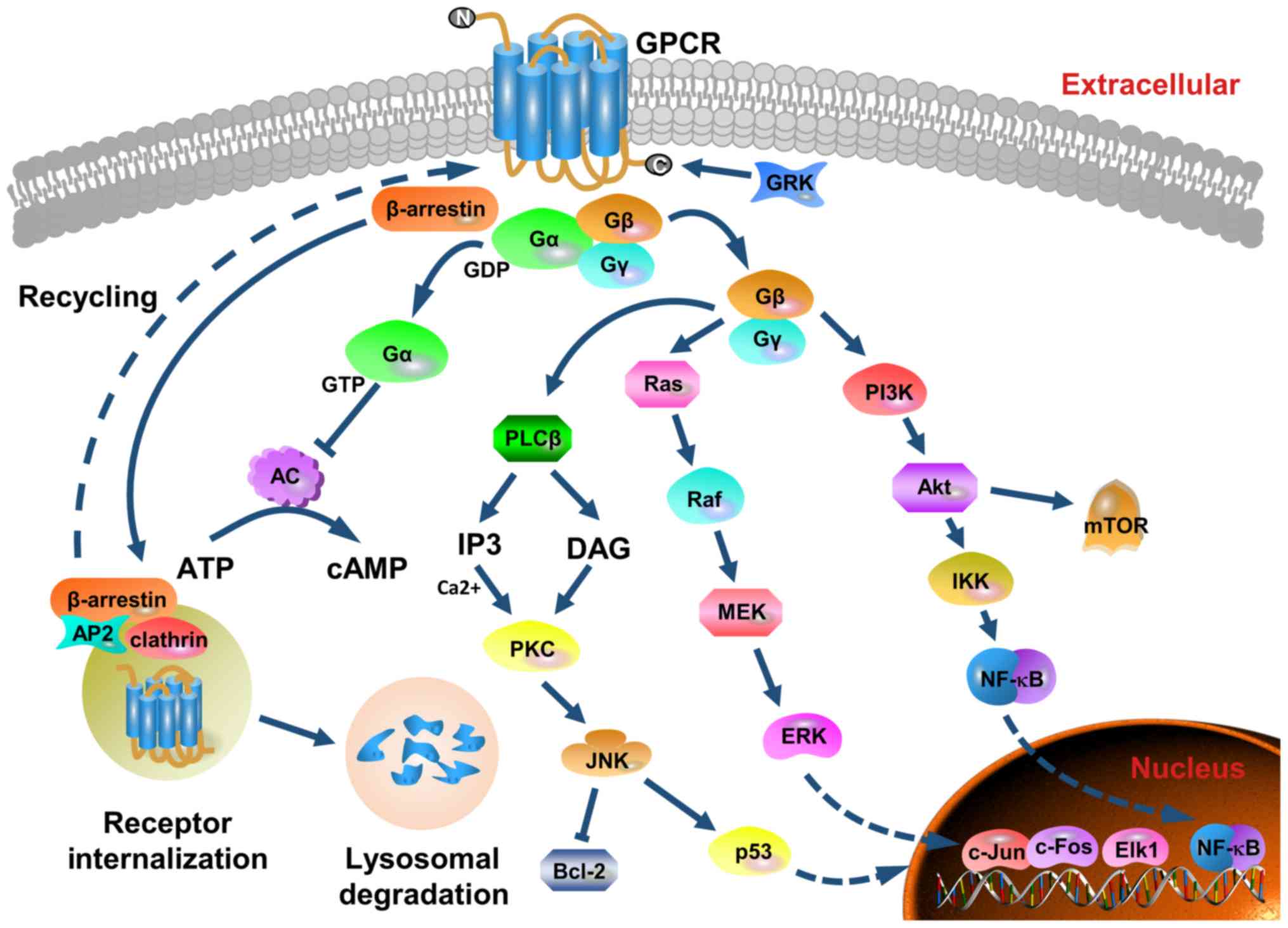 | Figure 1.Structure, major signaling cascades
and receptor recycling of CXCR2. CXCR2 belongs to the G
protein-coupled receptor family that possesses seven transmembrane
structures. It contains an extracellular N-terminus, an
intracellular C-terminus, three extracellular loops and three
cytoplasmic loops. Following ligand binding, CXCR2 physically
couples to the G protein. Subsequently, CXCR2 is activated and the
GDP linked to the Gα subunit of the G protein complex is converted
to GTP. The Gα subunit coupled to the inner cell membrane
dissociates from CXCR2 and the Gβγ subunits. Several downstream
pathways are induced, and the main three signaling pathways are via
PI3K/Akt, PLC/PKC and Ras/Raf/ERK1/2. Moreover, the Gα subunit
inhibits adenylate cyclase activity and decreases the efficiency of
ATP conversion to cAMP. GRK phosphorylates the C-terminus of the
receptor, and mediates the desensitization and endocytosis of the
receptor via β-arrestin recruitment of endocytic components. AP-2
also regulates CXCR2 internalization and sequestration.
Internalized CXCR2 is subjected to degradation by lysosomes, or
recycled to the outer membrane surface. These pathways modulate
cell metabolism, survival, proliferation, apoptosis, angiogenesis,
transduction and motility. As a positive feedback loop to enhance
CXCR2 functionality is formed by upregulating the expression of
cytokines and chemokines. CXCR2, CXC chemokine receptor 2; PI3K,
phosphatidylinositol-3 kinase; PLC, phospholipase C; PKC, protein
kinase C; ERK, extracellular signal related kinase; cAMP, cyclic
AMP; GRK, G-protein coupled receptor kinase; GPCR, G-protein
coupled receptor; GDP, guanosine disphosphate; AC, adenylyl
cyclase. |
These pathways act as modulators in breast cancer
cell metabolism, survival, proliferation, apoptosis, angiogenesis,
transduction and motility. These form a positive feedback loop to
enhance CXCR2 functionality by upregulating the expression of
cytokines and chemokines, such as CXCL8 (19).
Biological characteristics of CXCR2 in
breast cancer
CXCR2 and breast cancer growth
Tumorigenesis is a process that involves multiple
genes, several steps and various signaling pathways, including the
complex regulatory network of chemokines and their receptors
(35,36). Studies have found that the
CXCL8-CXCR2 axis is closely associated with multiple stages of
breast cancer growth. These include the regulation of transcription
of CDK inhibitors p21Cip1, p27Kip1 and p57Kip2, and the induction
of tumor cell proliferation, differentiation, stress response and
apoptosis (15,20,37).
Paradoxically, CXCL8 enhances the immune system response and
increases its ability to execute anti-tumor effects, whereas it
also transforms the tumor microenvironment to promote tumor growth.
Compared with healthy volunteers, patients with breast cancer have
elevated levels of CXCL8, and the severity of this overexpression
is positively correlated with disease stage (38). The levels of CXCR2 in malignant
tissues are higher than those in benign and normal tissues
(39). Polymorphisms in the CXCL8
and CXCR2 genes also suggest that elevated CXCL8 and CXCR2
expression may be risk factors of breast cancer (38).
However, the biological significance of CXCR2 in
cancer cell proliferation remains controversial. Shao et al
(40) performed small interfering
RNA-mediated knockdown of endogenous CXCL8 that upregulated
p27Kip21 and downregulated cyclin D1. The decreased Akt
phosphorylation and NF-κB activation resulted in reduced cell
proliferation in both MDA-MB-231 and BT549 breast cancer cell
lines. This indicated that CXCL8 and CCL2 overexpression enhances
tumor proliferation (40). By
contrast, other studies have shown that the overexpression of CXCR2
induces premature senescence, and silencing of CXCR2 prolongs cell
passage via p53, NF-κB or C/EBPβ-associated pathways (39,41).
Overall, several studies have reported that CXCR2 is a
tumor-stimulating receptor that could be exploited as a marker of
poor prognosis in a variety of cancer types. Thus, inhibiting CXCR2
production may promote cancer cell apoptosis (42,43).
Therefore, CXCR2 may have different functions in normal,
precancerous and tumor cells and requires further
investigation.
In the tumor microenvironment, breast cancer growth
in both autocrine and paracrine manners are regulated by CXCR2 and
its ligands produced by stromal cells (44). Furthermore, neutrophils, myeloid
cells and bone marrow-derived suppressor cells express CXCR2 and
assist in tumor cell proliferation (44). Following the entry of neutrophils
into the tumor site, an increase in cytokine secretion contributes
to the production of an inflammatory microenvironment (45). Additionally, bone marrow-derived
suppressor cells differentiate into M2-type macrophages, which
facilitate cancer cell growth (46).
Previous studies have demonstrated the knockout of the CXCR2 gene
in host cells to inhibit tumor growth and increased tumor cell
apoptosis (47–49).
CXCR2 and breast cancer
angiogenesis
Once tumors exceed 1–2 mm in diameter, angiogenesis
is initiated for growth and metastasis (50,51).
CXCR2 affects angiogenesis in breast cancer primarily by
interacting with CXCL8 and CXCL1, however the specific mechanism is
yet to be determined (52–54).
Addison et al (53) detected the expression of CXCR2 using
a CXCR2 antibody in human microvascular endothelial cells and
confirmed that the chemotaxis of ELR+CXC
chemokine-mediated microvascular endothelial cell was obstructed,
and was sensitive to pertussis toxins (53). Studies in CXCR2-deficient mice
indicated that CXCL8 is the strongest ligand for CXCR2, and is
mediated by the activation of the ELR+CXC chemokine
(52). In cancer cells, CXCL8 and
vascular endothelial growth factor (VEGF) cooperate to establish
and expand tumor neovascularization. Furthermore, glucose
deprivation and endoplasmic reticulum stress effectively induce the
upregulation of CXCL8 (55). CXCL8
and VEGF are regulated by distinct pathways in different cell
lines. MDA-MB-231 cells mainly activates the MAPK-ERK pathway, and
the activity of the PI3K/Akt pathway is increased in GI101A cells.
Both signaling pathways are activated in MDA-MB-468 and Hs578T cell
lines (56). CXCL8 generated by
endothelial cells binds to CXCR2 to mediate interactions between
CXCR2 and VEGFR receptor 2 (VEGFR2). This includes the
transactivation of VEGFR2 via Src kinase-mediated receptor
phosphorylation, which is required for CXCL8 to induce endothelial
cell permeability (56). The
CXCL8-CXCR2 axis also induces VEGF transcription and stimulates
VEGFR2 activation through the NF-κB pathway in endothelial cells
(57). Moreover, the CXCL8-CXCR2
axis activates the expression of EGFR to mediate endothelial cell
migration and capillary formation (58). It also elevates integrin αvβ3 levels,
which serve a key role in endothelial cell survival and cancer cell
migration during tumor angiogenesis (59). Another study revealed that the
expression of CXCL8 in ER+ cells was lower than that in
ER− cells, and exogenous ERα substantially interfered
with CXCL8 expression. This suggests that the inactivation of ERα
and upregulation of CXCL8 could promote angiogenesis in human
breast cancer (60). The silencing
of CXCR2 further indicated the importance of CXCL8-mediated
angiogenesis. Nannuru et al (61) analyzed the microvessel density of
primary tumor sections, and found that silencing CXCR2 in Cl66
cells considerably decreased tumor angiogenesis compared with the
control group.
Furthermore, thrombin stimulates tumors to secrete
CXCL1 in endothelial cells, which reinforces tumor angiogenesis.
Thus, thrombin-induced angiogenesis could be perturbed by the CXCL1
antibody (54). In 4T1 cells,
shRNA-knockdown of CXCL1 impeded tumor growth and angiogenesis
(54).
CXCR2 and breast cancer
metastasis
Metastasis is a basic biological characteristic of
malignant neoplasia. Distant metastasis confers breast cancer a
worse prognosis, with the five-year survival rate of 27% in the
United States between 2008 and 2014, whereas the five-year survival
rate of the localized stage was of 99% (62). Metastasis occurs predominantly
through the lymphatic system, blood, direct infiltration and
planting. This process is extremely complex, dynamic and
continuous, and contains several independent processes. For
example, when tumors metastasize via the blood circulation, the
cancer cells proliferate within the primary lesion and form new
blood vessels. Subsequently, the aggressive cells detach from the
primary tumor. This is followed by epithelial-mesenchymal
transition (EMT), which results in cells with properties similar to
interstitial cells. Epithelial cells subjected to EMT, lose their
intercellular connections and polarity, and experience changes in
cell morphology and increased migration capacity. Tumor cells grow
into the surrounding interstitial space by infiltration, and are in
close contact with local capillaries and lymphatic endothelial
cells. Once cells have invaded this interstitial space, they
subsequently penetrate the walls of the vessel and protrude into
the lumen. This enables transport to the target tissue, where the
cells then proliferate in the matrix to form a new secondary tumor
(63,64).
CXCR2 is involved in the migration, invasion and
metastasis of breast cancer cells in various ways (61). A number of studies have investigated
the effects of CXCL8 on cancer invasion and migration (65–68).
High levels of CXCL8 expression promote angiogenesis and attract
neutrophils to release enzymes involved in tissue remodeling and
tumor formation. Thus, ectopic expression of CXCL8, stimulated by
IL-1β and TNF-α, could exacerbate the metastatic potential of
breast cancer (65). A CpG island
located upstream of the CXCL8 promoter in the highly metastatic
cell lines MDA-231 and MDA-345, results in the upregulation of
CXCL8 expression (66). However, the
loss of CXCL8 production in MDA-MB-231 and BT549 cells consequently
attenuates migration and invasion, which may be due to decreased
integrin β3 transcription (40).
COX-2-mediated CXCL8 production in ER-negative breast cancer cells
promotes osteoclast formation and bone metastasis (67). Compared with patients with no
metastasis, those with breast cancer and bone metastases
demonstrate increased CXCL8 levels. Furthermore, there is a
significant positive correlation between plasma CXCL8 levels and
bone resorption (68). In addition,
CXCL8 regulates the actin cytoskeleton through
Ca2+-activated PLC-dependent PKC and Rho-GTPase
(69). Additionally, CXCL8 also
promotes cell migration through the stimulation of the
intracellular Akt pathway (27).
Triple-negative breast cancer (TNBC) cells secrete CXCL8, which
activates CXCR2 in tumor-associated fibroblasts and
tumor-associated macrophages. This results in STAT3 phosphorylation
and upregulation of CXCL8 transcription and translation. The
binding of CXCL8 to CXCR2 in cancer cells facilitates metastasis
(70), which was also observed in
MDA-MB-231 cells and xenograft mouse models with CXCR2-knockout, as
well as decreased migration, compared with wild-type cells
(70). These data indicate that the
CXCR2-CXCL8 axis is multifaceted in tumor progression and
metastasis, and renders cancer cells invasive.
Bone marrow-derived mesenchymal stem cells (MSCs)
express CXCL1 and CXCL5 to recruit PyMT breast cancer cells and
prompt the migration in a CXCR2-dependent manner in vitro.
The CXCR2-inhibitory antibody, SB265610, substantially curbs the
migration of cancer cells to MSC-conditioned media (71). Additionally, it was proposed that
TNFα-activated MSCs secrete CXCR2 ligands to recruit
CXCR2+ neutrophils to tumor sites (72). Tumor-associated neutrophils stimulate
metastasis through matrix metalloproteinases and other soluble
molecules. A co-culture system, consisting of tumor cells and
neutrophils, substantiated to higher expression levels of
metastasis-associated genes in tumor cells, which identified an
MSCs/neutrophil/tumor cell axis associated with cancer metastasis
(72).
The silencing of CXCR2 in breast cancer cell lines
using short hairpin RNA results in attenuated cell invasion.
Moreover, when these shRNA-treated cells were transplanted into an
orthotopic mouse xenograft model, spontaneous lung metastases were
decreased by 40% compared with the control group (61). In addition, the loss of CXCR2
expression in stroma cells (neutrophils, macrophages and
endothelial cells) in the tumor microenvironment also prevented
cancer cell migration (47). It was
hypothesized that CXCR2 is involved in the paracrine loop between
tumor cells and its surroundings, which potentiates invasion and
metastasis.
CXCR2 and drug resistance in breast
cancer
Drug resistance is one of the challenges of advanced
breast cancer therapy. Tumors gradually become less responsive to
chemoradiotherapy during treatment and therefore, the mortality
rate of breast cancer remains high. As a consequence of drug
resistance, there are few recognized therapeutic strategies
(73,74).
A wealth of evidence has shown that malignant cells
that survive primary chemoradiotherapy express higher levels of
CXCR2 ligands (65,75). Additionally, high levels of CXCL1,
CXCL3, CXCL5, CXCL6, CXCL7 and CXCL8 are observed in drug-resistant
breast cancer cells, which diminishes the effectiveness of other
medical interventions (7,76). Shi et al (76) revealed that the multidrug-resistant
human breast cancer cell line, MCF-7/R, has higher expression
levels of CXCL6 and CXCL8 compared with the sensitive control cell
line MCF-7/S. The inhibition of CXCL6 and CXCL8 with antibodies
enhances the sensitivity to paclitaxel and doxorubicin treatment in
MCF-7/R cells. The inhibition of CXCL6 and CXCL8 expression
reverses the chemoresistance of MCF-7/R cells, and the
overexpression of CXCL6 and CXCL8 potentiates the resistance of
MCF-7/S cells to doxorubicin (76).
Similarly, other studies demonstrated that the inhibition of CXCL8
in MDA-MB-231 and BT549 cells improved the efficacy of chemotherapy
(40). The inhibition of IL-8 or
CXCR2 was shown to prevent the paclitaxel-induced autocrine
inflammatory feed-forward loop (77). Sharma et al (75) used Cl66-wt, 4T1-wt, Cl66sh-CXCR2 and
4T1sh-CXCR2 cells that expressed varying levels of CXCR2, to assess
the effect of CXCR2 levels on chemosensitivity. Furthermore, it was
reported that the silencing of CXCR2 could increase the
cytotoxicity of paclitaxel and doxorubicin, and intensify the
antitumor activity of these drugs (75).
Regarding the intracellular mechanism of
CXCR2-mediated drug resistance, previous studies suggested that AKT
regulates chemical resistance through a broad anti-apoptotic
molecule, such as PED (78–80). Patients with breast cancer and Akt
phosphorylation at serine 473 are more sensitive to paclitaxel
treatment (81). The overexpression
of COX2 results in chemical resistance to breast cancer by
generating prostaglandin H2 and activating NF-κB (82), which was also observed in a study by
Xu et al (39). These results
suggest that CXCR2-dependent regulatory pathways are crucial for
cancer cells to maintain chemoresistance, and provide evidence that
supports the targeting of the CXCR2 axis as an adjunctive therapy
to prevent drug resistance.
CXCR2 and breast cancer stem
cells
Cancer stem cells (CSCs) were first identified in
breast cancer (83), and the
presence of CSCs may be responsible for the high level of
heterogeneity observed in this disease (84). Breast cancer stem cells (BCSCs) are
resistant to the effects of chemoradiotherapy and endocrine therapy
due to their self-renewal and differentiation potential. This
allows BCSBCs to initiate and maintain tumor growth, invasion,
metastasis, drug resistance and recurrence (85). The importance of CSCs was
demonstrated in several cancer types (86). Trastuzumab prolongs survival in
patients with HER2-positive breast cancer treatment, which is in
part due to its ability to limit CSCs (87). Neoadjuvant lapatinib was shown to
suppress BCSCs in HER2-positive tumors and its combination with
lapatinib prolongs the time to progression in patients with
trastuzumab resistance (88).
CXCL8 transmits signals through CXCR2, which induces
EMT (89). EMT is a process that
regulates invasion and metastasis, and enables cells to obtain stem
cell characteristics (36). Singh
et al (90) demonstrated a
positive correlation between the level of CXCL8 in metastatic
pleural and peritoneal effusion and the ability of these tumor
cells to form mammospheres in vitro. The inhibition of CXCR2
using a small molecule CXCR1/2 antagonist, SCH563705, abolished the
impact of CXCL8 expression. This was quantified by enzyme-linked
immunosorbent assay (90). The
suppression of CXCR2 expression limits tumor spheroid formation and
aldefluor-positive rate in breast cancer cells, and increases the
efficacy of anti-HER2 therapy in HER2-positive patients (91). Furthermore, CXCR2 regulates the
activity of CSCs through both HER2-dependent and HER2-independent
pathways. One such pathway is through FAK, which is also associated
with BCSC maintenance (92). In
vivo studies have shown that MSCs are recruited to the tumor
site, co-localized with CSCs in the microenvironment and secreted
cytokines. This increases the number of CSCs present in the tumor,
thereby accelerating tumor growth (93). Liu et al (93) suggested that this effect is initiated
by CXCL6 secreted by cancer cells, and is maintained by CXCL8 and
other CXCR2 ligands (GCP-2 and NAP-2) that are secreted by cancer
cells and stromal cells. In addition, cancer cell-derived CXCL1
induces the expression of CXCL8 and GRO-α in MSCs, which promotes
the formation and maintenance of CSCs (94). The inhibition of CXCR2 or downstream
pathways can decrease the number and activity of CSCs in
vitro and in xenografts, and increase the efficacy of docetaxel
to decrease tumor volume (95).
Recently, studies have identified CXCR2 as a novel BCSC biomarker
of only TNBC. CXCR2 was observed to be co-expressed with
CSC-associated proteins, such as NANOG and SOX2 (96,97).
Furthermore, 4T1 cells that express CXCR2 have characteristics of
CSCs, including a low proportion of CXCR2-positive cells (~1%),
hypoxia, elevated expression of CSC-associated mRNA, increased
tumor spherule formation, tumor xenografts, resistance to
radiotherapy and chemotherapy (96).
In addition to CXCR2, CXCR1 was also found to be
present on the surface of BCSCs, which facilitates the growth of
BCSCs stimulated by tissue damage or inflammation (98). CXCR1 is also known as IL-8 receptor
A, and is an active receptor selectively expressed by BCSCs
(99). CXCR1 belongs to the same
chemokine CXC receptor family as CXCR2, and CXCR1 has 75% sequence
similarity with CXCR2 (100). It
was proposed that CXCR1 has a prominent role in both the initiation
and therapy of breast cancer (101). The inhibition of CXCR1 results in
the depletion of BCSCs in vitro (102). CXCR1 inhibition combined with
chemotherapy, results in the release of CXCL8 by dying bulk
(non-CSC) tumor cells and binds with CXCR1 on the surface of BCSCs.
As a result, the BCSCs are protected from apoptotic signals
triggered by the Fas ligand (102).
Several CXCR2 inhibitors, such as Reparixin, also block CXCR1. This
further decreases the enrichment of BCSCs and prevents tumor
recurrence (77,79).
The aforementioned results uncover the integral role
played by CXCR2 signaling in the complex inflammatory cytokine
response. Furthermore, the role of CXCR1 in maintaining the
activity of BCSCs via autocrine or paracrine pathways, and its
effects on treatment efficacy and disease prognosis have been
uncovered (87).
Preclinical and clinical evidence of
targeting CXCR2 in breast cancer
Targeted therapy offers a unique opportunity to
suppress the activity of key genes associated with tumorigenesis
(103). CXCR2 and its ligands are
influential targets involved in tumor growth and regulation
(Fig. 2) (19). Additionally, preclinical studies on
other malignant neoplasms have revealed that targeting CXCR2 alone,
or in combination with other regimens, may provide an effective,
novel option for cancer management (104–107).
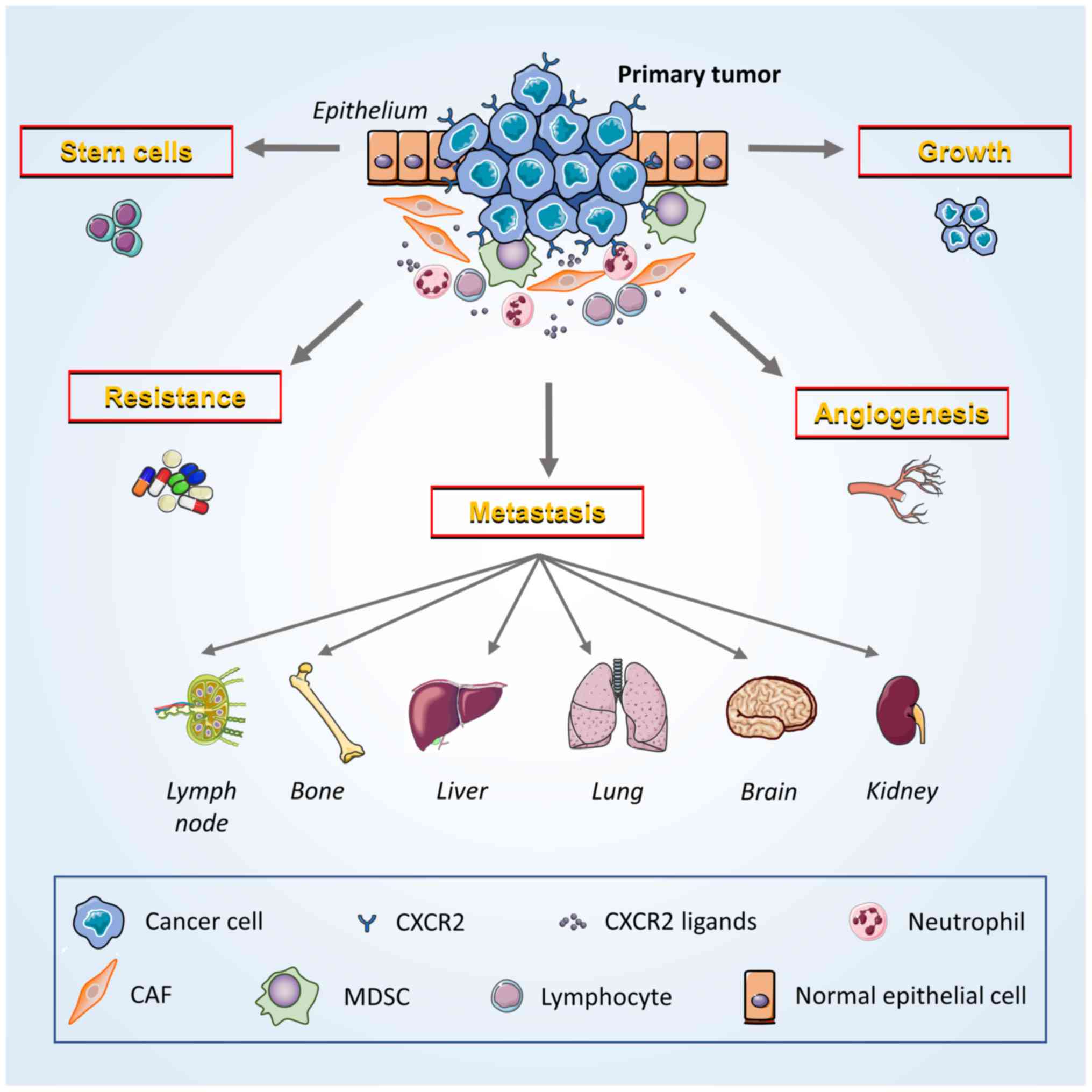 | Figure 2.Potential contribution of CXCR2 to
breast cancer development and progression. Breast cancer cells and
multiple host cells (MDSCs, neutrophils and lymphocytes) in the
tumor microenvironment exert various cancer-promoting functions
that are induced by the interaction of CXCR2 and its ligands. Tumor
cells express both CXCR2 and specific ligands to promote the growth
of neoplasms via autocrine and paracrine signaling. CXCR2 also
facilitate crosstalk with vascular endothelial cells, induce
angiogenesis and enhance the ability of cancer cells to invade and
migrate. CXCR2 promotes the metastasis of cancer cells to other
locations throughout the body, including lymph nodes, bones, liver,
lungs, brain and kidneys. Moreover, CXCR2 decreases the sensitivity
of cancer cells to chemoradiotherapy by enabling them to survive
treatment. Furthermore, the maintenance of cancer stem cell
activity may depend on the ability of CXCR2 to induce the stemness
of breast cancer cells. CXCR2, CXC chemokine receptor 2; MDSCs,
myeloid-derived suppressor cells; CAFs, cancer-associated
fibroblasts. |
Although CXCL8 is the most commonly studied ligand
of CXCR2 in breast cancer, other CXCR2 receptor agonists, such as
GRO-α, GRO-β, GRO-γ and CXCL5, are co-regulated with CXCL8. This
co-regulation of CXCR2 ligands may limit the effectiveness of
targeting CXCL8 alone (108), which
could be circumvented by suppressing CXCR2 and its downstream
signaling pathways. Blocking the function of CXCR2, by impeding
ligand binding or subsequent pathway activation, essentially
prevents the biological effects of multiple ELR+ CXC
chemokines with a single treatment. This intervention may be more
tumor-specific than the direct inhibition of cytokines, and could
decrease the severity of adverse events.
There are various orally active small-molecules that
are non-competitive antagonists of CXCR2, including Repertaxin,
SCH479833, SCH527123 and SCH563705. These antagonists exhibit
anti-tumor properties in breast cancer (95), colorectal cancer (105) and melanoma (106) xenograft models. These properties
include the obstruction of spontaneous hepatic metastasis of colon
carcinomas (109). Moreover, the
combination of paclitaxel treatment and the inhibition of CXCR2
slows breast tumor growth, attenuates tumor angiogenesis and lung
metastasis, and diminishes the number of CSCs. Additionally,
targeting CXCR2 reduces drug resistance (75,110).
Since CXCL8 is a prominent regulator of BCSC activity, a combined
treatment of CXCR2 inhibitors and current HER2-targeted therapies
is predicted to be an effective regimen to decrease CSC activity
and increase survival in HER2-positive patients with breast cancer
(90). A recent study in pancreatic
ductal adenocarcinomas demonstrated that inhibition of CXCR2
predominantly in neutrophils/MDSCs confers T cell entry to the
tumor site (111). The inhibition
of CXCR2 also enhances the sensitivity to anti-programmed death 1
therapy, which reinforces the endogenous anti-tumor immune response
(111). Moreover, Uddin et
al (98) revealed that the
inhibition of proteasome significantly upregulates the levels of
CXCL8 and its receptors CXCR1/2, through IKK. The inhibition of the
induced CXCL8 expression or IKK activity may improve the
effectiveness of proteasome inhibitors as a therapy for TNBC.
Overall, CXCR2 and its ligands are potential therapeutic
targets.
Due to the various functions of CXCL8 in homeostatic
processes, the side effects of CXCR2 inhibitors cannot be ignored.
Neutrophils are part of the immune surveillance system that
monitors and destroys transformed cells, and CXCL8 exerts an
anti-tumor effect by recruiting concentrated granulocytes (112). Therefore, the anti-tumor effects of
CXCR2 inhibitors that influence neutrophil infiltration may
unintentionally promote tumor growth. The solution to this dilemma
depends on the new technologies of targeted drug therapies for
cancer cells. In addition, CXCR2 inhibitors cause a marked decrease
in the number of circulating neutrophils combined with
chemotherapeutic agents, and this could cause a potentially
synergistic increase in myelotoxicity (113).
Currently, CXCR2 antagonists are used in phase II
clinical trials for chronic obstructive pulmonary disease (114), and obvious adverse reactions were
reported in patients with severe asthma (113). Repertaxin was originally used to
prevent CXCL8-induced reperfusion injury, and was the only
clinically-tested CXCR2 inhibitor (21). Interfering with CXCR2 is particularly
advantageous in breast cancer compared with other tumors. Phase I
clinical trials have shown that Repertaxin, in combination with
paclitaxel, is safe and well tolerated (99). Phase II studies on this combination
are currently ongoing (99). The
principle data on the efficacy and safety lay an essential
foundation for the accelerated development of CXCR2 inhibitors as a
treatment for breast cancer.
Conclusions and outlooks
Currently, controversies still exist in the field of
CXCR2 research. One controversy is the association between CXCR2
and the expression of ER. Other controversies include the
characterization of CXCR2 in cancer cell senescence, and its role
in Akt-associated tumor metastasis (39,115–117).
These controversies may be attributed to the limitations of the
pathological tissue, cell type and animal model used.
The association between inflammation and tumor
development is widely recognized, and the expression of many
inflammatory cytokines in the tumor microenvironment has previously
been reported (19). The poor
efficacy of current breast cancer treatments may be associated with
the molecular typing, and the function of CXCR2 in breast cancer.
The anti-tumor effect of targeting CXCR2 was reflected in both
in vitro and in vivo models of breast cancer, and
further supports the development of CXCR2 inhibitors as a clinical
therapy in breast cancer. Further exploration of whether CXCR2
inhibition plays a vital role in targeted therapies for breast
cancer, based on molecular typing, is warranted.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
FG, LL and YW were significant contributors to the
conception and design of this review article. LL, JW and YL were
involved in the literature search. JW, YL and FG analyzed and
summarized critically the existing knowledge obtained from the
literature. FG and LL drafted the manuscript text. LW, FL, LL and
FG designed and generated the figures. YW, LW and FL made
substantial contributions to the manuscript revision regarding
important intellectual content. All authors have read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Sun K, Zheng R, Zeng H, Zhang S,
Xia C, Yang Z, Li H, Zou X and He J: Cancer incidence and mortality
in China, 2014. Chin J Cancer Res. 30:1–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harbeck N, Thomssen C and Gnant M: St.
Gallen 2013: Brief preliminary summary of the consensus discussion.
Breast Care (Basel). 8:102–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Edenfield J, Schammel C, Collins J,
Schammel D and Edenfield WJ: Metaplastic breast cancer: Molecular
typing and identification of potential targeted therapies at a
single institution. Clin Breast Cancer. 17:e1–e10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma F, Guan Y, Yi Z, Chang L, Li Q, Chen S,
Zhu W, Guan X, Li C, Qian H, et al: Assessing tumor heterogeneity
using ctDNA to predict and monitor therapeutic response in
metastatic breast cancer. Int J Cancer. Jun 26–2019.(Epub ahead of
print). View Article : Google Scholar
|
|
6
|
Lee KL, Kuo YC, Ho YS and Huang YH:
Triple-negative breast cancer: Current understanding and future
therapeutic breakthrough targeting cancer stemness. Cancers.
11(pii): E13342019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sharma B, Varney ML, Saxena S, Wu L and
Singh RK: Induction of CXCR2 ligands, stem cell-like phenotype and
metastasis in chemotherapy-resistant breast cancer cells. Cancer
Lett. 372:192–200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang Y, Luo B, An Y, Sun H, Cai H and Sun
D: Systematic review and meta-analysis of the prognostic value of
CXCR2 in solid tumor patients. Oncotarget. 8:109740–109751. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murdoch C and Finn A: Chemokine receptors
and their role in inflammation and infectious diseases. Blood.
95:3032–3043. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang Y, Feng Y, Wu W, Chang C, Chen D,
Chen S and Zhen G: MicroRNA-218-5p plays a protective role in
eosinophilic airway inflammation via targeting δ-catenin, a novel
catenin in asthma. Clin Exp Allergy. Sep 12–2019.(Epub ahead of
print). View Article : Google Scholar
|
|
11
|
Wang X, Iyer A, Lyons AB, Körner H and Wei
W: Emerging roles for G-protein coupled receptors in development
and activation of macrophages. Front Immunol. 10:20312019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rollins BJ: Chemokines. Blood. 90:909–928.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Debnath B, Xu S, Grande F, Garofalo A and
Neamati N: Small molecule inhibitors of CXCR4. Theranostics.
3:47–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
O'Hayer KM, Brady DC and Counter CM: ELR+
CXC chemokines and oncogenic Ras-mediated tumorigenesis.
Carcinogenesis. 30:1841–1847. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lloyd A, Modi W, Sprenger H, Cevario S,
Oppenheim J and Kelvin D: Assignment of genes for interleukin-8
receptors (IL8R) A and B to human chromosome band 2q35. Cytogenet
Cell Genet. 63:238–240. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kobilka BK: G protein coupled receptor
structure and activation. Biochim Biophys Acta. 1768:794–807. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prado GN, Suetomi K, Shumate D, Maxwell C,
Ravindran A, Rajarathnam K and Navarro J: Chemokine signaling
specificity: Essential role for the N-terminal domain of chemokine
receptors. Biochemistry. 46:8961–8968. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moussouras NA, Getschman AE, Lackner ER,
Veldkamp CT, Dwinell MB and Volkman BF: Differences in
sulfotyrosine binding amongst CXCR1 and CXCR2 chemokine ligands.
Int J Mol Sci. 18(pii): E18942017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng Y, Ma XL, Wei YQ and Wei XW:
Potential roles and targeted therapy of the CXCLs/CXCR2 axis in
cancer and inflammatory diseases. Biochim Biophys Acta Rev Cancer.
1871:289–312. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ahuja SK and Murphy PM: The CXC chemokines
growth-regulated oncogene (GRO) alpha, GRObeta, GROgamma,
neutrophil-activating peptide-2, and epithelial cell-derived
neutrophil-activating peptide-78 are potent agonists for the type
B, but not the type A, human interleukin-8 receptor. J Biol Chem.
271:20545–20550. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ha H, Debnath B and Neamati N: Role of the
CXCL8-CXCR1/2 axis in cancer and inflammatory diseases.
Theranostics. 7:1543–1588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Damaj BB, McColl SR, Mahana W, Crouch MF
and Naccache PH: Physical association of Gi2alpha with
interleukin-8 receptors. J Biol Chem. 271:12783–12789. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu D, LaRosa GJ and Simon MI: G
protein-coupled signal transduction pathways for interleukin-8.
Science. 261:101–103. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Knall C, Young S, Nick JA, Buhl AM,
Worthen GS and Johnson GL: Interleukin-8 regulation of the
Ras/Raf/mitogen-activated protein kinase pathway in human
neutrophils. J Biol Chem. 271:2832–2838. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Knall C, Worthen GS and Johnson GL:
Interleukin 8-stimulated phosphatidylinositol-3-kinase activity
regulates the migration of human neutrophils independent of
extracellular signal-regulated kinase and p38 mitogen-activated
protein kinases. Proc Natl Acad Sci USA. 94:3052–3057. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oeckinghaus A and Ghosh S: The NF-kappaB
family of transcription factors and its regulation. Cold Spring
Harb Perspect Biol. 1:a0000342009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cheng GZ, Park S, Shu S, He L, Kong W,
Zhang W, Yuan Z, Wang LH and Cheng JQ: Advances of AKT pathway in
human oncogenesis and as a target for anti-cancer drug discovery.
Curr Cancer Drug Targets. 8:2–6. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hoffmann E, Dittrich-Breiholz O, Holtmann
H and Kracht M: Multiple control of interleukin-8 gene expression.
J Leukoc Biol. 72:847–855. 2002.PubMed/NCBI
|
|
29
|
Tang H, Sun Y, Shi Z, Huang H, Fang Z,
Chen J, Xiu Q and Li B: YKL-40 induces IL-8 expression from
bronchial epithelium via MAPK (JNK and ERK) and NF-κB pathways,
causing bronchial smooth muscle proliferation and migration. J
Immunol. 190:438–446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Clapham DE: Calcium signaling. Cell.
80:259–268. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lang K, Niggemann B, Zanker KS and
Entschladen F: Signal processing in migrating T24 human bladder
carcinoma cells: Role of the autocrine interleukin-8 loop. Int J
Cancer. 99:673–680. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schraufstatter IU, Chung J and Burger M:
IL-8 activates endothelial cell CXCR1 and CXCR2 through Rho and Rac
signaling pathways. Am J Physiol Lung Cell Mol Physiol.
280:L1094–L1103. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cohen-Hillel E, Yron I, Meshel T, Soria G,
Attal H and Ben-Baruch A: CXCL8-induced FAK phosphorylation via
CXCR1 and CXCR2: Cytoskeleton- and integrin-related mechanisms
converge with FAK regulatory pathways in a receptor-specific
manner. Cytokine. 33:1–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Britschgi A, Andraos R, Brinkhaus H,
Klebba I, Romanet V, Müller U, Murakami M, Radimerski T and
Bentires-Alj M: JAK2/STAT5 inhibition circumvents resistance to
PI3K/mTOR blockade: A rationale for cotargeting these pathways in
metastatic breast cancer. Cancer Cell. 22:796–811. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sotiriou C, Neo SY, McShane LM, Korn EL,
Long PM, Jazaeri A, Martiat P, Fox SB, Harris AL and Liu ET: Breast
cancer classification and prognosis based on gene expression
profiles from a population-based study. Proc Natl Acad Sci USA.
100:10393–10398. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hebert CA, Vitangcol RV and Baker JB:
Scanning mutagenesis of interleukin-8 identifies a cluster of
residues required for receptor binding. J Biol Chem.
266:18989–18994. 1991.PubMed/NCBI
|
|
38
|
Snoussi K, Mahfoudh W, Bouaouina N, Fekih
M, Khairi H, Helal AN and Chouchane L: Combined effects of IL-8 and
CXCR2 gene polymorphisms on breast cancer susceptibility and
aggressiveness. BMC Cancer. 10:2832010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu H, Lin F, Wang Z, Yang L, Meng J, Ou Z,
Shao Z, Di G and Yang G: CXCR2 promotes breast cancer metastasis
and chemoresistance via suppression of AKT1 and activation of COX2.
Cancer Lett. 412:69–80. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shao N, Chen LH, Ye RY, Lin Y and Wang SM:
The depletion of interleukin-8 causes cell cycle arrest and
increases the efficacy of docetaxel in breast cancer cells. Biochem
Biophys Res Commun. 431:535–541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ruan JW, Liao YC, Lua I, Li MH, Hsu CY and
Chen JH: Human pituitary tumor-transforming gene 1 overexpression
reinforces oncogene-induced senescence through CXCR2/p21 signaling
in breast cancer cells. Breast Cancer Res. 14:R1062012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee YS, Choi I, Ning Y, Kim NY,
Khatchadourian V, Yang D, Chung HK, Choi D, LaBonte MJ, Ladner RD,
et al: Interleukin-8 and its receptor CXCR2 in the tumour
microenvironment promote colon cancer growth, progression and
metastasis. Br J Cancer. 106:1833–1841. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Desurmont T, Skrypek N, Duhamel A,
Jonckheere N, Millet G, Leteurtre E, Gosset P, Duchene B, Ramdane
N, Hebbar M, et al: Overexpression of chemokine receptor CXCR2 and
ligand CXCL7 in liver metastases from colon cancer is correlated to
shorter disease-free and overall survival. Cancer Sci. 106:262–269.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Katoh H, Wang D, Daikoku T, Sun H, Dey SK
and Dubois RN: CXCR2-expressing myeloid-derived suppressor cells
are essential to promote colitis-associated tumorigenesis. Cancer
Cell. 24:631–644. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou SL, Dai Z, Zhou ZJ, Wang XY, Yang GH,
Wang Z, Huang XW, Fan J and Zhou J: Overexpression of CXCL5
mediates neutrophil infiltration and indicates poor prognosis for
hepatocellular carcinoma. Hepatology. 56:2242–2254. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sharma B, Nannuru KC, Varney ML and Singh
RK: Host Cxcr2-dependent regulation of mammary tumor growth and
metastasis. Clin Exp Metastasis. 32:65–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Singh S, Varney M and Singh RK: Host
CXCR2-dependent regulation of melanoma growth, angiogenesis and
experimental lung metastasis. Cancer Res. 69:411–415. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cardona AE, Sasse ME, Liu L, Cardona SM,
Mizutani M, Savarin C, Hu T and Ransohoff RM: Scavenging roles of
chemokine receptors: Chemokine receptor deficiency is associated
with increased levels of ligand in circulation and tissues. Blood.
112:256–263. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Folkman J: Angiogenesis. Annu Rev Med.
57:1–18. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Stadtmann A and Zarbock A: CXCR2: From
bench to bedside. Front Immunol. 3:2632012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Addison CL, Daniel TO, Burdick MD, Liu H,
Ehlert JE, Xue YY, Buechi L, Walz A, Richmond A and Strieter RM:
The CXC chemokine receptor 2, CXCR2, is the putative receptor for
ELR+ CXC chemokine-induced angiogenic activity. J Immunol.
165:5269–5277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Caunt M, Hu L, Tang T, Brooks PC, Ibrahim
S and Karpatkin S: Growth-regulated oncogene is pivotal in
thrombin-induced angiogenesis. Cancer Res. 66:4125–4132. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Marjon PL, Bobrovnikova-Marjon EV and
Abcouwer SF: Expression of the pro-angiogenic factors vascular
endothelial growth factor and interleukin-8/CXCL8 by human breast
carcinomas is responsive to nutrient deprivation and endoplasmic
reticulum stress. Mol Cancer. 3:42004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Petreaca ML, Yao M, Liu Y, Defea K and
Martins-Green M: Transactivation of vascular endothelial growth
factor receptor-2 by interleukin-8 (IL-8/CXCL8) is required for
IL-8/CXCL8-induced endothelial permeability. Mol Biol Cell.
18:5014–5023. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Martin D, Galisteo R and Gutkind JS:
CXCL8/IL8 stimulates vascular endothelial growth factor (VEGF)
expression and the autocrine activation of VEGFR2 in endothelial
cells by activating NFkappaB through the CBM (Carma3/Bcl10/Malt1)
complex. J Biol Chem. 284:6038–6042. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kyriakakis E, Cavallari M, Pfaff D, Fabbro
D, Mestan J, Philippova M, De Libero G, Erne P and Resink TJ:
IL-8-mediated angiogenic responses of endothelial cells to lipid
antigen activation of iNKT cells depend on EGFR transactivation. J
Leukoc Biol. 90:929–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Niu G and Chen X: Why integrin as a
primary target for imaging and therapy. Theranostics. 1:30–47.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lin Y, Huang R, Chen L, Li S, Shi Q,
Jordan C and Huang RP: Identification of interleukin-8 as estrogen
receptor-regulated factor involved in breast cancer invasion and
angiogenesis by protein arrays. Int J Cancer. 109:507–515. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nannuru KC, Sharma B, Varney ML and Singh
RK: Role of chemokine receptor CXCR2 expression in mammary tumor
growth, angiogenesis and metastasis. J Carcinog. 10:402011.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Welch DR and Hurst DR: Defining the
hallmarks of metastasis. Cancer Res. 79:3011–3027. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
van der Horst G, Bos L and van der Pluijm
G: Epithelial plasticity, cancer stem cells, and the
tumor-supportive stroma in bladder carcinoma. Mol Cancer Res.
10:995–1009. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
De Larco JE, Wuertz BR, Rosner KA,
Erickson SA, Gamache DE, Manivel JC and Furcht LT: A potential role
for interleukin-8 in the metastatic phenotype of breast carcinoma
cells. Am J Pathol. 158:639–646. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
De Larco JE, Wuertz BR, Yee D, Rickert BL
and Furcht LT: Atypical methylation of the interleukin-8 gene
correlates strongly with the metastatic potential of breast
carcinoma cells. Proc Natl Acad Sci USA. 100:13988–13993. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Singh B, Berry JA, Vincent LE and Lucci A:
Involvement of IL-8 in COX-2-mediated bone metastases from breast
cancer. J Surg Res. 134:44–51. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kamalakar A, Bendre MS, Washam CL, Fowler
TW, Carver A, Dilley JD, Bracey JW, Akel NS, Margulies AG, Skinner
RA, et al: Circulating interleukin-8 levels explain breast cancer
osteolysis in mice and humans. Bone. 61:176–185. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Waugh DJ and Wilson C: The interleukin-8
pathway in cancer. Clin cancer Res. 14:6735–6741. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Jin K, Pandey NB and Popel AS: Crosstalk
between stromal components and tumor cells of TNBC via secreted
factors enhances tumor growth and metastasis. Oncotarget.
8:60210–60222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Halpern JL, Kilbarger A and Lynch CC:
Mesenchymal stem cells promote mammary cancer cell migration in
vitro via the CXCR2 receptor. Cancer Lett. 308:91–99. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yu PF, Huang Y, Han YY, Lin LY, Sun WH,
Rabson AB, Wang Y and Shi YF: TNFα-activated mesenchymal stromal
cells promote breast cancer metastasis by recruiting
CXCR2+ neutrophils. Oncogene. 36:482–490. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Marquette C and Nabell L:
Chemotherapy-resistant metastatic breast cancer. Curr Treat Options
Oncol. 13:263–275. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chen Y and Zhang Y: Application of the
CRISPR/Cas9 system to drug resistance in breast cancer. Adv Sci
(Weinh). 5:17009642018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Sharma B, Nawandar DM, Nannuru KC, Varney
ML and Singh RK: Targeting CXCR2 enhances chemotherapeutic
response, inhibits mammary tumor growth, angiogenesis, and lung
metastasis. Mol Cancer Ther. 12:799–808. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Shi Z, Yang WM, Chen LP, Yang DH, Zhou Q,
Zhu J, Chen JJ, Huang RC, Chen ZS and Huang RP: Enhanced
chemosensitization in multidrug-resistant human breast cancer cells
by inhibition of IL-6 and IL-8 production. Breast Cancer Res Treat.
135:737–747. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Jia D, Li L, Andrew S, Allan D, Li X, Lee
J, Ji G, Yao Z, Gadde S, Figeys D and Wang L: An autocrine
inflammatory forward-feedback loop after chemotherapy withdrawal
facilitates the repopulation of drug-resistant breast cancer cells.
Cell Death Dis. 8:e29322017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Stassi G, Garofalo M, Zerilli M,
Ricci-Vitiani L, Zanca C, Todaro M, Aragona F, Limite G, Petrella G
and Condorelli G: PED mediates AKT-dependent chemoresistance in
human breast cancer cells. Cancer Res. 65:6668–6675. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Festuccia C, Gravina GL, D'Alessandro AM,
Millimaggi D, Di Rocco C, Dolo V, Ricevuto E, Vicentini C and
Bologna M: Downmodulation of dimethyl transferase activity enhances
tumor necrosis factor-related apoptosis-inducing ligand-induced
apoptosis in prostate cancer cells. Int J Oncol. 33:381–388.
2008.PubMed/NCBI
|
|
80
|
Zanca C, Cozzolino F, Quintavalle C, Di
Costanzo S, Ricci-Vitiani L, Santoriello M, Monti M, Pucci P and
Condorelli G: PED interacts with Rac1 and regulates cell
migration/invasion processes in human non-small cell lung cancer
cells. J Cell Physiol. 225:63–72. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Yang SX, Costantino JP, Kim C, Mamounas
EP, Nguyen D, Jeong JH, Wolmark N, Kidwell K, Paik S and Swain SM:
Akt phosphorylation at Ser473 predicts benefit of paclitaxel
chemotherapy in node-positive breast cancer. J Clin Oncol.
28:2974–2981. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zatelli MC, Molé D, Tagliati F, Minoia M,
Ambrosio MR and Degli Uberti E: Cyclo-oxygenase 2 modulates
chemoresistance in breast cancer cells involving NF-kappaB. Cell
Oncol. 31:457–465. 2009.PubMed/NCBI
|
|
83
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
van Nijnatten TJA, Moossdorff M, de Munck
L, Goorts B, Vane MLG, Keymeulen KBMI, Beets-Tan RGH, Lobbes MBI
and Smidt ML: TNM classification and the need for revision of pN3a
breast cancer. Eur J Cancer. 79:23–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Charafe-Jauffret E, Ginestier C, Iovino F,
Wicinski J, Cervera N, Finetti P, Hur MH, Diebel ME, Monville F,
Dutcher J, et al: Breast cancer cell lines contain functional
cancer stem cells with metastatic capacity and a distinct molecular
signature. Cancer Res. 69:1302–1313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Chen L, Fan J, Chen H, Meng Z, Chen Z,
Wang P and Liu L: The IL-8/CXCR1 axis is associated with cancer
stem cell-like properties and correlates with clinical prognosis in
human pancreatic cancer cases. Sci Rep. 4:59112014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Magnifico A, Albano L, Campaner S, Delia
D, Castiglioni F, Gasparini P, Sozzi G, Fontanella E, Menard S and
Tagliabue E: Tumor-initiating cells of HER2-positive carcinoma cell
lines express the highest oncoprotein levels and are sensitive to
trastuzumab. Clin Cancer Res. 15:2010–2021. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Cameron D, Casey M, Press M, Lindquist D,
Pienkowski T, Romieu CG, Chan S, Jagiello-Gruszfeld A, Kaufman B,
Crown J, et al: A phase III randomized comparison of lapatinib plus
capecitabine versus capecitabine alone in women with advanced
breast cancer that has progressed on trastuzumab: Updated efficacy
and biomarker analyses. Breast Cancer Res Treat. 112:533–543. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Fernando RI, Castillo MD, Litzinger M,
Hamilton DH and Palena C: IL-8 signaling plays a critical role in
the epithelial-mesenchymal transition of human carcinoma cells.
Cancer Res. 71:5296–5306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Singh JK, Farnie G, Bundred NJ, Simões BM,
Shergill A, Landberg G, Howell SJ and Clarke RB: Targeting CXCR1/2
significantly reduces breast cancer stem cell activity and
increases the efficacy of inhibiting HER2 via HER2-dependent and
-independent mechanisms. Clin Cancer Res. 19:643–656. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Harrison H, Farnie G, Howell SJ, Rock RE,
Stylianou S, Brennan KR, Bundred NJ and Clarke RB: Regulation of
breast cancer stem cell activity by signaling through the Notch4
receptor. Cancer Res. 70:709–718. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Luo M, Fan H, Nagy T, Wei H, Wang C, Liu
S, Wicha MS and Guan JL: Mammary epithelial-specific ablation of
the focal adhesion kinase suppresses mammary tumorigenesis by
affecting mammary cancer stem/progenitor cells. Cancer Res.
69:466–474. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Liu S, Ginestier C, Ou SJ, Clouthier SG,
Patel SH, Monville F, Korkaya H, Heath A, Dutcher J, Kleer CG, et
al: Breast cancer stem cells are regulated by mesenchymal stem
cells through cytokine networks. Cancer Res. 71:614–624. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Li HJ, Reinhardt F, Herschman HR and
Weinberg RA: Cancer-stimulated mesenchymal stem cells create a
carcinoma stem cell niche via prostaglandin E2 signaling. Cancer
Discov. 2:840–855. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Ginestier C, Liu S, Diebel ME, Korkaya H,
Luo M, Brown M, Wicinski J, Cabaud O, Charafe-Jauffret E, Birnbaum
D, et al: CXCR1 blockade selectively targets human breast cancer
stem cells in vitro and in xenografts. J Clin Invest. 120:485–497.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wang Y, Tu L, Du C, Xie X, Liu Y, Wang J,
Li Z, Jiang M, Cao D, Yan X and Luo F: CXCR2 is a novel cancer
stem-like cell marker for triple-negative breast cancer. Onco
Targets Ther. 11:5559–5567. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kumar S, Wilkes DW, Samuel N, Blanco MA,
Nayak A, Alicea-Torres K, Gluck C, Sinha S, Gabrilovich D and
Chakrabarti R: ΔNp63-driven recruitment of myeloid-derived
suppressor cells promotes metastasis in triple-negative breast
cancer. J Clin Invest. 128:5095–5109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Uddin MM, Zou Y, Sharma T, Gatla HR and
Vancurova I: Proteasome inhibition induces IKK-dependent
interleukin-8 expression in triple negative breast cancer cells:
Opportunity for combination therapy. PLoS One. 13:e02018582018.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Schott AF, Goldstein LJ, Cristofanilli M,
Ruffini PA, McCanna S, Reuben JM, Perez RP, Kato G and Wicha M:
Phase Ib pilot study to evaluate reparixin in combination with
weekly paclitaxel in patients with HER-2-negative metastatic breast
cancer. Clin Cancer Res. 23:5358–5365. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Jones SA, Moser B and Thelen M: A
comparison of post-receptor signal transduction events in Jurkat
cells transfected with either IL-8R1 or IL-8R2. Chemokine mediated
activation of p42/p44 MAP-kinase (ERK-2). FEBS Lett. 364:211–214.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xue MQ, Liu J, Sang JF, Su L and Yao YZ:
Expression characteristic of CXCR1 in different breast tissues and
the relevance between its expression and efficacy of neo-adjuvant
chemotherapy in breast cancer. Oncotarget. 8:48930–48937.
2017.PubMed/NCBI
|
|
102
|
Ruffini PA: The CXCL8-CXCR1/2 axis as a
Therapeutic target in breast cancer stem-like cells. Front Oncol.
9:402019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Murugan AK, Grieco M and Tsuchida N: RAS
mutations in human cancers: Roles in precision medicine. Semin
Cancer Biol. Jun 27–2019.(Epub ahead of print). View Article : Google Scholar
|
|
104
|
Kufareva I, Salanga CL and Handel TM:
Chemokine and chemokine receptor structure and interactions:
Implications for therapeutic strategies. Immunol Cell Biol.
93:372–383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Ning Y, Labonte MJ, Zhang W, Bohanes PO,
Gerger A, Yang D, Benhaim L, Paez D, Rosenberg DO, Nagulapalli
Venkata KC, et al: The CXCR2 antagonist, SCH-527123, shows
antitumor activity and sensitizes cells to oxaliplatin in
preclinical colon cancer models. Mol Cancer Ther. 11:1353–1364.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Singh S, Sadanandam A, Nannuru KC, Varney
ML, Mayer-Ezell R, Bond R and Singh RK: Small-molecule antagonists
for CXCR2 and CXCR1 inhibit human melanoma growth by decreasing
tumor cell proliferation, survival, and angiogenesis. Clin Cancer
Res. 15:2380–2386. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Liu X, Peng J, Sun W, Yang S, Deng G, Li
F, Cheng JW and Gordon JR: G31P, an antagonist against CXC
chemokine receptors 1 and 2, inhibits growth of human prostate
cancer cells in nude mice. Tohoku J Exp Med. 228:147–156. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Bieche I, Chavey C, Andrieu C, Busson M,
Vacher S, Le Corre L, Guinebretière JM, Burlinchon S, Lidereau R
and Lazennec G: CXC chemokines located in the 4q21 region are
up-regulated in breast cancer. Endocr Relat Cancer. 14:1039–1052.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Varney ML, Singh S, Li A, Mayer-Ezell R,
Bond R and Singh RK: Small molecule antagonists for CXCR2 and CXCR1
inhibit human colon cancer liver metastases. Cancer Lett.
300:180–188. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Brandolini L, Cristiano L, Fidoamore A, De
Pizzol M, Di Giacomo E, Florio TM, Confalone G, Galante A, Cinque
B, Benedetti E, et al: Targeting CXCR1 on breast cancer stem cells:
Signaling pathways and clinical application modelling. Oncotarget.
6:43375–43394. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Steele CW, Karim SA, Leach JDG, Bailey P,
Upstill-Goddard R, Rishi L, Foth M, Bryson S, McDaid K, Wilson Z,
et al: CXCR2 inhibition profoundly suppresses metastases and
augments immunotherapy in pancreatic ductal adenocarcinoma. Cancer
Cell. 29:832–845. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Hirose K, Hakozaki M, Nyunoya Y, Kobayashi
Y, Matsushita K, Takenouchi T, Mikata A, Mukaida N and Matsushima
K: Chemokine gene transfection into tumour cells reduced
tumorigenicity in nude mice in association with neutrophilic
infiltration. Br J Cancer. 72:708–714. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Nair P, Gaga M, Zervas E, Alagha K,
Hargreave FE, O'Byrne PM, Stryszak P, Gann L, Sadeh J and Chanez P;
Study Investigators, : Safety and efficacy of a CXCR2 antagonist in
patients with severe asthma and sputum neutrophils: A randomized,
placebo-controlled clinical trial. Clin Exp Allergy. 42:1097–1103.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Allegretti M, Cesta MC, Garin A and
Proudfoot AE: Current status of chemokine receptor inhibitors in
development. Immunol Lett. 145:68–78. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Maroulakou IG, Oemler W, Naber SP and
Tsichlis PN: Akt1 ablation inhibits, whereas Akt2 ablation
accelerates, the development of mammary adenocarcinomas in mouse
mammary tumor virus (MMTV)-ErbB2/neu and MMTV-polyoma middle T
transgenic mice. Cancer Res. 67:167–177. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Liu H, Radisky DC, Nelson CM, Zhang H,
Fata JE, Roth RA and Bissell MJ: Mechanism of Akt1 inhibition of
breast cancer cell invasion reveals a protumorigenic role for TSC2.
Proc Natl Acad Sci USA. 103:4134–4139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Li B, Hou D, Guo H, Zhou H, Zhang S, Xu X,
Liu Q, Zhang X, Zou Y, Gong Y and Shao C: Resveratrol sequentially
induces replication and oxidative stresses to drive p53-CXCR2
mediated cellular senescence in cancer cells. Sci Rep. 7:2082017.
View Article : Google Scholar : PubMed/NCBI
|














