Introduction
Colorectal cancer (CRC) has become the third most
common type of cancer and the second leading cause of
cancer-associated mortality worldwide (1). In the United States, the 5-year
relative survival rate is 90.1 or 69.2% for patients with CRC with
localized (stage I–II) or regional metastasis (stage III),
respectively. However, the 5-year relative survival rate is only
11.7% for patients with distant metastasis (stage IV) (2). Therefore, improving the diagnosis and
treatment efficacy of patients with stage IV CRC is the key to
ameliorating the overall survival of CRC.
The American Society of Clinical Oncology (ASCO)
Annual Meeting in 2016 named ‘Immunotherapy’ as the ‘Primary
Progress’ in Cancer Research in 2015 (3). The Federal Drug Agency and China Food
and Drug Administration (CFDA) have approved pembrolizumab, an
anti-PD-1 inhibitor, for the treatment of microsatellite
instability-high (MSI-H) or mismatch repair-deficient (dMMR)
metastatic CRC (mCRC) (4). Numerous
studies have reported that 44.8–89% of patients with CRC express
PD-L1, thus suggesting that abnormal expression of PD-L1 is an
independent poor prognostic indicator in CRC (5,6).
However, data from clinical trials revealed that PD-1/PD-L1
blockades are only effective in 2.5–5% of patients with mCRC (MSI-H
or dMMR), and the vast majority of patients [microsatellite stable
(MSS)] does not benefit from this treatment (7–9). This is
a great obstacle to the application of immunotherapy in CRC.
Chen and Mellman (10) analysed data from patients undergoing
immunotherapy. According to the clinical efficacy and the
immunophenotype of the patients, the tumor immuno-microenvironment
was divided into three basic types: The immune-desert phenotype,
immune-excluded tumor and inflamed tumor (10). Based on this classification, the
effectiveness of PD-L1 inhibitors depends not only on the
expression of intra-tumor PD-L1 but also on the existence of
sufficient immune effector cells, particularly CD8+ T
cells, in the tumor microenvironment (TME). Without the
infiltration of CD8+ T cells, even if elevated
expression of PD-L1 is detected in tumor tissues, a therapeutic
effect could not be achieved; this is the case for patients with
the immune-desert phenotype (10).
It has previously been reported that the number of
tumor-infiltrating CD4+ and CD8+ T cells in
patients with MSI-H CRC who benefit from pembrolizumab
immunotherapy is significantly higher than that in MSS CRC
(11). This may be due to patients
with MSI-H CRC producing a large number of mutant proteins in tumor
tissues, which can stimulate the immune response and promote the
local lymphocyte infiltration of tumors (7,12).
Therefore, promoting the infiltration of immune effector cells,
particularly CD8+ T cells, into tumor tissues and
improving their immune function are effective measures to increase
the efficacy of immunotherapy in the majority of patients with CRC
(MSS phenotype).
Cyclin-dependent kinase 9 (CDK9) is a key regulator
of transcriptional elongation, which is a promising therapeutic
target in cancer, particularly for types of cancer driven by
transcriptional dysregulation (13,14).
However, the mechanism and clinical transformation of CDK9 in CRC
have been rarely reported (15). In
addition, it has been demonstrated that APC/BRAF/SMAD4 gene
mutations lead to upregulation of transcription, which leads to the
occurrence and development of CRC (16–18).
Positive transcription elongation factor (P-TEFb)/CDK9 triggers the
release and nuclear export of β-catenin by the α-catenin:APC
complex (19). CDK8 and CDK9 provide
the coordinated regulation of SMAD transcriptional activators in
the bone morphogenetic protein and transforming growth factor β1
signalling pathways (20).
Therefore, CDK9 serves an important role in the development of CRC.
In addition, studies have demonstrated that CDK9 promotes the
proliferation and differentiation of immune cells, including
neutrophils, macrophages and lymphocytes, alters the ratio of
CD4+CD25+ forkhead box P3+ Tregs,
and regulates the expression of chemokines, thereby affecting the
infiltration of effector T cells in an inflammatory environment
(21–23). The present study utilized human
cancer datasets and clinically resected CRC specimens to assess
CDK9 expression and CD8+ T cell infiltration, and
examined their alterations in mCRC. The results of the present
study suggested that CDK9 may be a promising therapeutic target for
patients with MSS mCRC.
Materials and methods
Analysis of public datasets
RNA sequencing-based gene expression data in CRC was
obtained from the Gene Expression Profiling Interactive Analysis
(GEPIA) database for cancer genomics (24). The association between CDK9
expression and the survival of 605 patients with CRC, including 121
patients who died, was analysed using the Tumor Immune Estimation
Resource (TIMER) database (25). The
co-expression of proteins was analysed by the Search Tool for the
Retrieval of Interacting Genes/Proteins (STRING) database (26). For gene expression, P<0.01 was
used as the cut-off. The Kaplan-Meier method with a log-rank test
was used for estimating patient survival rates. P<0.05 was
considered to indicate a statistically significant difference.
Patient tissue samples
A total of 35 patients with CRC who were admitted
and treated at Tianjin Medical University Cancer Institute and
Hospital between May 2018 and September 2018 were enrolled
randomly. The patients comprised 16 men and 19 women, aged 27–79
years. During surgical resection, specimen (both tumor and paired
control tissues) were obtained from the treatment-naïve patients
with CRC. The gene stability of BAT-25, BAT-26, NR-21, NR-24
and Mono-27 in the specimens from all patients was examined
using next-generation sequencing (Hongzhong Precision Medicine).
All patients had MSS CRC according to the definition of the
National Cancer Institute (there was no ‘instability’ in the
results of the five aforementioned loci) (27). All samples and clinical data were
collected after ethical approval was granted by the Ethics
Committee of Tianjin Medical University Cancer Institute and
Hospital. Written informed consent was obtained from all patients
for participation in the present study. The clinical features of
the patients are presented in Table
I. The records of all of the patients contained basic
information, including Tumor-Node-Metastasis (TNM) stage, the
degree of differentiation, lymph node metastasis and distant
metastasis, according to the 2002 International Cancer Alliance TNM
staging criteria (28).
 | Table I.Clinical and pathological
characteristics of the patients with colorectal cancer. |
Table I.
Clinical and pathological
characteristics of the patients with colorectal cancer.
| Patient no. | Sex | Age, years | Primary tumor
location | TNM stage | Tumor stage
(28) |
|---|
| P#1 | Male | 61 | Ascending
colon | T4aN1bM0 | IIIB |
| P#2 | Female | 77 | Rectum | T3N1bM0 | IIIB |
| P#3 | Female | 64 | Descending
colon | T4bN2bM0 | IIIC |
| P#4 | Male | 79 | Ascending
colon | T3N2aM0 | IIIB |
| P#5 | Female | 60 | Rectum | T3N0M1c | IVC |
| P#6 | Female | 60 | Ascending
colon | T3N2bM1a | IVA |
| P#7 | Female | 49 | Descending
colon | T3N2bM0 | IIIB |
| P#8 | Female | 41 | Rectum | T3N2bM0 | IIIB |
| P#9 | Male | 67 | Sigmoid | T3N2bM1b | IVB |
| P#10 | Male | 65 | Ascending
colon | T2N1bM0 | IIIA |
| P#11 | Female | 27 | Rectum | T4aN1bM0 | IIIB |
| P#12 | Female | 54 | Rectum | T2N1cM0 | IIIA |
| P#13 | Female | 70 | Rectum | T3N1cM0 | IIIB |
| P#14 | Male | 56 | Descending
colon | T3N2aM0 | IIIB |
| P#15 | Female | 68 | Sigmoid | T3N1bM0 | IIIB |
| P#16 | Male | 73 | Rectum | T3N2aM1a | IVA |
| P#17 | Male | 50 | Descending
colon | T3N2aM1a | IVA |
| P#18 | Female | 65 | Ascending
colon | T4bN0M1a | IVA |
| P#19 | Female | 53 | Ascending
colon | T4aN2aM1a | IVA |
| P#20 | Male | 76 | Ascending
colon | T4bN2bM1a | IVA |
| P#21 | Female | 48 | Ileocecus | T4bN0M1b | IVB |
| P#22 | Male | 48 | Ascending
colon | T4bN2aM1b | IVB |
| P#23 | Male | 70 | Descending
colon | T4aN2M1a | IVA |
| P#24 | Female | 65 | Ascending
colon | T3N2M1a | IVA |
| P#25 | Male | 65 | Ascending
colon | T4aN0M1b | IVB |
| P#26 | Female | 60 | Transverse
colon | T4bN0M1b | IVB |
| P#27 | Male | 77 | Transverse
colon | T3N1aM1b | IVB |
| P#28 | Female | 29 | Ascending
colon | T3N1M1b | IVB |
| P#29 | Male | 59 | Ascending
colon | T3N1bM0 | IIIB |
| P#30 | Female | 79 | Rectum | T2N2bM0 | IIIB |
| P#31 | Male | 60 | Sigmoid | T4aN2bM0 | IIIC |
| P#32 | Male | 52 | Ascending
colon | T4aN2bM0 | IIIC |
| P#33 | Female | 68 | Ascending
colon | T3N1cM0 | IIIB |
| P#34 | Male | 62 | Sigmoid | T4aN1bM0 | IIIB |
| P#35 | Female | 65 | Ascending
colon | T3N1aM0 | IIIB |
Isolation of tissue-infiltrating
cells
Fresh colorectal tumor tissues and paired control
tissues from patients with CRC were prepared by mechanical
disruption, followed by digestion with 0.5 mg/ml collagenase type
IV (cat. no. C5138; Sigma-Aldrich; Merck KGaA) in 10% FBS with 10
U/ml DNase I in RPMI-1640 medium (both from Gibco; Thermo Fisher
Scientific, Inc.) at 37°C for 30 min. Digested tissues were
incubated for 5 min at 37°C with EDTA (0.5 M) to prevent dendritic
cell/T-cell aggregation and filtered through a 100-µm and a 40-µm
filter. The isolation and culture of tissue/tumor-infiltrating
cells were performed as previously described (29).
Immunohistochemistry (IHC)
Colon and rectal tumor tissues, and paired normal
tissues were were fixed with 4% formaldehyde for 24 h at room
temperature (RT), embedded in paraffin and cut into 4-µm sections.
The sections were dewaxed with xylene then hydrated through a
graded series of ethanol (100, 95, 90, 80 and 70%) for 5 min in
each percentage at RT. Slides were boiled in 10 mM sodium citrate
buffer pH 6.0 at 95°C for 10 min and subsequently incubated in 3%
hydrogen peroxide for 10 min. Sections were blocked with 100–400 µl
5% normal goat serum in TBS-Tween (cat. no. 5425; Cell Signaling
Technology, Inc.) for 1 h at RT. To assess CDK9 expression in the
MSS phenotype mCRC specimens, sections were incubated with a CDK9
antibody (dilution, 1:100; cat. no. 2316; Cell Signaling
Technology, Inc.) overnight at 4°C, and were then incubated with a
horseradish peroxidase-conjugated secondary antibody (dilution,
1:1,000; cat. no. ab6721; Abcam) at room temperature for 1 h.
Following washing with PBS, the sections were incubated for 30 min
at RT with streptavidin-biotin conjugated with horseradish
peroxidase [dilution, 1:100; cat. no. KIT-0305; UltraSensitive™ SP
(Mouse/Rabbit) IHC kit]. Subsequently, the slides were stained with
3,3′-diaminobenzidine (Fuzhou Maixin Biotech Co., Ltd.), and the
nuclei were counterstained with haematoxylin for 5 min at RT.
Morphometric analyses of the tumor and paired normal tissues were
performed using an Olympus BX51 light microscope (magnification,
×20; Olympus Corporation). Images were obtained from 10 randomly
selected areas.
Staining intensity was scored as follows: 0,
Negative; 1, weakly positive (<25% of cells stained); and 2,
positive (>25% of cells stained). Samples were subsequently
grouped into 2 categories: Low expression (0 and 1) and high
expression (2).
Flow cytometric analysis
Tumor/paired control tissue-infiltrating cells
(1×106 cells) were incubated with human FITC anti-CD3
monoclonal antibody (mAb) (1:20; cat. no. 300452; BioLegend, Inc.),
APC anti-CD8 mAb (1:20; cat. no. 344722; BioLegend, Inc.),
phycoerythrin (PE) anti-CD39 mAb (1:20; cat. no. 328208; BioLegend,
Inc.), APC/Cy7 anti-PD-1 mAb (1:20; cat. no. 329921; BioLegend,
Inc.), and PE/Cy7 anti-T-cell immunoglobulin mucin family member 3
(Tim-3) mAb (1:20; cat. no. 345013; BioLegend, Inc.) in cell
staining buffer (cat. no. 420201; BioLegend, Inc.) for 15 min at
RT. After washing with PBS and centrifugation at 400 × g for 5 min
at 4°C, 1×106 cells were suspended in 300 µl cell
staining buffer (cat. no. 420201; BioLegend, Inc.) and analysed on
a BD FACSCalibur (BD Biosciences) flow cytometer. The data were
analysed using FlowJo v10 software (FlowJo, LLC).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism v5 software (GraphPad Software, Inc.). Data are presented as
the mean ± standard error of the mean of three repeats. A
χ2 test was used to evaluate the association between
CDK9 expression and the clinicopathological parameters. A log-rank
test was used to compare the survival curves of two groups. Cox
regression analysis was performed for multivariate analysis of
prognostic variables. A correlation between transcripts per million
(TPM) of CDK9 and other genes was calculated for statistical
significance, correlation co-efficient, and is represented using a
scatter plot. One-way ANOVA followed by Tukey's multiple comparison
test was used for multiple-group analyses. P<0.05 was considered
to indicate a statistically significant difference.
Results
CDK9 significantly shortens the
survival of patients with colon cancer
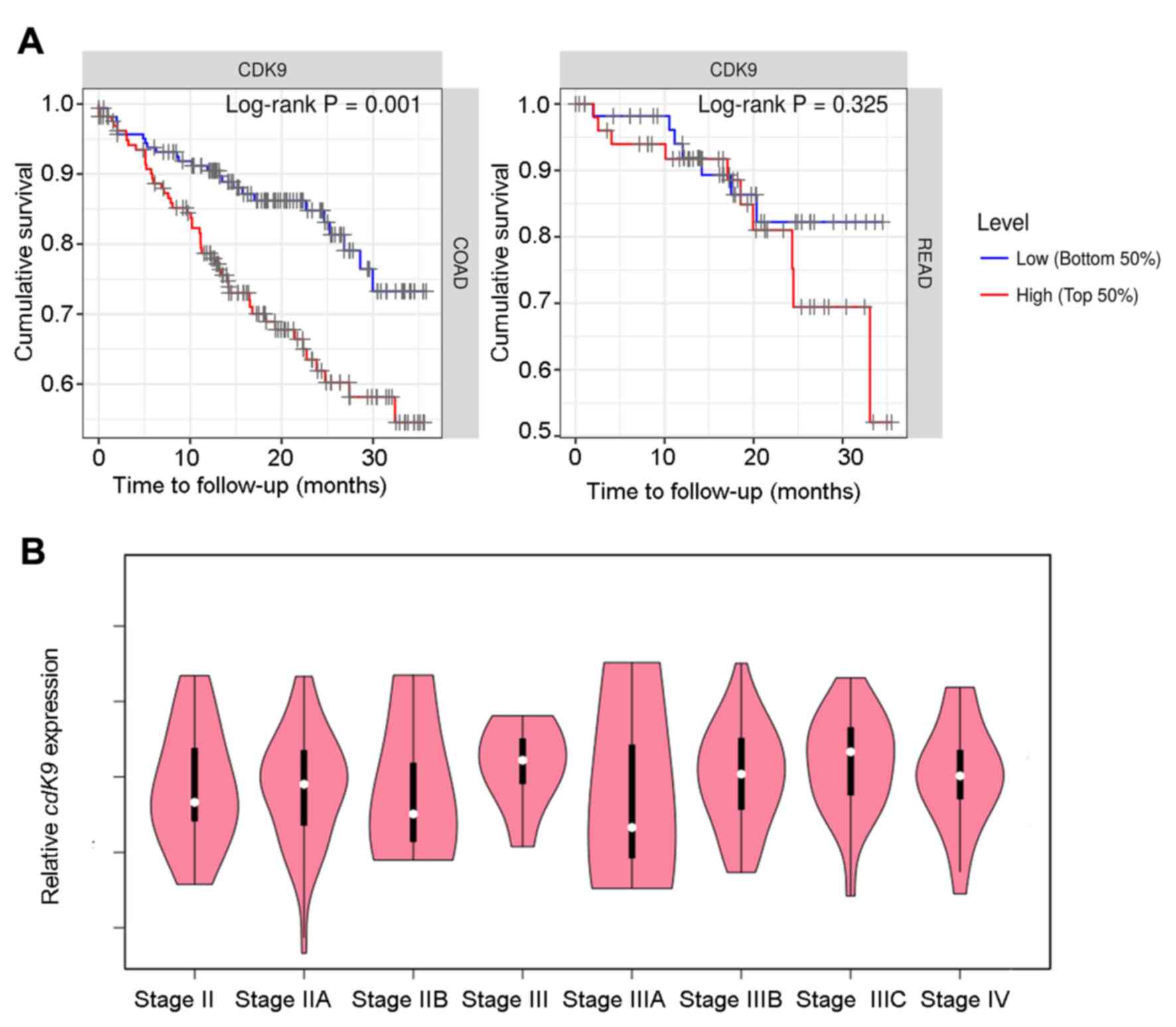
To examine the association between CDK9 and
prognosis in patients with colon and rectal cancer, the present
study analysed the TIMER database, and found that high CDK9
expression was significantly associated with a shortened survival
of patients with colon cancer (P=0.001; 445 cases total; 98 cases
of mortality). The same trend was observed in patients with rectal
cancer; however, this was not statistically significant (P=0.325;
160 cases total; 23 cases of mortality; Fig. 1A). As shown in Table II, CDK9 was a risk factor for
survival in patients with colon cancer based on a Cox proportional
hazard model analysis. However, Cox model analysis demonstrated
that CDK9 did not affect rectal cancer progression (Table III). In addition, CDK9 expression
had no significant effect on prognosis when the survival time was
>3 years (data not shown). Therefore, CDK9 may serve an
important role in the progression of advanced colon cancer. The
GEPIA database was searched and CDK9 mRNA expression in
stage III–IV patients was upregulated compared with that in stage
II patients, and the expression level was positively associated
with the clinical tumor stage in stage IIIA-IIIC patients although
the association was not significant (Fig. 1B). These data suggested that CDK9 may
promote lymph node metastasis in colon cancer.
| Table II.Multivariate survival analyses of
stage and CDK9 with overall survival of patients with COAD using
the Cox proportional hazards model. |
Table II.
Multivariate survival analyses of
stage and CDK9 with overall survival of patients with COAD using
the Cox proportional hazards model.
| Factor | Coefficient | HR | 95% CI_l | 95% CI_u | P-value |
|---|
| Stage II | 0.646 | 1.908 | 0.737 | 4.941 | 0.183 |
| Stage III | 1.166 | 3.208 | 1.243 | 8.277 | 0.016a |
| Stage IV | 2.241 | 9.406 | 3.629 | 24.381 | 0.000b |
| CDK9 | 0.625 | 1.868 | 1.151 | 3.033 | 0.011a |
| Table III.Multivariate survival analyses of
stage and CDK9 with overall survival of patients with READ using
the Cox proportional hazards model. |
Table III.
Multivariate survival analyses of
stage and CDK9 with overall survival of patients with READ using
the Cox proportional hazards model.
| Factor | Coefficient | HR | 95% CI_l | 95% CI_u | P-value |
|---|
| Stage II | 0.156 | 1.169 | 0.225 | 6.090 | 0.853 |
| Stage III | 0.726 | 2.066 | 0.426 | 10.026 | 0.368 |
| Stage IV | 1.657 | 5.244 | 1.104 | 24.907 | 0.037a |
| CDK9 | 0.823 | 2.277 | 0.775 | 6.696 | 0.135 |
CDK9 is negatively associated with the
infiltration of CD8+ T cells in CRC
By analysing RNA sequencing (RNA-seq) data obtained
from GEPIA, and the pathological estimation data from human CRC
tumors, the present study revealed that patients with high
expression levels of CDK9 and fewer CD8+ T cells in the
TME exhibited poorer prognoses (Figs.
1A and 2A). Next it was
determined whether CDK9 affected infiltration of CD8+ T
cells in CRC. Specimens were collected from 35 patients with stage
III–IV MSS CRC, and the clinical and pathological characteristics
of the patients are shown in Table
III. CDK9 expression in these patients was detected by
immunohistochemical staining. The results demonstrated that the
CDK9-positive expression rate (a score of 2) in stage III
right-sided colon cancer was 85.7% (6/7), which was significantly
higher than that in paracancerous tissues, with a positivity rate
of 28.6% (2/7; χ2=4.667; P=0.03; Fig. 2B and C). However, there was no
significant difference identified between CDK9 expression in all
cases of left-sided colon cancer and rectal cancer (3/12) compared
with the corresponding right-sided cancer (4/12; Fig. 2C). CDK9 expression in stage IV colon
cancer was much higher than that in the paracancerous region,
regardless of the primary tumor site (Fig. 2D). Clinically, the survival of
patients with stage III–IV right-sided colon cancer was
significantly shorter than that of patients with left-sided colon
cancer (30,31). Therefore, these results suggested
that abnormally high CDK9 expression predicted poor prognosis, and
this may be a significant factor in the prognosis of patients with
left/right-sided colon cancer.
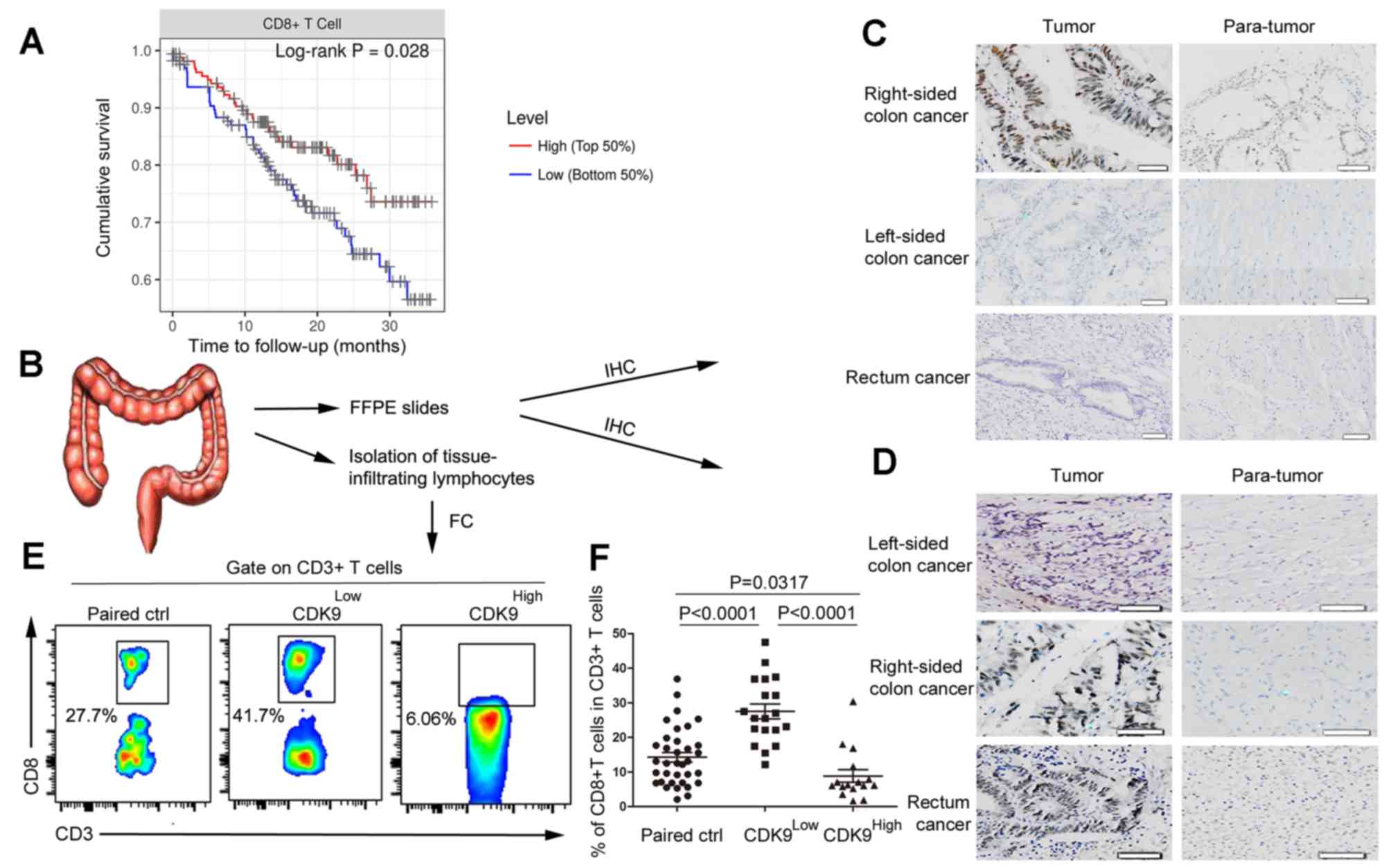 | Figure 2.CDK9 is negatively associated with
the infiltration of CD8+ T cells in colorectal cancer.
(A) Kaplan-Meier curves for CD8+ T cell infiltration in
colorectal cancer. Levels were divided into low and high levels
(cut-off, 50%). The P-value was calculated using a log-rank test.
(B) Experimental scheme. (C) Immunohistochemistry of CDK9
expression in stage III colorectal cancer tissues and adjacent
non-cancerous tissues. Scale bar, 50 µm. (D) Immunohistochemistry
of CDK9 expression in stage IV colorectal cancer tissues and
adjacent non-cancerous tissues. Scale bar, 50 nm. (E) Flow
cytometric analysis of tumor-infiltrating CD8+ T cells
in stage III–IV colorectal cancer tissues and paired normal
tissues. (F) Frequency of tumor-infiltrating CD8+ T
cells in stage III–IV colorectal cancer tissues and paired normal
tissues. Each dot represents data generated from one patient
(n=35). CDK9, cyclin-dependent kinase 9; IHC, immunohistochemistry;
FFPE, formalin-fixed and paraffin-embedded; FC, flow cytometry;
Paired ctrl, paired normal control tissues; CDK9Low T,
tumor tissues with low expression of cyclin-dependent kinase 9;
CDK9High T, tumor tissues with high expression of
cyclin-dependent kinase 9. |
Additionally, cells were isolated from the fresh
tumor samples and matched normal tissues of these patients. Flow
cytometry was utilized to detect tumor infiltration by
CD8+ T cells. According to the immunohistochemical
staining results, patients with the MSS-phenotype CRC were divided
into the CDK9 high- and low-expression groups, and the results of
flow cytometry were analysed. The frequency of tumor-infiltrating
CD8+ T cells in the CDK9 low-expression group was
markedly higher than that in the CDK9 high-expression group
(Fig. 2E and F). This result
demonstrated that CDK9 inhibited the recruitment and infiltration
of CD8+ T cells in the TME.
CDK9 is associated with genes
responsible for immune cell migration and exhaustion in CRC
Subsequently, it was explored how CDK9 inhibited the
recruitment and migration of CD8+ T cells to the TME by
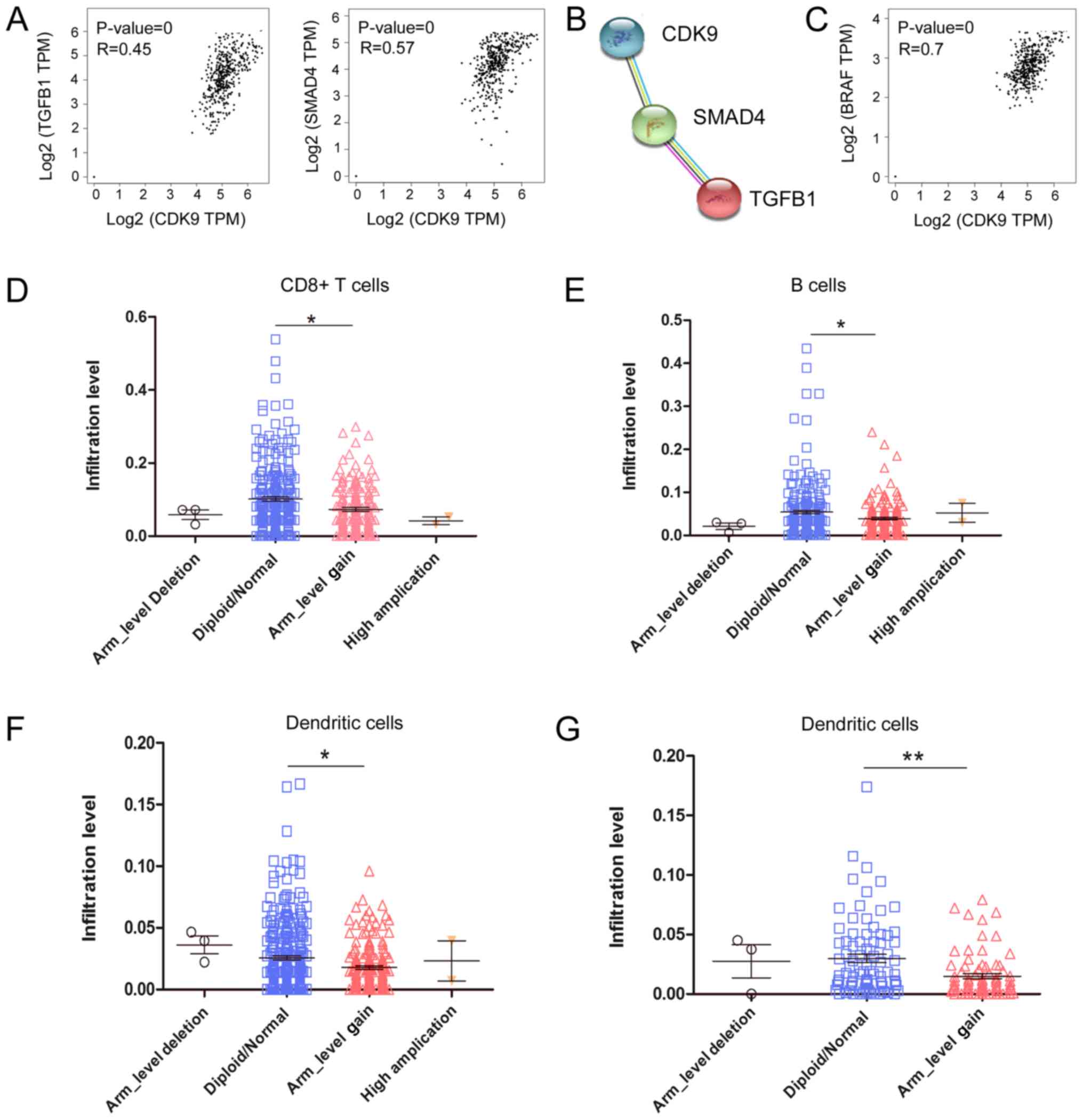
analysing RNA-seq data from CRC samples. As shown in Fig. 3A, CDK9 mRNA expression was
positively correlated with the expression of C-X-C motif chemokine
ligand 12 (CXCL12), C-C motif chemokine ligand 21
(CCL21) and atypical chemokine receptor 3 (ACKR3;
also referred to as CXCR7), which are associated with
lymphocyte migration. It has been reported that CXCL12 decreases
the number of tumor-infiltrating natural killer (NK) cells and
CD8+ T cells (32,33).
These data indicated that CDK9 may be involved in
CXCL12/CCL21/CXCR7-axis-mediated negative immune regulation in the
TME. Since CDK9 was associated with the expression of numerous
chemokines in the TME and affected the migration of immune cells,
the present study investigated whether CDK9 mediated ‘editing’ of
the TME, and the function and phenotype of immune cells. To explore
this, the present study analysed the association between CDK9 and
CD8+ T cell exhaustion-associated genes by analysing
RNA-seq data from CRC. Notably, there was a significant positive
correlation between the transcription of CDK9 and several
genes that induce T lymphocyte exhaustion, including nuclear factor
of activated T cells 1 (NFATC1), V-set immunoregulatory
receptor, nuclear factor of activated T cells 5 (NFAT5) and
ectonucleoside triphosphate diphosphohydrolase 1 (ENTPD1;
also referred to as CD39; Fig.
3B). Additionally, the anti-inflammatory molecules transforming
growth factor β1 (TGFB1) and SMAD4 were correlated
with CDK9 expression in CRC tumors (Fig.
4A). Furthermore, CDK9 was co-expressed with
TGFB1 and SMAD4 according to STRING database analysis
(Fig. 4B). Interestingly,
CDK9 was significantly positively correlated with
BRAF mRNA expression, which serves an important role in the
development of CRC (Fig. 4C). It was
revealed that 26.2% (96/367) of patients with CRC had BRAF
mutations, and their antitumor immunity was more inhibited than
that of wild-type patients, based on analysis of the TIMER
database. The decrease in tumor-infiltrating immune cells caused by
the BRAF mutation was significantly different between colon
and rectal cancer (Fig. 4D and E).
Compared with wild-type rectal cancer (diploid/normal), the
arm-level gain (affecting ≥50% of the chromosome) of BRAF
led to decreased tumor-infiltrating CD8+ T cells
(P<0.001), B cells (P<0.05) and dendritic cells (P<0.05)
in colon cancer (Fig. 4D-F), whereas
only tumor-infiltrating dendritic cells (P<0.01) were decreased
in rectal cancer (Fig. 4G). These
data indicated that CDK9 was involved in BRAF-mediated
immunosuppression, which was affected by the site of the primary
tumor. Overall, these data implied that CDK9 may contribute to CRC
immune escape by promoting CD8+ T cell exhaustion and
inhibiting the infiltration of numerous immune cell
populations.
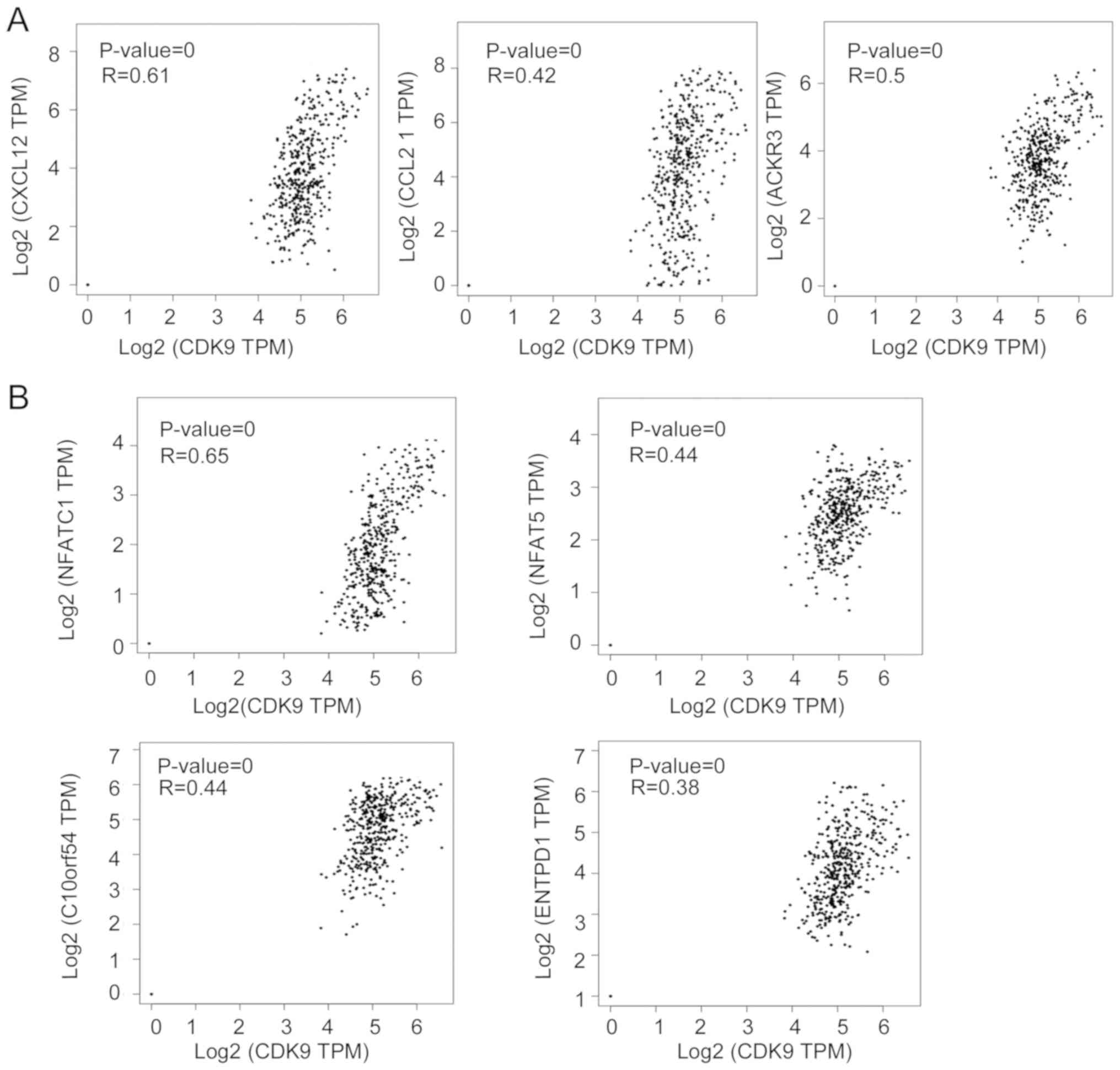 | Figure 3.CDK9 is correlated with genes
responsible for immune cell migration and exhaustion in colorectal
cancer. (A) Correlations of CDK9 expression with CXCL12,
CCL21 and ACKR3 mRNA expression in CRC. (B) Correlations
of CDK9 with NFATC1, NFAT5, C10orf54 and
ENTPD1 mRNA expression in CRC. CRC, colorectal cancer;
CDK9, cyclin-dependent kinase 9; CXCL12, C-X-C motif
chemokine ligand 12; CCL21, C-C motif chemokine ligand 21;
ACKR3, atypical chemokine receptor 3; NFATC1, nuclear
factor of activated T cells 1; NFAT5, nuclear factor of
activated T cells 5; C10orf54 V-set immunoregulatory
receptor; ENTPD1, ectonucleoside triphosphate
diphosphohydrolase 1; TPM, transcripts per million. |
Association of CDK9 expression with
tumor-infiltrating CD8+ T cell exhaustion in CRC
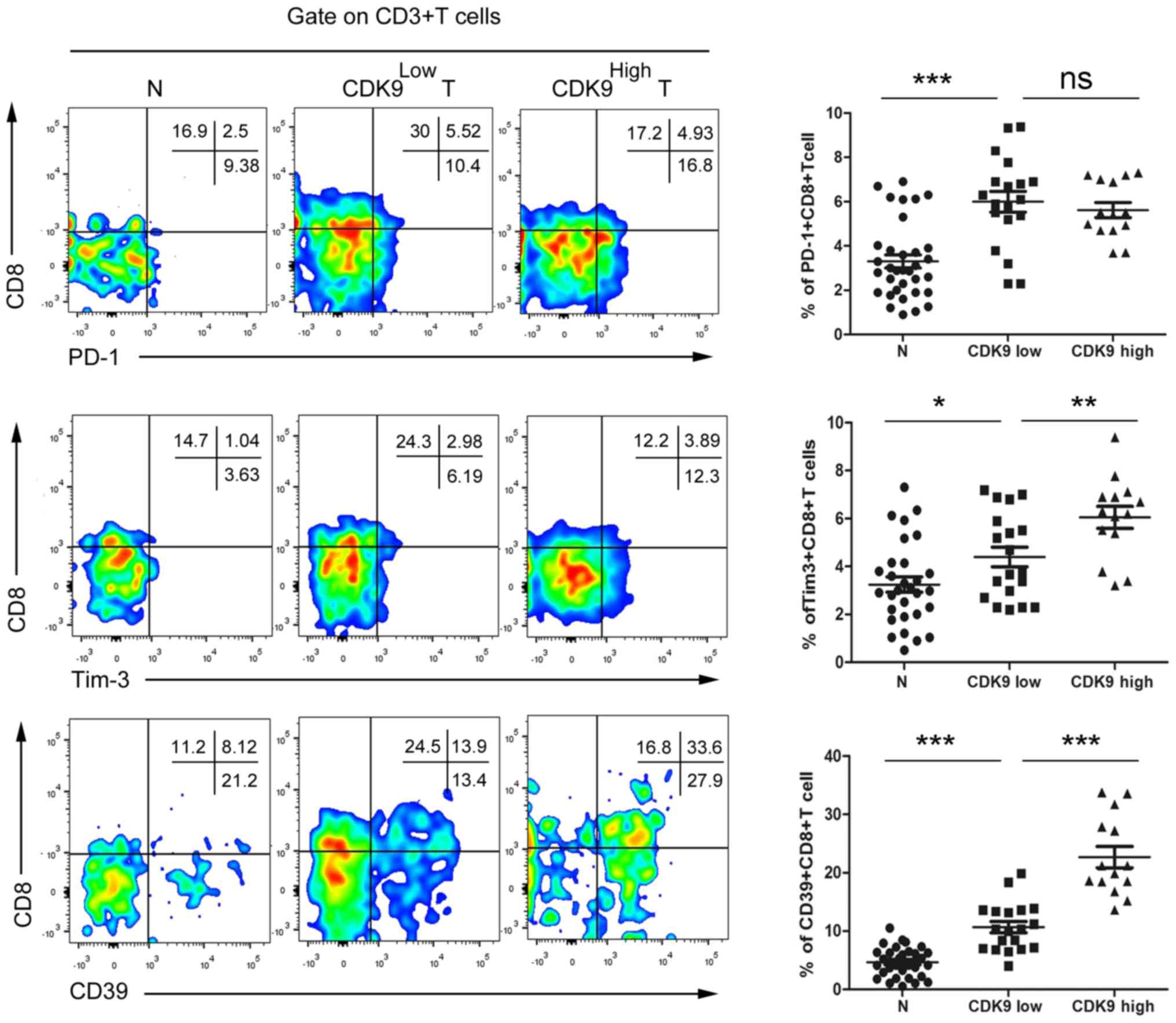
To confirm the association between CDK9 and
tumor-infiltrating CD8+ T cell exhaustion,
tumor-infiltrating cells were isolated from fresh CRC tissues and
paired normal tissues from advanced CRC tumors, and the cells were
co-cultured with antibodies and analysed by flow cytometry.
According to the aforementioned results of the CDK9
immunohistochemistry, patients with MSS phenotype CRC were divided
into two groups, a CDK9 high- and a CDK9 low-expression group, for
statistical analysis. The results revealed that there were no
significant differences identified in the
CD8+PD-1+ T cell frequency between the
CDK9high group and the CDK9low group
(Fig. 5). Compared with the
CDK9low group, the frequency of
CD8+Tim-3+ T cells was increased by 37.5% in
the CDK9high group (Fig.
5). Notably, the frequency of infiltrating
CD8+CD39+ T cells in CDK9high CRC
tumors was increased 1.12-fold compared with that in
CDK9low tumors (Fig. 5).
These data indicated that CDK9 was positively associated with
tumor-infiltrating CD8+ T cell exhaustion, independent
of the PD-1/PD-L1 signal, and may be used to evaluate the
immune-type of the TME in patients with MSS mCRC.
Discussion
CDK9 has been widely used in the research and
development of antitumor drugs due to its key regulatory role in
transcription, and its contribution to the progression and
maintenance of several types of cancer (34–37). A
preclinical in vivo study demonstrated that treatment with a
small-molecule CDK9 inhibitor impairs the growth of human melanoma
xenografts (38). BAY 1143572, a
novel and highly selective CDK9/P-TEFb inhibitor that is currently
being investigated in phase I studies, decreases c-Myc and MCL1
apoptosis regulator levels in ATL-derived or HTLV-1-transformed
lines, which inhibits their growth (39). It has been reported that CDK9
functions in immune responses and that its inhibition effectively
suppresses the inflammatory response in chondrocytes (40). However, the regulatory effect of CDK9
on the antitumor immune response in CRC remains to be elucidated.
The present study investigated CDK9 expression in different primary
tumor sites in CRC and provided evidence that CDK9 expression was
negatively associated with CD8+ T cell antitumor
function in MSS mCRC.
With the development of precision medicine, it has
become gradually understood that the prognoses of left- and
right-sided CRC are different. Venook's CALGB/SWOG 80405 clinical
data were published in ASCO in 2016 and revealed that the median
survival of KRas wild-type right-sided and left-sided mCRCs was
19.4 and 34.2 months, respectively, and that of KRas mutant
right-sided and left-sided mCRCs was 23.1 and 30.3 months,
respectively (3). Therefore, the
difference in primary tumor location has become a novel focus in
CRC research. The results of the present study demonstrated that
CDK9 expression in advanced left-sided CRC was significantly higher
than that in early-stage left-sided CRC, and CDK9 expression in
stage III–IV right-sided CRC was significantly higher than that in
stage III–IV left-sided CRC. Notably, the prognostic difference
between left-sided and right-sided CRC is only seen in tumors that
are at least at stage III (30).
Therefore, CDK9 may be a prognostic factor for left/right-sided
colon cancer, which broadens the knowledge for future research.
Since the 5-year relative survival of patients with
stage IV CRC is only 11.7%, regardless of primary tumor location,
improving the diagnosis and treatment efficacy of stage IV patients
is the key to ameliorating the survival of CRC (41). Anti-PD-1 immunotherapy has been
demonstrated to be effective only in patients with MSI-H phenotype
stage IV CRC, since they have more tumor-infiltrating immune cells
(11). However, only 3.5–5% of all
stage IV CRC cases are MSI-H (9).
The present study determined CDK9 expression and the frequency of
CD8+ T cells in tumor and paired normal control
specimens of 35 patients with MSS-phenotype CRC, which accounts for
>95% of all advanced CRC (9), and
identified that CDK9 was negatively associated with
tumor-infiltrating CD8+ T cells. This result suggested
that CDK9 inhibition may benefit patients with MSS phenotype CRC in
combination with anti-PD-1 immunotherapy.
The recruitment and migration of immune cells upon
stimulation are regulated by numerous factors, of which the
chemokine family serves an important role. It has been reported
that fibroblasts help pancreatic cancer to develop chemotherapy
resistance, and the chemokine CXCL12 is secreted by fibroblasts and
prevents the infiltration of CD8+ T cells into the tumor
(42). Zboralski et al
(33) demonstrated that inhibition
of CXCL12 by ‘NOX-A12’ can promote the infiltration of T cells and
NK cells, thereby increasing the efficacy of anti-PD-1 drugs in
colon cancer models. CXCR7/C-X-C motif chemokine receptor 4
heterodimer-induced histone demethylation promotes colorectal
tumorigenesis (43). In addition,
CCL21 is involved in T-cell migration and trafficking to secondary
lymphoid organs (44). The present
study revealed that CDK9 expression was positively
correlated with CXCL12, ACKR3 (also referred to as
CXCR7) and CCL21 expression, suggesting that CDK9 may
inhibit CD8+ T cell infiltration via the CXCL12/CXCR7
axis.
The present study investigated how the TME ‘edits’ T
cells in CRC and whether CDK9 serves a role in this process. tumor
antigens are weakly immunogenic self-molecules, and the majority of
tumor-specific T cells have a low T-cell receptor (TCR) affinity,
since tumor-specific T cells with high avidity are cleared during
the thymic selection process (45).
Therefore, the process of antigen presentation is impaired in the
TME, leading to insufficient priming and boosting of T cells
(45). Consequently, the complex
components in the TME drive T cells to terminally differentiate
into ‘exhausted’ T cells (46).
‘Exhausted’ CD8+ T cells overexpress a large number of
cell surface inhibitory receptors, including PD-1 and Tim-3. NFAT
activates the nuclear factor protein of T cells, which results in
inability of the NFATC1 gene to synergize with AP-1 during
gene modification, resulting in loss of the TCR signal and
upregulation of the expression of inhibitory receptors on the cell
surface, thereby weakening the antitumor ability of CD8+
T cells (47). In addition, CD39 may
be a functional surface marker for identifying exhausted
CD8+ T cell subsets and regulating purine signalling
pathways, and the combined blockade of the Tim-3 and PD-1
signalling pathways reverses the exhaustion of T cells that is
induced by rectal cancer (48). The
present study revealed that CDK9 expression was positively
correlated with NFATC1, ENTPD1 (also referred to as
CD39) and NFAT5 expression, suggesting that CDK9 may
be involved in the ‘cancer immunoediting’ of T cells by the TME.
Furthermore, CDK9 promoted CD39 and Tim-3 expression in
tumor-infiltrating CD8+ T cells in MSS CRC tumors,
whereas CDK9 did not significantly affect PD-1 expression.
To the best of our knowledge, the present study was
the first to evaluate the association of CDK9 and
tumor-infiltrating CD8+ T cells in MSS CRC. A number of
studies have demonstrated that CDK9/P-TEFb is involved in the cell
growth and survival of several types of cancer, including CRC
(15,49,50).
However, these studies have focused on the cancer cells themselves,
ignoring the effect of CDK9 on the TME. The present study explored
the association between CDK9 expression and CD8+ T cells
in the TME of MSS CRC. Our previous study demonstrated that
PHA767491, a selective CDK9 inhibitor, could impair the activation
and proliferation of effector T cells but preserve the function of
Treg cells in a mouse inflammatory model (51). To the best of our knowledge, no study
has reported that a CDK9 inhibitor could alter anti-inflammatory
molecules or T cells in CRC. The present study focused on the
immune microenvironment of MSS CRC. In future studies, the effect
of CDK9 inhibitors on tumor-infiltrating lymphocytes in CRC will be
explored.
In conclusion, the present study demonstrated that
CDK9 expression was positively associated with poor survival among
patients with colon cancer and was negatively associated with
CD8+ T cell antitumor function in MSS mCRC. These
findings suggested that CDK9 may be utilized to evaluate the
prognosis and the immune-type of the TME in patients with MSS
mCRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from The
Science and Technology Development Fund of Tianjin Education
Commission for Higher Education (grant no. 2017KJ204) and
Scientific Research Foundation of Tianjin Medical University Cancer
Institute and Hospital (grant no. B1704), awarded to YZ.
Availability of data and materials
The datasets generated and analyzed during the
current study are available in the GEPIA repository, http://gepia.cancer-pku.cn/detail.php?gene=CDK9.
Authors' contributions
YZ and JL performed the majority of the experiments.
FT collected clinical specimens. JW performed data analysis and
interpretation. YZ and DK designed and supervised the study. DK
wrote the manuscript. YZ revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Patient samples were collected at the Tianjin
Medical University Cancer Institute and Hospital, and ethical
approval was granted by the Ethics Committee of Tianjin Medical
University Cancer Institute and Hospital. Written informed consent
was obtained from all patients for participation in the present
study.
Patient consent for publication
Written consent was obtained from all participants
for the publication of data.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
mCRC
|
metastatic colorectal cancer
|
|
CDK9
|
cyclin-dependent kinase 9
|
|
ASCO
|
American Society of Clinical
Oncology
|
|
CFDA
|
China Food and Drug Administration
|
|
MSI-H
|
microsatellite instability-high
|
|
MSS
|
microsatellite stable
|
|
dMMR
|
mismatch repair deficient
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, DeSantis C, Virgo K, Stein K,
Mariotto A, Smith T, Cooper D, Gansler T, Lerro C, Fedewa S, et al:
Cancer treatment and survivorship statistics, 2012. CA Cancer J
Clin. 62:220–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kibble A, Al-Shamahi A, Kuennemann K,
Marqués F, Tremosa L and Cole P: Highlights from the 52nd Annual
Meeting of the American Society of Clinical Oncology (ASCO) (June
3–7, 2016-Chicago, Illinois, USA). Drugs Today (Barc). 52:407–423.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xie S, Huang J, Qiao Q, Zang W, Hong S,
Tan H, Dong C, Yang Z and Ni L: Expression of the inhibitory B7
family molecule VISTA in human colorectal carcinoma tumors. Cancer
Immunol Immunother. 67:1685–1694. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu J, Chen L, Zou L, Yang P, Wu R, Mao Y,
Zhou H, Li R, Wang K, Wang W, et al: MiR-20b, −21, and −130b
inhibit PTEN expression resulting in B7-H1 over-expression in
advanced colorectal cancer. Hum Immunol. 75:348–353. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Masugi Y, Nishihara R, Yang J, Mima K, da
Silva A, Shi Y, Inamura K, Cao Y, Song M, Nowak JA, et al: Tumour
CD274 (PD-L1) expression and T cells in colorectal cancer. Gut.
66:1463–1473. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Koopman M, Kortman GA, Mekenkamp L,
Ligtenberg MJ, Hoogerbrugge N, Antonini NF, Punt CJ and van Krieken
JH: Deficient mismatch repair system in patients with sporadic
advanced colorectal cancer. Br J Cancer. 100:266–273. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Venderbosch S, Nagtegaal ID, Maughan TS,
Smith CG, Cheadle JP, Fisher D, Kaplan R, Quirke P, Seymour MT,
Richman SD, et al: Mismatch repair status and BRAF mutation status
in metastatic colorectal cancer patients: A pooled analysis of the
CAIRO, CAIRO2, COIN, and FOCUS studies. Clin Cancer Res.
20:5322–5330. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Le DT, Uram JN, Wang H, Bartlett BR,
Kemberling H, Eyring AD, Skora AD, Luber BS, Azad NS, Laheru D, et
al: PD-1 blockade in tumors with mismatch-repair deficiency. N Engl
J Med. 372:2509–2520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen DS and Mellman I: Elements of cancer
immunity and the cancer-immune set point. Nature. 541:321–330.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Smedt L, Lemahieu J, Palmans S, Govaere
O, Tousseyn T, Van Cutsem E, Prenen H, Tejpar S, Spaepen M,
Matthijs G, et al: Microsatellite instable vs. stable colon
carcinomas: Analysis of tumor heterogeneity, inflammation and
angiogenesis. Br J Cancer. 113:500–509. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Llosa NJ, Cruise M, Tam A, Wicks EC,
Hechenbleikner EM, Taube JM, Blosser RL, Fan H, Wang H, Luber BS,
et al: The vigorous immune microenvironment of microsatellite
instable colon cancer is balanced by multiple counter-inhibitory
checkpoints. Cancer Discov. 5:43–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Beauchamp EM, Abedin SM, Radecki SG,
Fischietti M, Arslan AD, Blyth GT, Yang A, Lantz C, Nelson A, Goo
YA, et al: Identification and targeting of novel CDK9 complexes in
acute myeloid leukemia. Blood. 133:1171–1185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma H, Seebacher NA, Hornicek FJ and Duan
Z: Cyclin-dependent kinase 9 (CDK9) is a novel prognostic marker
and therapeutic target in osteosarcoma. EBioMedicine. 39:182–193.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rahaman MH, Lam F, Zhong L, Teo T, Adams
J, Yu M, Milne RW, Pepper C, Lokman NA, Ricciardelli C, et al:
Targeting CDK9 for treatment of colorectal cancerMol Oncol;
2019
|
|
16
|
Chaiyapan W, Duangpakdee P,
Boonpipattanapong T, Kanngern S and Sangkhathat S: Somatic
mutations of K-ras and BRAF in Thai colorectal cancer and their
prognostic value. Asian Pac J Cancer Prev. 14:329–332. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Novellasdemunt L, Antas P and Li VS:
Targeting Wnt signaling in colorectal cancer. A Review in the
Theme: Cell Signaling: Proteins, Pathways and Mechanisms. Am J
Physiol Cell Physiol. 309:C511–C521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fleming NI, Jorissen RN, Mouradov D,
Christie M, Sakthianandeswaren A, Palmieri M, Day F, Li S, Tsui C,
Lipton L, et al: SMAD2, SMAD3 and SMAD4 mutations in colorectal
cancer. Cancer Res. 73:725–735. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Choi SH, Estarás C, Moresco JJ, Yates JR
III and Jones KA: α-Catenin interacts with APC to regulate
β-catenin proteolysis and transcriptional repression of Wnt target
genes. Genes Dev. 27:2473–2488. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alarcón C, Zaromytidou AI, Xi Q, Gao S, Yu
J, Fujisawa S, Barlas A, Miller AN, Manova-Todorova K, Macias MJ,
et al: Nuclear CDKs drive Smad transcriptional activation and
turnover in BMP and TGF-beta pathways. Cell. 139:757–769. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
De Falco G, Leucci E, Onnis A, Bellan C,
Tigli C, Wirths S, Cerino G, Cocco M, Crupi D, De Luca A, et al:
Cdk9/Cyclin T1 complex: A key player during the
activation/differentiation process of normal lymphoid B cells. J
Cell Physiol. 215:276–282. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kanazawa S, Okamoto T and Peterlin BM: Tat
competes with CIITA for the binding to P-TEFb and blocks the
expression of MHC class II genes in HIV infection. Immunity.
12:61–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marshall RM, Salerno D, Garriga J and
Graña X: Cyclin T1 expression is regulated by multiple signaling
pathways and mechanisms during activation of human peripheral blood
lymphocytes. J Immunol. 175:6402–6411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li T, Fan J, Wang B, Traugh N, Chen Q, Liu
JS, Li B and Liu XS: TIMER: A web server for comprehensive analysis
of tumor-infiltrating immune cells. Cancer Res. 77:e108–e110. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Szklarczyk D, Morris JH, Cook H, Kuhn M,
Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, et al:
The STRING database in 2017: Quality-controlled protein-protein
association networks, made broadly accessible. Nucleic Acids Res.
45:D362–D368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gupta S, Provenzale D, Regenbogen SE,
Hampel H, Slavin TP, Hall MJ, Llor X, Chung DC, Ahnen DJ, Bray T,
et al: NCCN guidelines insights: Genetic/Familial high-risk
assessment: Colorectal, version 3.2017. J Natl Compr Canc Netw.
15:1465–1475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu H, Krasinskas A and Willis J:
Perspectives on current tumor-node-metastasis (TNM) staging of
cancers of the colon and rectum. Semin Oncol. 38:500–510. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee YH, Martin-Orozco N, Zheng P, Li J,
Zhang P, Tan H, Park HJ, Jeong M, Chang SH, Kim BS, et al:
Inhibition of the B7-H3 immune checkpoint limits tumor growth by
enhancing cytotoxic lymphocyte function. Cell Res. 27:1034–1045.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park JH, Kim MJ, Park SC, Kim MJ, Hong CW,
Sohn DK, Han KS and Oh JH: Difference in time to locoregional
recurrence between patients with right-sided and left-sided colon
cancers. Dis Colon Rectum. 58:831–837. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Meguid RA, Slidell MB, Wolfgang CL, Chang
DC and Ahuja N: Is there a difference in survival between right-
versus left-sided colon cancers? Ann Surg Oncol. 15:2388–2394.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Feig C, Jones JO, Kraman M, Wells RJ,
Deonarine A, Chan DS, Connell CM, Roberts EW, Zhao Q, Caballero OL,
et al: Targeting CXCL12 from FAP-expressing carcinoma-associated
fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic
cancer. Proc Natl Acad Sci USA. 110:20212–20217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zboralski D, Hoehlig K, Eulberg D,
Frömming A and Vater A: Increasing tumor-infiltrating T cells
through inhibition of CXCL12 with NOX-A12 synergizes with PD-1
blockade. Cancer Immunol Res. 5:950–956. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Minzel W, Venkatachalam A, Fink A, Hung E,
Brachya G, Burstain I, Shaham M, Rivlin A, Omer I, Zinger A, et al:
Small molecules Co-targeting CKIα and the transcriptional kinases
CDK7/9 control AML in preclinical models. Cell.
175:171.e25–185.e25. 2018. View Article : Google Scholar
|
|
35
|
Rahaman MH, Yu Y, Zhong L, Adams J, Lam F,
Li P, Noll B, Milne R, Peng J and Wang S: CDKI-73: An orally
bioavailable and highly efficacious CDK9 inhibitor against acute
myeloid leukemia. Invest New Drugs. 37:625–635. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Morales F and Giordano A: Overview of CDK9
as a target in cancer research. Cell Cycle. 15:519–527. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin T, Lallena MJ, Kreklau EL, Fales KR,
Carballares S, Torrres R, Wishart GN, Ajamie RT, Cronier DM,
Iversen PW, et al: A novel CDK9 inhibitor shows potent antitumor
efficacy in preclinical hematologic tumor models. Mol Cancer Ther.
13:1442–1456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abdullah C, Wang X and Becker D:
Expression analysis and molecular targeting of cyclin-dependent
kinases in advanced melanoma. Cell Cycle. 10:977–988. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Narita T, Ishida T, Ito A, Masaki A,
Kinoshita S, Suzuki S, Takino H, Yoshida T, Ri M, Kusumoto S, et
al: Cyclin-dependent kinase 9 is a novel specific molecular target
in adult T-cell leukemia/lymphoma. Blood. 130:1114–1124. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yik JH, Hu Z, Kumari R, Christiansen BA
and Haudenschild DR: Cyclin-dependent kinase 9 inhibition protects
cartilage from the catabolic effects of proinflammatory cytokines.
Arthritis Rheumatol. 66:1537–1546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sleightholm RL, Neilsen BK, Li J, Steele
MM, Singh RK, Hollingsworth MA and Oupicky D: Emerging roles of the
CXCL12/CXCR4 axis in pancreatic cancer progression and therapy.
Pharmacol Ther. 179:158–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song ZY, Wang F, Cui SX, Gao ZH and Qu XJ:
CXCR7/CXCR4 heterodimer-induced histone demethylation: A new
mechanism of colorectal tumorigenesis. Oncogene. 38:1560–1575.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu L, Zhao L, Yang Y, Gao J, Hu C, Guo B
and Zhu B: Cytotoxic chemotherapy reduces T cell trafficking to the
spleen by downregulating the expression of C-C motif chemokine
ligand 21 and C-C motif chemokine ligand 19. Oncol Lett.
16:5013–5019. 2018.PubMed/NCBI
|
|
45
|
Kim PS and Ahmed R: Features of responding
T cells in cancer and chronic infection. Curr Opin Immunol.
22:223–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Baitsch L, Fuertes-Marraco SA, Legat A,
Meyer C and Speiser DE: The three main stumbling blocks for
anticancer T cells. Trends Immunol. 33:364–372. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Martinez GJ, Pereira RM, Äijö T, Kim EY,
Marangoni F, Pipkin ME, Togher S, Heissmeyer V, Zhang YC, Crotty S,
et al: The transcription factor NFAT promotes exhaustion of
activated CD8+ T cells. Immunity. 42:265–278. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu J, Zhang S, Hu Y, Yang Z, Li J, Liu X,
Deng L, Wang Y, Zhang X, Jiang T and Lu X: Targeting PD-1 and Tim-3
pathways to reverse CD8 T-cell exhaustion and enhance ex
vivo T-cell responses to autologous dendritic/tumor vaccines. J
Immunother. 39:171–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Franco LC, Morales F, Boffo S and Giordano
A: CDK9: A key player in cancer and other diseases. J Cell Biochem.
119:1273–1284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Boquoi A, Chen T and Enders GH:
Chemoprevention of mouse intestinal tumorigenesis by the
cyclin-dependent kinase inhibitor SNS-032. Cancer Prev Res (Phila).
2:800–806. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhan Y, Han Y, Sun H, Liang T, Zhang C,
Song J and Hou G: Down-regulating cyclin-dependent kinase 9 of
alloreactive CD4+ T cells prolongs allograft survival. Oncotarget.
7:24983–24994. 2016. View Article : Google Scholar : PubMed/NCBI
|