Introduction
Leukemia is a type of hematological disease
characterized by the malignant proliferation of leukocytes. It is
mainly divided into chronic lymphocytic leukemia, chronic myeloid
leukemia (CML), acute lymphoblastic leukemia and acute myeloid
leukemia (1). The treatment of
leukemia has always been based on chemotherapy; however, multidrug
resistance (MDR) that occurs during chemotherapy has seriously
impeded the efficacy of leukemia treatment (2,3). Based
on extensive research, the mechanisms of the MDR of leukemia are
mainly divided into the following categories: i) Overexpression of
transporters; ii) evasion of apoptosis-associated signaling
pathways; and iii) changes in the intracellular microenvironment,
all of which lead to an increase in the tolerance of cells to
chemotherapeutic drugs (2,4). The MDR phenomenon of leukemia cells is
widespread, and its mechanisms are diverse (5). Therefore, seeking reversal strategies
or reversal agents to overcome MDR has become a research hotspot
(6). Chemical reversal agents
usually have serious toxicity and side effects; for instance,
verapamil (VRP) and cyclosporin A, which target p-glycoprotein
(P-gp), although producing good reversal effects in vitro,
cannot be applied in a clinical setting due to their serious toxic
side effects. Furthermore, they only have a single mechanism of
action and poor selectivity (7).
Traditional Chinese medicine has a long history and
abundant resources, and it provides a large number of drugs
targeting multiple factors and pathways (8). An increasing amount of research has
been conducted regarding potential MDR reversal agents for leukemia
(9). Curcumin (Fig. 1A) is a polyphenolic compound
extracted from the rhizome of Zingiberaceae and
Araceae, and has numerous biological effects with
pharmaceutical applications, including analgesic, antioxidant,
anti-inflammatory, antiseptic, antirheumatic and
antiatherosclerotic actions. Due to its multiple actions and
multi-targeting characteristics, curcumin has attracted widespread
attention (10). Previous studies
have indicated curcumin's antitumor activity (11,12). An
early study indicated that curcumin modulates the function and
expression of P-gp in vitro, potentially sensitizing
P-gp-overexpressing cell lines to chemotherapeutics (13). A large number of subsequent studies
have also supported this hypothesis (14,15);
however, the majority of studies on curcumin have primarily focused
on its effects on the function and expression of P-gp (16). Due to its multiple targets, curcumin
may possess further molecular mechanisms that are worth examining
within the context of MDR.
The S100 proteins are a family of low molecular
weight (9–13 kDa) calcium-binding proteins featuring an EF-hand
structure with 21 members, and are widely distributed in various
types of tissue, such as brain, heart, kidneys and skin, and is
highly expressed in the brain and heart (17,18).
Upon binding to calcium ions, the conformation of the protein
changes, exposing its binding site to the target protein, and
thereby exerting its biological effects via the corresponding
target protein (19). Therefore,
S100 protein is considered to be a calcium sensor protein, which
has an important role in cell proliferation, differentiation,
muscle contraction, gene expression, secretion and apoptosis
through the calcium signal transduction pathway. S100
calcium-binding protein A8 (S100A8), also known as myeloid-related
protein 8 or calgranulin A, is a member of the S100 multigene
subfamilies. Studies have indicated that S100A8 is associated with
the progression of multiple tumor types and mediates apoptosis
through the B-cell lymphoma 2 (Bcl-2) family of proteins (20,21).
Recent studies have also shown that S100A8 is associated with drug
resistance (22,23). The present study examined two
possible targets, P-gp and S100A8, to elucidate the mechanisms via
which curcumin reverses the drug resistance of doxorubicin
(DOX)-resistant K562 (K562/DOX) cells. The study aimed to provide a
sufficient basis for the clinical application of curcumin to
improve the efficacy of chemotherapy for drug-resistant
leukemia.
Materials and methods
Cell culture
The human CML cell lines K562 and K562/DOX were
obtained from Nanjing KeyGen Biotech Co., Ltd. Cells were cultured
in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 100 U/ml penicillin/streptomycin and 10% fetal
calf serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. and passaged every 2–3 days to maintain logarithmic
growth. In order to maintain DOX resistance, K562/DOX cells were
cultured in medium with 2 mg/ml DOX (Sigma-Aldrich; Merck KGaA).
K562/DOX cells were grown in DOX-free culture medium for >2
weeks prior to the assays.
MTT assay
K562 or K562/DOX cells were seeded in 96-well plates
at a density of 5,000 cells/well. After 24 h, cells were incubated
with various drugs. The concentration of DOX was 0, 0.8, 1.6, 3.13,
6.25, 12.5, 25, 50 µM and the concentration of curcumin used was 0,
0.5, 1, 2, 4, 8, 16, 32 µM. The MTT assay was then performed
according to the manufacturer's protocol (BioVision, Inc.). The
absorbance was measured at a wavelength of 570 nm using a
microplate reader (Bio-Rad Laboratories, Inc.). The IC50
value was calculated with SPSS 23.0 (IBM Corp.). Resistance Index
was calculated by dividing the IC50 values of K562/DOX
cells in the presence or absence of curcumin by the IC50
values of K562 cells without curcumin. Reversal Index was
calculated by dividing the values of Resistance Index without
curcumin by the Resistance Index in the presence or absence of
curcumin.
Western blot analysis
A total of 1×106 K562 or K562/DOX cells
were collected, washed thrice with ice-cold PBS and lysed with
ice-cold RIPA lysis buffer (Beyotime Institute of Biotechnology).
The protein content was detected using the BCA protein
concentration determination kit (cat. no. PC0020; Beijing Solarbio
Science & Technology Co., Ltd.). A total of 30 µg protein was
loaded per lane and subjected to SDS-PAGE (upper gel 5%, separation
gel 10–15%). The nitrocellulose membranes were blocked by 5% BSA
blocking buffer for 1 h at room temperature and incubated with
primary antibodies overnight at 4°C after washing three times with
0.1% Tween-20 in Tris-buffered saline; membranes were then
incubated with alkaline phosphatase-conjugated anti-rabbit (cat.
no. 7054; 1:5,000 dilution) or mouse (cat. no. 7056; 1:5,000
dilution) antibodies (Cell Signaling Technology, Inc.) for 1 h at
room temperature, and then developed with enhanced
chemiluminescence reagent (Beyotime Institute of Biotechnology).
Band density was quantified using ImageJ software 1.46r (National
Institutes of Health). Antibodies against P-gp (cat. no. 13342;
1:1,500 dilution) and CCAAT/enhancer-binding protein homologous
protein (CHOP; cat. no. 2895; 1:2,000 dilution) were obtained from
Cell Signaling Technology, Inc.; anti-S100A8 antibody (cat. no.
15792-1-AP; 1:1,500 dilution) was obtained from ProteinTech Group,
Inc., whereas antibodies specific for Bcl extra-large (Bcl-xL; cat.
no. bs-5234R; 1:1,000 dilution), Bcl-2-associated X protein (Bax;
cat. no. bs-0127R; 1:1,000 dilution) and β-actin (cat. no.
bs-0061R; 1:1,000 dilution) were purchased from Bioss
Antibodies.
Determination of intracellular
rhodamine (RHO)-123 retention
Intracellular retention of RHO-123 (Beyotime
Institute of Biotechnology) was detected in order to determine P-gp
efflux function (24). K562 or
K562/DOX Cells were seeded in 6-well plates at 5×105
cells/well. The cells were pre-treated with the indicated drugs
(VRP 10 µM; curcumin 0.5,1 and 2µM) for 48 h at 37°C with 5%
CO2. and subsequently VRP(10 µM) was used as a positive
control. After pre-treatment, the cells were incubated with 10
µg/ml RHO-123 in RPMI-1640 medium at 37°C for 45 min. The cells
were then collected, washed with ice-cold PBS twice and suspended
in 0.5 ml RPMI-1640 medium. The mean fluorescence intensity of the
samples was determined via flow cytometric analysis (FACSCalibur™;
BD Biosciences) using a 530-nm long band-pass filter. The data of
flow cytometric was analyzed by FlowJo X 10.0.7r2 (FlowJo,
LLC).
Intracellular accumulation of DOX
The intracellular accumulation of DOX was examined
by measuring the fluorescence intensity of DOX. The K562 or
K562/DOX cells were seeded in 6-well plates at 5×105
cells/well. After 24 h, the cells were pre-treated for 48 h at 37°C
with 5% CO2. and then 2 ml DOX (4 µM) was added to each
well, followed by incubation at 37°C for 60 min in the dark.
Following incubation, the cells were immediately washed with
ice-cold PBS three times and lysed with RIPA lysis buffer, and the
fluorescent signal of the cells was detected with a multimode
microplate reader (TriStar2; Berthold Bio) using a 595-nm long
band-pass filter.
Apoptosis detection assay
Caspase-3 activity was detected with a caspase-3
activity assay kit (cat. no. BC3830; Beijing Solarbio Science &
Technology Co., Ltd.) (25). In
brief, 5×106 pre-treated K562 or K562/DOX cells were
collected and lysed. After centrifugation at 4°C at 12,000 × g for
10 min, the supernatant was collected and according to the
manufacturer's protocol, the corresponding reagent was added. The
absorbance was measured at a wavelength of 405 nm using a
microplate reader (Bio-Rad Laboratories, Inc.). The caspase-3
activity was quantified as the absorbance of the drug-treated cells
relative to that of the vehicle-treated control cells.
The stage of apoptosis was analyzed by staining with
DAPI (cat. no. C1002; Beyotime Institute of Biotechnology) as
described previously (26). DAPI can
be used for the detection of apoptosis, because it is a blue
fluorescent dye that can penetrate cell membranes. Binding to
double-stranded DNA produces a >20-fold increase in fluorescence
compared with DAPI itself (27).
According to the recommended DAPI concentration of 0.5–10 µg/ml, a
final concentration of 5 µg/ml was selected (28). The cells were seeded in 6-well plates
at 5×105 cells/well and subjected to the designated
treatments for 48 h, and subsequently, 0.5 ml DAPI at a final
concentration of 5 µg/ml was added to each well, followed by
incubation at 37°C for 30 min in the dark. The cells were then
immediately washed with ice-cold PBS and observed under an inverted
fluorescence microscope (magnification, ×100; OIS IX81; Olympus
Corporation) using a 454-nm long band-pass filter.
Determination of the intracellular
concentration of Ca2+
For calcium determination, 5×105 K562 or
K562/DOX cells that had been subjected to the aforementioned
treatments were collected and lysed with lysis buffer. A Calcium
Colorimetric Assay kit (cat. no. MAK022; Sigma-Aldrich; Merck KGaA)
was used to analyze the intracellular calcium ion concentration,
according to the manufacturer's protocol. The absorbance was
measured at a wavelength of 575 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
RNA knockdown
Small interfering (si)RNA oligos for P-gp and
S100A8, and no-targeting si-RNA controls (ctrl) for S100A8 and P-gp
were synthesized by General Biosystems, Inc. For knockdown
experiments, siRNA oligos were diluted to a concentration of 50 nM
and transfected into the cells with Lipofectamine™ 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The sequences for P-gp siRNA and sictrl
P-gp were as follows: P-gp siRNA, forward,
5′-CGGAAGGCCUAAUGCCGAA-3′ and reverse, 5′-UUCGGCAUUAGGCCUUCCG-3′;
sictrl P-gp, forward, 5′-UGUAGAUGGACUUGAACUC-3′, and reverse,
5′-GAGUUCAAGUCCAUCUACA3′ (29). The
following sequences were used for S100A8 siRNA and sictrl S100A8:
S100A8 siRNA, forward, 5′-AGACCGAGUGUCCUCAGUA-3′ and reverse,
5′-UACUGAGGACACUCGGUCU-3′; sictrl S100A8, forward,
5′-GCGACGAUCUGCCUAAGAU-3′ and reverse, 5′-AUCUUAGGCAGAUCGUCGC-3′
(30). The K562/DOX cells were
treated with different concentrations of drugs. After 48 h the
non-transfection group cells were treated with 4 µM DOX, 2 µM
curcumin alone, or in combination for 48 h, and the transfection
group cells were treated with 4 µM DOX for 48 h.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Statistical significance between groups was evaluated
with a two-tailed Student's t-test using GraphPad Prism 7 (GraphPad
Software, Inc.) and SPSS 23.0 (IBM Corp.). Multiple comparison
between the groups was performed using one-way analysis of variance
with Student-Newman-Keuls method post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Curcumin sensitizes K562/DOX cells to
DOX
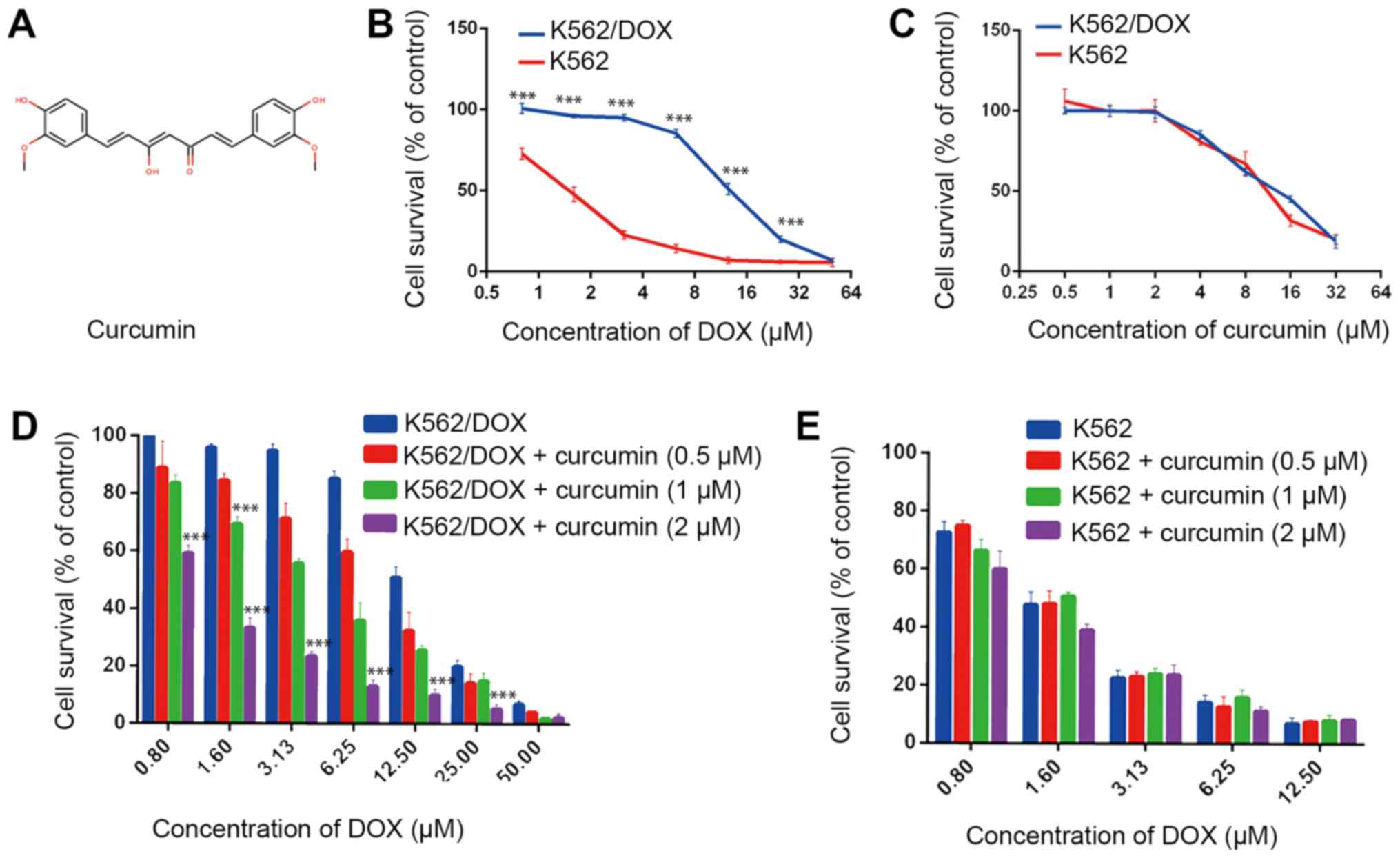
To investigate the sensitization effect of curcumin,
an MTT cytotoxicity assay was performed. First the results
indicated that the resistant K562/DOX cell line exhibited
significant drug resistance to DOX compared with the native K562
cell line, with a resistance index up to 10 (Fig. 1B; Table
I). Furthermore, curcumin at 0.5–2 µM was non-toxic to K562 and
K562/DOX cells (Fig. 1C; Table I). To determine whether the decrease
in the survival rate of K562/DOX and K562 cells was due to the
toxicity of curcumin, a non-toxic concentration of curcumin (0.5, 1
and 2 µM) was used in combination with DOX. Of note, curcumin
significantly enhanced the sensitivity of K562/DOX and cells to
DOX. Curcumin reduced the survival rate of K562/DOX at different
concentrations of DOX in a dosage-depended manner; the reversal
indices of 0.5, 1 and 2 µM curcumin were 1.3, 2.43 and 9.3,
respectively (Fig. 1D; Table I). However, for the K562 cells,
curcumin did not significantly affect the decrease in the cell
survival rate caused by DOX (Fig.
1E; Table I). Therefore, it may
be concluded that curcumin at non-toxic concentrations reverses the
drug resistance of K562/DOX cells.
 | Table I.IC50 values and reversal
effect of curcumin on DOX in K562 and K562/DOX cells. |
Table I.
IC50 values and reversal
effect of curcumin on DOX in K562 and K562/DOX cells.
|
|
IC50±standard
deviationa (µM) |
|
|
|---|
|
|
|
|
|
|---|
| Drugs | K562 | K562/DOX | Reversal index | Resistance
index |
|---|
| DOX | 1.38±0.22 |
13.83±4.43c | 10 | 1 |
|
Curcuminb | 2.3±0.17
(IC10) | 2.5±0.24
(IC10) | / | / |
| DOX + curcumin (0.5
µM) | 1.4±0.28 |
10.91±2.84c | 7.9 | 1.3 |
| DOX + curcumin (1
µM) | 1.373±0.18 |
5.72±0.83c | 4.1 | 2.43 |
| DOX + curcumin (2
µM) | 1.052+0.24 |
1.48±0.13c | 1.07 | 9.3 |
S100A8 and P-gp are involved in the
reversal effects of curcumin on the drug resistance of K562/DOX
cells
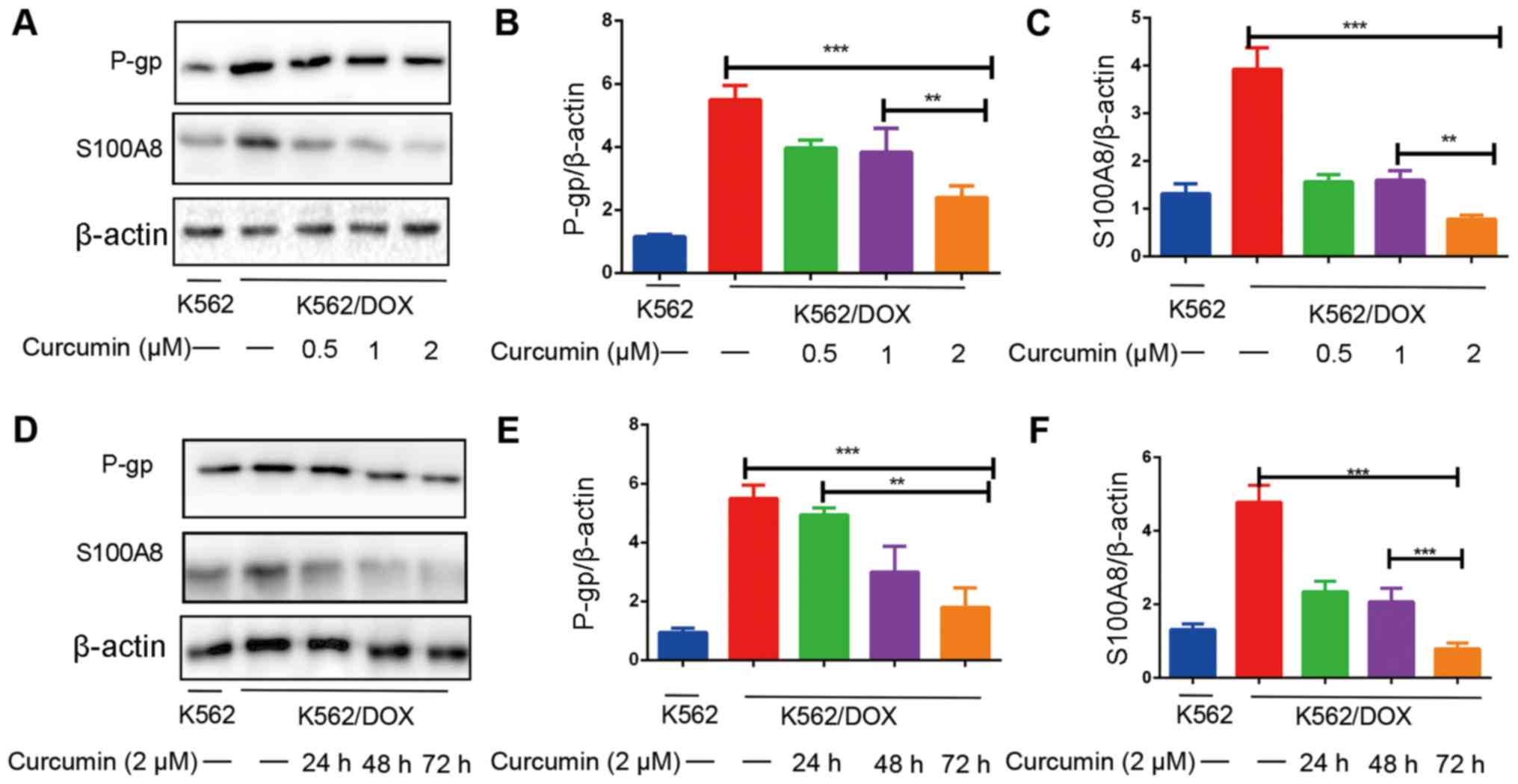
To investigate the expression of S100A8 and P-gp in
K562 and K562/DOX cells, a western blot assay was performed. The
results indicated that the expression of S100A8 and P-gp in
K562/DOX cells was significantly increased compared with in K562
cells (Fig. 2A and D). The effects
of different concentrations of curcumin on the expression of S100A8
and P-gp were then assessed. The results suggested that when
curcumin was administered at 0.5, 1 and 2 µM for 48 h, the
expression levels of S100A8 and P-gp were significantly reduced
(Fig. 2A-C). Next, the
time-dependence of the effect of curcumin on the downregulation of
S100A8 and P-gp in K562/DOX cells was assessed. The results
indicated that the expression of P-gp decreased with prolonged
incubation with curcumin. The expression of S100A8 following 24 and
48 h of curcumin treatment was similar, however it was still
downregulated compared with that in the control group, and at 72 h,
the downregulation effect was most notable (Fig. 2D-F). These results indicated that the
dose- and time-dependent downregulation of S100A8 and P-gp
expression by curcumin may be associated with its reversal effect
on drug resistance. The downregulated expression of P-gp and S100A8
was evident up to the 48-h time point; therefore, the shortest time
that induced changes in protein expression, 48-h, was selected for
the duration of pre-treatment for subsequent experiments.
Curcumin inhibits the expression and
function of P-gp, and increases the intracellular accumulation of
DOX
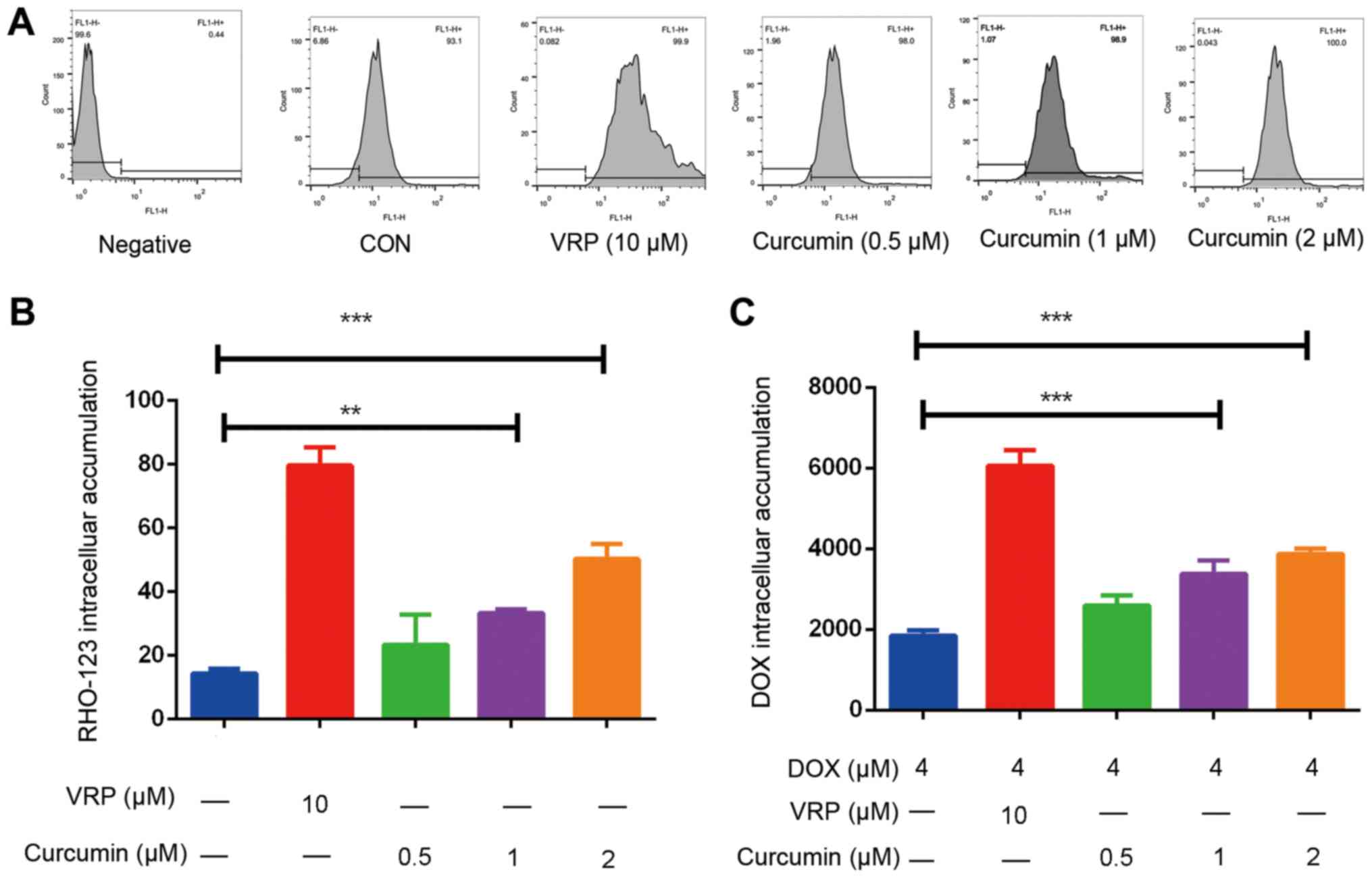
The results in Fig. 2
indicated that curcumin treatment causes a decrease in the
expression levels of P-gp; it was further investigated whether it
inhibits the efflux function of P-gp. To determine this, a RHO-123
assay was performed. The results suggested that curcumin increased
the intracellular accumulation of RHO-123 in a dose-dependent
manner (Fig. 3A and B). This
indicates that the efflux function of P-gp was inhibited when
curcumin was administered. In addition, the intracellular
accumulation of DOX was assayed; it was revealed that curcumin
dose-dependently increased the intracellular accumulation of DOX
(Fig. 3C). DOX is a substrate for
P-gp (31); however, when P-gp is
overexpressed, intracellular DOX is transported to the outside of
the cell via P-gp, and the resulting intracellular drug
concentration becomes too low to induce cancer cell death, thereby
producing resistance (32). It is
proposed that as curcumin inhibits the expression and function of
P-gp, the intracellular drug concentration increases and the drug
resistance effect is reversed.
S100A8 and P-gp are involved in the
curcumin-induced drug sensitization of K526/DOX cells independent
of each other
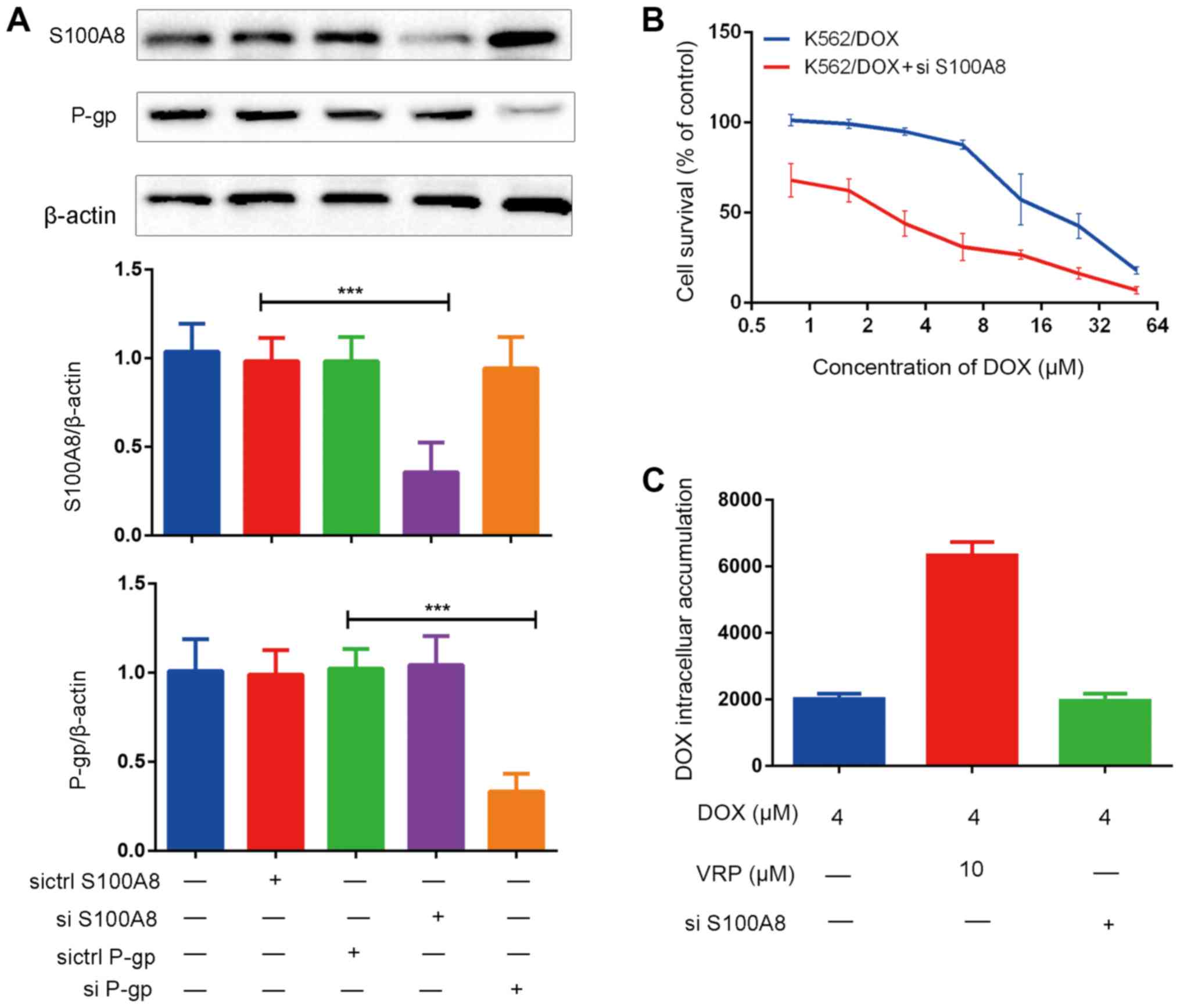
To elucidate the association between S100A8 and
P-gp, gene silencing technology was used. The results indicated
that when S100A8 was silenced, the levels of P-gp were not
affected, and in turn, gene silencing of P-gp did not affect the
expression of S100A8 (Fig. 4A). This
result indicated that S100A8 and P-gp mediated curcumin-induced
reversal of K526/DOX cell resistance to DOX independent of each
other.
In order to further investigate the role of S100A8
in the curcumin-induced reversal of drug resistance, the expression
of S100A8 in K562/DOX cells was inhibited by gene silencing, and
the sensitivity of K562/DOX to DOX was assessed via a cytotoxicity
assay. The results indicated that at the same DOX concentration,
the cell survival rate was significantly reduced after silencing
S100A8 (Fig. 4B). In addition, the
effects of S100A8-knockdown on the intracellular accumulation of
DOX were investigated. Of note, after gene silencing of S100A8, the
intracellular accumulation of DOX did not significantly increase
(Fig. 4C). These results suggest
that the role of S100A8 in the curcumin-induced reversal of
K562/DOX cell resistance to DOX is not associated with increased
intracellular accumulation of DOX.
Curcumin inhibits the expression of
S100A8 and increases apoptosis induced by endoplasmic reticulum
stress (ERS)
S100A8 is a calcium ion-binding protein, and calcium
ions are involved in the transduction of various intracellular
signaling pathways, such as Ras/ERK/CREB pathway (33–35). The
ER is a calcium reservoir in the cell that is sensitive to changes
in the intracellular calcium levels (36). To clarify the association between
S100A8, calcium ions and the ER, intracellular free calcium ion
levels were first examined. The results indicated that the
intracellular calcium levels in K562/DOX and K562 cells were
equivalent. After administration of 4 µM DOX or 2 µM curcumin
alone, the intracellular calcium levels did not increase; however,
when DOX was combined with curcumin or S100A8 was silenced, the
intracellular calcium levels were significantly elevated (Fig. 5A). In addition, the expression of the
ERS-associated protein CHOP was increased (Fig. 5C). To investigate the effects on
apoptosis, the expression of the apoptosis-associated proteins
Bcl-xL and Bax, and the activity of the apoptosis executioner
protein caspase-3 were assessed. The results indicated that DOX
combined with curcumin or S100A8 silencing significantly decreased
the expression of antiapoptotic protein Bcl-xL, whereas the
expression of the proapoptotic protein Bax and the activity of
caspase-3 were significantly upregulated (Fig. 5B and D). In addition, the results of
a DAPI staining assay revealed that when DOX was combined with
curcumin or silencing of S100A8, an increase in nuclear
fragmentation was observed, indicative of the occurrence of
apoptosis (Fig. 5E). A possible
explanation for this observation is that due to the downregulation
of S100A8, the increase in intracellular calcium levels induced by
the chemotherapeutic drug cannot be cleared in time, leading to
apoptosis induced by ERS.
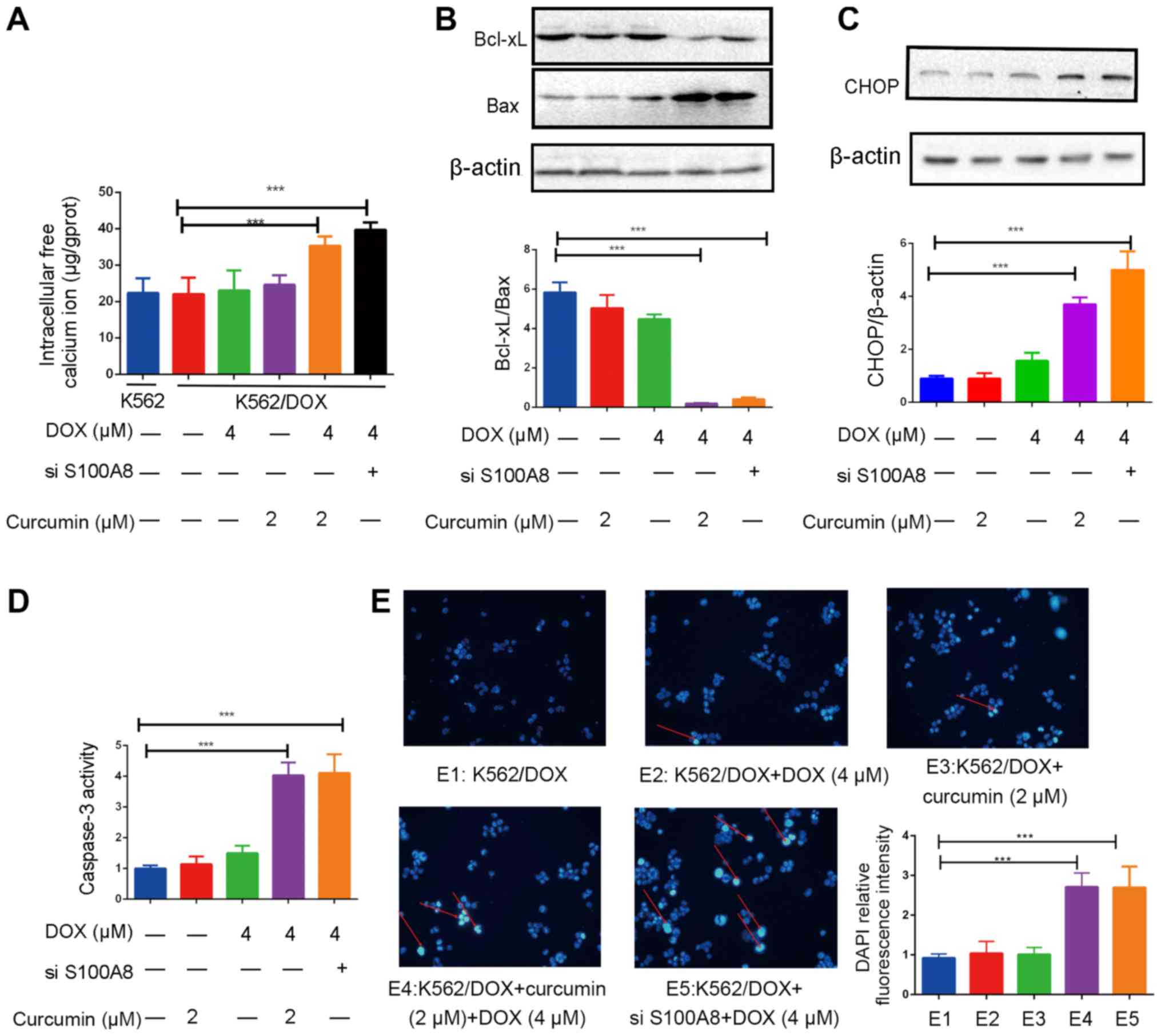 | Figure 5.Addition of curcumin enhances
apoptosis induced by DOX in K562/DOX cells. (A) Effects of curcumin
(2 µM), DOX (4 µM) and si-S100A8 used alone or in combination on
the intracellular free calcium ion concentration in K562/DOX cells.
Effects of curcumin (2 µM), DOX (4 µM) and si-S100A8 used alone or
in combination on the protein expression of (B) Bcl-xL, Bax and (C)
CHOP in K562/DOX cells. (D) Effects of curcumin (2 µM), DOX (4 µM)
and si-S100A8 used alone or in combination on caspase-3 activity.
(E) K562/DOX cells were stained with DAPI after pre-treatment and
observed under an inverted fluorescence microscope (magnification,
×40) and the relative fluorescence intensity analysis of DAPI was
determined. DNA fragmentation is indicated by red arrows.
***P<0.001. DOX, doxorubicin; K562/DOX, DOX-resistant K562 cell
line; S100A8, S100 calcium-binding protein A8; Bcl-xL, B-cell
lymphoma extra-large; Bax, Bcl-2-associated X protein; CHOP,
CCAAT/enhancer-binding protein homologous protein; si-S100A8, small
interfering RNA targeting S100A8. |
Discussion
The emergence of MDR has seriously impeded the
efficacy of chemotherapy for leukemia. Previous studies have
indicated that curcumin and its derivatives are capable of
reversing the MDR of various types of cancer, including non-small
cell lung cancer, liver cancer and colon cancer (37–39).
Although the ability of curcumin to reverse drug resistance in
solid tumors has been reported in a large number of studies, few
studies have examined the reversal of MDR of leukemia by curcumin,
and mechanistic investigations mostly focused on the inhibition of
P-gp expression (40–43). The aforementioned was confirmed by
this study's results. It is well known that P-gp is involved in the
development of drug resistance in most cell types, partly due the
specific physiological functions of P-gp, and the fact that P-gp
has a particularly broad substrate range (44–46).
However, it may not be feasible to reverse MDR by inhibiting P-gp
alone. The present study revealed that the reversal effects of
curcumin are not only associated with P-gp, but also with the
inhibition of S100A8 expression. To the best of our knowledge,
S100A8 has not been reported prior to this study as a target of
curcumin.
To investigate the underlying mechanisms, the
present study examined two proteins, P-gp and S100A8, and aimed to
identify an association between the two. First, the effects of
curcumin on the expression of S100A8 and P-gp were investigated.
The results suggested that curcumin decreased the expression of
P-gp and S100A8 in a time- and dose-dependent manner.
The ability of curcumin to reverse resistance and
downregulate S100A8 expression after administration was observed.
To demonstrate that S100A8 downregulation was associated with the
reverse of drug-resistance, S100A8 gene silencing was performed
using siRNAs inhibiting S100A8 expression. The results showed that
after inhibiting S100A8 expression, drug-resistant cells were more
sensitive to DOX, the expression and activity of proapoptotic
proteins was increased, and the ratio of Bcl-xl and Bax was
decreased. The results indicated that the alterations in S100A8
expression level were important for the reversal of drug
resistance. To identify the potential association between P-gp and
S100A8, the expression of each was separately inhibited by gene
silencing, with the expression of the other protein then evaluated.
The results indicated that the two proteins did not directly
regulate the expression of the alternate protein; therefore,
curcumin may act independently on them.
The molecular mechanisms via which curcumin acted on
P-gp and S100A8 individually were subsequently examined. K562/DOX
cells overexpress of P-gp and S100A8 compared with K562 cells.
Curcumin did not affect DOX sensitivity in the K562 model, likely
due to low expression of P-gp and S100A8. For P-gp, in order to
investigate the effect of curcumin on its transport function,
curcumin and RHO-123 were applied to K562/DOX cells simultaneously.
Following incubation for 45 min, the intracellular RHO-123 levels
were detected, with the P-gp-specific inhibitor VRP used as a
positive control (47,48). The results suggested that when
curcumin was applied to K562/DOX cells, the intracellular
accumulation of RHO-123 increased significantly, indicating that
curcumin inhibited the function of P-gp.
For the calcium ion-binding protein S100A8, the
results indicated that the downregulation of S100A8 by curcumin may
enhance ERS induced by DOX and thereby increase cell apoptosis.
Calcium ions have an important role in intracellular signal
transduction (49). They are able to
activate a variety of different protein kinases involved in various
biological processes, including cell proliferation,
differentiation, invasion and apoptosis (50,51). The
ER is the major site for intracellular protein synthesis,
secretion, modification and transport, and is also the major
organelle for the maintenance of the intracellular calcium balance
(52). Under physiological
conditions, the levels of free calcium ions in the cytoplasm are
much lower than those in the ER (53). The calcium ion pump moves calcium
ions from the cytosol into the ER, whereas the ER releases calcium
ions from the cytosol via ryanodine receptors and inositol
1,4,5-trisphosphate receptors to maintain the homeostasis of
intracellular calcium ions (54–56). An
increase in the levels of intracellular free calcium ions causes an
imbalance of intracellular calcium ion homeostasis that leads to
calcium overload in the ER, resulting in ERS and the induction of
apoptosis (54–56). In the present study, changes in
intracellular calcium in K562/DOX cells were examined. It was
revealed that, while 4 µM DOX did not significantly change
intracellular calcium levels, they were significantly increased
when DOX was combined with curcumin or S100A8 siRNA
transfection.
CHOP is a specific transcription factor for ERS.
When ERS induces apoptosis, the expression of CHOP is notably
increased and the protein accumulates in the nucleus, where it
induces apoptosis. In addition, CHOP also affects the expression of
Bcl-2 family proteins; specifically, it inhibits the expression of
the antiapoptotic Bcl protein and increases the sensitivity of
mitochondria to proapoptotic factors (57). The results of the present study
suggested that the combination of DOX and curcumin or silencing of
S100A8 causes an upregulation of CHOP expression, a downregulation
of the Bcl-xL/Bax ratio and an increase in caspase activity, which
is expected to be associated with marked ERS and cell apoptosis. In
addition, to evaluate cell death, nuclear staining with DAPI was
performed. It was revealed that in the DOX + curcumin and the DOX +
S100A8 siRNA groups, a significant increase in nuclear
fragmentation was present, suggesting that apoptosis occurred. It's
worth mentioning that when designing the assays, the drug to treat
K562 as a control was not designed by the present study. There were
two reasons for this: The concentration of DOX (4 µM) far exceeds
the IC50 (1.38 µM) value of K562 for DOX, which
inevitably causes a large number of cell deaths, and for K562/DOX
cells, the concentration of 4 µM is much lower than its
IC50 (13.83 µM) value, due to drug resistance (Fig. 1B; Table
I).
In conclusion, the present study indicated that
curcumin reversed the resistance of K562/DOX cells to DOX, and
S100A8 and P-gp were identified as two independent targets.
Curcumin was demonstrated to inhibit the expression and function of
P-gp, and to increase the intracellular accumulation of DOX.
Furthermore, curcumin downregulated the expression of S100A8,
causing an intracellular calcium ion imbalance that resulted in
increased ERS, thereby accelerating cell apoptosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LY completed the major assays such as RNA knockdown,
western blot analysis and flow cytometry. DL analyzed the data and
was responsible for the cell culture. LY and DL contributed to the
writing of this manuscript, while PT and YZ contributed the
conception and design of the study, and editing of the manuscript.
All authors have read and approved the final version of this
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DOX
|
doxorubicin
|
|
K562/DOX
|
DOX-resistant K562 cell line
|
|
S100A8
|
S100 calcium-binding protein A8
|
|
Bcl-xL
|
B-cell lymphoma extra-large
|
|
Bax
|
Bcl-2-associated X protein
|
|
CHOP
|
CCAAT/enhancer-binding protein
homologous protein
|
|
si-S100A8
|
small interfering RNA targeting
S100A8
|
References
|
1
|
Swerdlow S, Campo E, Harris N, Jaffe E,
Pileri S, Stein H, Thiele J and Vardiman J: WHO Classification of
Tumours of Haematopoietic and Lymphoid TissuesIARC; Lyon: 2008
|
|
2
|
Agrawal M, Hanfstein B, Erben P, Wolf D,
Ernst T, Fabarius A, Saussele S, Purkayastha D, Woodman RC, Hofmann
WK, et al: MDR1 expression predicts outcome of Ph+ chronic phase
CML patients on second-line nilotinib therapy after imatinib
failure. Leukemia. 28:1478–1485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Q, Feng Y and Kennedy D:
Multidrug-resistant cancer cells and cancer stem cells hijack
cellular systems to circumvent systemic therapies, can natural
products reverse this? Cell Mol Life Sci. 74:777–801. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xia CQ and Smith PG: Drug efflux
transporters and multidrug resistance in acute leukemia:
Therapeutic impact and novel approaches to mediation. Mol
Pharmacol. 82:1008–1021. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Briot T, Roger E, Thépot S and Lagarce F:
Advances in treatment formulations for acute myeloid leukemia. Drug
Discov. Today. 23:1936–1949. 2018.
|
|
6
|
Salerno L, Romeo G, Modica MN, Amata E,
Sorrenti V, Barbagallo I and Pittalà V: Heme oxygenase-1: A new
druggable target in the management of chronic and acute myeloid
leukemia. Eur J Med Chem. 142:163–178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tsuruo T, Iida H, Tsukagoshi S and Sakurai
Y: Overcoming of vincristine resistance in P388 leukemia in vivo
and in vitrothrough enhanced cytotoxicity of vincristine and
vinblastine by verapamil. Cancer Res. 41:1967–1972. 1981.PubMed/NCBI
|
|
8
|
Batra P and Sharma AK: Anti-cancer
potential of flavonoids: Recent trends and future perspectives. 3
Biotech. 3:439–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jacquemin G, Granci V, Gallouet AS,
Lalaoui N, Morlé A, Iessi E, Morizot A, Garrido C, Guillaudeux T
and Micheau O: Quercetin-mediated Mcl-1 and survivin downregulation
restores TRAIL-induced apoptosis in non-Hodgkin's lymphoma B cells.
Haematologica. 97:38–46. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goel A, Kunnumakkara AB and Aggarwal BB:
Curcumin as ‘Curecumin’: From kitchen to clinic. Biochem Pharmacol.
75:787–809. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ravindran J, Prasad S and Aggarwal BB:
Curcumin and cancer cells: How many ways can curry Kill tumor cells
selectively? AAPS J. 11:495–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Banerjee S, Ji C, Mayfield JE, Goel A,
Xiao J, Dixon JE and Guo X: Ancient drug curcumin impedes 26S
proteasome activity by direct inhibition of dual-specificity
tyrosine-regulated kinase 2. Proc Natl Acad Sci USA. 115:8155–8160.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Romiti N, Tongiani R, Cervelli F and
Chieli E: Effects of curcumin on P-glycoprotein in primary cultures
of rat hepatocytes. Life Sci. 62:2349–2358. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gu Y, Li J, Li Y, Song L, Li D, Peng L,
Wan Y and Hua S: Nanomicelles loaded with doxorubicin and curcumin
for alleviating multidrug resistance in lung cancer. Int J
Nanomedicine. 11:5757–5770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mapoung S, Pitchakarn P, Yodkeeree S,
Ovatlarnporn C, Sakorn N and Limtrakul P: Chemosensitizing effects
of synthetic curcumin analogs on human multi-drug resistance
leukemic cells. Chem Biol Interact. 244:140–148. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maia RC, Vasconcelos FC, Souza PS and
Rumjanek VM: Towards Comprehension of the ABCB1/P-glycoprotein role
in chronic myeloid leukemia. Molecules. 23(pii): E1192018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lalioti VS, Ilari A, O'Connell DJ, Poser
E, Sandoval IV and Colotti G: Sorcin links calcium signaling to
vesicle trafficking, regulates Polo-like kinase 1 and is necessary
for mitosis. PLoS One. 9:e854382014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bresnick AR, Weber DJ and Zimmer DB: S100
proteins in cancer. Nat Rev Cancer. 15:96–109. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salama I, Malone PS, Mihaimeed F and Jones
JL: A review of the S100 proteins in cancer. Eur J Surg Oncol.
34:357–364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nicolas E, Ramus C, Berthier S, Arlotto M,
Bouamrani A, Lefebvre C, Morel F, Garin J, Ifrah N, Berger F, et
al: Expression of S100A8 in leukemic cells predicts poor survival
in de novo AML patients. Leukemia. 25:57–65. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Spijkers-Hagelstein JA, Schneider P,
Hulleman E, de Boer J, Williams O, Pieters R and Stam RW: Elevated
S100A8/S100A9 expression causes glucocorticoid resistance in
MLL-rearranged infant acute lymphoblastic leukemia. Leukemia.
26:1255–1265. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang XY, Zhang MY, Zhou Q, Wu SY, Zhao Y,
Gu WY, Pan J, Cen JN, Chen ZX, Guo WG, et al: High expression of
S100A8 gene is associated with drug resistance to etoposide and
poor prognosis in acute myeloid leukemia through influencing the
apoptosis pathway. Onco Targets Ther. 9:4887–4899. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang M, Zeng P, Kang R, Yu Y, Yang L, Tang
D and Cao L: S100A8 contributes to drug resistance by promoting
autophagy in leukemia cells. PLoS One. 9:e972422014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lampidis TJ, Shi YF, Calderon CL, Kolonias
D, Tapiero H and Savaraj N: Accumulation of simple organic cations
correlates with differential cytotoxicity in multidrug-resistant
and -sensitive human and rodent cells. Leukemia. 11:1156–1159.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Y, Zhang SP and Cai YQ: Cytoprotective
effects of selenium on cadmium-induced LLC-PK1 cells apoptosis by
activating JNK pathway. Toxicol In Vitro. 21:677–684. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou H, Qian J, Wang J, Yao W, Liu C, Chen
J and Cao X: Enhanced bioactivity of bone morphogenetic protein-2
with low dose of 2-N, 6-O-sulfated chitosan in vitro and in vivo.
Biomaterials. 30:1715–1724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kapuscinski J: DAPI: A DNA-specific
fluorescent probe. Biotech Histochem. 70:220–233. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan P, Li T, Bo M, Die L and Xing L:
Inhibition of bone resorption by econazole in rat osteoclast-like
cells through suppressing TRPV5. Arch Pharm Res. 34:1007–1013.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yhee JY, Song S, Lee SJ, Park SG, Kim KS,
Kim MG, Son S, Koo H, Kwon IC, Jeong JH, et al: Cancer-targeted
MDR-1 siRNA delivery using self-cross-linked glycol chitosan
nanoparticles to overcome drug resistance. J Control Release.
198:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yan LL, Huang YJ, Yi X, Yan XM, Cai Y, He
Q and Han ZJ: Effects of silencing S100A8 and S100A9 with small
interfering RNA on the migration of CNE1 nasopharyngeal carcinoma
cells. Oncol Lett. 9:2534–2540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Morsy MA, El-Sheikh AAK, Ibrahim ARN,
Khedr MA and Al-Taher AY: In silico comparisons between natural
inhibitors of ABCB1/P-glycoprotein to overcome
doxorubicin-resistance in the NCI/ADR-RES cell line. Eur J Pharm
Sci. 112:87–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fletcher JI, Williams RT, Henderson MJ,
Norris MD and Haber M: ABC transporters as mediators of drug
resistance and contributors to cancer cell biology. Drug Resist
Updat. 26:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Servili E, Trus M, Maayan D and Atlas D:
β-Subunit of the voltage-gated Ca2+ channel Cav1.2 drives signaling
to the nucleus via H-Ras. Proc Natl Acad Sci USA. 115:E8624–E8633.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
DeCaen PG, Delling M, Vien TN and Clapham
DE: Direct recording and molecular identification of the calcium
channel of primary cilia. Nature. 504:315–318. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Monteith GR, Prevarskaya N and
Roberts-Thomson SJ: The calcium-cancer signalling nexus. Nat Rev
Cancer. 17:367–380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hajnóczky G, Booth D, Csordás G,
Debattisti V, Golenár T, Naghdi S, Niknejad N, Paillard M, Seifert
EL and Weaver D: Reliance of ER-mitochondrial calcium signaling on
mitochondrial EF-hand Ca 2+ binding proteins: Miros, MICUs, LETM1
and solute carriers. Curr Opin Cell Biol. 29:133–141. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Andjelkovic T, Pesic M, Bankovic J, Tanic
N, Markovic ID and Ruzdijic S: Synergistic effects of the purine
analog sulfinosine and curcumin on the multidrug resistant human
non-small cell lung carcinoma cell line (NCI-H460/R). Cancer Biol
Ther. 7:1024–1032. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chiu LY, Ko JL, Lee YJ, Yang TY, Tee YT
and Sheu GT: L-type calcium channel blockers reverse docetaxel and
vincristine-induced multidrug resistance independent of ABCB1
expression in human lung cancer cell lines. Toxicol Lett.
192:408–418. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Prasad N, Sudhakar YA and Kanwar JR:
Curcumin regulates colon cancer by inhibiting P-Glycoprotein in
In-situ Cancerous colon perfusion rat model. J Cancer Sci Ther.
5:313–319. 2013.PubMed/NCBI
|
|
40
|
Shah K, Mirza S, Desai U, Jain N and Rawal
R: Synergism of curcumin and cytarabine in the down regulation of
Multi-Drug resistance genes in acute myeloid leukemia. Anticancer
Agents Med Chem. 16:128–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Misra R and Sahoo SK: Coformulation of
doxorubicin and curcumin in poly(D,L-lactide-co-glycolide)
nanoparticles suppresses the development of multidrug resistance in
K562 cells. Mol Pharm. 8:852–866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Anuchapreeda S, Thanarattanakorn P,
Sittipreechacharn S, Tima S, Chanarat P and Limtrakul P: Inhibitory
effect of curcumin on MDR1 gene expression in patient leukemic
cells. Arch Pharm Res. 29:866–873. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Choi BH, Kim CG, Lim Y, Shin SY and Lee
YH: Curcumin down-regulates the multidrug-resistance mdr1b gene by
inhibiting the PI3K/Akt/NF kappa B pathway. Cancer Lett.
259:111–118. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Robey RW, Pluchino KM, Hall MD, Fojo AT,
Bates SE and Gottesman MM: Revisiting the role of ABC transporters
in multidrug-resistant cancer. Nat Rev Cancer. 18:452–464. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yang SS, Liu AL, Shan LL, Zeng TC, Zhou Q
and Li YB: Pharmacokinetics mechanism of ABC efflux
proteins-mediated seven features of compatibility. Zhongguo Zhong
Yao Za Zhi. 43:676–683. 2018.(In Chinese). PubMed/NCBI
|
|
46
|
Efferth T and Volm M: Multiple resistance
to carcinogens and xenobiotics: P-glycoproteins as universal
detoxifiers. Arch Toxicol. 91:2515–2538. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huang J, Guo L, Tan R, Wei M, Zhang J,
Zhao Y, Gong L, Huang Z and Qiu X: Interactions between emodin and
efflux transporters on rat enterocyte by a validated ussing chamber
technique. Front Pharmacol. 9:6462018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ampasavate C, Sotanaphun U, Phattanawasin
P and Piyapolrungroj N: Effects of curcuma spp. on P-glycoprotein
function. Phytomedicine. 17:506–512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Monteith GR, Prevarskaya N and
Roberts-Thomson SJ: The calcium-cancer signalling nexus. Nat Rev
Cancer. 17:367–380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kania E, Roest G, Vervliet T, Parys JB and
Bultynck G: IP3Receptor-mediated calcium signaling and its role in
autophagy in cancer. Front Oncol. 7:1402017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Dubois C, Vanden Abeele F and Prevarskaya
N: Targeting apoptosis by the remodelling of calcium-transporting
proteins in cancerogenesis. FEBS J. 280:5500–5510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Roderick HL and Bootman MD: Redoxing
calcium from the ER. Cell. 120:4–5. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Clapham DE: Calcium signaling. Cell.
131:1047–1058. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Luciani DS, Gwiazda KS, Yang TL, Kalynyak
TB, Bychkivska Y, Frey MH, Jeffrey KD, Sampaio AV, Underhill TM and
Johnson JD: Roles of IP3R and RyR Ca2+Channels in endoplasmic
reticulum stress and β-cell death. Diabetes. 58:422–432. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dubois C, Vanden Abeele F, Sehgal P,
Olesen C, Junker S, Christensen SB, Prevarskaya N and Møller JV:
Differential effects of thapsigargin analogues on apoptosis of
prostate cancer cells: Complex regulation by intracellular calcium.
FEBS J. 280:5430–5440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ansari N, Hadi-Alijanvand H, Sabbaghian M,
Kiaei M and Khodagholi F: Interaction of 2-APB, dantrolene, and
TDMT with IP3R and RyR modulates ER stress-induced programmed cell
death I and II in neuron-like PC12 cells: An experimental and
computational investigation. J Biomol Struct Dyn. 32:1211–1230.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bouman L, Schlierf A, Lutz AK, Shan J,
Deinlein A, Kast J, Galehdar Z, Palmisano V, Patenge N, Berg D, et
al: Parkin is transcriptionally regulated by ATF4: Evidence for an
interconnection between mitochondrial stress and ER stress. Cell
Death Differ. 18:769–782. 2011. View Article : Google Scholar : PubMed/NCBI
|