Approximately 93% of human genome DNA is transcribed
into RNAs, but <2% of these nucleotide sequences can code for
proteins, while the other 98% are non-coding RNAs (ncRNAs) that
partially or completely lack the ability to be coded into proteins.
The majority of these ncRNAs are known as long non-coding RNAs
(lncRNAs) whose length exceeds 200 nucleotides (1). LncRNAs were at first regarded as the
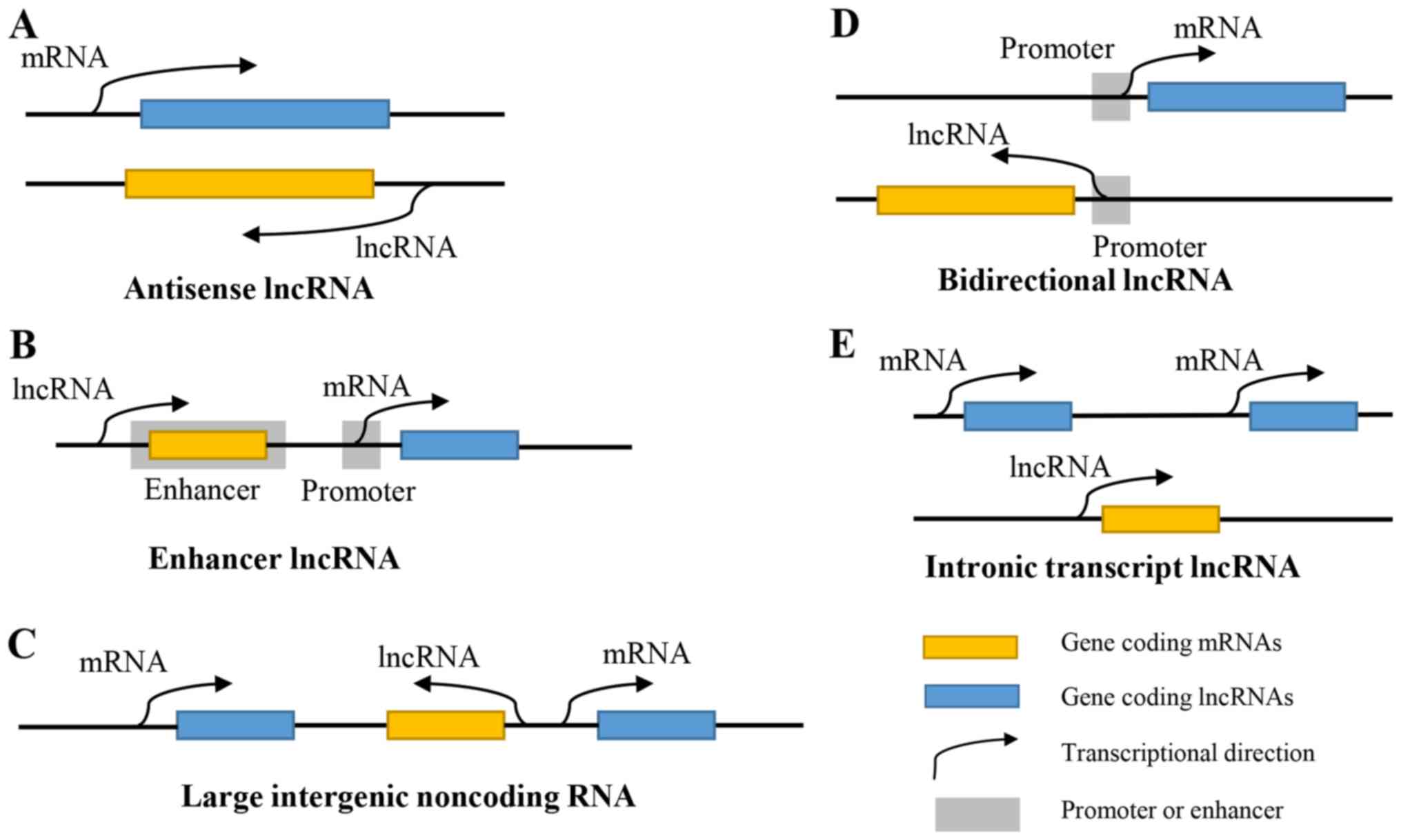
‘noise’ of gene transcription. According to the position where they
are relative to the protein-coding genes, lncRNAs can be roughly
divided into antisense lncRNAs, enhancer lncRNAs, large intergenic
non-coding RNAs, bidirectional lncRNAs and intronic transcript
lncRNAs (2) (Fig. 1). With the existence and development
of next-generation sequencing, lncRNAs have been demonstrated to
affect multiple biological processes, including housekeeping
functions and specialized functions, such as genomic imprinting and
dosage compensation (3). LncRNAs can
regulate gene expression via multiple mechanisms, including at the
epigenetic, transcriptional and post-transcriptional levels
(4). They participate in the
regulation of a variety of cell activities, such as cell
differentiation, proliferation, invasion, apoptosis and autophagy.
through interacting with RNAs, DNAs or proteins (5).
The H19 gene is located in 11p15.5, an imprinted
region of chromosome 11, being adjacent to the insulin-like growth
factor 2 (IGF2) gene, and is expressed only from the maternally
inherited chromosome, while IGF2 is expressed only from the
paternally inherited chromosome. H19, a lncRNA, is the
transcription product of the H19 gene, and diversified transcript
variants exist due to alternative splicing (6). Although H19 RNA molecules can be
detected in both the cytoplasm and nucleus, H19 RNA primarily
exists in cytoplasm. H19 functions in the form of regulatory RNA or
ribosome regulators. H19 promotes biological processes such as
apoptosis, angiogenesis, inflammation and cell death (7). Furthermore, Gene Ontology (GO) analyses
predicted that H19 is connected with neurogenesis, angiogenesis and
inflammation through DNA transcription, RNA folding, methylation
and gene imprinting (8). The
aberrant expression of H19 is associated with multiple diseases,
including carcinoma, sarcoma, type 2 diabetes and hypertrophic
cardiomyopathy (9–12). The present review primarily
summarizes the role of H19 in the carcinogenesis and development of
breast cancer.
Abnormal expression of H19 has been demonstrated in
a variety different types of cancer cells, such as gastric cancer
(13), pancreatic cancer (14), liver cancer (15) and breast cancer (16), affecting the development and
progression of cancer through various mechanisms. H19 is an
estrogen-regulated transcript, and a wide array of studies have
confirmed that H19 regulates cell differentiation and proliferation
(6). In addition, research into its
application in the clinical setting revealed that H19 had the
potential to serve as a biomarker of cancer, and had promise as a
novel therapeutic option in oncotherapy (6,7).
However, whether H19 functions as a tumor promoter or a tumor
suppressor remains controversial (17). It is possible that H19 plays
differential roles depending on the tissue type and developmental
stage (6). Therefore, the clear-cut
mechanisms underlying how H19, as an lncRNA, regulates the
biological progression of cancer requires further investigation.
The various roles of H19 in different kinds of cancers are briefly
summarized in Table I.
Breast cancer is a malignant tumor deriving from the
epithelium of the mammary gland, and >99% of breast cancer cases
occur in women worldwide; less than 1% (0.02%) of all breast cancer
cases develop in men (18).
According to data released by Globocan in 2012, the standardized
mortality rate of breast cancer in the developed world was
149/100,000, and that in the less developed world was 115/100,000
(19). The global incidence and
mortality rate of breast cancer increased in the late 1970s, while
the mortality rate indicated signs of decline in the 1990s due to
the introduction of breast cancer screening and comprehensive
treatment. Metastasis is the leading cause of mortality in patients
with breast cancer. The H19 gene is an oncogene in breast cancer
and is highly expressed in cancer tissues compared with normal
tissues. The expression level of H19 is associated with the
oncogenesis, proliferation, invasion, metastasis and drug
resistance of breast cancer, but the underlying molecular
mechanisms are very different (7).
Thus, serum H19 levels may possess clinical significance in the
early diagnosis, treatment and prognosis of breast cancer.
A recent case-control study in China revealed that
high expression levels of H19 were associated with an increased
risk of breast carcinogenesis in both codominant and dominant
models, and the association was more apparent in patients with
estrogen receptor-positive (ER+), human epidermal growth factor
receptor 2-negative (HER2-), and ER+-HER2-negative (HER2-)
molecular subtypes (20). In
addition, two single nucleotide polymorphisms (SNPs) were
identified, and another study suggested that the expression levels
of H19 in breast cancer cells with the rs2071095 CC genotype were
significantly higher than those with the AA genotype, and that the
CC genotype was markedly associated with an increased risk of
breast cancer compared with the CA + AA genotypes (21). Furthermore, animal experiments
demonstrated that the probability of breast carcinogenesis was
increased in severe combined immunodeficiency mice injected with
cells overexpressing the H19 gene (22).
The molecular mechanisms underlying H19-associated
carcinogenesis may involve several aspects. The H19 promoter was
activated by E2F transcription factor 1 (E2F1), which promoted cell
cycle progression (particularly in the S-phase) of MCF-7 cells
(23). Furthermore, H19 contributed
to the epigenetic regulation of gene expression in breast cancer.
H19 bound to and inhibited S-adenosylhomocysteine hydrolase, the
sole enzyme that can hydrolyze S-adenosylhomocysteine (SAH) in
humans. SAH can markedly suppress S-adenosylmethionine-dependent
methyltransferases, which can methylate multiple cellular
components, including DNA, RNAs and proteins, through a feedback
mechanism. H19 knockdown increased the DNMT3B-mediated methylation
of Nctc1, a gene encoding lncRNAs, within the Igf2-H19-Nctc1 locus.
Thus, H19 altered DNA methylation and led to breast tumorigenesis
(24). A new lncRNA within the
H19/IGF2 locus named 91H is an antisense gene to H19. The 91H
lncRNA also regulated the expression levels of H19 and IGF2 by
epigenetic modifications and increased the tumorigenic properties
of MDA-MB-231 cells both in vitro and in vivo
(25). In summary, H19 plays an
oncogenic role in breast tumorigenesis.
H19 is a precursor for miR-675 and generates two
functional miRNAs, miR-675-5p and miR-675-3p (26). miR-675 is harbored in the first exon
of H19 and miR-675 regulated by H19 was originally demonstrated to
restrict the growth of the placenta prior to birth (27). A study of miR-675 in formalin-fixed
paraffin-embedded (FFPE) tissues demonstrated that the expression
of miR-675 was significantly higher in the FFPE tissues of patients
with breast cancer compared with the control group. However, the
expression levels were not associated with age, lymph node stage,
ER status or progestrogen receptor (PR) status. The frequency of
miR-675 overexpression was higher in patients with low histological
grade (histological grade I–II) cancer. In addition, there was a
strong association between miR-675 and miR-24/93/98/378 levels in
patients with breast cancer (28).
In a prospective cohort study, DNA methylation levels of 517
miRNA-encoding genes were analyzed in the prediagnostic peripheral
leukocytes of patients with colorectal cancer (n=159) or breast
cancer (n=166). In the subjects who developed breast cancer, 8
miRNAs, including miR-675, were aberrantly methylated, while there
were no significant associations in those with colorectal cancer.
This aberrant methylation significantly affected the development of
breast cancer (29). A luciferase
assay revealed that H19/miR-675-5p decreased the expression levels
of the ubiquitin ligase E3 family (c-Cbl and Cbl-b), and activated
epidermal growth factor receptor (EGFR) and c-Met, enhancing the
proliferation and migration of MDA-MB-231 breast cancer cells
through the protein kinase B (Akt) and extracellular
signal-regulated kinase (Erk) pathways (30). Furthermore, miR-675 was also involved
in the mechanism through which H19 promoted Slug expression
accompanied by the suppression of E-cadherin, and thereby increased
the potential of cell invasion and metastasis in breast cancer
(31). Huaier extract, a traditional
Chinese medicine, decreased the expression levels of H19 and
miR-675-5p and increased the expression levels of calcineurin
B-like (CBL) protein. Huaier extract decreased the viability and
increased the apoptosis rates of MDA-MB-231 and MDA-MB-468 cells by
regulating the approach of the H19/miR-675-5p/CBL axis. Huaier
extract was expected to be a novel clinical treatment for breast
cancer (32). In summary, the
ectopic expression of H19/miR-675 reinforced the aggressive
phenotype of breast cancer cells. This abnormal expression
aggrandized cell proliferation, cell migration, tumor growth and
metastasis.
Multitudinous RNA transcripts have miRNA binding
sites, which are also known as miRNA response elements (MREs).
These RNA transcripts include mRNAs, pseudogene transcripts,
lncRNAs and circular RNAs (circRNAs). They can compete with each
other to bind to a shared miRNA through a common MRE, functioning
as ceRNAs. There are multiple MREs on mRNA transcripts of
protein-coding genes, and miRNAs can bind to corresponding mRNAs
through MREs, leading to the degradation of mRNAs or inhibition of
mRNA translation. Thus, RNA transcripts acting as ceRNAs can weaken
the negative regulation of miRNA on target genes, promoting the
expression of target genes. The example of lncRNAs as ceRNAs was
first described in plants. LncRNAs indirectly regulate the
expression level and translation of mRNAs through the bridge of
MREs, so as to regulate the biological process of cells (33–36). H19
acted as a sponge for miR-200b/c and let-7b and promoted the
expression of miRNA targets GIT2 and CYTH3, which motivated breast
cancer cell migration through the epithelial-to-mesenchymal
transition (EMT) and dissemination of breast cancer cells (37). In addition, H19 functioned as a ceRNA
and bound to miR-152, which directly targeted DNA methyltransferase
1 (DNMT1). The overexpression of H19 diminished the inhibitory
effect of miR-152 on DNMT1 expression and thereby promoted the
proliferation and invasion of MCF-7 and MDA-MB-231 cells (38). H19 was indicated to be a target of
miR-93-5p, and this molecule upregulated the expression levels of
signal transducers and activators of transcription 3 (STAT3), a
target of miR-93-5p. STAT3 significantly promotes the
proliferation, migration and invasion of MCF-7 and MDA-MB-231 cells
(39). H19 may also function as a
ceRNA to sponge miRNA let-7a/b, resulting in the high expression
level of the transcriptional factor LIN28. H19, let-7a/b and LIN28
formed a double-negative feedback loop that profoundly influenced
the migration of breast cancer stem cells (BCSCs) (40). This ceRNA regulating effect released
hypoxia-inducible factor 1α, leading to an increase of pyruvate
dehydrogenase kinase 1, which influenced BCSC reprogramming and
malignancy (41). In addition,
let-7-regulated-Wnt signaling pathway activity was enhanced by the
inhibitory effect of H19 on miR-146, which decreased asymmetric
division and increased the self-renewal of BCSCs. H19 had a
positive feedback regulatory effect on let-7 (42). Inversely, the knockdown of H19
reactivated let-7c, restraining the oestrogen receptor activated
Wnt signaling pathway and inhibited the symmetric division of BCSCs
(43).
Certain pseudogene transcripts also act as ceRNAs
and affect H19 expression in breast cancer cells. For instance,
HMGA1P7 mRNA, a transcription product of the HMGA1 gene, may
increase the expression levels of H19 and IGF2 by a ceRNA mechanism
and consequently accelerate cancer progression (44).
Similar regulatory mechanisms exist in other types
of tumor, such as H19/miR-29b-3p in lung adenocarcinoma (45) and multiple myeloma (46), H19/miR-324-5p in ovarian carcinoma
(47), H19/miR-194 in pancreatic
carcinoma (48), H19/miR-106b-5p in
seminoma (49), H19/miR-3126-5p in
papillary thyroid carcinoma (50),
H19/miR-326 in hepatocellular carcinoma (51) and H19/miR-194-5p in colorectal
adenocarcinoma (52).
Thus, the ceRNA regulatory network of H19 plays a
pivotal role in the proliferation, invasion and metastasis of
breast cancer.
The oncogene MYC, including c-Myc, n-Myc and l-Myc,
is deregulated in various types of tumor cell, including breast
cancer cells. The overexpression of H19 influenced the
let-7-mediated regulation of certain metastasis-promoting genes,
including c-Myc (53). c-Myc protein
bound to evolutionarily conserved E-boxes close to the imprinting
control region to promote the histone acetylation and
transcriptional initiation of the H19 promoter, distinctly inducing
H19 expression (54). Furthermore,
the c-Myc mRNA coding region instability determinant binding
protein (CRD-BP) may bind to c-Myc and H19 and influence RNA
stability, localization and translation. CRD-BP has been
demonstrated to be associated with tumorigenesis, and experimental
evidence suggested that CRD-BP was a proto-oncogene (55). Vigilin, a multi-K homology-domain
protein, was present at several target sites of a zinc finger
protein named CCCTC-binding factor, including promoter regions of
c-Myc and several regions within the Igf2/H19 locus (56). Apart from vigilin,
double-strand-break repair protein RAD21 and structural maintenance
of chromosome protein 1 also bound to such regions and were
associated with complicated chromatin organization and gene
regulation (57). Furthermore, n-Myc
has been demonstrated as a forceful glycogen synthase
kinase-3-dependent regulator of DNA methyltransferase 3α 2
expression, changing the expression of H19 and the DNA methylation
of corresponding imprinted loci via influencing signal transduction
(58), which altered the biological
process of breast cancer cells. In conclusion, the association
between oncogene MYC and H19 expression plays a large role in the
oncogenesis, proliferation, invasion and metastasis of breast
cancer.
The overexpression of H19 is associated with cells
exhibiting higher tumorigenic phenotypes (22), which indicates that H19 expression
levels can be used in the clinical diagnosis of breast cancer. A
statistical analysis of clinical data revealed that H19 expression
was significantly increased in breast cancer biopsies and plasma
compared with that in the control group, and there was a
significant correlation between the levels of plasma H19 and ER,
PR, c-erbB-2 and lymph node metastasis in breast cancer. The
diagnostic value of H19 for breast cancer was higher than
carbohydrate antigen 153 and carcinoembryonic antigen (59). Reverse transcription-quantitative PCR
was conducted in 12 lncRNAs that were confirmed as abnormally
expressed in breast cancer cells, and the experimental results
demonstrated that H19 was one of three lncRNAs (H19, HOTAIR and
RP11-445H22.4) that notably increased in patients with breast
cancer compared with the other nine lncRNAs (60). Furthermore, chromogenic in
situ hybridization (CISH) was performed using the breast cancer
tissues of 52 patients. The expression levels of H19 were disparate
in different types of breast cancer. H19 was expressed in invasive
breast carcinoma, ductal carcinoma in situ and normal
adjacent tissue from higher to lower level, respectively (61). These data suggested that CISH with
H19 is a potential clinical diagnostic assay with the ability to
accurately classify breast cancer. In short, H19 possesses marked
diagnostic significance and may serve as a potential biomarker for
the early diagnosis of breast cancer.
The expression levels of H19 were associated with
tumor size, lymph node status and hormone negativity. Patients with
breast cancer who exhibited high H19 expression levels had
relatively unfavorable disease-free survival and overall survival
(OS). The most prominent effect of H19 expression on clinical
cancer prognosis was observed in triple-negative breast cancer
(TNBC) (62). Another study examined
the association between SNPs tagging the low-risk breast cancer
loci in or near eight lncRNAs and the prognosis of breast cancer.
The results revealed that only SNP rs2107425 near H19 were
significantly associated with shorter metastasis-free survival
(P=0.006 in univariate analysis; P=0.004 in multivariate analysis)
(63). The postsurgical expression
levels of H19 were significantly decreased in 82.1% of patients
with breast cancer (32/39) compared with those in paired
preoperative plasma (64), which
indicated that the expression levels of H19 may reflect the
curative effect of radical mastectomy. In summary, a negative
correlation exists between the expression levels of H19 and the
prognosis of patients with breast cancer.
H19 is a downstream target molecule of ERα, and the
expression of ERα has been demonstrated to alter H19 levels, which
decreases cell apoptosis in response to paclitaxel (PTX) treatment
by suppressing the expression of the pro-apoptotic genes BIK and
NOXA (65). Concrete mechanisms may
include the inhibiting effect of H19 on the promoter activity of
BIK via recruiting enhancer of zeste homolog 2 and trimethylating
the lysine 27 of histone H3 (65).
Endocrine therapies are widely used as an effective treatment for
ERα+ breast cancer, such as Luminal A and Luminal B.
However, the frequent occurrence of endocrine therapy resistance
(ETR) leads to increased disease relapse and decreased OS. The
expression levels of H19 were positively correlated with the
expression of ERα in primary breast tumors (66). H19 regulated ERα expression at the
transcript and protein levels and H19/ERα levels could be
significantly downregulated by blocking the Notch and c-MET
signaling pathway. Pharmacological inhibitors against this
signaling pathway helped reverse the resistance to endocrinotherapy
such as Tamoxifen and Fulvestrant (67). In addition, the expression levels of
H19 were significantly increased in PTX-resistant TNBC cells
compared with that in PTX-sensitive TNBC cells. Inversely, the
knockdown of H19 can recover the chemosensitivity of PTX-resistant
TNBC by mediating the Akt signaling pathway (16). Furthermore, overexpression of H19
enhanced breast cancer cell resistance to doxorubicin (Dox). The
major effector proteins of H19-induced Dox resistance may include
multidrug resistance protein 1 (MDR1) and multidrug resistance
protein 4. The ubiquitin ligase component cullin 4A (CUL4A) has
been regarded as an important factor that connects H19 and MDR1
expression, and the high expression levels of CUL4A compromised the
cell chemical sensitivity to Dox and led to lower survival rates of
patients with breast cancer undergoing chemotherapy (68). In summary, H19 affects the drug
resistance and chemosensitivity of breast cancer cells. As a
consequence, the response of breast cancer cells to
chemotherapeutic drugs weakens, which leads to a decrease in the
curative effect for the tumor.
Breast cancer is the most common malignant tumor in
women in the majority of countries and usually occurs in women at
35–70 years of age (69,70). In recent years, breast cancer
incidence has demonstrated a younger trend (71). The incidence of breast cancer had
been demonstrated to be associated with multiple factors, such as a
family history of breast cancer, early menarche, not being married,
not having had children, radiographic exposure, long-term use of
exogenous estrogen, postmenopausal obesity and alcohol consumption
(72). Therefore, the tertiary
prevention of breast cancer appears to be particularly important.
In addition, primary and secondary prevention, including decreasing
risk factors, early diagnosis and timely treatment, are key ways to
decrease the mortality rate of patients with breast cancer.
According to a number of studies, many lncRNAs can
be used as potential biomarkers in the diagnosis and prognostic
evaluation of tumors. Excluding H19, there are a number of lncRNAs
recently investigated in breast cancer, including NORAD (73), MIR2052HG (74), NAMPT-AS (75), HISLA (76), TROJAN (77), TINCR (78) and LINC00511 (79), some of which have also been reported
as potential biomarkers. Among these lncRNAs, H19 is abnormally
expressed in various cancer cells and is involved in the regulation
of multiple physiological and pathological processes, affecting the
occurrence and development of different types of cancer, including
breast cancer. Likewise, the development of human breast cancer is
influenced by polygenes and multifactor regulation, including H19
regulation (6,7). The regulatory mechanisms involve
numerous aspects, such as encoding miR-675, regulating ceRNA,
regulating the EMT and regulating tumor angiopoiesis.
The primary mechanisms underlying the effect that
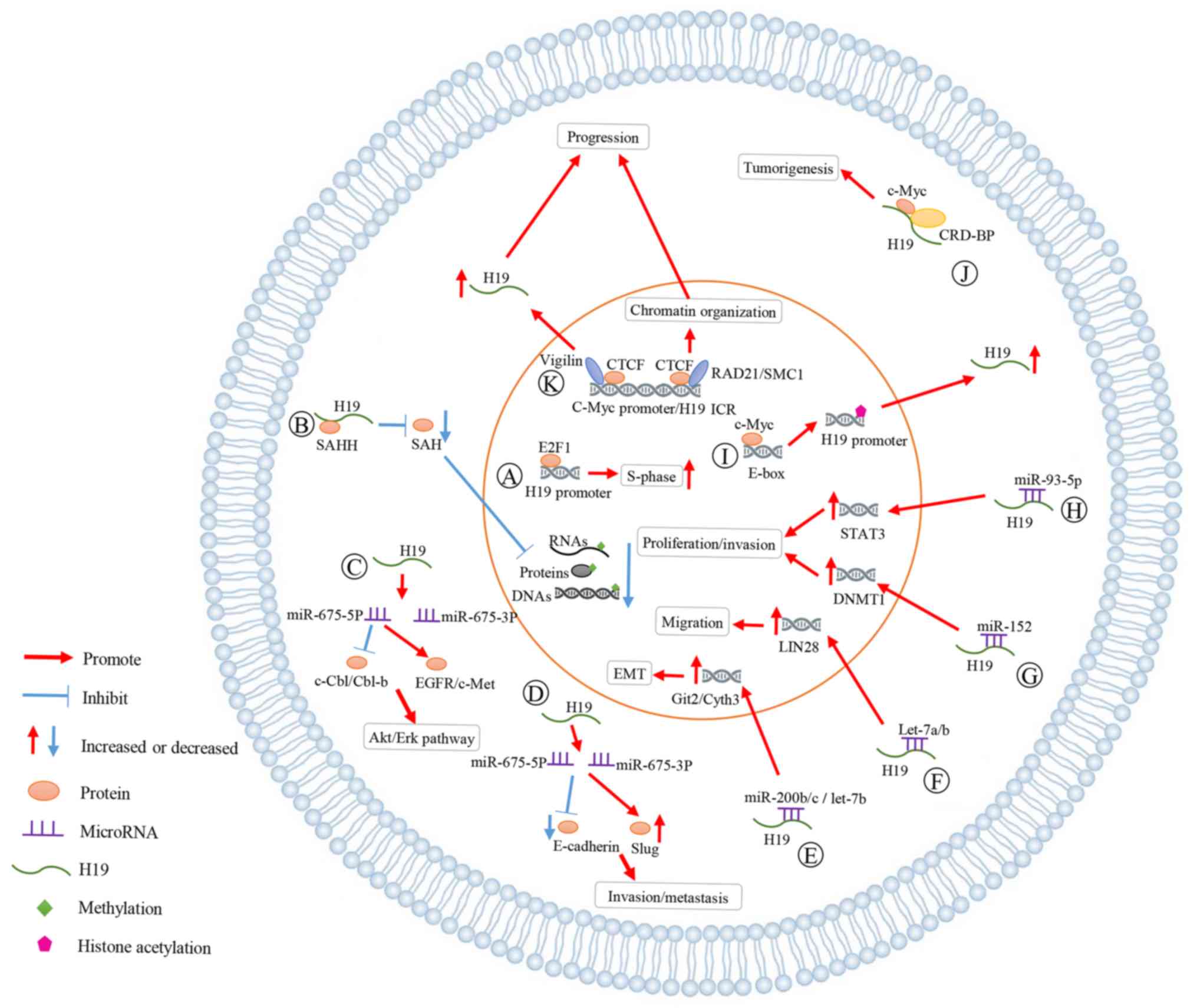
H19 has on breast cancer are summarized in Fig. 2. A: As indicated, E2F1 activated H19
promoter to accelerate cell cycle progression (particularly the
S-phase) of breast cancer cells. B: H19 bound SAHH to increase SAH
levels and suppressed the methylation of DNA, RNAs and proteins. C:
H19 generated miR-675-5p and miR-675-3p. H19/miR-675-5p decreased
the expression levels of c-Cbl/Cbl-b and activated EGFR/c-Met,
enhancing the proliferation and migration of breast cancer via Akt
and Erk pathways. D: miR-675 was involved in the process in which
H19 increased Slug expression and inhibited E-cadherin, promoting
the cell invasion and metastasis in breast cancer. E-H: H19 acted
as a ceRNA for miR-200b/c, let-7a/b, miR-152 and miR-93-5p,
increasing the expression of related genes. The interaction between
MYC and H19 has great influences on the development of breast
cancer. I: C-Myc bound evolutionarily conserved E-boxes close to
ICR to promote histone acetylation and transcriptional initiation
of the H19 promoter, increasing the H19 expression. J: The complex
of CRD-BP, c-Myc and H19 promoted tumorigenesis of breast cancer.
K: Vigilin was present at several CTCF target sites, such as c-Myc
promoter regions and IGF2/H19 locus, upregulating the H19
expression, and affecting the chromatin organization.
Currently, certain deficiencies and problems remain
in the studies that focus on the association between H19 and breast
cancer. For instance, the potential mechanisms underlying
H19/miR-675 regulation in gene expression and any specific
molecular mechanisms underlying the interaction between H19 and MYC
remain to be elucidated. In addition, the combination of basic
research into H19 and the clinical diagnosis of breast cancer could
be more proficient, and a unified standard for judging the
expression level of H19 in the clinical setting should be
available. Furthermore, to the best of our knowledge, there are few
studies that have investigated whether H19 can be used in the
treatment of breast cancer. In order to apply the studies on H19 to
the clinical treatment of breast cancer, further research is
required to comprehensively understand the complicated underlying
mechanisms through which H19 regulates the biological
characteristics of human breast cancer. As research on H19
progresses, anticancer drugs targeting H19 will be used to treat
breast cancer more safely and effectively.
Not applicable.
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 31401094)
and the Natural Science Foundation of Jiangsu Province, China
(grant no. BK20181365).
Not applicable.
JW and JS conceived the study and drafted the
manuscript. FY critically revised the manuscript. All authors read
and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Kapranov P, Cheng J, Dike S, Nix DA,
Duttagupta R, Willingham AT, Stadler PF, Hertel J, Hackermuller J,
Hofacker IL, et al: RNA maps reveal new RNA classes and a possible
function for pervasive transcription. Science. 316:1484–1488. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dahariya S, Paddibhatla I, Kumar S,
Raghuwanshi S, Pallepati A and Gutti RK: Long non-coding RNA:
Classification, biogenesis and functions in blood cells. Mol
Immunol. 112:82–92. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Charles Richard JL and Eichhorn PJA:
Platforms for investigating LncRNA functions. SLAS Technol.
23:493–506. 2018.PubMed/NCBI
|
|
4
|
Jia B, Xie T, Qiu X, Sun X, Chen J, Huang
Z, Zheng X, Wang Z and Zhao J: Long noncoding RNA FALEC inhibits
proliferation and metastasis of tongue squamous cell carcinoma by
epigenetically silencing ECM1 through EZH2. Aging (Albany NY).
11:4990–5007. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lim LJ, Wong S, Huang F, Lim S, Chong SS,
Ooi LL, Kon OL and Lee CG: Roles and regulation of long non-coding
RNAs in hepatocellular carcinoma. Cancer Res. 79:5131–5139. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raveh E, Matouk IJ, Gilon M and Hochberg
A: The H19 Long non-coding RNA in cancer initiation, progression
and metastasis-a proposed unifying theory. Mol Cancer. 14:1842015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoshimura H, Matsuda Y, Yamamoto M, Kamiya
S and Ishiwata T: Expression and role of long non-coding RNA H19 in
carcinogenesis. Front Biosci (Landmark Ed). 23:614–625. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bao MH, Szeto V, Yang BB, Zhu SZ, Sun HS
and Feng ZP: Long non-coding RNAs in ischemic stroke. Cell Death
Dis. 9:2812018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zeng Y, Li TL, Zhang HB, Deng JL, Zhang R,
Sun H, Wan ZR, Liu YZ, Zhu YS and Wang G: Polymorphisms in IGF2/H19
gene locus are associated with platinum-based chemotherapeutic
response in Chinese patients with epithelial ovarian cancer.
Pharmacogenomics. 20:179–188. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ghaedi H, Zare A, Omrani MD, Doustimotlagh
AH, Meshkani R, Alipoor S and Alipoor B: Genetic variants in long
noncoding RNA H19 and MEG3 confer risk of type 2 diabetes in an
Iranian population. Gene. 675:265–271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gomez J, Lorca R, Reguero JR, Martín M,
Morís C, Alonso B, Iglesias S, Díaz-Molina B, Avanzas P and Coto E:
Genetic variation at the long noncoding RNA H19 gene is associated
with the risk of hypertrophic cardiomyopathy. Epigenomics.
10:865–873. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tarnowski M, Tkacz M, Czerewaty M,
Poniewierska-Baran A, Grymuła K and Ratajczak MZ: 5-Azacytidine
inhibits human rhabdomyosarcoma cell growth by downregulating
insulin-like growth factor 2 expression and reactivating the H19
gene product miR-675, which negatively affects insulin-like growth
factors and insulin signaling. Int J Oncol. 46:2241–2250. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei Y, Liu Z and Fang J: H19 functions as
a competing endogenous RNA to regulate human epidermal growth
factor receptor expression by sequestering let7c in gastric cancer.
Mol Med Rep. 17:2600–2606. 2018.PubMed/NCBI
|
|
14
|
Ma L, Tian X, Guo H, Zhang Z, Du C, Wang
F, Xie X, Gao H, Zhuang Y, Kornmann M, et al: Long noncoding RNA
H19 derived miR-675 regulates cell proliferation by down-regulating
E2F-1 in human pancreatic ductal adenocarcinoma. J Cancer.
9:389–399. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Han C, Ungerleider N, Chen W,
Song K, Wang Y, Kwon H, Ma W and Wu T: A transforming growth
factor-β and H19 signaling axis in tumor-initiating hepatocytes
that regulates hepatic carcinogenesis. Hepatology. 69:1549–1563.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han J, Han B, Wu X, Hao J, Dong X, Shen Q
and Pang H: Knockdown of lncRNA H19 restores chemo-sensitivity in
paclitaxel-resistant triple-negative breast cancer through
triggering apoptosis and regulating Akt signaling pathway. Toxicol
Appl Pharmacol. 359:55–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
El Hajj J, Nguyen E, Liu Q, Bouyer C,
Adriaenssens E, Hilal G and Ségal-Bendirdjian E: Telomerase
regulation by the long non-coding RNA H19 in human acute
promyelocytic leukemia cells. Mol Cancer. 17:852018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jemal A, Siegel R, Ward E, Murray T, Xu J,
Smigal C and Thun MJ: Cancer statistics, 2006. CA Cancer J Clin.
56:106–130. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin Y, Fu F, Chen Y, Qiu W, Lin S, Yang P,
Huang M and Wang C: Genetic variants in long noncoding RNA H19
contribute to the risk of breast cancer in a southeast China Han
population. Onco Targets Ther. 10:4369–4378. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui P, Zhao Y, Chu X, He N, Zheng H, Han
J, Song F and Chen K: SNP rs2071095 in LincRNA H19 is associated
with breast cancer risk. Breast Cancer Res Treat. 171:161–171.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lottin S, Adriaenssens E, Dupressoir T,
Berteaux N, Montpellier C, Coll J, Dugimont T and Curgy JJ:
Overexpression of an ectopic H19 gene enhances the tumorigenic
properties of breast cancer cells. Carcinogenesis. 23:1885–1895.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Berteaux N, Lottin S, Monte D, Pinte S,
Quatannens B, Coll J, Hondermarck H, Curgy JJ, Dugimont T and
Adriaenssens E: H19 mRNA-like noncoding RNA promotes breast cancer
cell proliferation through positive control by E2F1. J Biol Chem.
280:29625–29636. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou J, Yang L, Zhong T, Mueller M, Men Y,
Zhang N, Xie J, Giang K, Chung H, Sun X, et al: H19 lncRNA alters
DNA methylation genome wide by regulating S-adenosylhomocysteine
hydrolase. Nat Commun. 6:102212015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vennin C, Spruyt N, Robin YM, Chassat T,
Le Bourhis X and Adriaenssens E: The long non-coding RNA 91H
increases aggressive phenotype of breast cancer cells and
up-regulates H19/IGF2 expression through epigenetic modifications.
Cancer Lett. 385:198–206. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Collette J, Le Bourhis X and Adriaenssens
E: Regulation of human breast cancer by the long non-coding RNA
H19. Int J Mol Sci. 18(pii): E23192017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Keniry A, Oxley D, Monnier P, Kyba M,
Dandolo L, Smits G and Reik W: The H19 lincRNA is a developmental
reservoir of miR-675 that suppresses growth and Igf1r. Nat Cell
Biol. 14:659–665. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhai LL, Wang P, Zhou LY, Yin JY, Tang Q,
Zhang TJ, Wang YX, Yang DQ, Lin J and Deng ZQ: Over-expression of
miR-675 in formalin-fixed paraffin-embedded (FFPE) tissues of
breast cancer patients. Int J Clin Exp Med. 8:11195–11201.
2015.PubMed/NCBI
|
|
29
|
Cordero F, Ferrero G, Polidoro S, Fiorito
G, Campanella G, Sacerdote C, Mattiello A, Masala G, Agnoli C,
Frasca G, et al: Differentially methylated microRNAs in
prediagnostic samples of subjects who developed breast cancer in
the European Prospective Investigation into Nutrition and Cancer
(EPIC-Italy) cohort. Carcinogenesis. 36:1144–1153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vennin C, Spruyt N, Dahmani F, Julien S,
Bertucci F, Finetti P, Chassat T, Bourette RP, Le Bourhis X and
Adriaenssens E: H19 non coding RNA-derived miR-675 enhances
tumorigenesis and metastasis of breast cancer cells by
downregulating c-Cbl and Cbl-b. Oncotarget. 6:29209–29223. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matouk IJ, Raveh E, Abu-Lail R, Mezan S,
Gilon M, Gershtain E, Birman T, Gallula J, Schneider T, Barkali M,
et al: Oncofetal H19 RNA promotes tumor metastasis. Biochim Biophys
Acta. 1843:1414–1426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang J, Wang X, Chen T, Jiang L and Yang
Q: Huaier extract inhibits breast cancer progression through a
lncRNA-H19/miR-675-5p pathway. Cell Physiol Biochem. 44:581–593.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ebert MS and Sharp PA: Emerging roles for
natural microRNA sponges. Curr Biol. 20:R858–R861. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Franco-Zorrilla JM, Valli A, Todesco M,
Mateos I, Puga MI, Rubio-Somoza I, Leyva A, Weigel D, Garcia JA and
Paz-Ares J: Target mimicry provides a new mechanism for regulation
of microRNA activity. Nat Genet. 39:1033–1037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou W, Ye XL, Xu J, Cao MG, Fang ZY, Li
LY, Guan GH, Liu Q, Qian YH and Xie D: The lncRNA H19 mediates
breast cancer cell plasticity during EMT and MET plasticity by
differentially sponging miR-200b/c and let-7b. Sci Signal. 10(pii):
eaak95572017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Z, Li Y, Li Y, Ren K, Li X, Han X and
Wang J: Long non-coding RNA H19 promotes the proliferation and
invasion of breast cancer through upregulating DNMT1 expression by
sponging miR-152. J Biochem Mol Toxicol. 312017.doi:
10.1002/jbt.21933.
|
|
39
|
Li JP, Xiang Y, Fan LJ, Yao A, Li H and
Liao XH: Long noncoding RNA H19 competitively binds miR-93-5p to
regulate STAT3 expression in breast cancer. J Cell Biochem.
120:3137–3148. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peng F, Li TT, Wang KL, Xiao GQ, Wang JH,
Zhao HD, Kang ZJ, Fan WJ, Zhu LL, Li M, et al: H19/let-7/LIN28
reciprocal negative regulatory circuit promotes breast cancer stem
cell maintenance. Cell Death Dis. 8:e25692017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Peng F, Wang JH, Fan WJ, Meng YT, Li MM,
Li TT, Cui B, Wang HF, Zhao Y, An F, et al: Glycolysis gatekeeper
PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene.
37:1062–1074. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liang R, Li Y, Wang M, Tang SC, Xiao G,
Sun X, Li G, Du N, Liu D and Ren H: MiR-146a promotes the
asymmetric division and inhibits the self-renewal ability of breast
cancer stem-like cells via indirect upregulation of Let-7. Cell
Cycle. 17:1445–1456. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang M, Li Y, Xiao GD, Zheng XQ, Wang JC,
Xu CW, Qin S, Ren H, Tang SC and Sun X: H19 regulation of oestrogen
induction of symmetric division is achieved by antagonizing Let-7c
in breast cancer stem-like cells. Cell Prolif. 52:e125342019.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
De Martino M, Forzati F, Marfella M,
Pellecchia S, Arra C, Terracciano L, Fusco A and Esposito F:
HMGA1P7-pseudogene regulates H19 and Igf2 expression by a
competitive endogenous RNA mechanism. Sci Rep. 6:376222016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu L, Liu L and Lu S: lncRNA H19 promotes
viability and epithelial-mesenchymal transition of lung
adenocarcinoma cells by targeting miR-29b-3p and modifying STAT3.
Int J Oncol. 54:929–941. 2019.PubMed/NCBI
|
|
46
|
Pan Y, Zhang Y, Liu W, Huang Y, Shen X,
Jing R, Pu J, Wang X, Ju S, Cong H and Chen H: LncRNA H19
overexpression induces bortezomib resistance in multiple myeloma by
targeting MCL-1 via miR-29b-3p. Cell Death Dis. 10:1062019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zheng X, Zhou Y, Chen W, Chen L, Lu J, He
F, Li X and Zhao L: Ginsenoside 20(S)-Rg3 prevents PKM2-targeting
miR-324-5p from H19 sponging to antagonize the warburg effect in
ovarian cancer cells. Cell Physiol Biochem. 51:1340–1353. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun Y, Zhu Q, Yang W, Shan Y, Yu Z, Zhang
Q and Wu H: LncRNA H19/miR-194/PFTK1 axis modulates the cell
proliferation and migration of pancreatic cancer. J Cell Biochem.
120:3874–3886. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wei J, Gan Y, Peng D, Jiang X, Kitazawa R,
Xiang Y, Dai Y, Tang Y and Yang J: Long non-coding RNA H19 promotes
TDRG1 expression and cisplatin resistance by sequestering
miRNA-106b-5p in seminoma. Cancer Med. 7:6247–6257. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li M, Chai HF, Peng F, Meng YT, Zhang LZ,
Zhang L, Zou H, Liang QL, Li MM, Mao KG, et al: Estrogen receptor β
upregulated by lncRNA-H19 to promote cancer stem-like properties in
papillary thyroid carcinoma. Cell Death Dis. 9:11202018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wei LQ, Li L, Lu C, Liu J, Chen Y and Wu
H: Involvement of H19/miR-326 axis in hepatocellular carcinoma
development through modulating TWIST1. J Cell Physiol.
234:5153–5162. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li CF, Li YC, Wang Y and Sun LB: The
Effect of LncRNA H19/miR-194-5p axis on the Epithelial-mesenchymal
transition of colorectal adenocarcinoma. Cell Physiol Biochem.
50:196–213. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yan L, Zhou J, Gao Y, Ghazal S, Lu L,
Bellone S, Yang Y, Liu N, Zhao X, Santin AD, et al: Regulation of
tumor cell migration and invasion by the H19/let-7 axis is
antagonized by metformin-induced DNA methylation. Oncogene.
34:3076–3084. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Barsyte-Lovejoy D, Lau SK, Boutros PC,
Khosravi F, Jurisica I, Andrulis IL, Tsao MS and Penn LZ: The c-Myc
oncogene directly induces the H19 noncoding RNA by allele-specific
binding to potentiate tumorigenesis. Cancer Res. 66:5330–5337.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tessier CR, Doyle GA, Clark BA, Pitot HC
and Ross J: Mammary tumor induction in transgenic mice expressing
an RNA-binding protein. Cancer Res. 64:209–214. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu Q, Yang B, Xie X, Wei L, Liu W, Yang
W, Ge Y, Zhu Q, Zhang J, Jiang L, et al: Vigilin interacts with
CCCTC-binding factor (CTCF) and is involved in CTCF-dependent
regulation of the imprinted genes Igf2 and H19. FEBS J.
281:2713–2725. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Stedman W, Kang H, Lin S, Kissil JL,
Bartolomei MS and Lieberman PM: Cohesins localize with CTCF at the
KSHV latency control region and at cellular c-myc and H19/Igf2
insulators. EMBO J. 27:654–666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Popkie AP, Zeidner LC, Albrecht AM,
D'Ippolito A, Eckardt S, Newsom DE, Groden J, Doble BW, Aronow B,
McLaughlin KJ, et al: Phosphatidylinositol 3-kinase (PI3K)
signaling via glycogen synthase kinase-3 (Gsk-3) regulates DNA
methylation of imprinted loci. J Biol Chem. 285:41337–41347. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang K, Luo Z, Zhang Y, Zhang L, Wu L,
Liu L, Yang J, Song X and Liu J: Circulating lncRNA H19 in plasma
as a novel biomarker for breast cancer. Cancer Biomark. 17:187–194.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jiao ZY, Tian Q, Li N, Wang HB and Li KZ:
Plasma long non-coding RNAs (lncRNAs) serve as potential biomarkers
for predicting breast cancer. Eur Rev Med Pharmacol Sci.
22:1994–1999. 2018.PubMed/NCBI
|
|
61
|
Zhang Z, Weaver DL, Olsen D, DeKay J, Peng
Z, Ashikaga T and Evans MF: Long non-coding RNA chromogenic in situ
hybridisation signal pattern correlation with breast tumour
pathology. J Clin Pathol. 69:76–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Shima H, Kida K, Adachi S, Yamada A, Sugae
S, Narui K, Miyagi Y, Nishi M, Ryo A, Murata S, et al: Lnc RNA H19
is associated with poor prognosis in breast cancer patients and
promotes cancer stemness. Breast Cancer Res Treat. 170:507–516.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Riaz M, Berns EM, Sieuwerts AM,
Ruigrok-Ritstier K, de Weerd V, Groenewoud A, Uitterlinden AG, Look
MP, Klijn JG, Sleijfer S, et al: Correlation of breast cancer
susceptibility loci with patient characteristics, metastasis-free
survival, and mRNA expression of the nearest genes. Breast Cancer
Res Treat. 133:843–851. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Han L, Ma P, Liu SM and Zhou X:
Circulating long noncoding RNA GAS5 as a potential biomarker in
breast cancer for assessing the surgical effects. Tumour Biol.
37:6847–6854. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Si X, Zang R, Zhang E, Liu Y, Shi X, Zhang
E, Shao L, Li A, Yang N, Han X, et al: LncRNA H19 confers
chemoresistance in ERα-positive breast cancer through epigenetic
silencing of the pro-apoptotic gene BIK. Oncotarget. 7:81452–81462.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Basak P, Chatterjee S, Weger S, Bruce MC,
Murphy LC and Raouf A: Estrogen regulates luminal progenitor cell
differentiation through H19 gene expression. Endocr Relat Cancer.
22:505–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Basak P, Chatterjee S, Bhat V, Su A, Jin
H, Lee-Wing V, Liu Q, Hu P, Murphy LC and Raouf A: Long non-coding
RNA H19 acts as an estrogen receptor modulator that is required for
endocrine therapy resistance in ER+ breast cancer cells. Cell
Physiol Biochem. 51:1518–1532. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhu QN, Wang G, Guo Y, Peng Y, Zhang R,
Deng JL, Li ZX and Zhu YS: LncRNA H19 is a major mediator of
doxorubicin chemoresistance in breast cancer cells through a
cullin4A-MDR1 pathway. Oncotarget. 8:91990–92003. 2017.PubMed/NCBI
|
|
69
|
Abdel Hadi M: Breast cancer in developing
countries: The shrinking age gap. Breast J. 25:795–797. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Sinaga ES, Ahmad RA, Shivalli S and
Hutajulu SH: Age at diagnosis predicted survival outcome of female
patients with breast cancer at a tertiary hospital in Yogyakarta,
Indonesia. Pan Afr Med J. 31:1632018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Fazel A, Hasanpour-Heidari S, Salamat F,
Rajaie S, Kazeminezhad V, Naeimi-Tabiei M, Jahangirrad A, Sedaghat
S, Hosseinpoor R, Ghasemi-Kebria F, et al: Marked increase in
breast cancer incidence in young women: A 10-year study from
Northern Iran, 2004–2013. Cancer Epidemiol. 62:1015732019.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Kaminska M, Ciszewski T, Łopacka-Szatan K,
Miotla P and Staroslawska E: Breast cancer risk factors. Prz
Menopauzalny. 14:196–202. 2015.PubMed/NCBI
|
|
73
|
Tan BS, Yang MC, Singh S, Chou YC, Chen
HY, Wang MY, Wang YC and Chen RH: LncRNA NORAD is repressed by the
YAP pathway and suppresses lung and breast cancer metastasis by
sequestering S100P. Oncogene. 38:5612–5626. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Cairns J, Ingle JN, Kalari KR, Shepherd
LE, Kubo M, Goetz MP, Weinshilboum RM and Wang L: The lncRNA
MIR2052HG regulates ERα levels and aromatase inhibitor resistance
through LMTK3 by recruiting EGR1. Breast Cancer Res. 21:472019.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhang H, Zhang N, Liu Y, Su P, Liang Y, Li
Y, Wang X, Chen T, Song X, Sang Y, et al: Epigenetic regulation of
NAMPT by NAMPT-AS drives metastatic progression in triple-negative
breast cancer. Cancer Res. 79:3347–3359. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chen F, Chen J, Yang L, Liu J, Zhang X,
Zhang Y, Tu Q, Yin D, Lin D, Wong PP, et al: Extracellular
vesicle-packaged HIF-1α-stabilizing lncRNA from tumour-associated
macrophages regulates aerobic glycolysis of breast cancer cells.
Nat Cell Biol. 21:498–510. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Jin X, Xu XE, Jiang YZ, Liu YR, Sun W, Guo
YJ, Ren YX, Zuo WJ, Hu X, Huang SL, et al: The endogenous
retrovirus-derived long noncoding RNA TROJAN promotes
triple-negative breast cancer progression via ZMYND8 degradation.
Sci Adv. 5:eaat98202019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Dong H, Hu J, Zou K, Ye M, Chen Y, Wu C,
Chen X and Han M: Activation of LncRNA TINCR by H3K27 acetylation
promotes Trastuzumab resistance and epithelial-mesenchymal
transition by targeting MicroRNA-125b in breast Cancer. Mol Cancer.
18:32019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Lu G, Li Y, Ma Y, Lu J, Chen Y, Jiang Q,
Qin Q, Zhao L, Huang Q, Luo Z, et al: Long noncoding RNA LINC00511
contributes to breast cancer tumourigenesis and stemness by
inducing the miR-185-3p/E2F1/Nanog axis. J Exp Clin Cancer Res.
37:2892018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yu G, Zhang W, Zhu L and Xia L:
Upregulated long non-coding RNAs demonstrate promising efficacy for
breast cancer detection: A meta-analysis. Onco Targets Ther.
11:1491–1499. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Ren J, Fu J, Ma T, Yan B, Gao R, An Z and
Wang D: LncRNA H19-elevated LIN28B promotes lung cancer progression
through sequestering miR-196b. Cell Cycle. 17:1372–1380. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Huang Z, Lei W, Hu HB, Zhang H and Zhu Y:
H19 promotes non-small-cell lung cancer (NSCLC) development through
STAT3 signaling via sponging miR-17. J Cell Physiol. 233:6768–6776.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhang Q, Li X, Li X, Li X and Chen Z:
LncRNA H19 promotes epithelial-mesenchymal transition (EMT) by
targeting miR-484 in human lung cancer cells. J Cell Biochem.
119:4447–4457. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wang Q, Cheng N, Li X, Pan H, Li C, Ren S,
Su C, Cai W, Zhao C, Zhang L and Zhou C: Correlation of long
non-coding RNA H19 expression with cisplatin-resistance and
clinical outcome in lung adenocarcinoma. Oncotarget. 8:2558–2567.
2017.PubMed/NCBI
|
|
85
|
Yan J, Zhang Y, She Q, Li X, Peng L, Wang
X, Liu S, Shen X, Zhang W, Dong Y, et al: Long noncoding RNA
H19/miR-675 axis promotes gastric cancer via FADD/caspase 8/caspase
3 signaling pathway. Cell Physiol Biochem. 42:2364–2376. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Ishii S, Yamashita K, Harada H, Ushiku H,
Tanaka T, Nishizawa N, Yokoi K, Washio M, Ema A, Mieno H, et al:
The H19-PEG10/IGF2BP3 axis promotes gastric cancer progression in
patients with high lymph node ratios. Oncotarget. 8:74567–74581.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ma C, Nong K, Zhu H, Wang W, Huang X, Yuan
Z and Ai K: H19 promotes pancreatic cancer metastasis by
derepressing let-7's suppression on its target HMGA2-mediated EMT.
Tumour Biol. 35:9163–9169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ma H, Yuan L, Li W, Xu K and Yang L: The
LncRNA H19/miR-193a-3p axis modifies the radio-resistance and
chemotherapeutic tolerance of hepatocellular carcinoma cells by
targeting PSEN1. J Cell Biochem. 119:8325–8335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Pope C, Mishra S, Russell J, Zhou Q and
Zhong XB: Targeting H19, an imprinted long non-coding RNA, in
hepatic functions and liver diseases. Diseases. 5:E112017.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yang W, Redpath RE, Zhang C and Ning N:
Long non-coding RNA H19 promotes the migration and invasion of
colon cancer cells via MAPK signaling pathway. Oncol Lett.
16:3365–3372. 2018.PubMed/NCBI
|
|
91
|
Yang W, Ning N and Jin X: The lncRNA H19
promotes cell proliferation by competitively binding to miR-200a
and derepressing β-catenin expression in colorectal cancer. Biomed
Res Int. 2017:27674842017.PubMed/NCBI
|
|
92
|
Yang Q, Wang X, Tang C, Chen X and He J:
H19 promotes the migration and invasion of colon cancer by sponging
miR-138 to upregulate the expression of HMGA1. Int J Oncol.
50:1801–1809. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Chen SW, Zhu J, Ma J, Zhang JL, Zuo S,
Chen GW, Wang X, Pan YS, Liu YC and Wang PY: Overexpression of long
non-coding RNA H19 is associated with unfavorable prognosis in
patients with colorectal cancer and increased proliferation and
migration in colon cancer cells. Oncol Lett. 14:2446–2452. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang L, Wang DL and Yu P: LncRNA H19
regulates the expression of its target gene HOXA10 in endometrial
carcinoma through competing with miR-612. Eur Rev Med Pharmacol
Sci. 22:4820–4827. 2018.PubMed/NCBI
|
|
95
|
Zhao L, Li Z, Chen W, Zhai W, Pan J, Pang
H and Li X: H19 promotes endometrial cancer progression by
modulating epithelial-mesenchymal transition. Oncol Lett.
13:363–369. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Ghazal S, McKinnon B, Zhou J, Mueller M,
Men Y, Yang L, Mueller M, Flannery C, Huang Y and Taylor HS: H19
lncRNA alters stromal cell growth via IGF signaling in the
endometrium of women with endometriosis. EMBO Mol Med. 7:996–1003.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhu Z, Xu L, Wan Y, Zhou J, Fu D, Chao H,
Bao K and Zeng T: Inhibition of E-cadherin expression by lnc-RNA
H19 to facilitate bladder cancer metastasis. Cancer Biomark.
22:275–281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lv M, Zhong Z, Huang M, Tian Q, Jiang R
and Chen J: lncRNA H19 regulates epithelial-mesenchymal transition
and metastasis of bladder cancer by miR-29b-3p as competing
endogenous RNA. Biochim Biophys Acta Mol Cell Res. 1864:1887–1899.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Liu C, Chen Z, Fang J, Xu A, Zhang W and
Wang Z: H19-derived miR-675 contributes to bladder cancer cell
proliferation by regulating p53 activation. Tumour Biol.
37:263–270. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Luo M, Li Z, Wang W, Zeng Y, Liu Z and Qiu
J: Upregulated H19 contributes to bladder cancer cell proliferation
by regulating ID2 expression. FEBS J. 280:1709–1716. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ramnarine VR, Alshalalfa M, Mo F, Nabavi
N, Erho N, Takhar M, Shukin R, Brahmbhatt S, Gawronski A, Kobelev
M, et al: The long noncoding RNA landscape of neuroendocrine
prostate cancer and its clinical implications. Gigascience. Jun
1–2018.(Epub ahead of print). doi: 10.1093/gigascience/giy050.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zhu M, Chen Q, Liu X, Sun Q, Zhao X, Deng
R, Wang Y, Huang J, Xu M, Yan J and Yu J: lncRNA H19/miR-675 axis
represses prostate cancer metastasis by targeting TGFBI. FEBS J.
281:3766–3775. 2014. View Article : Google Scholar : PubMed/NCBI
|