Introduction
Urothelial bladder cancer is the 10th most commonly
diagnosed carcinoma, with an estimated 549,000 new cases and
200,000 cancer-related deaths worldwide in 2018 (1). Among newly diagnosed cases, >70% are
non-muscle-invasive bladder cancer, which exhibits a satisfactory
5-year survival rate, but is prone to relapse (2). Up to 10–15% of bladder cancers progress
to muscle invasion, which has a poor prognosis despite treatment,
including radical cystectomy, adjuvant chemotherapy, radiation
therapy and immunotherapy (3).
Cisplatin (CDDP)-based chemotherapy is the standard first-line
treatment for patients with muscle-invasive bladder cancer.
Cisplatin is a platinum-based compound used to treat various
malignancies such as lung, ovarian, head and neck and bladder
cancer (4). However, its therapeutic
efficacy in the clinic is inevitably reduced by intrinsic or
acquired chemoresistance that develops by poorly understood
molecular mechanisms.
Elemene (ELE) (1-methyl-1-vinyl-2,
4-diisopropenyl-cyclohexane) is a novel broad-spectrum antitumor
agent extracted from the traditional Chinese medical herb
Rhizoma zedoariae (5). The
antitumor activity of β-ELE, a class II antitumor drug, has been
evaluated in human solid tumors; combined with other chemotherapy
drugs, β-ELE can reduce the dose of cytotoxic agents and the
occurrence of side effects (6).
β-ELE was approved by the State Food and Drug Administration of
China for the treatment of certain types of cancer, including
malignant brain tumors, in clinical practice (7). β-ELE suppresses tumor activity by
decreasing the mitochondrial membrane potential to promote
apoptosis, causing cell cycle arrest, promoting necrosis and
inhibiting angiogenesis (6,8). β-ELE has also been reported to inhibit
cell proliferation and enhance cisplatin-induced cell death in
bladder cancer, but the mechanisms have not been described
(9,10).
Apoptosis, or programmed cell death, eliminates
defective cells to maintain a balance between cell proliferation
and differentiation (11).
Dysfunction of apoptosis can result in tumor growth and distant
metastasis and may be involved in drug resistance. Apoptosis occurs
by either an extrinsic pathway (mediated by the death receptor) or
an intrinsic pathway (mediated by loss of mitochondrial membrane
potential and translocation of cytochrome c) (11). Generation of reactive oxygen species
(ROS) results in a reduction in the mitochondrial membrane
potential, imbalance of the anti-apoptotic and pro-apoptotic
members of the Bcl-2 family, release of cytochrome c from the
mitochondria to the cytoplasm and activation of the downstream
caspase cascades (12). A previous
study reported that β-ELE promoted platinum intake and
oxaliplatin-induced intrinsic apoptosis in hepatocellular carcinoma
cells (13). The effects of β-ELE
and its mechanism of action with regards to bladder cancer have not
been reported. Hence, the present study investigated the antitumor
activity of β-ELE alone or in combination with cisplatin in
cultured bladder cancer cells.
Materials and methods
Reagents and cell culture
Human bladder cancer cell lines SW780, 5637, J82,
UM-UC-3, TCCSUP and T24 and human ureteral epithelium immortalized
cell line SV-HUC-1 were purchased from the American Type Culture
Collection; RT4 cells were purchased from Procell Life Science
& Technology Co., Ltd. The cells were cultured in RPMI-1640
medium (Corning, Inc.) with 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.), 100 µg/ml penicillin and 100 µg/ml
streptomycin (cat. no. C0222; Beyotime Institute of Biotechnology)
at 37°C with 5% CO2. β-ELE was obtained from Dalian
Holley Kingkong Pharmaceutical Co., Ltd. Monoclonal antibodies
specific for p21 (cat. no. 2947), p27 (cat. no. 3686), cyclin D1
(cat. no. 2978), CDK4 (cat. no. 2906), CDK6 (cat. no. 3136),
phospho-Akt (Thr308) (cat. no. 13038), total Akt (cat. no. 4685),
phospho-STAT3 (cat. no. 9145), total STAT3 (cat. no. 12640), Bax
(cat. no. 5023), Bcl-2 (cat. no. 4223), cytochrome c (cat. no.
11940), caspase-9 (cat. no. 9508), caspase-3 (cat. no. 9662),
cleaved caspase-3 (cat. no. 9664), cleaved caspase-9 (cat. no.
7237), cleaved poly (ADP-ribose) polymerase (PARP; cat. no. 9548),
PARP (cat. no. 9532), phospho-AMPKα (cat. no. 2535) and total AMPK
(cat. no. 5832) were purchased from Cell Signaling Technology, Inc.
N-acetyl-l-cysteine (NAC; ROS scavenger) and compound C (a specific
inhibitor of AMPK) were purchased from Selleck Chemicals. Cell
Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular
Technologies, Inc.
Cell viability
SV-HUC-1 and seven types of bladder cancer cells
(T24, 5637, TCCSUP, J82, UM-UC-3, RT4 and SW780 cells) were seeded
into 96-well plates at density of 5×103 cells/well and
cultured at 37°C with 5% CO2 for 24 h. Following
treatment with β-ELE at varying concentrations (0, 6.25, 12.5, 25,
50, 100, 150 and 200 µg/ml) for 24 h, 10 µl CCK-8 solution was
added into each well. The cells were incubated for 2 h at 37°C.
Absorbance at 450 nm was measured using a Tecan Infinite F200/M200
type multifunction microplate reader (Tecan Group, Ltd.). Cell
viability was calculated as follows: Cell viability=[mean optical
density (OD) of experimental group/mean OD of control group]
×100%.
Assessment of apoptosis by Hoechst
33258 staining
Hoechst 33258 staining (Beyotime Institute of
Biotechnology) was performed according to the manufacturer's
instructions. T24 cells were seeded into 6-well plates at a density
of 5×104 cells/well and cultured at 37°C with 5%
CO2 for 24 h. Cells were treated with cisplatin (20 µM)
and/or β-ELE (50 µg/ml) at 37°C for 24 h, washed three times with
PBS and incubated with Hoechst 33258 (10 µg/ml) for 10 min at room
temperature (RT) in the dark. Morphological changes in nuclei for
the evaluation of apoptosis were observed under a fluorescence
microscope using a blue filter.
Cell cycle analysis
T24 and 5637 cells were seeded into 6-well plates at
density of 5×104 cells/well and cultured at 37°C with 5%
CO2 for 24 h. Cells were treated with different
concentrations of β-ELE (0, 25, 50 and 75 µg/ml) at 37°C for 24 h,
washed twice with PBS and fixed in cold 70% (v/v) ethanol for 48 h
at 4°C. Prior to analysis, cells were washed with PBS, incubated
with 100 µl RNase A (0.1 mg/ml) for 30 min at 37°C, stained with 2
µl propidium iodide (PI; Sigma-Aldrich; Merck KGaA) and incubated
for another 30 min at RT in the dark according to the
manufacturer's instructions. Samples were analyzed by flow
cytometry using CellQuest™ Pro software (version 5.1 BD CellQuest
Pro Software, BD Biosciences).
Flow cytometry for apoptosis
detection
T24 cells were seeded into 6-well plates at density
of 5×104 cells/well and cultured at 37°C with 5%
CO2 for 24 h. Cells were incubated with cisplatin (20
µM) and/or β-ELE (50 µg/ml) for 24 h at 37°C, harvested and washed
with PBS, re-suspended in binding buffer and stained with Annexin
V-FITC and PI (cat. no. C1062M; all Beyotime Institute of
Biotechnology) according to the manufacturer's instructions. The
stained samples were analyzed by flow cytometry (BD Biosciences).
Data were analyzed using CellQuest™ Pro software (version 5.1 BD
CellQuest Pro Software, BD Biosciences).
Western blotting
T24 cells were seeded into 6-well plates at a
density of 5×104 cells/well, and cultured at 37°C with
5% CO2 for 24 h. Cells were pretreated with Compound c
(Selleck Chemicals) for 2 h, and treated with cisplatin (20 µM)
and/or β-ELE (50 µg/ml) for a further 12 h at 37°C. Total protein
of T24 cells were lysed in RIPA buffer (Beyotime Institute of
Biotechnology). Mitochondrial and cytoplasmic components were
isolated using the Cell Mitochondria Isolation kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
instructions. Protein concentration was measured using a BCA kit
(Beyotime Institute of Biotechnology). The prepared protein samples
(40 µg/ml) were loaded and subjected to 12% SDS-PAGE, transferred
to PVDF membranes (EMD Millipore), blocked with 5% skimmed milk for
1.5 h at room temperature and incubated with primary antibodies
overnight at 4°C. Following washing with TBST (10 mmol/l Tris, pH
7.5; 100 mmol/l NaCl; 0.1% Tween-20), the membranes were incubated
with horseradish peroxidase-conjugated goat anti-rabbit (cat. no.
7074; Cell Signaling Technology, Inc.) or goat anti-mouse (cat. no.
7076; Cell Signaling Technology, Inc.) secondary antibody for 1.5 h
at room temperature. Primary antibodies were diluted at 1:1,000 and
secondary antibodies were diluted at 1:3,000. The bands were
detected by enhanced chemiluminescence (EMD Millipore) and
visualized using an ECL system (Pierce Chemical Co.). The results
were quantified using Image J (version 2.0; National Institute of
Health).
Detection of mitochondrial membrane
potential by JC-1 staining
T24 cells were seeded into 6-well plates at a
density of 5×104 cells/well and cultured at 37°C with 5%
CO2 for 24 h. Cells were treated with cisplatin (20 µM)
and/or β-ELE (50 µg/ml) for 12 h at 37°C and stained with a JC-1
probe (Beyotime Institute of Biotechnology) for 20 min at 37°C
according to the manufacturer's instructions. The prepared cells
were observed by fluorescence microscopy and detected by flow
cytometry. Five visual fields were randomly selected for each
sample by fluorescence microscopy (magnification, ×400). Flow
cytometric data were analyzed using CellQuest™ Pro software
(version 5.;1 BD CellQuest Pro Software; BD Biosciences).
Detection of ROS levels by
dichloro-dihydro-fluorescein diacetate (DCFH-DA) staining
T24 cells were seeded into 6-well plates at a
density of 5×104 cells/well and cultured at 37°C with 5%
CO2 for 24 h. Cells were pretreated with
N-acetyl-L-cysteine (NAC, Sigma-Aldrich; Merck KGaA) for 3 h, and
treated with cisplatin (20 µM) and/or β-ELE (50 µg/ml) for another
12 h at 37°C, before being stained with DCFH-DA probe for 20 min at
37°C using the Reactive Oxygen Species Assay kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
instructions. Intracellular production of ROS was evaluated by flow
cytometry using CellQuest™ Pro software (version 5.1; BD CellQuest
Pro; BD Biosciences,).
Statistical analysis
Data are presented as the mean ± SD. Statistical
analysis was performed using SPSS 22.0 software (IBM Corp.).
One-way analysis of variance was used for comparisons among
multiple groups, followed by the Newman-Keuls post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
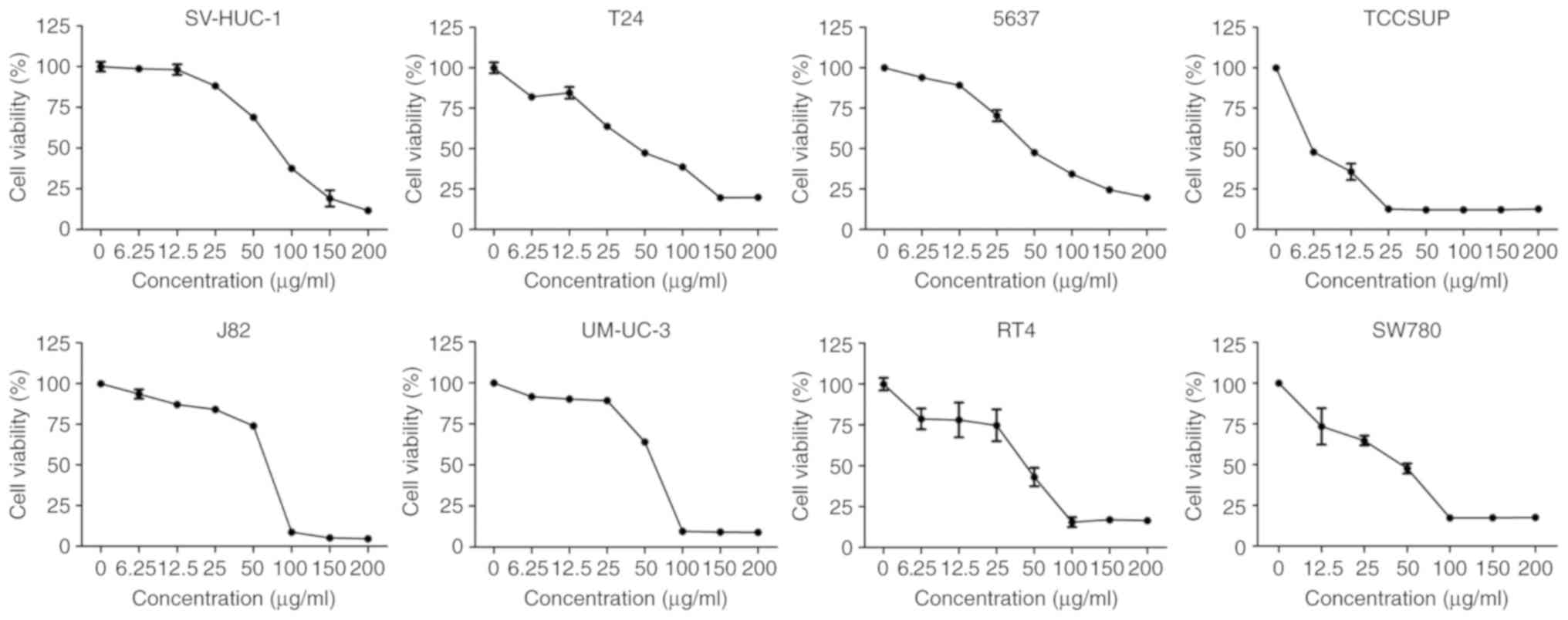
β-ELE inhibits the proliferation of
human bladder cancer cells
Exposure to 0, 6.25, 12.5, 25, 50, 100, 150 and 200
µg/ml β-ELE for 24 h inhibited the proliferation of RT4, SW780,
J82, UMUC-3, TCCSUP, 5637 and T24 human bladder cancer cells and
SV-HUC-1 human urothelial cells. The results of the CCK-8 assay
demonstrated that β-ELE effectively suppressed cell viability of
all tested bladder cancer cell lines (Fig. 1). The half-maximal inhibitory
concentrations (IC50) of β-ELE for each bladder cancer
cell line and SV-HUC-1 cells are presented in Table I. The aforementioned result indicated
that β-ELE had a notable anti-tumor effect on bladder cancer cells.
As T24 and 5637 are the most commonly used and representative cell
line in bladder cancer research, they were both selected for
subsequent experiments.
 | Table I.IC50 of β-elemene in
bladder cancer and normal urothelial cells. |
Table I.
IC50 of β-elemene in
bladder cancer and normal urothelial cells.
| Cell line | IC50,
µg/ml (mean ± SD) |
|---|
| SV-HUC-1 | 72.860±4.128 |
| T24 | 47.403±8.950 |
| 5637 | 61.553±4.070 |
| TCCSUP | 3.661±0.735 |
| J82 | 68.201±0.441 |
| UM-UC-3 | 72.129±1.121 |
| RT4 | 37.894±10.307 |
| SW780 | 37.703±7.323 |
β-ELE induces cell cycle arrest in T24
cells
Regulation of the cell cycle was investigated by
fluorescence microscopy and flow cytometry of T24 cells exposed to
0, 25, 50 or 75 µg/ml β-ELE. Cell numbers began to decrease at a
dose of 50 µg/ml and were notably decreased at 75 µg/ml (Fig. 2A). Flow cytometry revealed that
treatment with β-ELE induced dose-dependent G1-phase
arrest in T24 and 5637 cells and significantly reduced the
percentage of cells in the S-phase (Fig.
2B and C). Western blotting of proteins that modulate cell
survival, cell cycle and cancer progression demonstrated that the
expression levels of p-STAT3 and the cell cycle-related proteins
cyclin D1, CDK4 and CDK6 were downregulated, and p21 and p27were
upregulated by β-ELE. The phosphorylation of Akt (Thr 308) was
reduced by 75 µg/ml β-ELE (Fig.
2D).
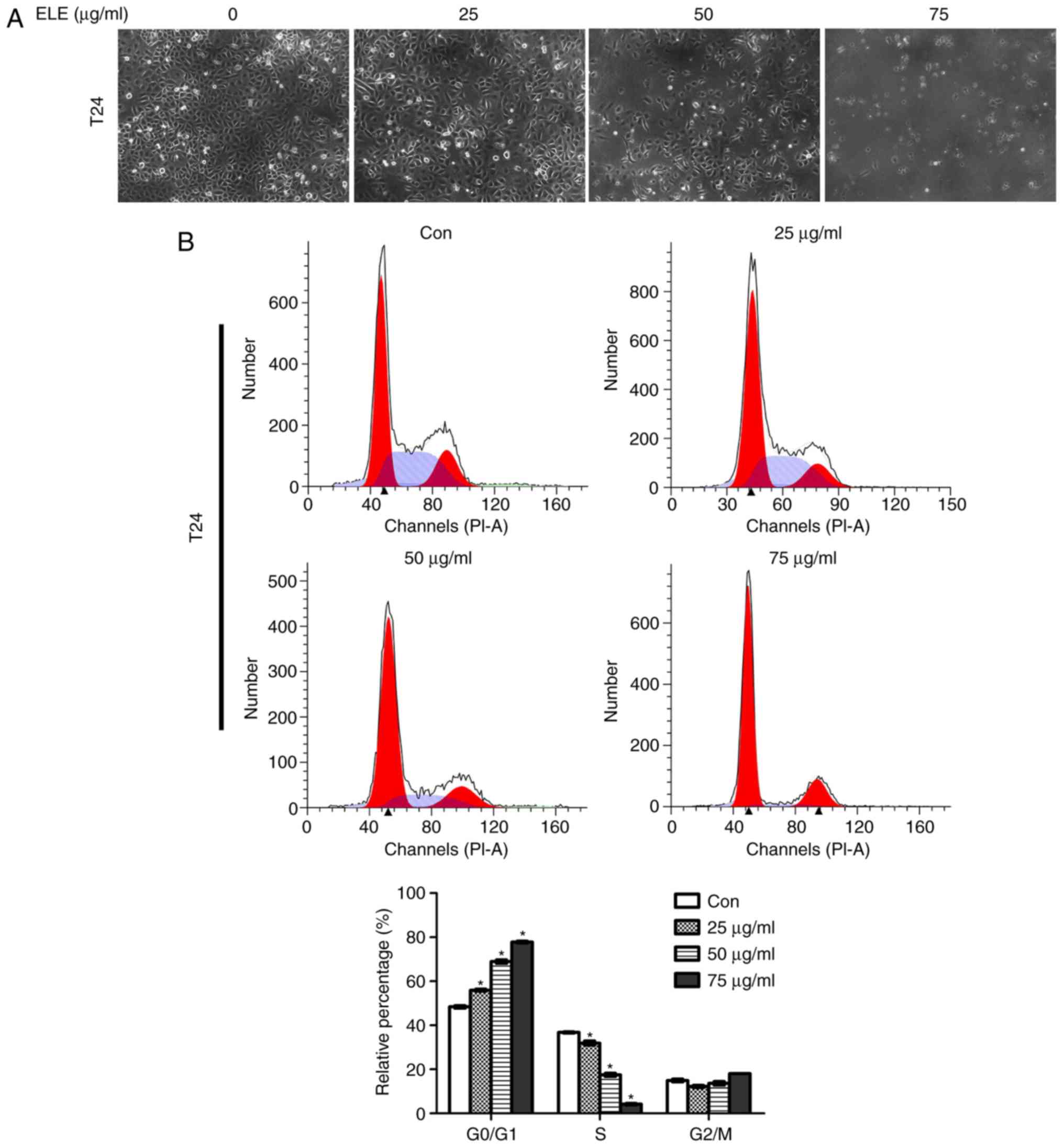 | Figure 2.β-ELE induces
G0/G1 cell cycle arrest in T24 and 5637
cells. (A) T24 cells were counted using an inverted microscope
following treatment with 0, 25, 50 or 75 µg/ml β-ELE for 24 h. (B
and C) The percentages of (B) T24 and (C) 5637 cells in
G0/G1, S and G2/M phase following
treatment with 0, 25, 50 or 75 µg/ml β-ELE for 24 h were determined
by flow cytometry. Data are presented as the mean ± SD, n=3.
*P<0.05 vs. control. (D) Western blot analysis of the expression
of cell cycle-related proteins and signaling molecules in T24 cells
following treatment with 0, 25, 50 and 75 µg/ml β-ELE for 24 h.
Con, control; β-ELE, β-elemene. |
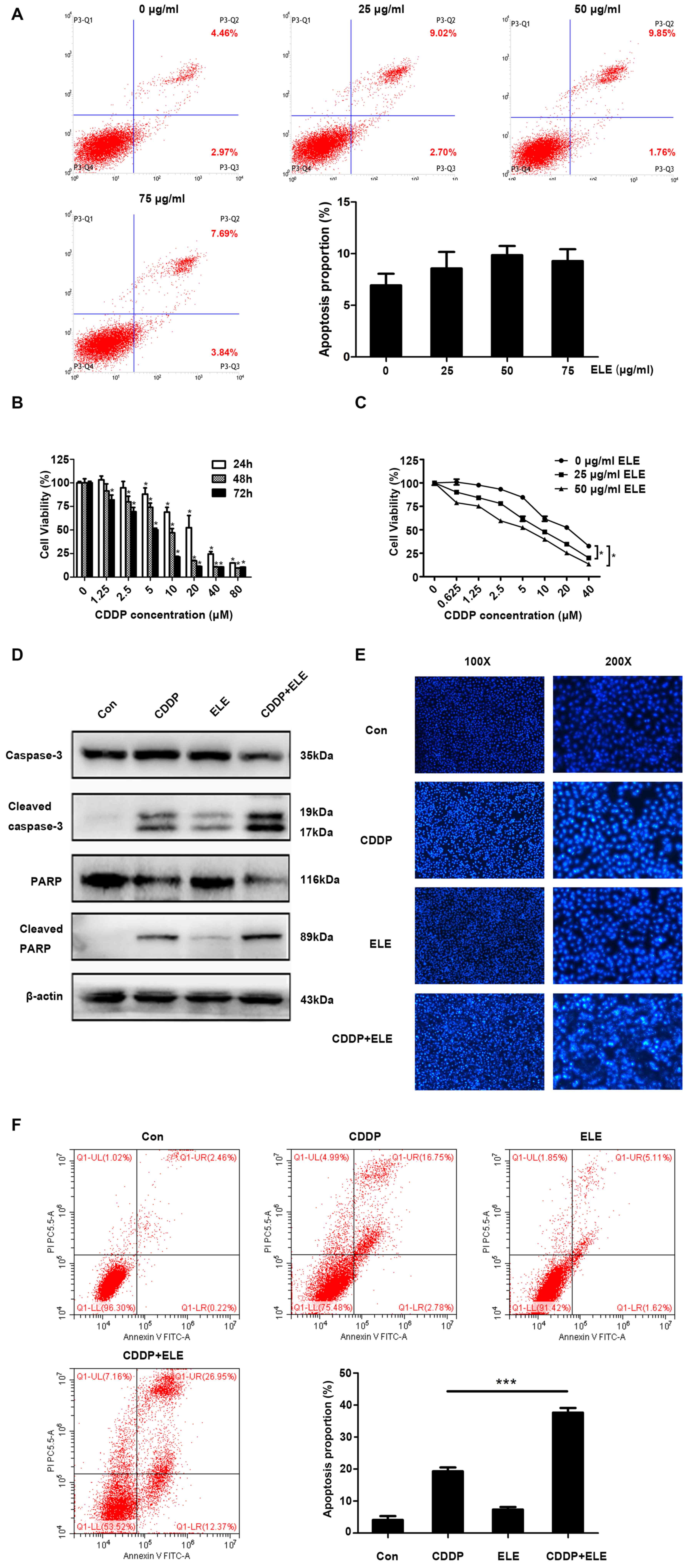
β-ELE potentiates cisplatin-induced
apoptosis in T24 cells
β-ELE was previously demonstrated to promote
apoptosis in T24 and 5637 human bladder carcinoma and primary
bladder cancer cells (14). In the
present study, β-ELE only induced a small increase in the apoptosis
of T24 cells (Fig. 3A). Cisplatin
significantly suppressed T24 cell viability in a time- and
dose-dependent manner (Fig. 3B). The
IC50 values were 20.07±2.18 µM at 24 h, 8.68±1.31 µM at
48 h and 4.63±0.40 µM at 72 h. The additive cytotoxicity of low (25
µg/ml) and moderate (50 µg/ml) dose β-ELE and cisplatin was
stronger compared with that of cisplatin treatment alone at 24 h
(Fig. 3C). Thus, 20 µM cisplatin and
50 µg/ml β-ELE were used in subsequent experiments. Western
blotting results revealed that caspase-3 and PARP cleavage was
increased by cisplatin combined with β-ELE compared with either
β-ELE of cisplatin alone (Fig. 3D).
Fluorescence microscopy of T24 cells stained with Hoechst 33258
(Fig. 3E) indicated the compounded
effect of cotreatment with cisplatin and β-ELE, compared with
either compound alone, which was consistent with the results of the
flow cytometry apoptosis assay of Annexin-V stained cells (Fig. 3F).
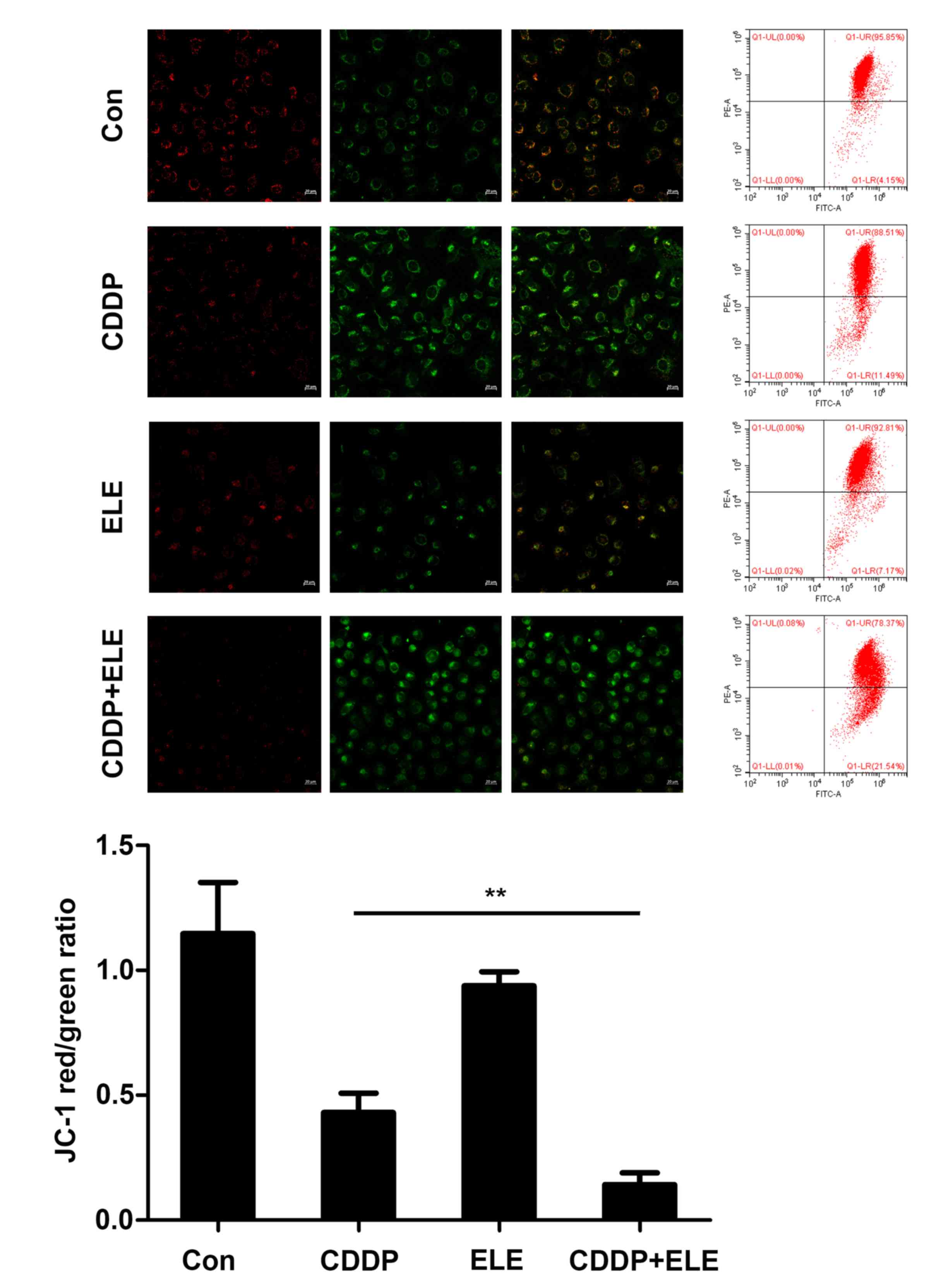
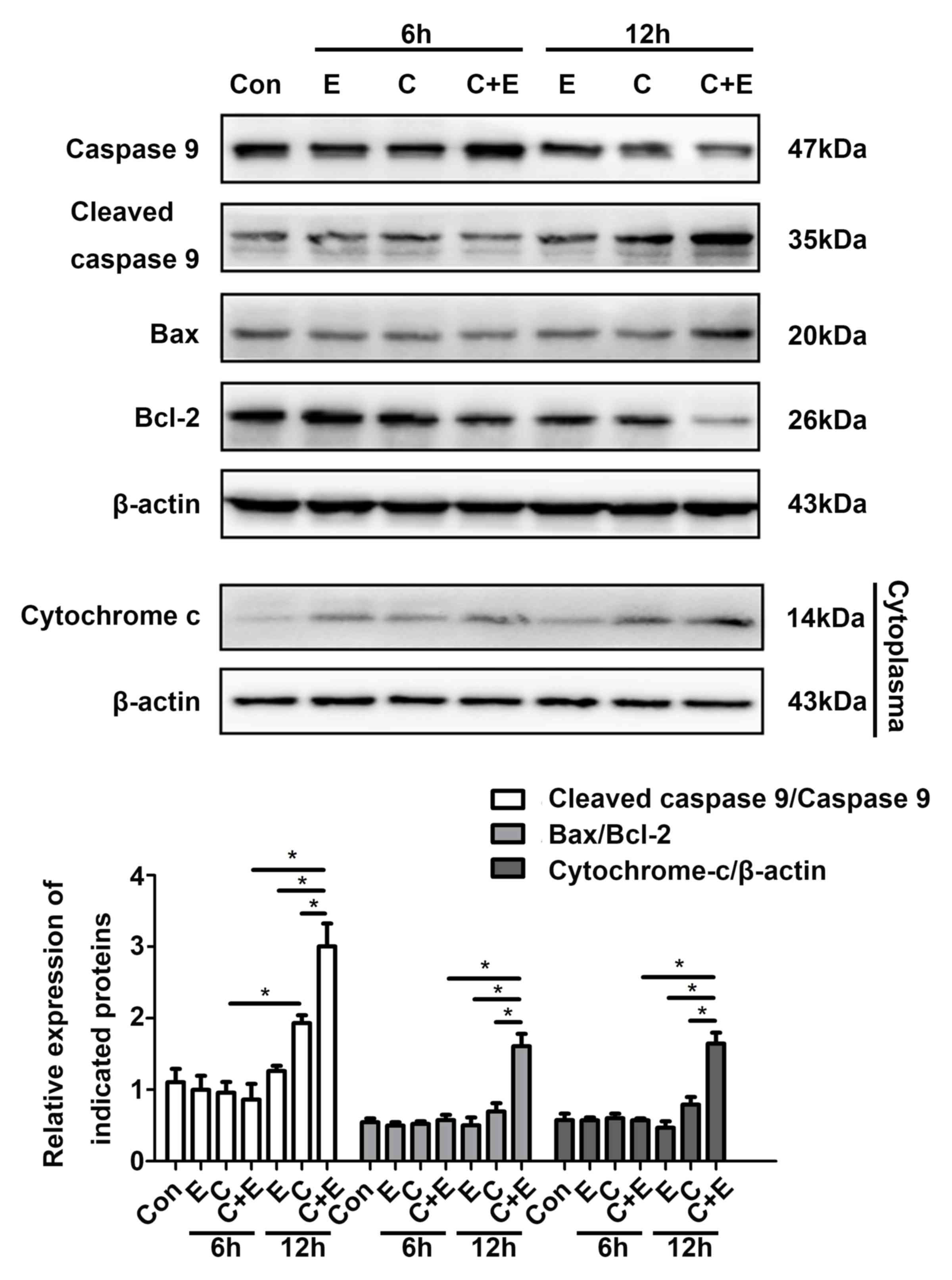
β-ELE promotes cisplatin-induced
apoptosis via the mitochondrial pathway
The augmentation of cisplatin cytotoxicity by β-ELE
in cisplatin-resistant ovarian cancer cells has been demonstrated
to be mediated in part by mitochondria-dependent apoptosis
(15). In the present study,
apoptotic T24 cell death induced by cisplatin and β-ELE was
determined by the change in mitochondrial membrane potential
(ΔΨm) indicated by the JC-1 fluorescent probe. The decrease
in the red/green fluorescence ratio observed by fluorescence
microscopy or flow cytometry was significantly larger in cells
co-treated with cisplatin and β-ELE compared with those treated
with cisplatin or β-ELE alone, and results of flow cytometry were
statistically analyzed and shown in the bar graph (Fig. 4). Western blotting revealed
upregulated expression of cleaved caspase-9, Bax and cytoplasmic
cytochrome c, as well as downregulated expression of Bcl-2
following cisplatin and β-ELE co-treatment for 12 h compared with
single drug treatment group (Fig.
5). Thus, the additive effect of β-ELE on cisplatin-induced
apoptosis in T24 cells may be associated with the activation of the
mitochondrial apoptosis pathway.
β-ELE enhances cisplatin-induced
apoptosis by activating the ROS-AMPK signaling pathway
ROS generation and the activation of ROS-dependent
pathways were associated with the induction of apoptosis in bladder
cancer cells co-treated with cisplatin and curcumin (16). The involvement of ROS-dependent
pathways in the regulation of apoptosis by co-treatment with
cisplatin and β-ELE was evaluated by DCFH-DA assay of intracellular
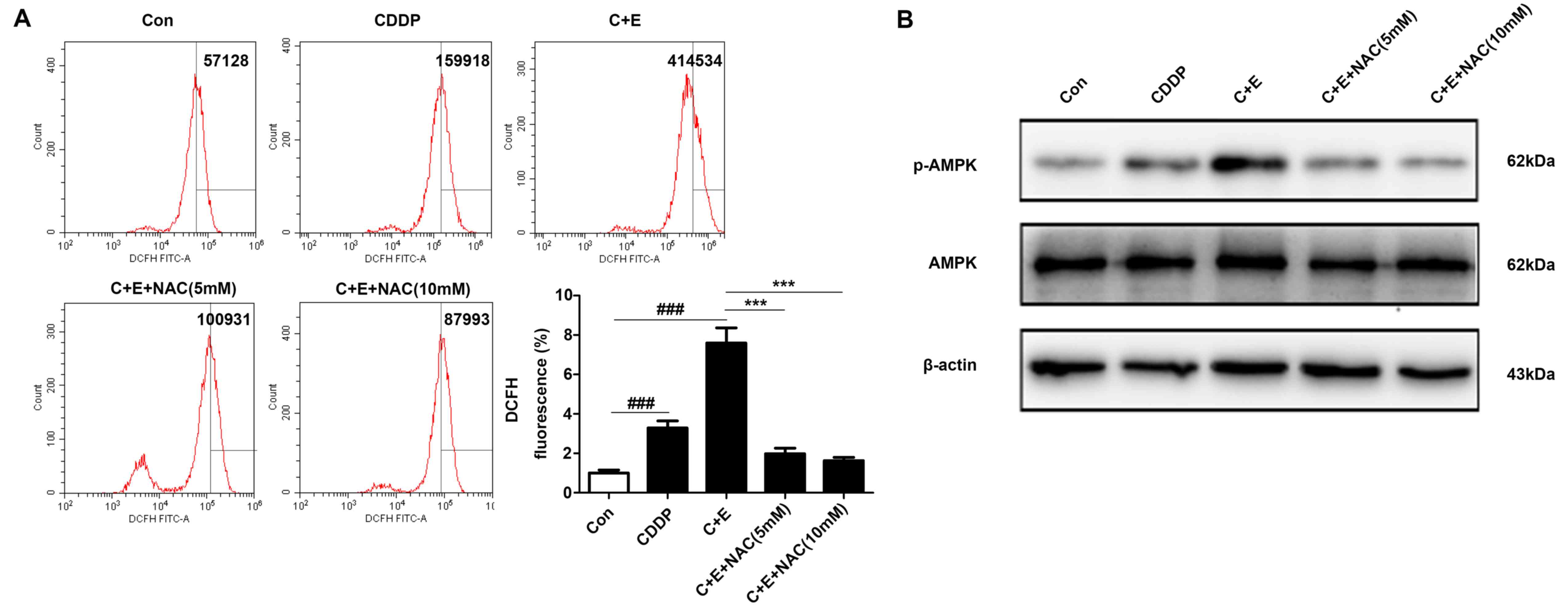
ROS in the present study. Flow cytometry results demonstrated that
the intracellular ROS levels were significantly higher in cells
co-treated with cisplatin and β-ELE compared with those in cells
treated with cisplatin alone; pre-treatment with 5 or 10 mM NAC
reversed ROS accumulation induced by the addition of cisplatin and
β-ELE (Fig. 6A). Western blotting
results demonstrated that AMPK phophorylation was increased by
cisplatin and β-ELE co-treatment and reversed by NAC pretreatment
(Fig. 6B). Considering the
complicated roles of the ROS-AMPK signaling pathway in maintaining
redox homeostasis and cell survival (17), it was speculated in the present study
that the ROS-mediated AMPK activation may contribute to regulating
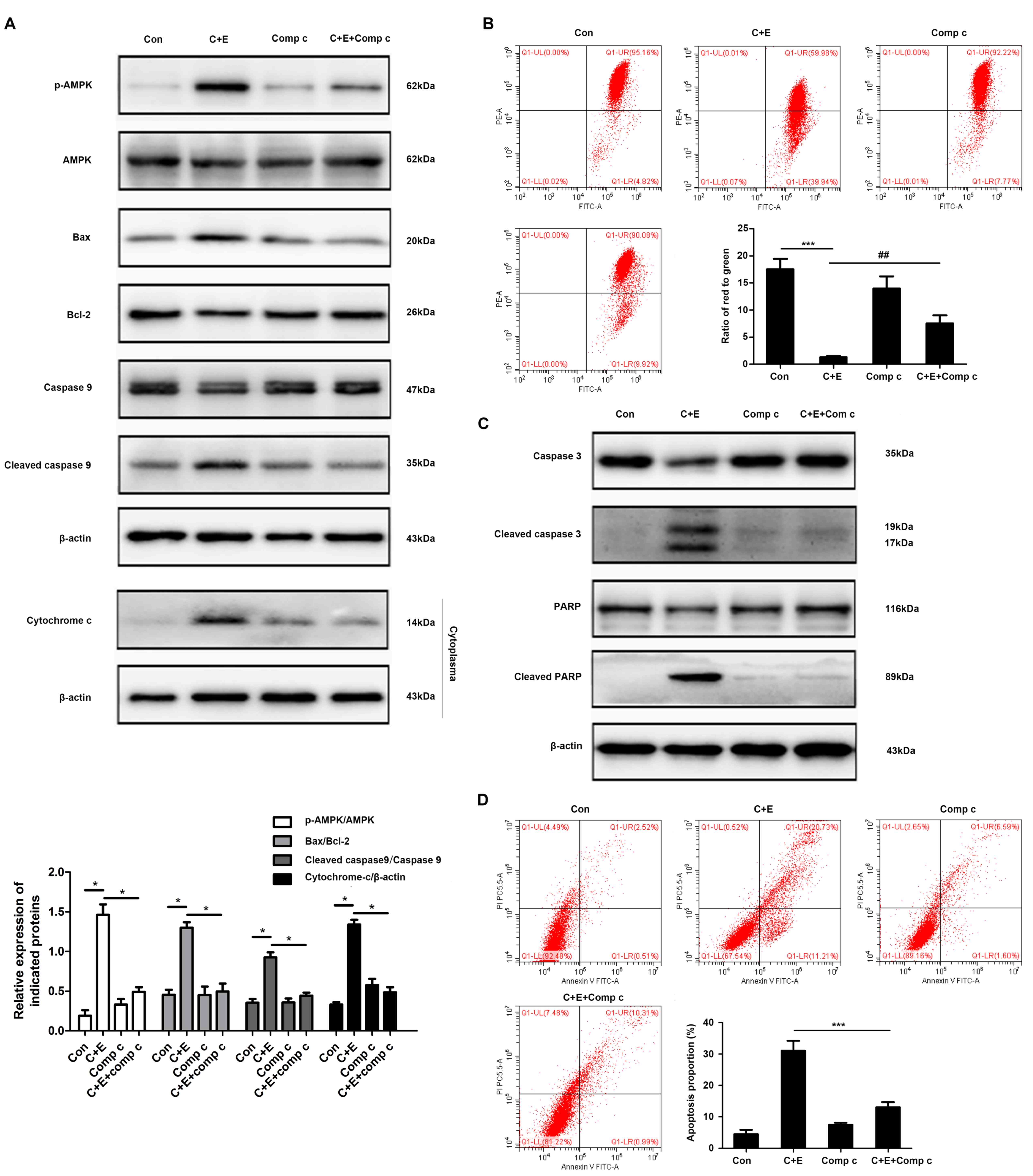
apoptosis via the mitochondrial pathway. T24 cells were pretreated
with 5 or 10 µM compound C, a specific inhibitor against AMPK, for
2 h and treated with cisplatin and β-ELE for 12 h. Pretreatment
with compound C reversed the effects of cisplatin and β-ELE
co-treatment on the expression of mitochondrial apoptosis-related
proteins, including Bax, Bcl-2, cleaved caspase-9 and cytoplasmic
cytochrome-c (Fig. 7A), as well as
the loss of mitochondrial membrane potential (Fig. 7B). In addition, the inhibition of
AMPK by compound C attenuated the levels of caspase-3 and PARP
cleavage (Fig. 7C) and apoptotic
rate (Fig. 7D) compared with
cisplatin and β-ELE co-treatment. These results suggested that the
ROS-AMPK signaling pathway may be involved in the intrinsic
apoptosis induced by cisplatin and β-ELE co-treatment.
Discussion
Cisplatin-based regimens are currently the standard
treatment of advanced bladder cancer, but the 5-year survival rate
of patients with muscle-invasive bladder cancer is <60%
(2). The effectiveness of cisplatin
is reduced by treatment-related side effects and the development of
chemoresistance (18). Traditional
Chinese medical herbs with anticancer activity, low toxicity and
low rates of adverse effects can be combined with chemotherapeutic
agents to improve clinical treatment response (6). β-ELE has previously been demonstrated
to synergistically enhance cisplatin cytotoxicity in drug-resistant
ovarian cancer cells by activating mitochondria-dependent signaling
(15). β-ELE has also been reported
to induce apoptosis in non-small cell lung cancer cells by
triggering endoplasmic reticulum (ER) stress, activating the
PRKR-like ER kinase/Ire1-α/activating transcription factor 6
pathway and downregulating Bcl-2 expression (19). In addition, β-ELE can induce
different types of cell cycle arrest, including
G0/G1, G2/M and S phase arrest,
depending on the tumor type. For example, β-ELE reduced the
expression of α-tubulin and interrupted microtubular polymerization
to induce G2/M and S phase arrest in human liver cancer
HepG2 cells. In addition, β-ELE can directly regulate the activity
of CDK and cell cycle checkpoints to suppress tumor growth
(20). β-ELE has been demonstrated
to induce a persistent arrest at the G2/M phase in human
platinum-sensitive and -resistant ovarian carcinoma cells by
reducing the expression of CDK1, cyclin A and cyclin B1 and
increasing that of p53 and p21, which are vital regulatory proteins
that target cell cycle progression (21). By contrast, β-ELE increased the
proportion of cells at the G0/G1 phase in a
time- and dose-dependent manner in human nasopharyngeal carcinoma
by inactivating phospho-STAT3, a transcription factor that
participates in tumor cell growth and progression, and inhibiting
the protein expression of DNA methyltransferase 1 (DNMT1) and
enhancer of zeste homolog 2 (22).
In the present study, β-ELE induced G0/G1
phase arrest in T24 and 5637 cells, which may have resulted from
β-ELE-induced inactivation of phospho-AKT and phospho-STAT3, which
are closely associated with tumor cell survival and cancer
progression (23). A similar
previous study demonstrated that the anticancer activity of β-ELE
in T24 and 5637 bladder cancer cells was modulated by the PTEN and
AKT pathway (14), which is in
agreement with the results of the present study. The mechanisms of
action of β-ELE used in combination with chemotherapeutic agents
have not been clearly described. β-ELE has been demonstrated to
enhance the radiosensitivity and temozolomide sensitivity of
glioblastoma cells through the inhibition of the ataxia
telangiectasia mutated-AKT and extracellular-signal-regulated
kinase (ERK) signaling pathways (24). In addition, β-ELE has been reported
to augment the cisplatin-induced inhibition of proliferation and
promotion of apoptosis in gingival squamous cell carcinoma by
blocking the activation of the JAK2-STAT3 signaling pathway
(25). In MCF-7 breast cancer cells,
β-ELE reversed Adriacin and docetaxel resistance by altering the
expression of multidrug resistance-specific microRNAs, increasing
PTEN and decreasing phosphoglycolate phosphatase gene expression in
exosomes (26). Another study
reported that β-ELE mediated an increase in cisplatin response of
platinum-resistant ovarian cancer cells by inducing
caspase-dependent apoptosis, decreasing mitochondrial transmembrane
potential and releasing cytochrome c into the cytoplasm (15). In the present study, intrinsic
apoptosis was triggered by the combined effects of cisplatin and
β-ELE on mitochondrial membrane potential, increased cytosolic
cytochrome c, increased Bax and cleaved caspase-9 expression and
decreased Bcl-2 expression.
The effects of ROS on cell metabolism, homeostasis
and survival, regulation of immune responses, induction of
autophagy, apoptosis and the chemosensitivity of cancer cells are
well-described (27,28). ROS is a positive regulator of AMPK,
which activates glucose and fatty acid uptake and metabolism.
Activation of AMPK phosphorylation by exogenous
H2O2 is mediated by liver kinase B1 or
calcium/calmodulin-dependent protein kinase β and an increase in
the AMP/ATP ratio (29,30). An increase in ROS and AMPK
phosphorylation in response to hypoxia can be abolished by
antioxidants such as EUK-134. Hypoxia does not activate AMPK in
cells deficient in mitochondrial DNA, with the exception of the
presence of exogenous H2O2 (31). In addition, AMPK signaling is
associated with increased cell survival. Activation of AMPKα and
ERK1/2 by β-ELE is associated with the suppression of transcription
factor Sp1 and DNMT1 protein expression, as well as the inhibition
of non-small cell lung carcinoma cell proliferation in vitro
(32). The blocking of AMPK activity
by compound C or AMPK-targeting small interfering RNA inhibited
hispidulin-induced ER stress and AMPK-mTOR signaling-promoted
apoptosis in hepatocellular carcinoma cells (33). Metformin can induce apoptosis by
activating the phosphorylation of AMPK at Thr172, and decreased
phospho-AMPK expression has been demonstrated to reverse the
inhibition of viability of gastric adenocarcinoma cells by
metformin (34). In the present
study, β-ELE promoted cisplatin-induced apoptosis accompanied by
accumulation of ROS and upregulation of phospho-AMPKα. The
apoptosis induced by co-treatment with β-ELE and cisplatin was
reversed by an ROS scavenger and an AMPK inhibitor.
The present study mainly focused on the role of
β-ELE in increasing the sensitivity of bladder cancer cells to
cisplatin and it was revealed that the ROS-AMPK pathway-mediated
mitochondrial dysfunction is considered to be involved in this
process. Since β-ELE improved cell chemosensitivity to cisplatin,
it may alleviate or reverse chemotherapy resistance by altering of
the expression of multidrug resistance (MDR)-associated genes and
proteins. In addition, a recent study revealed that β-ELE may
repress the MDR process by inhibiting the expression of ATP-binding
cassette transporters, such as P-glycoprotein (P-gp) and breast
cancer resistance protein, which can pump chemotherapeutic drugs
outside of cancer cells, leading to chemoresistance (35). β-ELE also regulates the expression of
certain microRNAs (miRs; miR-34a, miR-222, miR-452 and miR-29a)
that bind to the 3′-untranslated region of PTEN and P-gp to
attenuate MDR (35). Drug-resistant
bladder cancer cells exhibiting high expression levels of
MDR-related genes were not available in the present study to test
whether this process occurred in bladder cancer. Further specific
studies on MDR in bladder cancer are needed in the future.
In conclusion, the results of the present study
demonstrated that β-ELE suppressed the proliferation of bladder
cancer cells in vitro and induced
G0/G1 phase arrest in T24 and 5637 cells,
which may have involved the AKT and STAT3 signaling pathways. β-ELE
also enhanced cisplatin-induced mitochondrial apoptosis by
activating the ROS-AMPK pathway. β-ELE activity may offer benefits
that improve the response of bladder cancer to chemotherapeutic
agents and reverse chemoresistance.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China (grant no. 81874092 and 81801482) and
Chongqing Science & Technology Commission (grant no.
cstc2019jcyi-msxmX0126).
Availability of data and materials
The datasets generated and analyzed during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
DG designed and performed the experiments and wrote
the manuscript. HY analyzed and organized the data and revised the
manuscript. XG and WH assisted with the study design and obtained
funding.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yin H, He W, Li Y, Xu N, Zhu X, Lin Y and
Gou X: Loss of DUSP2 predicts a poor prognosis in patients with
bladder cancer. Hum Pathol. 85:152–161. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lamm D, Persad R, Brausi M, Buckley R,
Witjes JA, Palou J, Böhle A, Kamat AM, Colombel M and Soloway M:
Defining progression in nonmuscle invasive bladder cancer: It is
time for a new, standard definition. J Urol. 191:20–27. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tan W, Lu J, Huang M, Li Y, Chen M, Wu G,
Gong J, Zhong Z, Xu Z, Dang Y, et al: Anti-cancer natural products
isolated from chinese medicinal herbs. Chin Med. 6:272011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhai B, Zeng Y, Zeng Z, Zhang N, Li C,
Zeng Y, You Y, Wang S, Chen X, Sui X and Xie T: Drug delivery
systems for elemene, its main active ingredient β-elemene, and its
derivatives in cancer therapy. Int J Nanomedicine. 13:6279–6296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu J, Zhang Y, Qu J, Xu L, Hou K, Zhang
J, Qu X and Liu Y: β-Elemene-induced autophagy protects human
gastric cancer cells from undergoing apoptosis. BMC Cancer.
11:1832011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang Z, Jacob JA, Loganathachetti DS,
Nainangu P and Chen B: β-elemene: Mechanistic studies on cancer
cell interaction and its chemosensitization effect. Front
Pharmacol. 8:1052017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu X, Wang Y, Luo H, Qiu W, Han H, Chen X
and Yang L: β-elemene inhibits the proliferation of T24 bladder
carcinoma cells through upregulation of the expression of Smad4.
Mol Med Rep. 7:513–518. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li QQ, Wang G, Liang H, Li JM, Huang F,
Agarwal PK, Zhong Y and Reed E: β-elemene promotes
cisplatin-induced cell death in human bladder cancer and other
carcinomas. Anticancer Res. 33:1421–1428. 2013.PubMed/NCBI
|
|
11
|
Koosha S, Mohamed Z, Sinniah A, Ibrahim
ZA, Seyedan A and Alshawsh MA: Antiproliferative and apoptotic
activities of 8-prenylnaringenin against human colon cancer cells.
Life Sci. 232:1166332019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goldar S, Khaniani MS, Derakhshan SM and
Baradaran B: Molecular mechanisms of apoptosis and roles in cancer
development and treatment. Asian Pac J Cancer Prev. 16:2129–2144.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X, Lin Z, Zhang B, Guo L, Liu S, Li H,
Zhang J and Ye Q: β-elemene sensitizes hepatocellular carcinoma
cells to oxaliplatin by preventing oxaliplatin-induced degradation
of copper transporter 1. Sci Rep. 6:210102016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cai B, Ma L, Nong S, Wu Y, Guo X and Pu J:
β-elemene induced anticancer effect in bladder cancer through
upregulation of PTEN and suppression of AKT phosphorylation. Oncol
Lett. 16:6019–6025. 2018.PubMed/NCBI
|
|
15
|
Li QQ, Lee RX, Liang H, Zhong Y and Reed
E: Enhancement of cisplatin-induced apoptosis by β-elemene in
resistant human ovarian cancer cells. Med Oncol. 30:4242013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park BH, Lim JE, Jeon HG, Seo SI, Lee HM,
Choi HY, Jeon SS and Jeong BC: Curcumin potentiates antitumor
activity of cisplatin in bladder cancer cell lines via ROS-mediated
activation of ERK1/2. Oncotarget. 7:63870–63886. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren Y and Shen HM: Critical role of AMPK
in redox regulation under glucose starvation. Redox Biol.
1011542019.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
von der Maase H, Sengelov L, Roberts JT,
Ricci S, Dogliotti L, Oliver T, Moore MJ, Zimmermann A and Arning
M: Long-term survival results of a randomized trial comparing
gemcitabine plus cisplatin, with methotrexate, vinblastine,
doxorubicin, plus cisplatin in patients with bladder cancer. J Clin
Oncol. 23:4602–4608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Jiang ZY, Zhou YL, Qiu HH, Wang G,
Luo Y, Liu JB, Liu XW, Bu WQ, Song J, et al: β-elemene regulates
endoplasmic reticulum stress to induce the apoptosis of NSCLC cells
through PERK/IRE1α/ATF6 pathway. Biomed Pharmacother. 93:490–497.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mao Y, Zhang J, Hou L and Cui X: The
effect of beta-elemene on alpha-tubulin polymerization in human
hepatoma HepG2 cells. Chin J Cancer Res. 25:770–776.
2013.PubMed/NCBI
|
|
21
|
Lee RX, Li QQ and Reed E: β-elemene
effectively suppresses the growth and survival of both
platinum-sensitive and -resistant ovarian tumor cells. Anticancer
Res. 32:3103–3113. 2012.PubMed/NCBI
|
|
22
|
Wu J, Tang Q, Yang L, Chen Y, Zheng F and
Hann SS: Interplay of DNA methyltransferase 1 and EZH2 through
inactivation of Stat3 contributes to β-elemene-inhibited growth of
nasopharyngeal carcinoma cells. Sci Rep. 7:5092017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yao S, Xu J, Zhao K, Song P, Yan Q, Fan W,
Li W and Lu C: Down-regulation of HPGD by miR-146b-3p promotes
cervical cancer cell proliferation, migration and
anchorage-independent growth through activation of STAT3 and AKT
pathways. Cell Death Dis. 9:10552018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu S, Zhou L, Zhao Y and Yuan Y:
β-elemene enhances both radiosensitivity and chemosensitivity of
glioblastoma cells through the inhibition of the ATM signaling
pathway. Oncol Rep. 34:943–951. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang C and Yu Y: Synergistic cytotoxicity
of β-elemene and cisplatin in gingival squamous cell carcinoma by
inhibition of STAT3 signaling pathway. Med Sci Monit. 23:1507–1513.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang J, Zhang HD, Yao YF, Zhong SL, Zhao
JH and Tang JH: β-elemene reverses chemoresistance of breast cancer
cells by reducing resistance transmission via exosomes. Cell
Physiol Biochem. 36:2274–2286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kruk J, Aboul-Enein HY, Kładna A and
Bowser JE: Oxidative stress in biological systems and its relation
with pathophysiological functions: The effect of physical activity
on cellular redox homeostasis. Free Radic Res. 53:497–521. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li X, Wang H, Wang J, Chen Y, Yin X, Shi
G, Li H, Hu Z and Liang X: Emodin enhances cisplatin-induced
cytotoxicity in human bladder cancer cells through ROS elevation
and MRP1 downregulation. BMC Cancer. 16:5782016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woods A, Dickerson K, Heath R, Hong SP,
Momcilovic M, Johnstone SR, Carlson M and Carling D:
Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream
of AMP-activated protein kinase in mammalian cells. Cell Metab.
2:21–33. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Emerling BM, Weinberg F, Snyder C, Burgess
Z, Mutlu GM, Viollet B, Budinger GR and Chandel NS: Hypoxic
activation of AMPK is dependent on mitochondrial ROS but
independent of an increase in AMP/ATP ratio. Free Radic Biol Med.
46:1386–1391. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao S, Wu J, Zheng F, Tang Q, Yang L, Li
L, Wu W and Hann SS: β-elemene inhibited expression of DNA
methyltransferase 1 through activation of ERK1/2 and AMPKα
signalling pathways in human lung cancer cells: The role of Sp1. J
Cell Mol Med. 19:630–641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han M, Gao H, Xie J, Yuan YP, Yuan Q, Gao
MQ, Liu KL, Chen XH, Han YT and Han ZW: Hispidulin induces ER
stress-mediated apoptosis in human hepatocellular carcinoma cells
in vitro and in vivo by activating AMPK signaling pathway. Acta
Pharmacol Sin. 40:666–676. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lu CC, Chiang JH, Tsai FJ, Hsu YM, Juan
YN, Yang JS and Chiu HY: Metformin triggers the intrinsic apoptotic
response in human AGS gastric adenocarcinoma cells by activating
AMPK and suppressing mTOR/AKT signaling. Int J Oncol. 54:1271–1281.
2019.PubMed/NCBI
|
|
35
|
Zhai B, Zhang N, Han X, Li Q, Zhang M,
Chen X, Li G, Zhang R, Chen P, Wang W, et al: Molecular targets of
β-elemene, a herbal extract used in traditional Chinese medicine,
and its potential role in cancer therapy: A review. Biomed
Pharmacother. 114:1088122019. View Article : Google Scholar : PubMed/NCBI
|