Introduction
Gastric cancer (GC) is a common life-threatening
cancer type worldwide, with an increasing prevalence and high rate
of mortality, particularly in China (1). Etiological factors, and genetic and
epigenetic factors are all involved in the progression of GC
(2). At present, surgery is the most
effective treatment for GC, and the survival rate primarily depends
on the stage of GC (non-invasive detection of GC) (3). Therefore, identifying specific and
effective biomarkers in the early stage of GC is important for the
efficient treatment of patients with GC in order to increase the
survival rate. Previous studies have demonstrated that genetic
abnormalities, including aberrant genes, microRNAs (miRNAs) and
long non-coding RNAs (lncRNAs), participated in the initiation and
progression of GC (competing endogenous RNA networks) (4).
Long non-coding RNAs are defined as transcripts
>200 nucleotides, and are widely expressed in different tissues
(5,6).
lncRNAs are involved in a number of biological progresses,
including the proliferation and differentiation of cells (7). lncRNA genes resemble protein-coding
genes, and they have a variety of molecular functions across
numerous cellular pathways and processes, including oncogenic
signaling (8). Researchers have
identified an increasing number of aberrantly expressed lncRNAs in
GC, and have aimed to determine their role in the progression of GC
(9–12).
Epigenetic modifications serve causative roles in
the progression of GC (13). Histone
deacetylases (HDACs), a family of 18 molecules divided into four
sub-classes defined according to their structural similarities
(14), serve functional roles,
including modification of histone and non-histone proteins,
regulation of chromatin structure, repression or activation of gene
expression and regulation of cell metabolism (14). Previous studies have demonstrated that
HDACs are crucial for the progression of several types of disease,
including diabetes, melanoma, hepatocellular carcinoma and GC
(15–18). However, the precise role of HDACs in
the progression of GC remains unclear. HDACs have been demonstrated
to serve roles in regulating the expression of microRNAs. However,
little is known regarding whether HDACs may serve certain roles in
regulating the expression of lncRNAs during the progression of
GC.
The present study attempted to investigate the role
of abnormal expression of HDACs in the progression of GC, and to
determine the association between HDACs and lncRNAs. The present
study provided evidence of the importance of HDACs in the
progression of GC and identified them as promising targets for the
clinical treatment of GC.
Patients and methods
Patient population
Gastric cancer tissues from 63 patients (mean age 61
years, range 45–73 years; 45 males and 18 females) with gastric
cancer and normal tissues from 31 healthy individuals (mean age 58
years, range 40–75 years; 20 male and 11 female) were used in the
present study, which was obtained from Jiangxi Province People's
Hospital during the period from March 2015 to August 2016. Ethics
approval for this study was approved by the Institutional Review
Board and the Ethics Committee at Jiangxi Province People's
Hospital (ID no. 2015083). Written informed consent was obtained
from each patient. Ten cancer tissues and normal tissues were
randomly selected for examining the expression profile of lncRNAs
in the GSE64951 database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE64951).
Additionally, 9 cancer tissues and normal tissues were randomly
selected for examining the expression profile of lncRNAs in the
GSE19826 database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi). The
expression of lncRNAs in the tissues of patients with gastric
cancer and healthy donors was analyzed by Affymetrix Human Genome
U133 Plus 2.0 Array (platforms: GPL570, [HG-U133_Plus_2];
Affymetrix, Inc., California, USA).
Sample preparation for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
gene chip analysis
The differential genes were verified using RT-qPCR.
The procedure was as follows: Total RNA was isolated from the
cultured SC-M1 cells using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. Total RNA was used to synthesize cDNA
using the PrimeScriptTM RT reagent kit (cat. no. RR037A;
Takara Bio, Inc., Otsu, Japan), all processes were performed
according to the manufacturer's protocol, then the cDNA was
subjected to qPCR. The relative expression levels of lncRNAs were
measured using a SYBR® Premix Ex TaqTM (cat.
no. RR420A; Takara Bio, Inc.), according to the manufacturer's
protocol, using GAPDH as an internal control. RT-qPCR primers were
synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). The
5′-3′ primer sequences used for RT-qPCR were as follows: GAPDH
forward, GACTCATGACCACAGTCCATGC and reverse,
AGAGGCAGGGATGATGTTCTGRT-PCR was performed on an ABI 7500 Sequence
Detection system (ABI, Vernon, CA, USA). The thermocycling
conditions were as follows: 95°C for 30 sec, 40 cycles of 95°C for
5 sec and 60°C for 31 sec. The dissociation stage was 95°C for 15
sec, 60°C for 1 min and 95°C for 15 sec. The expression of target
genes was calculated based on the threshold cycle (CQ), and
relative expression levels were calculated as 2−[(Cq of target
genes)-(Cq of GAPDH)] (19)
following normalization with reference to the quantification of
GAPDH. Each experiment was repeated three times. The labeled target
was fragmented and hybridized to probe arrays, using Encore Biotin
Module from NuGEN Technologies, Inc. (San Carlos, CA, USA).
Stimulation of gastric cell line
The human gastric cancer SC-M1 cell line was
obtained from American Type Culture Collection (ATCC; Manassas, VA,
USA). The cells were cultured in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.), containing 10% fetal bovine serum (FBS;
Hyclone; GE Healthcare Life Sciences, Logan, UT, USA), 100 U/ml
penicillin and 100 mg/ml streptomycin. In certain experiments,
cells (1×105 cells/ml) were cultured in 96-well plates
for the examination of cell viability using an MTT assay (MTT Cell
Proliferation kit; Sangon Biotech Co., Ltd., Shanghai, China).
According to the manufacturer's protocol, formazan solubilization
solution was used to dissolve the formazan crystals and measure the
absorbance at 570 nm using a microplate reader (Thermo Fisher
Scientific, Inc.).
For the cell migration assay by Transwell method
(Transwell Permeable Supports, Corning, Cornstar, NY, USA), cells
(1×105 cells/ml) were transfected with HDAC siRNAs and
were cultured using Dulbecco's modified Eagle's medium (DMEM;
Hyclone; GE Healthcare Life Sciences) in the upper chamber of
24-well plates, and 0.5 ml DMEM containing 10% FBS were added to
the lower chambers. The total number of migrated cells were
calculated 48 h later by inverted microscope (OLYMPUS IX51; Olympus
Corporation, Tokyo, Japan. The magnification of the captured image
was ×400.
RNA interference
The siRNA sequence targeting HDAC1s (NCBI; GenBank
ID: NM_004964.2) and the scramble siRNA sequence were designed and
synthesized by GenePharma (Shanghai, CHINA). The sequences for the
si-HDAC1s were: siRNA1, 5′-gcaaagaaguccgaggc-3′; siRNA2,
5′-ucaaaggacacgccaagug-3′; and siRNA3, 5′-gagaagaaagaagucacc-3′.
For in vitro experiments, siRNAs were transfected into three
gastric cancer cell lines using Lipofectamine iMAX (Invitrogen;
Thermo Fisher Scientific, Inc.), the terminal concentration of
siRNA was 50 nM according to the manufacturer's protocol. At 48 h
following transfection, the efficiency of RNA interference was
determined by RT-qPCR that the method has been described in the
upper.
Cell apoptosis assay
Gastric cancer cells were stained using a
fluorescein isothiocyanate (FITC)-conjugated Annexin V and
propidium iodide (PI) kit (BD Biosciences, Franklin Lakes, NJ,
USA), according to the manufacturer's protocol. Cells
(1×106 cells/ml) were cultured in 12-well plates, and
following transfection with siRNAs for 24 h, the cells were
harvested and washed twice with cold phosphate-buffered saline with
gentle agitation. SC-M1 cells were then re-suspended and added to
binding buffer (1X), and cell density was adjusted to
2–5×105/ml. In the dark, 5 µl Annexin V-FITC was added
to the cell suspension volume of 195 µl and incubated for 10 min at
room temperature prior to the addition of 190 µl binding buffer and
10 µl PI. A total of 10,000 events per sample were acquired using a
FACS-scan flow cytometer (BD Biosciences, San Jose, CA, USA). The
percentage of apoptotic cells was analyzed by Acri (BD
Biosciences). The percentage of cells undergoing apoptosis was
analyzed using BD CellQuestTM Pro Software Analysis
Tutorial (BD Biosciences; version 5.1).
Western blot analysis
For western blotting, human gastric cancer SC-M1
cells were transfected with siRNAs in 12-well plates. The proteins
were extracted from SC-M1 cells according to the standard protocol.
Cells were lysed in cold lysis buffer (50 mM Tris, 150 mM NaCl, 10
mM ethylenediaminetetraacetic acid and 1% Triton X-100) containing
protease and phosphatase inhibitors. The protein determination
method was BCA (BCA Protein Assay kit, Sangon Biotech Co., Ltd.
Shanghai, China). Briefly, the loading of protein sample was 40 µg
each hole, the proteins were separated on 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels, and
subsequently transferred to nitrocellulose membranes. The membrane
was completely immersed in 5% skimmed milk powder for 60 min at
room temperature. The following primary antibodies were used: Total
p-21 (cat. no. PL0502562; dilution, 1:1,000; PLLABS, British
Columbia, Canada; http://www.pllabs.com/total-p53 (cat. no. ab131442;
dilution, 1:2,000: Epitomics; Abcam, Cambridge, MA, USA) and actin
(cat. no. PA1-183; dilution, 1:2,000: Thermo Fisher Scientific,
Inc.) antibodies. Incubate overnight at 4°C with primary
antibodies, respectively. Membranes were subsequently incubated
with horseradish peroxidase-conjugated secondary anti-rabbit
antibodies (cat. no. 7074; Cell Signaling Technology, Inc.,
Danvers, MA, USA) at a dilution of 1:4,000 for 1 h at room
temperature. Membranes were developed using the enhanced
chemiluminescence detection system (Pierce; Thermo Fisher
Scientific, Inc.).
Statistical analysis
Limma R package (1.10.1) was utilized to identify
the DEGs in GSE64951 and GSE19826 database, with a padj ≤0.05. SPSS
version 19.0 for Windows (IBM Corp., Armonk, NY, USA) was used for
all analyses. All the data are expressed as the mean ± standard
deviation. The correlation between HDAC1 and upregulated lncRNAs
(BC01600 and AF116637 and uc004ebu.1) in gastric cancer were
assessed using one-way analysis of variance followed by Tukey's
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Abnormal expression of lncRNA is
identified in gastric cancer
Accumulating evidence has demonstrated that the
abnormal expression of lncRNAs is associated with the progression
of certain diseases. In order to detect the expression profile of
lncRNAs in gastric cancer tissues, the expression of lncRNAs in the
tissues of patients with gastric cancer and healthy donors was
analyzed by Affymetrix Human Genome U133 Plus 2.0 Array. Compared
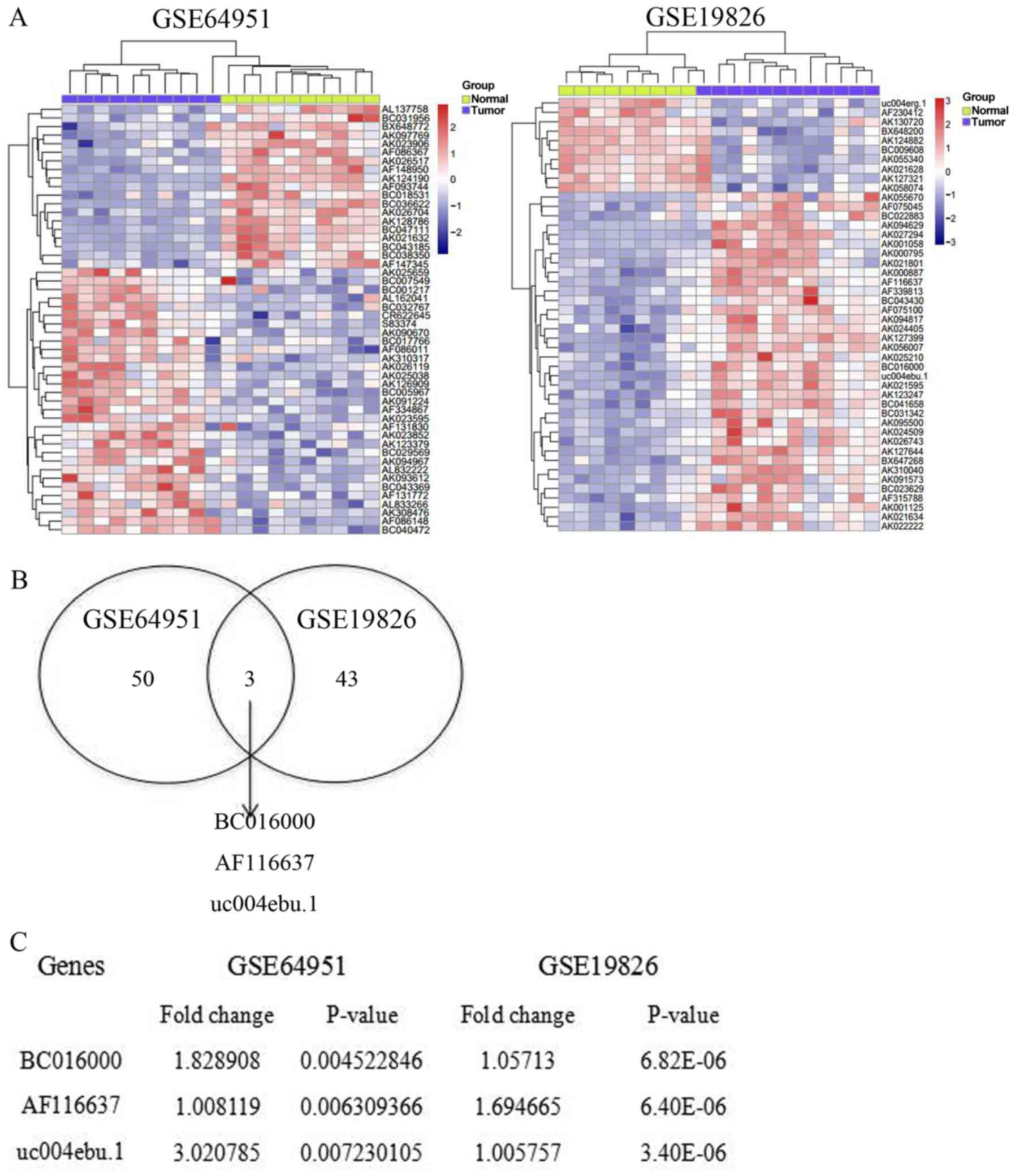
with normal controls, the expression profile of lncRNA is
completely different in gastric cancer. A total of 5,621 lncRNAs
were analyzed from the samples of patients with gastric cancer and
healthy donors. In database no. GSE64951, 53 lncRNAs were
significantly altered. Among these lncRNAs, 19 downregulated
lncRNAs and 34 upregulated lncRNAs were identified from the samples
of patients with gastric cancer, compared with matched samples from
healthy donors (Fig. 1A). Meanwhile,
in database no. GSE19826, 46 lncRNAs were abnormally expressed in
the samples from patients with gastric cancer. Ten of these lncRNAs
were upregulated, while 36 of these lncRNAs were downregulated
(Fig. 1A). In order to further
investigate the potential DE-lncRNAs, a Venn diagram was
constructed using the upregulated lncRNAs in these two databases.
The online software can be found at http://bioinfogp.cnb.csic.es/tools/venny/index.html.
It was revealed that 3 lncRNAs overlapped between these two
databases, GSE64951 and GSE19826 (Fig.
1B). The three significantly upregulated lncRNAs were BC016000,
AF116637 and uc004edu.1, with a fold-change of 1.83, 1.01 and 3.02,
respectively (Fig. 1C). Taken
together, the results of the present study revealed that the
expression profile is completely different in gastric cancer
tissues compared with the normal tissues, indicating the abnormal
expression profile of lncRNAs in the gastric cancer.
The correlation between HDAC1 and
upregulated lncRNAs in gastric cancer
HDACs are a family of multi-functional proteins,
which are involved in tumor suppression. HDAC1 is a very important
member of the HDAC family. Therefore, the present study
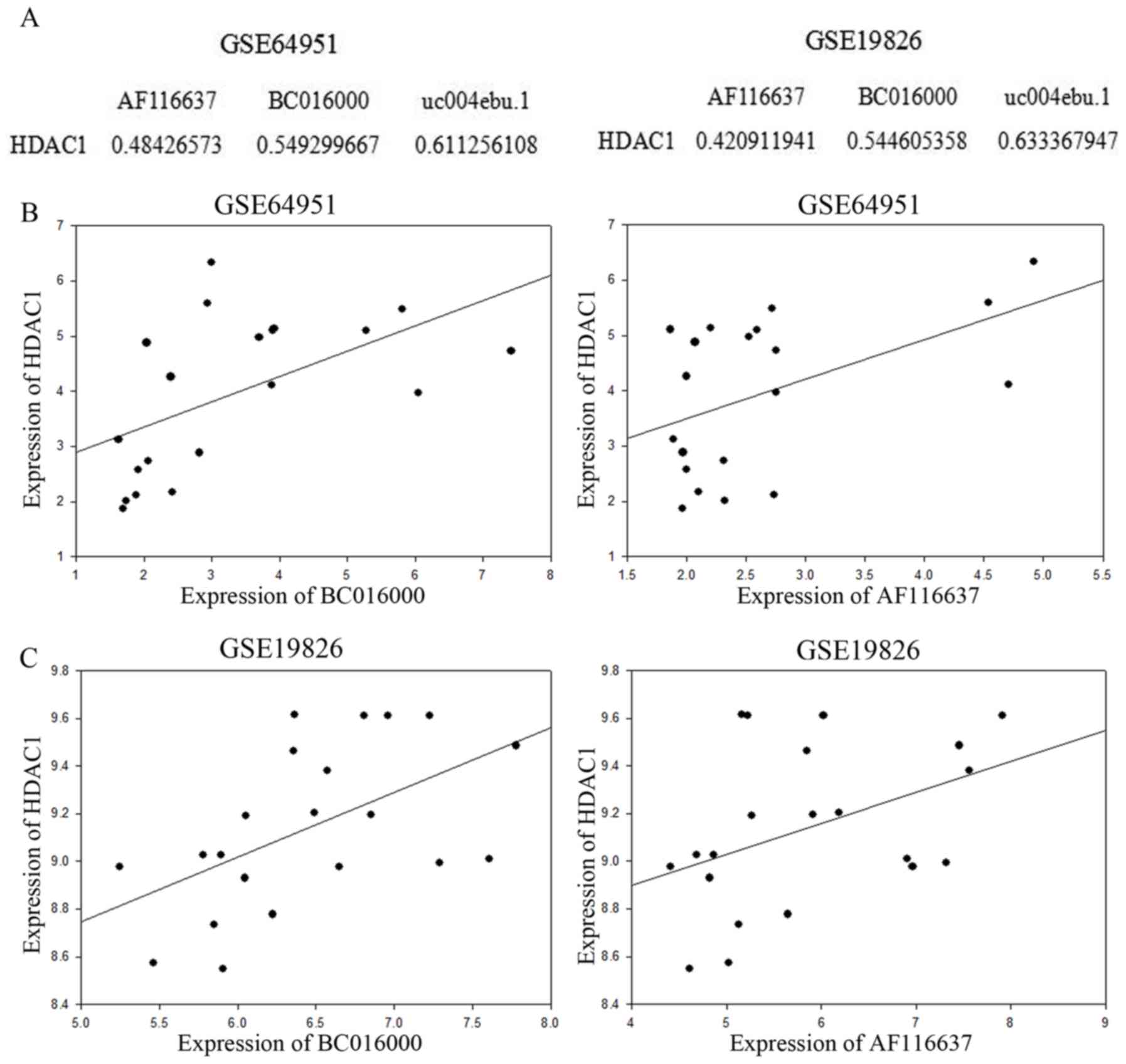
investigated the association between HDAC1 and the upregulated
lncRNAs in gastric cancer. Correlation analysis of HDAC1 and the
upregulated lncRNAs, BC016000 and AF116637, was conducted. The
Pearson's correlation coefficient (PCC) between HDAC1 and BC016000
and AF116637 was calculated using the expression value. The
absolute values of Correlation Coefficient were shown in Fig. 2A, for example: The absolute value of
Correlation Coefficient between HDAC1 expression value and AF116637
expression value was 0.48426573 in the GSE64951 database.
Correlation analysis was performed in the GSE64951 and GSE19826
databases. In the two aforementioned databases, lncRNAs BC016000
and AF116637 exhibited a moderate to strong positive correlation
with HDAC1 with Correlation Coefficients of 0.545 and 0.421,
respectively, in the GSE64951 database (Fig. 2A and B), and in the GSE19826 database,
the Correlation Coefficients were 0.549 and 0.484, respectively
(Fig. 2A and C).
Silencing of HDAC1 downregulates the
expression of BC01600 and AF116637
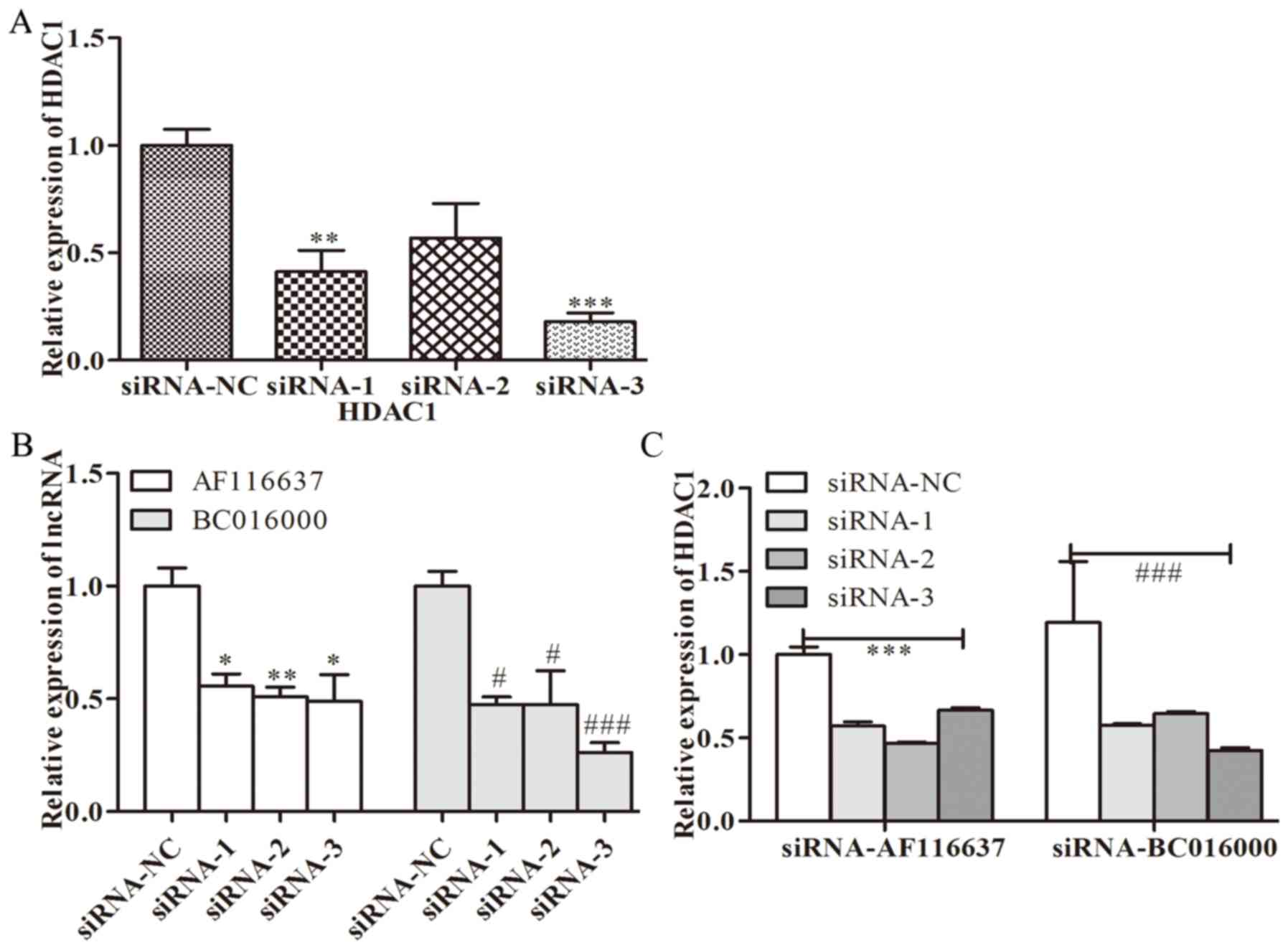
To further investigate whether BC01600 and AF116637
were correlated with HDAC1, the expression levels of BC01600 and
AF116637 were evaluated following HDAC1-silencing with its specific
siRNA. Three pairs of siRNAs targeting HDAC1 were designed. It was
revealed that all these three siRNA sequences could efficiently
inhibit the expression of HDAC1 (Fig.
3A). HDAC1 siRNA transfection with these three siRNAs
significantly inhibited the expression of BC01600 and AF116637
(Fig. 3B). Additionally, three pairs
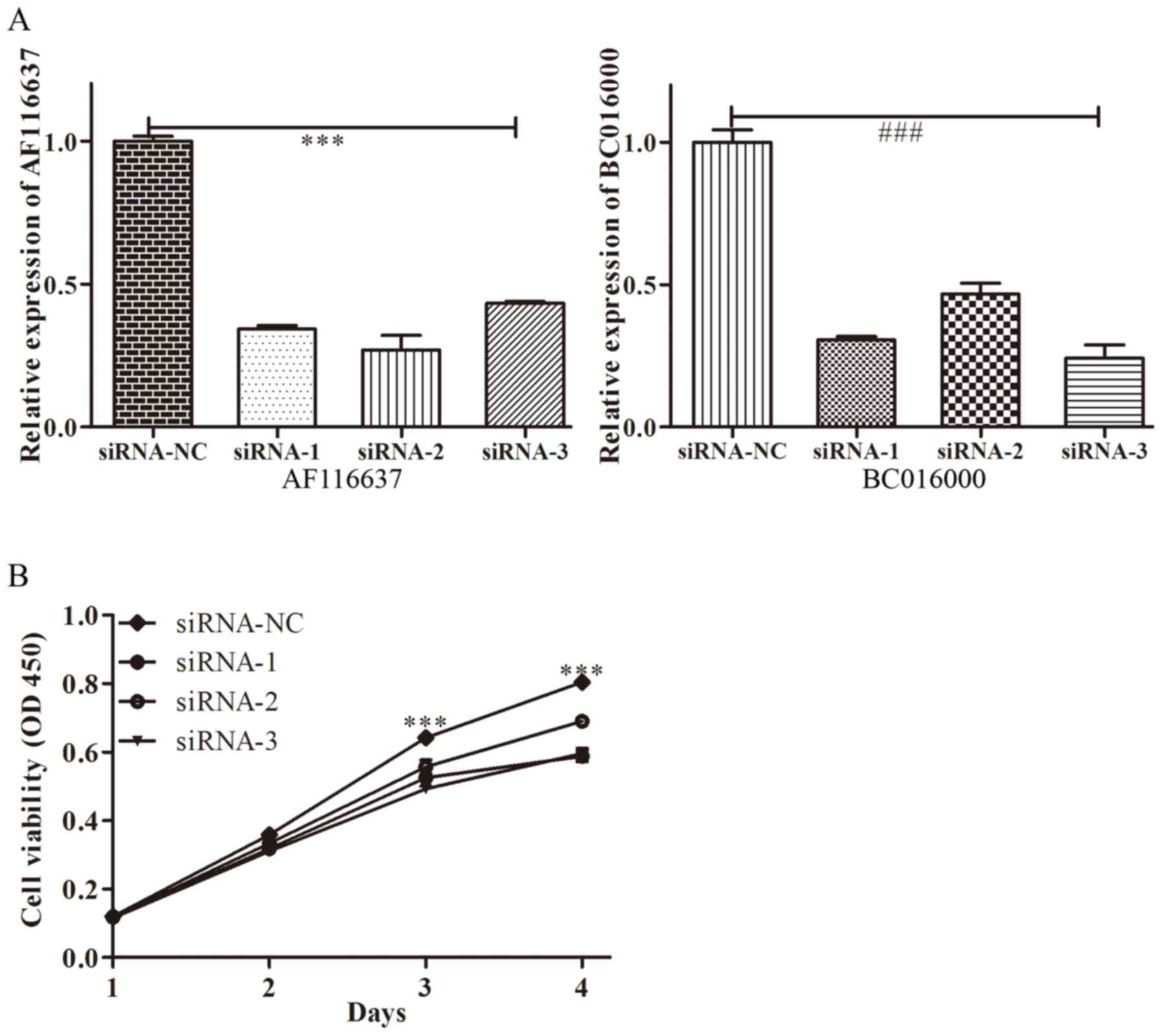
of siRNAs targeting lncRNA, BC01600 and AF116637 were designed.
Next, the expression of BC01600 and AF116637 was efficiently
decreased by their respective siRNA sequences (Fig. 4). Subsequently, it was demonstrated
that BC01600 and AF116637 lncRNA siRNA transfection with their
respective siRNAs significantly inhibited the expression of HDAC1
(Fig. 3B). Furthermore, the
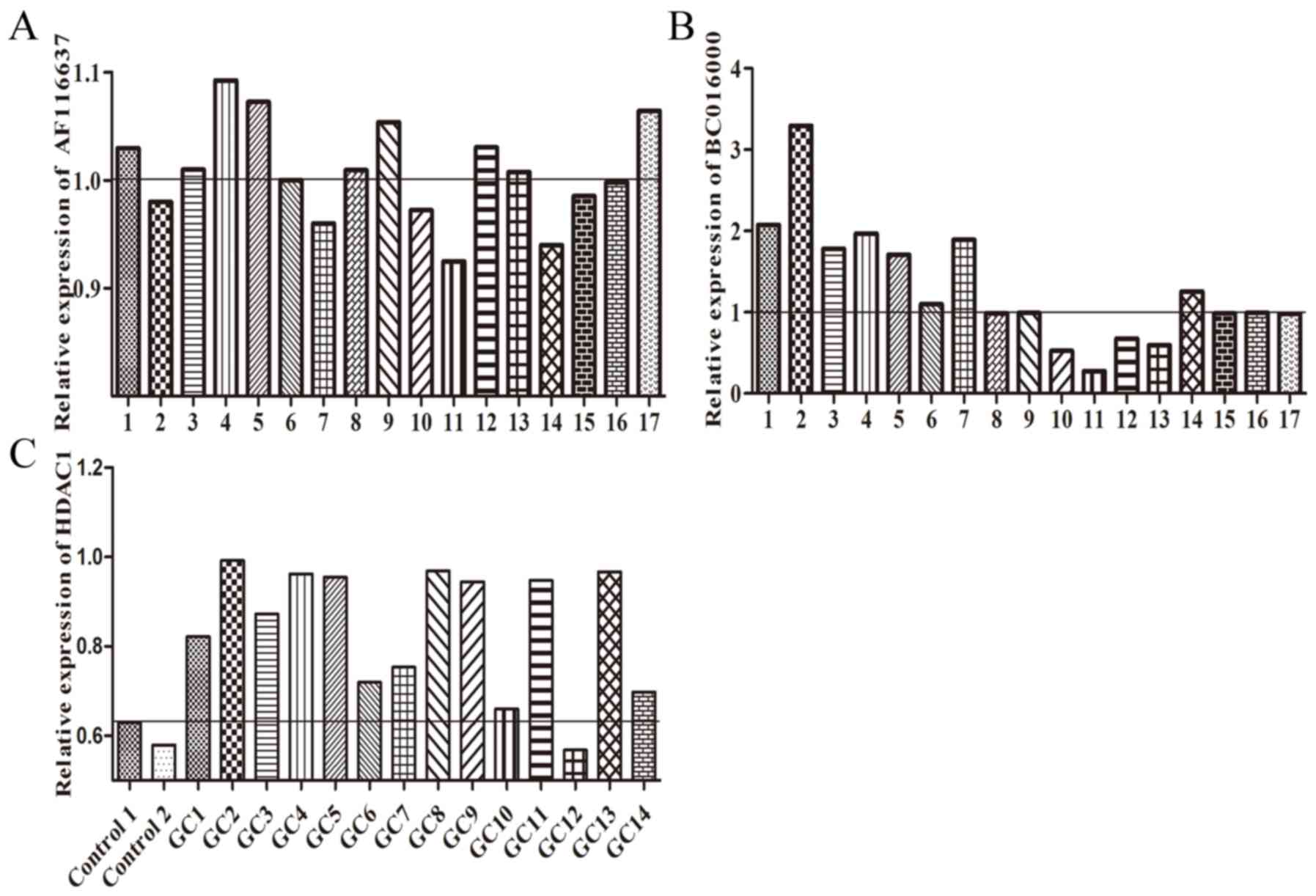
expression of BC01600, AF116637 and HDAC1 was analyzed in gastric
cancer tissues. It was revealed that these two lncRNAs and HDAC1
have an increased expression trend in gastric cancer tissue
(Fig. 5). Taken together, these
results indicated that HDAC1 could promote the expression of
BC01600 and AF116637 in gastric cancer.
HDAC1 promotes the proliferation of
gastric cancer cells
To further examine the role of HDACs in the
progression of gastric cancer, HDAC1 was silenced by transfecting
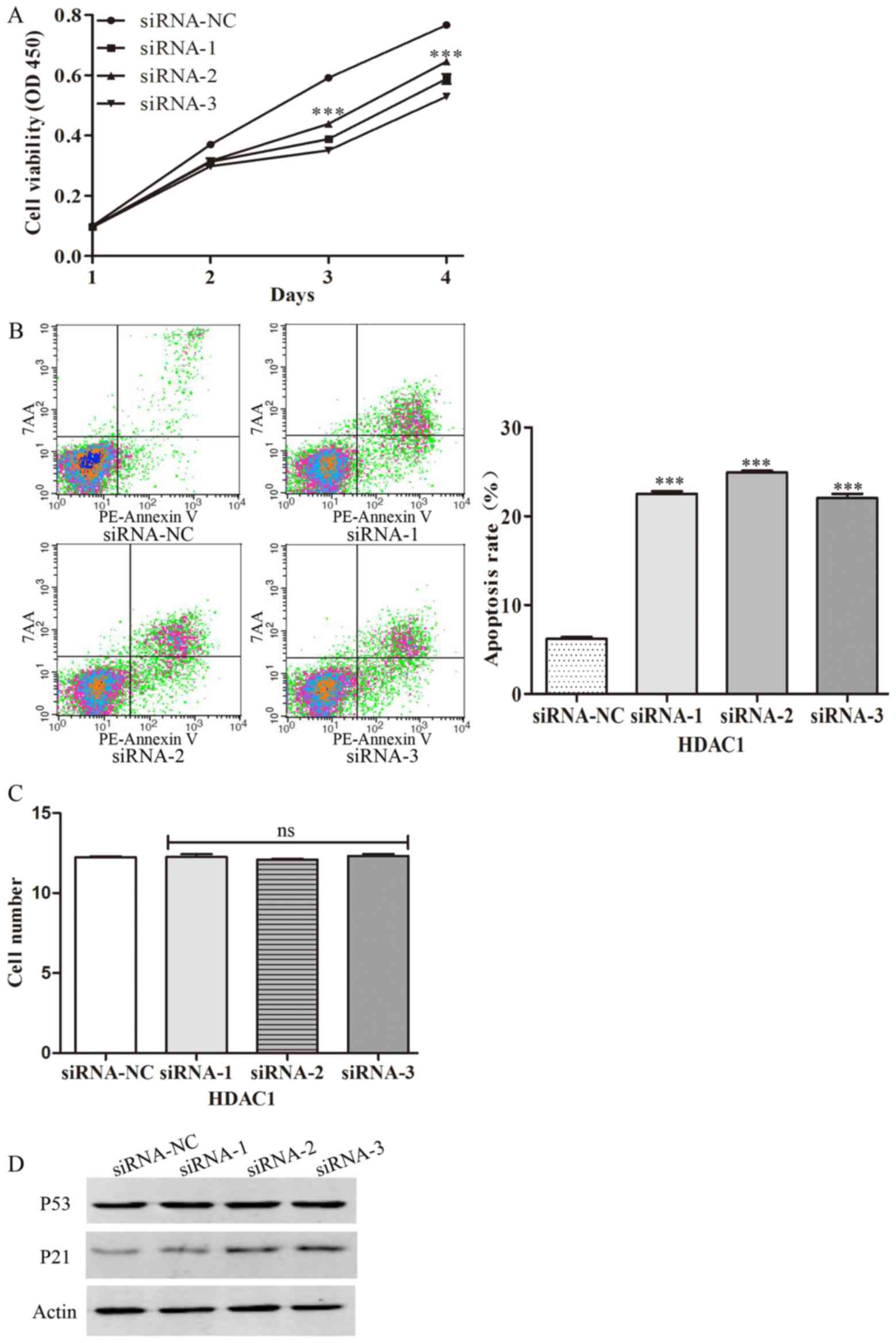
gastric cancer cells with specific siRNAs. It was revealed that
silencing of HDAC1 decreased cell viability, indicating that HDAC1
can inhibit the proliferation of gastric cancer cells (Fig. 6A). Furthermore, the present study also
examined the effect of HDAC1 on apoptosis by flow cytometry. It was
demonstrated that silencing of HDAC1 increased the percentage of
apoptotic cells, indicating that HDAC1 can decrease the apoptosis
of gastric cancer cells (Fig. 6B).
Following transfection of the gastric cancer cells with specific
siRNAs, the present study also examined the migration of gastric
cancer cells. The results demonstrated that silencing of HDAC1 with
specific siRNAs has no effect on the migration of gastric cancer
cells (Fig. 6C). Furthermore, the
role of HDAC1 in the activation of tissue-associated genes, p53 and
p21, was examined. It was demonstrated that transfection with HDAC1
siRNAs significantly increased the expression of total-p21, but had
no effect on the expression of total-p53 (Fig. 6D). Taken together, the results of the
present study demonstrated that HDACs could promote the
proliferation of gastric cancer.
Discussion
The present study demonstrated that HDAC1 could
promote the proliferation of GC cells, possibly by upregulating the
expression of lncRNAs, BC01600 and AF116637. High expression of
HDAC1 and abnormal expression of lncRNA was observed in the tissues
of patients with GC. It was demonstrated that the expression of
BC01600 and AF116637, two typically upregulated lncRNAs, is
correlated with the expression of HDAC1, as evidenced by the
decreased expression of these two lncRNAs in HDAC1-knockout GC cell
lines, and by reduced expression of HDAC1 in lncRNA, BC01600 and
AF116637, knockout GC cell lines, respectively. Silencing of HDACs
increased the p21-dependent apoptosis of the GC cell lines.
Investigating novel treatments for GC, one of the
most common types of cancer, is necessary. Limitations of current
treatment options include the following: The patients in the later
phase of GC experienced a lot of painful and a high mortality rate,
even after surgery (20). It is
useful to identify efficient biomarkers for the diagnosis of GC in
the early phase of GC. Epigenetics has become a popular field of
molecular biology recently and accumulating evidence has
demonstrated that epigenetic mechanisms contribute toward
differentiation, aging and disease development, particularly during
the progression of cancer (13,21,22).
lncRNAs serve important roles in the progression of
cancer and numerous hallmarks of cancer are controlled by lncRNAs
(8-10,23-25).
Various lncRNAs have been implicated in the regulation of
individual genes via epigenetic regulation (26). Previous studies have demonstrated that
overexpression of lncRNAs is implicated in the metastasis of human
tumors (27,28). An increasing volume of evidence has
indicated that specific lncRNA mutations are associated with the
progression of cancer (8). Therefore,
lncRNAs could be a promising candidate for the diagnosis and
therapy of cancer (28,29). The present study demonstrated that the
expression profile of lncRNAs is completely different in gastric
cancer, compared with normal controls by using the bioinformatics
methods according to the GSE64951 and GSE19826 databases. The
following three lncRNAs were selected: BC016000, AF116637 and
uc004edu.1, which were overlapped in these two databases, as
determined by constructing a Venn diagram using the upregulated
lncRNAs in these two databases. Therefore, the abnormal expression
profile of lncRNAs may be involved in or be a factor in promoting
the progression of GC. However, the way in which the expression of
these lncRNAs is regulated remains unknown.
HDACs are multi-functional proteins, which serve
critical roles in transcription regulation (30). Aberrant expression of HDACs is
frequently observed in various types of human cancer. In the
majority of cases, upregulation of HDACs is associated with
advanced disease and poor outcomes in patients (15). HDACs have also been revealed to be
broadly dysregulated in multiple myeloma (31). HDACs are also involved in the
progression of several types of cancer, including melanoma and
hepatocellular carcinoma (15,18).
Participating in a range of cellular and molecular pathways, HDACs
can either repress tumor suppressor gene expression or modify key
molecules in oncogenic cell-signaling pathways. However, the
precise role of HDACs has not yet been revealed. A previous study
demonstrated that HDACs could upregulate the expression of
microRNAs, indicating that HDACs may serve certain roles in
regulating the expression of lncRNAs (32). HDAC1 is an important member of the
HDACs family. In order to determine the role of HDAC1 in the
expression of lncRNAs, correlation analysis of HDAC1 and the
upregulated lncRNAs, BC016000 and AF116637, was conducted, and
BC016000 and AF116637 lncRNAs exhibited a positive and strong
correlation with HDAC1, indicating that HDAC1 serves roles in the
expression of lncRNAs. The results of the present study
demonstrated that HDACs may be involved in the progression of GC by
upregulating the expression of lncRNAs.
Cancer cells are abnormally proliferated and
differentiated cells. Proliferation, apoptosis and migration are
important for the progression of cancer. The role of HDAC1 in the
proliferation, apoptosis and migration of GC cancer cells was also
examined. It was demonstrated that silencing of HDAC1 may decrease
the proliferation and increase the apoptosis of gastric cancer
cells. However, HDAC1 has no effect on the migration of gastric
cancer cells. Therefore, HDAC1 is specifically involved in the
proliferation of gastric cancer cells.
In summary, HDAC1 is critical for the progression of
GC, by promoting the proliferation of cancer cells, which may be
regulated by the HDAC1-mediated decreased or increased expression
of lncRNAs. The present study demonstrated the correlation of HDACs
and lncRNAs, and provided a possible mechanism by which HDACs
regulate the progression of gastric cancer. The results of the
present study provided a possible target for the clinical treatment
of gastric cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Jiangxi Province Natural Science Foundation of China (grant no.
2010BSA14300) and Science and Technology Department of Jiangxi
Province, Key R&D Plan (grant no. 20171BBG700870).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC designed the research. ZhiY and JZ performed the
research. TW and ZiY were responsible for clinical diagnosis and
sample collection. HL contributed to data collection and
statistical analysis. ZhiY, JZ and JC wrote the manuscript. All the
authors read and approved the final manuscript.
Ethics statement and consent to
participate
Human samples used in the present study were
obtained from the patients with written informed consent. The
present study was approved by the Ethics Committee of Jiangxi
Province People's Hospital and was conducted according to The
Declaration of Helsinki.
Patient consent for publication
Consent for publication was obtained from all
patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Matsuda T and Saika K: Worldwide burden of
cancer incidence in 2002 extrapolated from cancer incidence in five
continents Vol.IX. Jpn J Clin Oncol. 42:1111–1112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guo LL, Song CH, Wang P, Dai LP, Zhang JY
and Wang KJ: Competing endogenous RNA networks and gastric cancer.
World J Gastroenterol. 21:11680–11687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu J, Huang CM, Zheng CH, Li P, Xie JW,
Wang JB and Lin JX: Consideration of tumor size improves the
accuracy of TNM predictions in patients with gastric cancer after
curative gastrectomy. Surg Oncol. 22:167–171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Müller S, Raulefs S, Bruns P, Afonso-Grunz
F, Plötner A, Thermann R, Jäger C, Schlitter AM, Kong B, Regel I,
et al: Next-generation sequencing reveals novel differentially
regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA in pancreatic
cancer. Mol Cancer. 14:942015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Müller S, Raulefs S, Bruns P, Afonso-Grunz
F, Plötner A, Thermann R, Jäger C, Schlitter AM, Kong B, Regel I,
et al: Erratum to: Next-generation sequencing reveals novel
differentially regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA
in pancreatic cancer. Mol Cancer. 14:1442015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao T, Xu J, Liu L, Bai J, Xu C, Xiao Y,
Li X and Zhang L: Identification of cancer-related lncRNAs through
integrating genome, regulome and transcriptome features. Mol
Biosyst. 11:126–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Evans JR, Feng FY and Chinnaiyan AM: The
bright side of dark matter: lncRNAs in cancer. J Clin Invest.
126:2775–2782. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gu Y, Chen T, Li G, Yu X, Lu Y, Wang H and
Teng L: LncRNAs: Emerging biomarkers in gastric cancer. Future
Oncol. 11:2427–2441. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang X, Zhi X, Gao Y, Ta N, Jiang H and
Zheng J: LncRNAs in pancreatic cancer. Oncotarget. 30:57379–57390.
2016.
|
|
11
|
Jonsson P, Coarfa C, Mesmar F, Raz T,
Rajapakshe K, Thompson JF, Gunaratne PH and Williams C:
Single-molecule sequencing reveals estrogen-regulated clinically
relevant lncRNAs in breast cancer. Mol Endocrinol. 29:1634–1645.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng L, Yuan X, Jiang B, Tang Z and Li GC:
LncRNAs: Key players and novel insights into cervical cancer.
Tumour Biol. 37:2779–2788. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ali Z, Deng Y, Tang Y, Zheng S, Ma N and
He N: Epigenetic deregulations in gastric cancer. J Nanosci
Nanotechnol. 13:40–51. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hildmann C, Riester D and Schwienhorst A:
Histone deacetylases-an important class of cellular regulators with
a variety of functions. Appl Microbiol Biotechnol. 75:487–497.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mithraprabhu S, Kalff A, Chow A, Khong T
and Spencer A: Dysregulated class I histone deacetylases are
indicators of poor prognosis in multiple myeloma. Epigenetics.
9:1511–1520. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Christensen DP, Dahllöf M, Lundh M,
Rasmussen DN, Nielsen MD, Billestrup N, Grunnet LG and
Mandrup-Poulsen T: Histone deacetylase (HDAC) inhibition as a novel
treatment for diabetes mellitus. Mol Med. 17:378–390. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mutze K, Langer R, Becker K, Ott K,
Novotny A, Luber B, Hapfelmeier A, Göttlicher M, Höfler H and
Keller G: Histone deacetylase (HDAC) 1 and 2 expression and
chemotherapy in gastric cancer. Ann Surg Oncol. 17:3336–3343. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Z, Zhou L, Wu LM, Xie HY, Zhang F and
Zheng SS: Combination of polymorphisms within the HDAC1 and HDAC3
gene predict tumor recurrence in hepatocellular carcinoma patients
that have undergone transplant therapy. Clin Chem Lab Med.
48:1785–1791. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wan QS and Zhang KH: Noninvasive detection
of gastric cancer. Tumour Biol. 37:11633–11643. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gigek CO, Chen ES, Calcagno DQ, Wisnieski
F, Burbano RR and Smith MA: Epigenetic mechanisms in gastric
cancer. Epigenomics. 4:279–294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hagelkruys A, Sawicka A, Rennmayr M and
Seiser C: The biology of HDAC in cancer: The nuclear and epigenetic
components. Handb Exp Pharmacol. 206:13–37. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang YN, Zhang K, Hu ZM, Qi HX, Shi ZM,
Han XH, Han YW and Hong W: Hypoxia-regulated lncRNAs in cancer.
Gene. 575:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cao B, Song N, Zhang M, Di C, Yang Y, Lu
Y, Chen R, Lu ZJ and Guo M: Systematic study of novel lncRNAs in
different gastrointestinal cancer cells. Discov Med. 21:159–171.
2016.PubMed/NCBI
|
|
25
|
Cheng WS, Tao H, Hu EP, Liu S, Cai HR, Tao
XL, Zhang L, Mao JJ and Yan DL: Both genes and lncRNAs can be used
as biomarkers of prostate cancer by using high throughput
sequencing data. Eur Rev Med Pharmacol Sci. 18:3504–3510.
2014.PubMed/NCBI
|
|
26
|
Cogill SB and Wang L: Co-expression
network analysis of human lncRNAs and cancer genes. Cancer Inform.
13 (Suppl 5):S49–S59. 2014.
|
|
27
|
Han J, Rong LF, Shi CB, Dong XG, Wang J,
Wang BL, Wen H and He ZY: Screening of lymph nodes metastasis
associated lncRNAs in colorectal cancer patients. World J
Gastroenterol. 20:8139–8150. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang F, Zhang L and Zhang C: Long
noncoding RNAs and tumorigenesis: Genetic associations, molecular
mechanisms, and therapeutic strategies. Tumour Biol. 37:163–175.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fayda M, Isin M, Tambas M, Guveli M, Meral
R, Altun M, Sahin D, Ozkan G, Sanli Y, Isin H, et al: Do
circulating long non-coding RNAs (lncRNAs) (LincRNA-p21, GAS 5,
HOTAIR) predict the treatment response in patients with head and
neck cancer treated with chemoradiotherapy? Tumour Biol.
37:3969–3978. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Seto E and Yoshida M: Erasers of histone
acetylation: The histone deacetylase enzymes. Cold Spring Harb
Perspect Biol. 6:a0187132014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Terui Y: The epigenetic alteration and the
effect of HDAC inhibitors in multiple myeloma. Nihon Rinsho.
73:124–129. 2015.(In Japanese). PubMed/NCBI
|
|
32
|
Srinivas C, Swathi V, Priyanka C, Anjana
Devi T, Subba Reddy BV, Janaki Ramaiah M, Bhadra U and Bhadra MP:
Novel SAHA analogues inhibit HDACs, induce apoptosis and modulate
the expression of microRNAs in hepatocellular carcinoma. Apoptosis.
21:1249–1264. 2016. View Article : Google Scholar : PubMed/NCBI
|