Introduction
The cell cycle involves a series of events
controlled by a complex network of proteins, while mitosis is a
cell division process of nuclear-envelope breakdown, chromosome
condensation and spindle assembly (1). Cyclin-dependent kinase 1 (Cdk1)-cyclin B
controls mitotic entry and exit. The inhibitory residues of Cdk1
are dephosphorylated by the dual-specificity phosphatase Cdc25,
which is subsequently activated at the G2-M transition. This in
turn phosphorylates multiple substrates by activating Cdk1-cyclin B
(2,3).
Previous studies have illustrated that protein phosphatase 2A
(PP2A) negatively regulates the activation of Cdk1-cyclin B
substrates, and promotes normal cell division (4). Greatwall (Gwl), a serine/threonine
kinase originally identified in Drosophila (5) is essential for mitosis progression
through PP2A inhibition by interacting with Alpha-endosulfine
(Ensa)/cAMP-regulated phosphoprotein 19 (Arpp19) (6,7).
As a mammalian ortholog of Gwl, human microtubule
associated serine/threonine kinase-like (MASTL) exhibits 50.2%
sequence homology with Drosophila Gwl and 65.7% with
Xenopus Gwl. Experiments in human cell lines, including HeLa
or U2OS, have illustrated that MASTL has an essential
function in regulating mitosis, similar to that of Gwl in
Drosophila (8); there are also
reports supporting the role of MASTL in lung cancer and
thrombocytopenia (9,10). Nevertheless, there is currently
limited knowledge of the potential functions of MASTL in
human diseases.
Liver cancer develops slowly from chronic liver
diseases, including chronic hepatitis caused by hepatitis B virus
(HBV) and hepatitis C virus (HCV) infection (11–13); as a
result, genetic alterations and the loss of cell cycle regulation
are two of the basic mechanisms of carcinogenesis (14–16).
Chronic inflammation associated with viral infection favors the
recruitment of macrophages that produce large quantities of
proinflammatory cytokines (17).
Studies have illustrated that TNF-α and IL-6 are closely associated
with the progression of liver cancer, and that the IκB kinase (IKK)
and c-Jun NH2-terminal kinase (JNK) signaling pathways are involved
in this process (18). However, the
detailed mechanisms of hepatocarcinogenesis remain to be
clarified.
In the present study, MASTL was induced by
TNF-α and IL-6 in liver cancer cell lines, promoting cell
proliferation by regulating the progression of the cell cycle.
MASTL was markedly overexpressed in human liver cancer
tissues compared with non-tumor liver tissues. TNF-α and IL-6
promote the trimethylation of H3K4 to facilitate NF-κB-induced
MASTL transcription. Therefore, the data provided novel
insights into the critical function of MASTL in the
progression of live cancer.
Materials and methods
Liver cancer samples and sera
A total of 20 human liver cancer tissues and 20
adjacent non-tumor liver tissue specimens and their sera were used
(Table I), we also recruited 20
healthy control subjects (Table II)
from physical examination and the donating blood were collected for
experiments. Patients with liver cancer were treated, and frozen
tissue samples were obtained from The Affiliated Hospital of Hebei
University, same as the healthy control subjects. The tumor type
was confirmed by a pathologist. The study was approved by the
Ethics Committee of The Affiliated Hospital of Hebei University,
and written informed consent was obtained from all participants.
All human materials were used in accordance with the Declaration of
Helsinki Principles and relevant policies and regulations of China
and the policies of the Institutional Review Board of the Hospital
of Hebei University.
 | Table I.Patient clinical characteristics. |
Table I.
Patient clinical characteristics.
| Patient no. | Age | Sex | Histology |
|---|
| 1 | 80 | Female | Hepatocellular
carcinoma |
| 2 | 32 | Female | Hepatocellular
carcinoma |
| 3 | 59 | Female | Hepatocellular
carcinoma |
| 4 | 61 | Female | Hepatocellular
carcinoma |
| 5 | 59 | Female | Hepatocellular
carcinoma |
| 6 | 67 | Female | Hepatocellular
carcinoma |
| 7 | 54 | Female | Hepatocellular
carcinoma |
| 8 | 56 | Male | Hepatocellular
carcinoma |
| 9 | 77 | Female | Hepatocellular
carcinoma |
| 10 | 60 | Female | Hepatocellular
carcinoma |
| 11 | 35 | Female | Hepatocellular
carcinoma |
| 12 | 57 | Female | Hepatocellular
carcinoma |
| 13 | 30 | Female | Hepatocellular
carcinoma |
| 14 | 67 | Female | Hepatocellular
carcinoma |
| 15 | 67 | Female | Hepatocellular
carcinoma |
| 16 | 60 | Male | Hepatocellular
carcinoma |
| 17 | 64 | Male | Hepatocellular
carcinoma |
| 18 | 43 | Male | Hepatocellular
carcinoma |
| 19 | 42 | Male | Hepatocellular
carcinoma |
| 20 | 81 | Male | Hepatocellular
carcinoma |
| Table II.The characteristics of healthy
subjects. |
Table II.
The characteristics of healthy
subjects.
| No. | Age | Sex |
|---|
| 1 | 35 | Female |
| 2 | 52 | Female |
| 3 | 40 | Female |
| 4 | 76 | Male |
| 5 | 65 | Male |
| 6 | 37 | Male |
| 7 | 37 | Female |
| 8 | 30 | Male |
| 9 | 58 | Male |
| 10 | 52 | Male |
| 11 | 50 | Female |
| 12 | 38 | Female |
| 13 | 59 | Male |
| 14 | 75 | Male |
| 15 | 35 | Male |
| 16 | 39 | Male |
| 17 | 68 | Male |
| 18 | 38 | Male |
| 19 | 40 | Female |
| 20 | 44 | Female |
Cell culture
Human cell lines HepG2 and SUN387 were purchased
from the Type Culture Collection of the Chinese Academy of Sciences
(Beijing, China). The cell lines were maintained in RPMI-1640
medium (cat no. 11875; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and supplemented with 10% fetal bovine serum (cat
no. 10437028; Gibco; Thermo Fisher Scientific, Inc.). Cells were
incubated at 37°C in a humidified incubator, supplemented with 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from liver cancer cells
using guanidinium thiocyanate (cat no. 5596026; Invitrogen; Thermo
Fisher Scientific, Inc.). cDNA was generated by RT using oligo (dT)
primers as outlined in the manufacturer's protocol (cat no.
R233-01; Vazyme, Piscataway, NJ, USA). For the thermocyling
conditions, 50°C 15 min, 85°C 5 sec. Target genes and controls were
treated under the same conditions and analyzed by qPCR using SYBR
Premix Ex Taq™ (cat. no. RR420L; Takara Biotechnology Co., Ltd.,
Dalian, China), according to the manufacturer's protocol. The
primers used were as follows: MASTL forward,
5′-ATCCTGTATGCCACATCAGAC-3′, and reverse,
5′-TTTCTTCACCTTCTGGCCAAG-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′, and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′.
These results were normalized by the Pfaffi method (19) using CFX96 C1000 Touch Thermal Cycler
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Transfection and RNA interference
(RNAi)
RNAi experiments were performed using 30 nM small
interfering RNA (siRNA) oligos (Shanghai GenePharma Co., Ltd,
Shanghai, China) by targeting human MASTL. Liver cancer
cells were seeded at 2×105 cells per well in a 6-well
plate. Control siRNAs consisting of a scrambled sequence were used
as a negative control. Transfection was performed with
Lipofectamine® 2000 reagent (cat no. 11668019;
Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The efficiency of RNAi was confirmed by
western blot analysis 48 h post-transfection, and the phenotype was
detected following confirmation of MASTL knockdown by siRNA.
The MASTL siRNA sequence was 5′-GGACAAGTGTTATCGCTTA-3′ (8).
Western blot analysis
Protein extracts were prepared with
radioimmunoprecipitation assay buffer according to the
manufacturer's protocol (cat no. 89900; Pierce; Thermo Fisher
Scientific, Inc.). The protein concentration was detected by BCA
assay (cat. no. CW0014S; CWBIO; Beijing, China). In brief, 40 µg of
whole cell lysate was separated by SDS-PAGE at 12% polyacrylamide
and then transferred on to a PVDF membrane. Then, the membrane was
blocked with 5% BSA (cat. no. A8010; Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) in PBST for 1h at room
temperature and incubated with the primary antibodies overnight at
4°C. Western blot analysis was performed using antibodies against
MASTL (1:1,000) (cat no. ab86387; Abcam; Cambridge, MA, USA) and
β-actin (1:5,000) (cat. no. 4967; Cell Signaling Technology, Inc.,
Danvers, MA, USA). The secondary antibodies conjugated with HRP
(1:10,000) for 90 min at room temperature. The blots were developed
using enhanced chemiluminescence reagent (cat. no. GERPN2109; GE
Healthcare, Chicago, IL, USA) by GeneGnome XRQ (Syngene, Syngene
Division of Synoptics Ltd; UK).
MTT assay
HepG2 and SUN3r7 cells were plated at
1×104 cells per well in 96-well plates and 10 µl (cat
no. C0009; Beyotime Institute of Biotechnology, Haimen, China) was
added to the cells. MTT was removed 4 h post incubation, followed
by pipetting of dimethyl sulfoxide to solubilize the formazan
products. Absorbance was measured with a microplate reader (Gen5;
BioTek Instruments, Inc., Winooski, VT, USA) at 570 nm.
Flow cytometric analysis
HepG2 cells transfected with siRNA were harvested,
washed with PBS, and fixed in 70% ethanol at 4°C overnight. The
fixed cells were stained with propidium iodide (PI; cat no. C0080;
Beijing Solarbio Science & Technology Co., Ltd.) at 37°C for 30
min prior to analysis via fluorescence-activated cell sorting
(FACS) (Becton, Dickinson and Company, Franklin Lakes, NJ, USA).
The cell cycle profiles were interpreted by BD CellQuest Pro
software version 5.1 (BD Biosciences; Becton, Dickinson and
Company).
Generation of vectors
Briefly, to create the MASTL kinase-dead mutation,
the nucleotide residues 718–816 of MASTL were deleted. The
DNA coding sequences of MASTL and the kinase-dead mutant of
MASTL were cloned into the pcDNA3.1 eukaryotic expression
vectors (Invitrogen; Thermo Fisher Scientific, Inc., USA) and were
confirmed by sequencing.
Immunocytochemistry (IHC) and
immunofluorescence histochemistry (IHF)
Cells were cultured overnight on a glass coverslip
and fixed using 4% paraformaldehyde for 30 min at room temperature.
The cells were incubated with primary anti-MASTL antibody and
secondary fluorescein isothiocyanate (FITC)-goat anti-rabbit IgG
antibody (cat no. ab6717; Abcam; USA). DAPI (cat. no. C1002;
Beyotime Institute of Biotechnology) was employed for nuclear
staining. Paraffin sections were compared with non-tumor liver
samples fixed using 4% paraformaldehyde overnight at room
temperature (tissue sections 8 mm), 0.5% Triton X-100 in PBS was
used for reducing non-specific interactions at room temperature for
1 h, and subsequently stained with anti-MASTL antibody (1:100) and
FITC-goat anti-rabbit IgG antibodies (1:50). Images were acquired
using a fluorescence microscope (magnification, ×100) (E600; Nikon
Corporation, Tokyo, Japan) and the results were evaluated by
fluorescence signal intensity using Zeiss 510 META software version
3.5.
Enzyme-linked immunosorbent assay
(ELISA)
IL-6 and TNF-α quantification in healthy and liver
cancer patient sera by ELISA (cat. no. EK106/2&EK1822; MULTI
SCIENCES, Beijing; China) according to the manufacturer's protocol.
The concentrations of IL-6 and TNF-α were calculated based on
standard curves provided with the kits, and the results of IL-6 and
TNF-α were expressed in pg/ml.
Chromatin immunoprecipitation (ChIP)
assay
Cells were processed according to the protocol
described in the ChIP Assay kit (cat no. P2078; Beyotime Institute
of Biotechnology, Haimen, China) with anti-H3K4Me3 antibody (1:100)
(cat no. 07-473; EMD Millipore, Billerica, MA, USA). Magnetic
protein G beads (cat no. 88847; Pierce; Thermo Fisher Scientific,
Inc.) were used to pull-down the antibody-chromatin complexes. The
ChIP primers used were as follows: MASTL pro (0–0.2 K) forward,
5′-TTAAGACTTTCTACAGCTT-3′, and reverse, 5′-TGAGCAGGAAGCGAGTCC-3′.
The NF-κB inhibitor pyrrolidine dithiocarbamate (PDTC; cat. no.
P8765) was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany). The STAT3 inhibitor NSC74859 (cat. no. SD4794) was
purchased from Beyotime (Beyotime Institute of Biotechnology).
Statistical analysis
One-way ANOVA followed by Newman-Keuls post-test was
used for multi-group comparisons. The two-tailed Student's t-test
of SPSS 18.0 (SPSS, Inc., Chicago, IL, USA) was used for two-group
comparisons. All the experiments were repeated at least three
times. Results are displayed as the mean ± standard error of the
mean. P<0.05 was considered to indicate a statistically
significant difference.
Results
Inducible expression of MASTL by
inflammatory cytokines
Cancer of the liver is one of the most common causes
of cancer-associated mortality worldwide (11); in China, the majority of liver cancer
cases occur due to sustained, chronic infection with HBV or HCV
(12,13,20).
Numerous studies have confirmed that inflammatory cytokines,
including IL-6 and TNF-α, secreted by Kupffer cells and
hepatocytes, contribute to the development of hepatocacinogenesis
(21–23). Additionally, these inflammatory
cytokines may activate signal transducer and activator of
transcription 3 (STAT3) and NF-κB to induce hepatocarcinogenesis
(24,25), though the detailed mechanisms are yet
to be clarified.
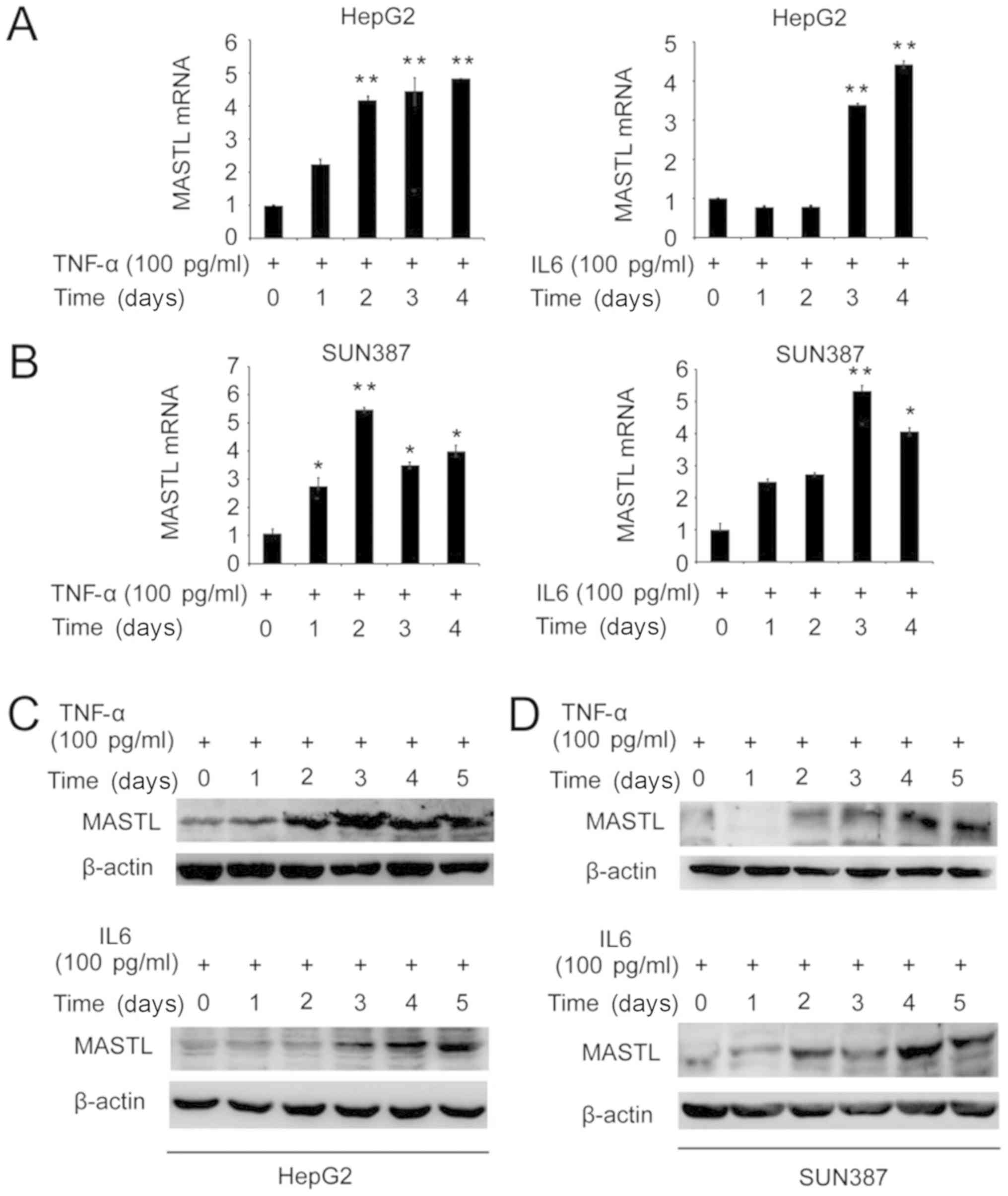
To investigate whether IL-6 or TNF-α may affect the
expression of MASTL in the human liver cancer cell lines
HepG2 and SUN387, these cells were stimulated with 100 pg/ml TNF-α
or IL-6 for 1–5 days. RT-qPCR and western blotting were performed
to determine the mRNA and protein expression levels of MASTL. MASTL
mRNA expression was induced in HepG2 and SUN387 cells by
stimulation with IL-6 or TNF-α (Fig. 1A
and B). Similarly, MASTL protein expression was increased by
the introduction of IL-6 and TNF-α (Fig.
1C and D).
Knockdown of MASTL leads to mitotic
arrest in liver cancer cell lines
A number of studies have demonstrated that Gwl
kinases serve a critical role in regulating mitosis in
Drosophila and Xenopus. In addition, Gwl kinases
inhibit the activation of PP2A by interacting with Ensa/Arpp19
(26,27). Previous studies of MASTL
confirmed that it had a similar function in regulating mitotic
entry and cytokinesis in human cell lines, including HeLa and U2OS
(8).
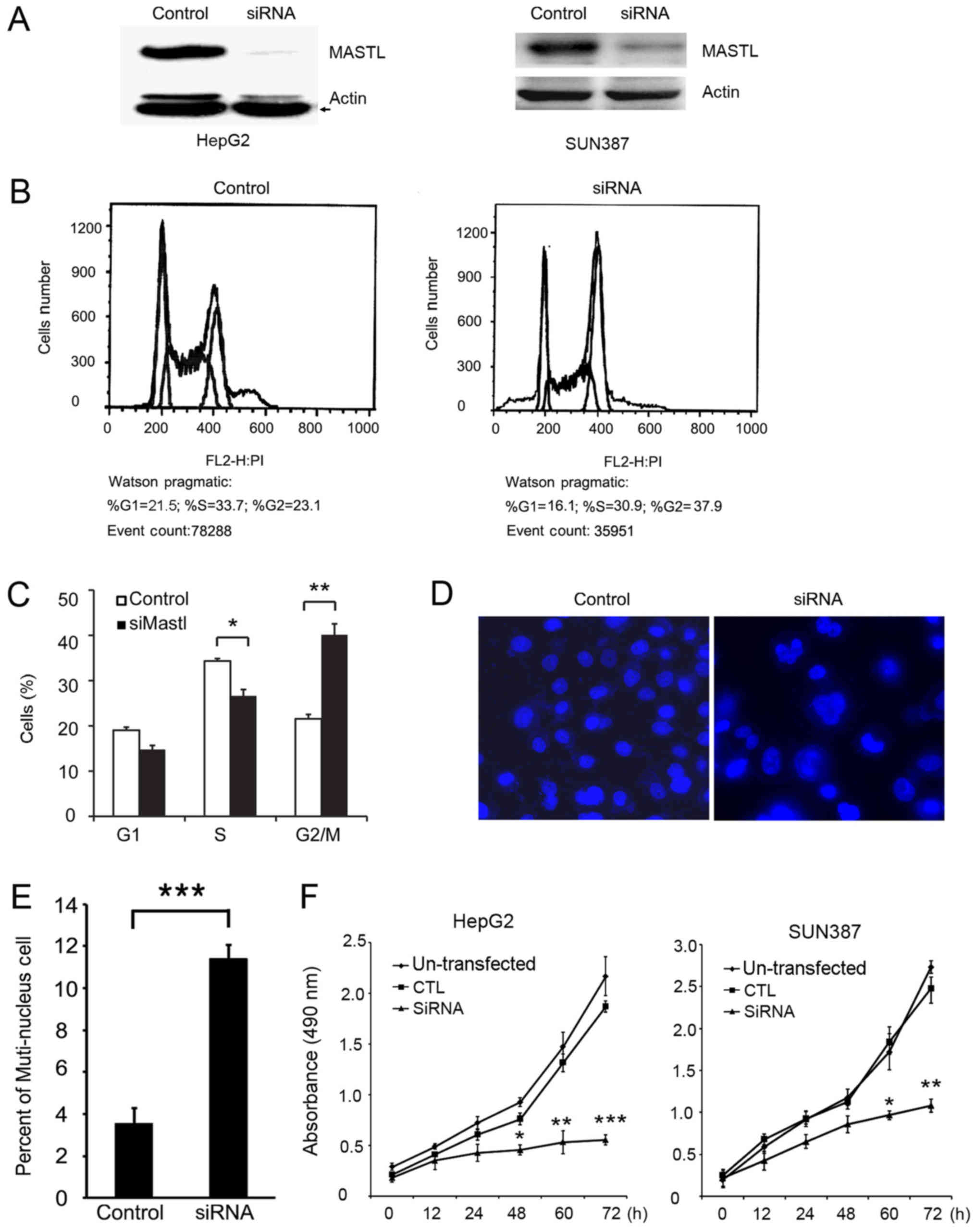
To demonstrate the function of MASTL in HepG2
cells, MASTL was knocked down with siRNA (Fig. 2A). FACS analysis was performed
following PI staining to examine whether MASTL silencing
altered cell cycle progression. The results indicated that cells
transfected with siRNA-MASTL were arrested at the G2/M phase
compared with the control cells (Fig. 2B
and C). Morphological differences were observed by
immunofluorescence. Specifically, HepG2 cells treated with siRNA
exhibited a greater proportion of multinuclear cells compared with
control cells (Fig. 2D and E). This
indicated that the cells failed to enter mitosis and complete cell
division. These results were consistent with previous reports
(8,28), which demenstrated that cells depleted
of MASTL by siRNA remain in the G2 phase, and exhibit slow
chromosomal condensation.
Based on the FACS data, it was hypothesized that
knockdown of MASTL may inhibit liver cancer cell
proliferation. To confirm this hypothesis, an MTT assay was
performed to examine the proliferation of HepG2 and SUN387 cells
transfected with MASTL siRNA or control siRNA, and western blotting
was performed to test the efficiency of silencing in SUN387 cells
(Fig. 2A, right). The results
indicated that the proliferation of MASTL-silenced HepG2 and SUN387
cells was significantly inhibited compared with the control samples
(Fig. 2F), suggesting that
MASTL influences the proliferation of liver cancer
cells.
Overexpression of MASTL promotes cell
proliferation and cell cycle progression
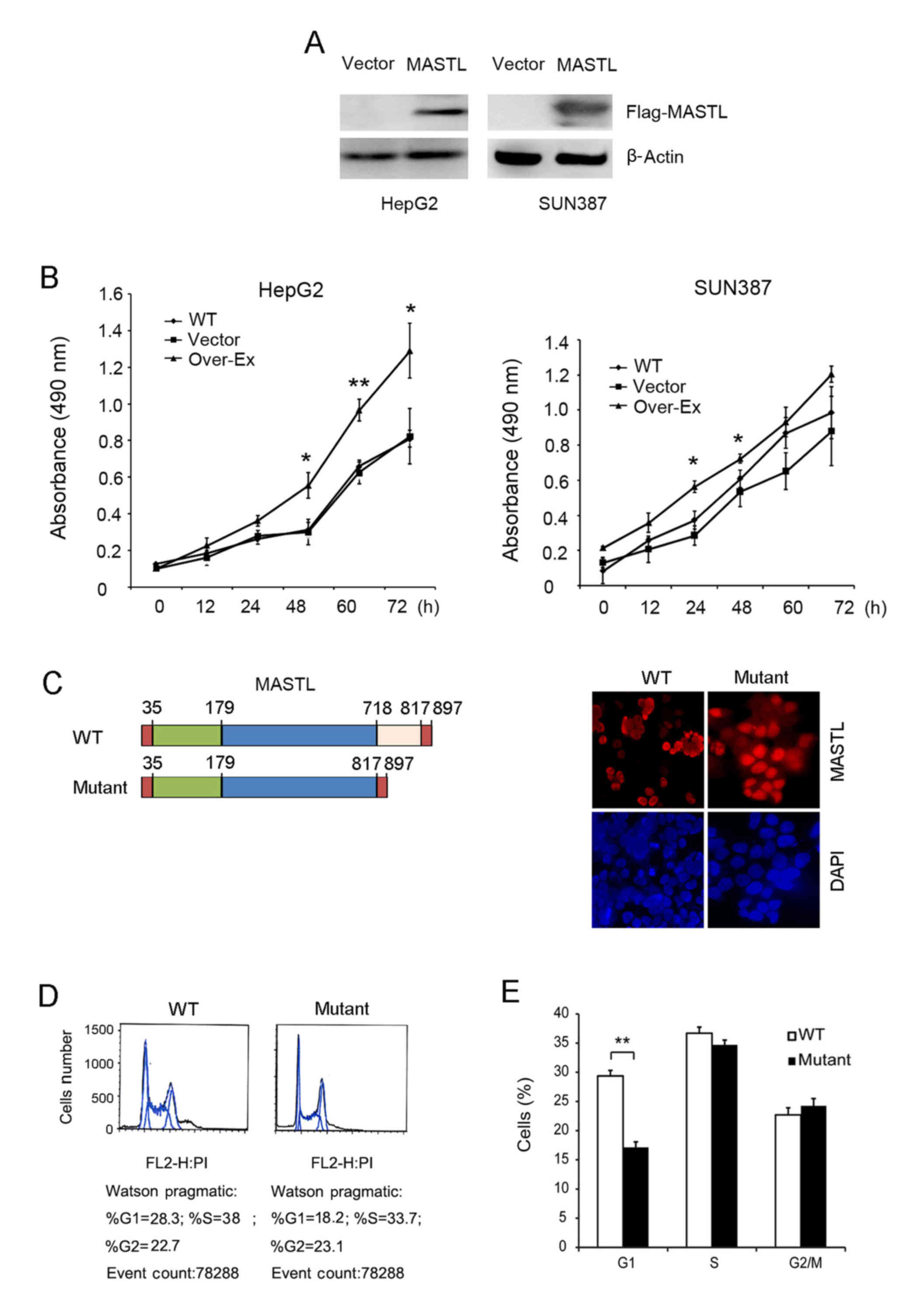
The function of MASTL in cell cycle
progression and cell proliferation was examined by overexpression
in HepG2 and SUN387 cell lines. Exogenous expression of
MASTL was confirmed using an anti-Flag antibody (Fig. 3A), and a subsequent MTT assay revealed
that overexpression of MASTL in HepG2 and SUN387 cells
resulted in more rapid proliferation, compared with the empty
vector controls (Fig. 3B). These
results are partially consistent with the MASTL silencing data in
these cell lines (predominantly in HepG2), further confirming the
role of MASTL in liver cancer proliferation.
Furthermore, MASTL wild type and kinase-dead
vectors were transfected into HepG2 cells to assess alterations in
morphology (Fig. 3C). The nuclei of
HepG2 cells transfected with the wild type MASTL vector were
much smaller, and aggregated together compared with those
transfected with kinase-dead vector, indicating that MASTL
promotes liver cancer cell proliferation. FACS analysis was
performed with cells expressing mutant MASTL and the
corresponding empty vector. The results indicated that the number
of cells expressing mutant MASTL in the G1 phase was significantly
decreased compared with the control (Fig.
3D and E). Taken together, this suggested the involvement of
MASTL in liver cancer cell proliferation by regulating cell
cycle progression. These results were consistent with previous
reports (8,28) which illustrated that MASTL
knockdown impaired cell proliferation.
Increased expression of MASTL in liver
cancer
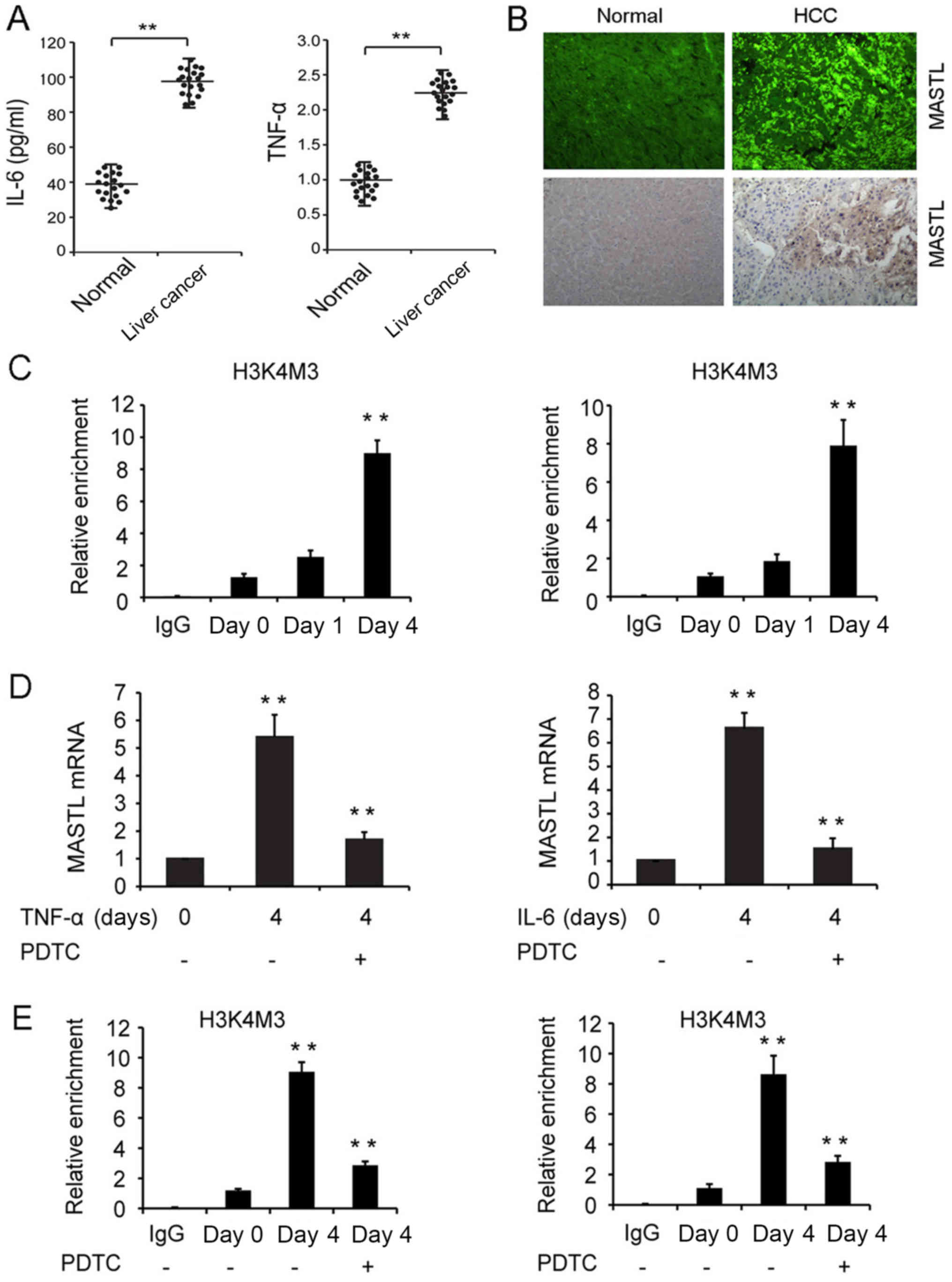
To confirm whether the inflammatory cytokines IL-6
and TNF-α induced MASTL expression in liver cancer tissues,
20 patients with liver cancer and 20 healthy control subjects were
recruited. The results indicated higher concentrations of IL-6 and
TNF-α in the sera of diseased patients compared with those of the
control subjects (Fig. 4A). IHF and
IHC results revealed that 14 of the 20 diseased patients (70%)
exhibited high expression of the MASTL protein, and all of these
patients were chronically infected with HBV. Chronic HBV infection
was determined by persistent HBV surface antigenemia lasting more
than six months. By contrast, 4 of the 20 matched non-tumor liver
tissues (20%) displayed increased MASTL protein expression
(Fig. 4B). Lower expression of the
MASTL protein was revealed in 16 of the 20 non-tumor liver tissues
(Table III). These data indicated
that the high expression of MASTL promoted liver cancer
carcinogenesis.
| Table III.Expression of MASTL protein in liver
cancer and non-tumor liver tissues by immunofluorescence
histochemistry. |
Table III.
Expression of MASTL protein in liver
cancer and non-tumor liver tissues by immunofluorescence
histochemistry.
|
| MASTL expression,
n |
|
|---|
|
|
|
|
|---|
| Histology | High (%) | Low (%) | Total cases, n |
|---|
| Tumor | 14 (70) | 6 (30) | 20 |
| Non-tumor | 4 (20) | 16 (80) | 20 |
IL-6 and TNF-α promote methylation of
H3K4 to facilitate NF-κB-mediated MASTL transcription
The mechanisms involved in the IL-6 and
TNF-α-induced MASTL expression in liver cancer cell lines
were investigated. A ChIP assay was performed to analyze whether
stimulation with cytokines effected the methylation of histone H3K4
at the MASTL promoter, which critically influences the
regulation of transcription by increasing chromatin accessibility.
The results revealed that by day 4, H3K4me3 at the MASTL
promoter was significantly increased in HepG2 cells treated with
IL-6 or TNF-α (Fig. 4C). This
suggested that cytokine stimulation of liver cancer cells promoted
chromatin accessibility at the MASTL promoter, inducing
MASTL expression.
Numerous studies have demonstrated that IL-6 and
TNF-α signaling promotes the proliferation of liver cancer cells by
NF-κB activation (29–32); therefore, the role of NF-κB in IL-6
and TNF-α-induced expression of MASTL was investigated.
HepG2 cells were pretreated with the NF-κB inhibitor PDTC, and the
MASTL mRNA expression level was measured in response to IL-6 or
TNF-α stimulation. The promotion of MASTL mRNA expression was
inhibited by PDTC (Fig. 4D). In
addition, a ChIP assay was performed, which revealed that on day 4,
H3K4me3 at the MASTL promoter was significantly decreased in
HepG2 cells treated with PDTC (Fig.
4E). Thus NF-κB activation was required to induce MASTL mRNA
expression by IL-6 or TNF-α.
In summary, stimulation of liver cancer cells with
IL-6 or TNF-α promoted trimethylation of histone H3 lysine 4 at the
MASTL promoter to facilitate chromatin accessibility, and
NF-κB was involved in cytokine-induced MASTL mRNA expression.
Discussion
Chronic hepatitis as a result of HBV or HCV
infection is associated with cancer of the liver. The
transformation and development of cancer occurs in response to
numerous pathological events, including cell damage, oxidative
stress, compensatory regeneration and proliferation (12). Various immune cells, including
lymphocytes, macrophages, natural killer cells, natural killer T
cells and dendritic cells are involved in chronic
hepatitis-associated liver cancer (27). TNF-α and IL-6 are secreted by immune
cells or hepatocytes, leading to either promotion or inhibition of
multiple hepatocarcinogenesis-associated genes (33). In addition, cell cycle dysregulation
may trigger carcinogenesis, and previous studies have illustrated
that MASTL, the mammalian ortholog of Gwl, has an essential
role in the entry to and exit from mitosis in human cell lines,
including HeLa and U2OS (8). In the
present study, the effects of the proinflammatory cytokines TNF-α
and IL-6 on MASTL expression were investigated in liver
cancer cell lines. The data revealed that mRNA and protein
expression were induced in response to TNF-α or IL-6 stimulation,
suggesting that MASTL serves a prominent role in chronic
hepatitis-associated liver cancer. Moreover, the NF-κB inhibitor
PDTC, and the STAT3 inhibitor NSC74859 were employed to investigate
which signaling pathway may be involved in MASTL expression
in liver cancer cells, in response to IL-6 and TNF-α stimulation.
PDTC, but not NSC74859 influenced MASTL expression,
suggesting a role for NF-κB in this process. The methylation of
H3K4 and H3K36 are associated with transcriptional activation;
however, H3K4me3 is close to the transcriptional start sites of
actively transcribed genes, and as such, is enriched in
transcriptionally active promoters (25,30). In
the present study it was confirmed that the stimulation of liver
cancer cells with IL-6 or TNF-α promoted H3K4Me3 at the
MASTL promoter to facilitate chromatin accessibility.
Further investigation is required to explain how IL-6 and TNF-α
signaling increases the H3K4Me3 level. It was also noted that NF-κB
was involved in the induction of MASTL mRNA expression.
Subsequently, future studies intend to investigate the underlying
mechanisms of NF-κB in this setting, and include the potential role
of NF-κB in methylation of H3K4 at the MASTL promoter.
Previous studies have illustrated that GWL regulates
mitosis in Drosophila and Xenopus eggs (5,34). An RNAi
assay was employed to investigate the function of MASTL in
the liver cancer cell lines HepG2 and SUN387. The results revealed
that HepG2 cells were arrested at the G2/M phase following
silencing of MASTL, and that an increase in the number of
multinuclear cells was apparent, compared with the control. These
results suggested that mitotic regulation of MASTL in human
cells was universal.
Uncontrolled proliferation is one of the hallmark
characteristics of cancer cells. Normal progression of the cell
cycle and subsequent cellular proliferation are gene regulated
(35,36), and abnormal expression of such genes
as cellular tumor antigen p53 results in uncontrolled cell
proliferation (37). The present
study revealed that MASTL had a kinase-dependent influence
on proliferation in liver cancer cell lines (HepG2 and SUN387).
Also, the effects of MASTL on cell proliferation were
confirmed by a marked inhibition following gene silencing. On the
contrary, cell proliferation was promoted by overexpression of
wild-type MASTL, but not the kinase-dead mutant. Further
morphological studies revealed that smaller nuclei were observed
following the overexpression of MASTL. This indicated that
the cells entered mitosis without completing the S phase, resulting
in abnormal cell division.
Though human MASTL has been extensively
studied, and numerous studies indicate its upregulation in human
oral squamous cell carcinoma (38),
little is known of its function in human digestive diseases.
Therefore, the role of MASTL in liver cancer was the focus
of the present study. IHF analysis revealed higher expression of
the MASTL protein in liver cancer tissues compared with those in
non-cancerous liver tissues. Moreover, these results indicated that
MASTL expression was induced by TNF-α and IL-6, which was in
accordance with the increased concentrations of IL-6 and TNF-α in
the sera of liver cancer patients with chronic HBV infection. These
inflammatory cytokines were elevated in chronic hepatitis and liver
cancer, supporting their role in the development of the
disease.
Taken together, the present study has revealed that
the proinflammatory cytokines TNF-α and IL-6 are elevated in
patients with chronic HBV infection. These cytokines induce
MASTL expression by increasing H3K4Me3 and activating NF-κB.
Abnormal expression of MASTL promotes the proliferation of
hepatocytes by regulating mitosis, which subsequently leads to the
carcinogenesis of liver cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Hebei
Province Financial Department Foundation of China (grant no.
361007).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LC, WL, SC and QZ were responsible for the
conceptualization of the study. LC, WL, JY, YW, ZH, YC and JZ
curated the data, and LC, WL, DL, HZ and RZ conducted the formal
analysis. Investigations were conducted by LC, WL, JY, YW, and ZH,
and the methods were designed by LC, WL, DL, YC, HZ, RZ and JZ.
Project administration was the responsibility of LC, WL, SC and QZ.
Resources were provided by SC and QZ, and the study was supervised
by WL, SC and QZ. The original article was written by LC, WL, SC
and QZ, and revised by WL, SC and QZ.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Affiliated Hospital of Hebei University, and
written informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nasmyth K: Viewpoint: Putting the cell
cycle in order. Science. 274:1643–1645. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Murray AW: Recycling the cell cycle:
Cyclins revisited. Cell. 116:221–234. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Davydenko O, Schultz RM and Lampson MA:
Increased CDK1 activity determines the timing of
kinetochore-microtubule attachments in meiosis I. J Cell Biol.
202:221–229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chang HY, Jennings PC, Stewart J, Verrills
NM and Jones KT: Essential role of protein phosphatase 2A in
metaphase II arrest and activation of mouse eggs shown by okadaic
acid, dominant negative protein phosphatase 2A, and FTY720. J Biol
Chem. 286:14705–14712. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu J, Fleming SL, Williams B, Williams EV,
Li Z, Somma P, Rieder CL and Goldberg ML: Greatwall kinase: A
nuclear protein required for proper chromosome condensation and
mitotic progression in Drosophila. J Cell Biol. 164:487–492. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang P, Malumbres M and Archambault V: The
Greatwall-PP2A axis in cell cycle control. Methods Mol Biol.
1170:99–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cundell MJ, Bastos RN, Zhang T, Holder J,
Gruneberg U, Novak B and Barr FA: The BEG (PP2A-B55/ENSA/Greatwall)
pathway ensures cytokinesis follows chromosome separation. Mol
Cell. 52:393–405. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Voets E and Wolthuis RM: MASTL is the
human orthologue of Greatwall kinase that facilitates mitotic
entry, anaphase and cytokinesis. Cell Cycle. 9:3591–3601. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagel R, Stigter-van Walsum M, Buijze M,
van den Berg J, van der Meulen IH, Hodzic J, Piersma SR, Pham TV,
Jiménez CR, van Beusechem VW and Brakenhoff RH: Genome-wide siRNA
screen identifies the radiosensitizing effect of downregulation of
MASTL and FOXM1 in NSCLC. Mol Cancer Ther. 14:1434–1444. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Johnson HJ, Gandhi MJ, Shafizadeh E,
Langer NB, Pierce EL, Paw BH, Gilligan DM and Drachman JG: In vivo
inactivation of MASTL kinase results in thrombocytopenia. Exp
Hematol. 37:901–908. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hoshida Y, Toffanin S, Lachenmayer A,
Villanueva A, Minguez B and Llovet JM: Molecular classification and
novel targets in hepatocellular carcinoma: Recent advancements.
Semin Liver Dis. 30:35–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hartwell LH and Kastan MB: Cell cycle
control and cancer. Science. 266:1821–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chaturvedi P, Eng WK, Zhu Y, Mattern MR,
Mishra R, Hurle MR, Zhang X, Annan RS, Lu Q, Faucette LF, et al:
Mammalian Chk2 is a downstream effector of the ATM-dependent DNA
damage checkpoint pathway. Oncogene. 18:4047–4054. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zeng JZ, Wang HY, Chen ZJ, Ullrich A and
Wu MC: Molecular cloning and characterization of a novel gene which
is highly expressed in hepatocellular carcinoma. Oncogene.
21:4932–4943. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bishayee A: The role of inflammation and
liver cancer. Adv Exp Med Biol. 816:401–435. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hodge DR, Hurt EM and Farrar WL: The role
of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer.
41:2502–2512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pfaffi MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Naugler WE, Sakurai T, Kim S, Maeda S, Kim
K, Elsharkawy AM and Karin M: Gender disparity in liver cancer due
to sex differences in MyD88-dependent IL-6 production. Science.
317:121–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park EJ, Lee JH, Yu GY, He G, Ali SR,
Holzer RG, Osterreicher CH, Takahashi H and Karin M: Dietary and
genetic obesity promote liver inflammation and tumorigenesis by
enhancing IL-6 and TNF expression. Cell. 140:197–208. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
He G, Dhar D, Nakagawa H, Font-Burgada J,
Ogata H, Jiang Y, Shalapour S, Seki E, Yost SE, Jepsen K, et al:
Identification of liver cancer progenitors whose malignant
progression depends on autocrine IL-6 signaling. Cell. 155:384–396.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Iliopoulos D, Hirsch HA and Struhl K: An
epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA, and
IL6 links inflammation to cell transformation. Cell. 139:693–706.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pilati C, Amessou M, Bihl MP, Balabaud C,
Nhieu JT, Paradis V, Nault JC, Izard T, Bioulac-Sage P, Couchy G,
et al: Somatic mutations activating STAT3 in human inflammatory
hepatocellular adenomas. J Exp Med. 208:1359–1366. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mochida S, Maslen SL, Skehel M and Hunt T:
Greatwall phosphorylates an inhibitor of protein phosphatase 2A
that is essential for mitosis. Science. 330:1670–1673. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gharbi-Ayachi A, Labbé JC, Burgess A,
Vigneron S, Strub JM, Brioudes E, Van-Dorsselaer A, Castro A and
Lorca T: The substrate of Greatwall kinase, Arpp19, controls
mitosis by inhibiting protein phosphatase 2A. Science.
330:1673–1677. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Burgess A, Vigneron S, Brioudes E, Labbé
JC, Lorca T and Castro A: Loss of human Greatwall results in G2
arrest and multiple mitotic defects due to deregulation of the
cyclin B-Cdc2/PP2A balance. Proc Natl Acad Sci USA.
107:12564–12569. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell 109 (Suppl). S81–S96. 2002.
|
|
30
|
Xing S, Zhang B, Hua R, Tai WC, Zeng Z,
Xie B, Huang C, Xue J, Xiong S, Yang J, et al: URG4/URGCP enhances
the angiogenic capacity of human hepatocellular carcinoma cells in
vitro via activation of the NF-κB signaling pathway. BMC Cancer.
15:3682015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Tu Q, Yan W, Xiao D, Zeng Z,
Ouyang Y, Huang L, Cai J, Zeng X, Chen YJ and Liu A: CXC195
suppresses proliferation and inflammatory response in LPS-induced
human hepatocellular carcinoma cells via regulating
TLR4-MyD88-TAK1-mediated NF-κB and MAPK pathway. Biochem Biophys
Res Commun. 456:373–379. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu X, Ma P, Shi Y, Yao M, Hou L, Zhang P
and Jiang L: NF-κB increased expression of 17β-hydroxysteroid
dehydrogenase 4 promotes HepG2 proliferation via inactivating
estradiol. Mol Cell Endocrinol. 401:1–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
He G and Karin M: NF-κB and STAT3-key
players in liver inflammation and cancer. Cell Res. 21:159–168.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu J, Zhao Y, Li Z, Galas S and Goldberg
ML: Greatwall kinase participates in the Cdc2 autoregulatory loop
in Xenopus egg extracts. Mol Cell. 22:83–91. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qian Y and Chen X: Tumor suppression by
p53: Making cells senescent. Histol Histopathol. 25:515–526.
2010.PubMed/NCBI
|
|
36
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hermeking H: MicroRNAs in the p53 network:
Micromanagement of tumour suppression. Nat Rev Cancer. 12:613–626.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lorca T and Castro A: The Greatwall
kinase: A new pathway in the control of the cell cycle. Oncogene.
32:537–543. 2013. View Article : Google Scholar : PubMed/NCBI
|