Introduction
Over recent years, the understanding of the causes
of pancreatic cancer (PAC) has improved, but this has not improved
the efficacy of treatment for patients with PAC (1). Surgical resection, which is effective
in alleviating clinical symptoms and results in improved long-term
survival, is only suitable for early pancreatic ductal carcinoma
(2). Although postoperative
radiotherapy and chemotherapy reduce metastatic development, no
notable improvement in patient survival has been observed (3). Therefore, PAC remains among the most
aggressive types of cancer with a 1-year survival rate of ~19% in
the USA, and a 5-year survival rate of only 5% worldwide according
to epidemiological data from 2010 (3). Therefore, there remains a need to
understand the pathogenesis of PAC, identify novel therapeutic
targets and develop new therapeutic strategies (4).
Keratin belongs to the superfamily of intermediate
filament proteins and can be categorized into two major classes:
Type I acidic protein and type II basic protein (5,6). The two
types of keratin provide structural support through the use of
heterodimers to assemble 10-nm filaments, thus maintaining cell
integrity (7). Keratin 17 (KRT17) is
a type I keratin and comprises 432 amino acids (8,9). The
KRT17 gene is located on chromosome 17q21.2 and is mainly
expressed in epithelial appendages, such as hair follicles and
sebaceous glands, but not on healthy skin (10). However, KRT17 expression can be
induced under specific circumstances, such as injury (11), viral infection (12) and psoriasis (13,14).
KRT17 is a multifunctional protein and is involved in the
regulation of cellular processes, including proliferation,
differentiation and inflammation (15). Increasing evidence has suggested that
mutations in KRT17 are associated with pachyonychia congenita type
2 (16–18) and steatocystoma multiplex (19).
Studies have demonstrated that the expression levels
of KRT17 are abnormally high in multiple types of cancer, such as
cervical (20–22), breast (23,24),
gastric (25) and oral (26–28)
cancer and lung adenocarcinoma (29), and have recently been reported in
pancreatic cancer (30). KRT17 is
considered to be a diagnostic and prognostic marker for a variety
of diseases, in particular PAC (12,20,21,31,32).
However, the underlying mechanism of KRT17 function in PAC remains
unknown.
In the present study, the role of KRT17 in PAC and
the potential mechanisms underlying KRT17 function were explored,
as KRT17 inhibition may be a novel target for PAC treatment.
Materials and methods
Human PAC samples
PAC tumor and adjacent non-tumor samples were
obtained from 18 patients (10 male and 8 female patients) that
underwent surgical resection at the Department of Hepatobiliary
Surgery, the First Affiliated Hospital of Wannan Medical College
(Wuhu, China) between November 2016 and December 2018. The Medical
Ethics Committee of the First Affiliated Hospital of Wannan Medical
College (Wuhu, China) approved the experimental protocols in the
present study and written informed consent was obtained from the
patients. All patients included in the present study had no history
of chemoradiotherapy or other pancreatic diseases. The ages of
patients at the time of diagnosis ranged between 46 and 74 years
(mean, 56). All patients were characterized clinically by two
pathologists in accordance with the criteria of the American Joint
Committee on Cancer (33).
Bioinformatics analysis of KRT
expression
KRT17 mRNA expression levels in PAC and non-tumor
tissues were analyzed using the GEPIA website according to the
operating instructions on the website (34). In addition, overall survival and
disease-free survival of patients with PAC were evaluated using
Log-rank test according to the default settings on the GEPIA
website online. The expression of KRT17 protein in PAC and
non-tumor tissues was also compared using The Human Protein Atlas
website (35–37).
Cell culture
The human PAC cell lines MIA PaCa-2, PANC-1 and
KP-3, the human immortalized pancreatic duct epithelial cell line
H6c7 and 293T cells were obtained from Guangzhou Cellcook Cell
Biotechnology, Ltd (http://www.cellcook.com). All cells were maintained in
RPMI-1640 medium (Sangon Biotech Co., Ltd.) containing 10% fetal
bovine serum (FBS; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin/streptomycin (Sangon Biotech Co., Ltd.) and 2%
L-glutamine (Sangon Biotech Co., Ltd.) at 37°C in a 5%
CO2 incubator.
Construction of the KRT17-shRNA
lentivirus
Two short hairpin RNA (shRNA) sequences specifically
targeting KRT17 (GeneBank ID: X62571.1;
5′-CAGTCGCGTTTGCGACTGG-3′ and 5′-CTTCCTTGCCTGATGACAA-3′) and a
scrambled (Scr) shRNA sequence (5′-TTCTCCGAACGTGTCACGT-3′) for use
as a negative control were designed and purchased from iGene
Biotechnology Co., Ltd. The KRT17 or Scr shRNA (2 ng of each
shRNA used) was cloned into the enhanced green fluorescent protein
(eGFP) containing pReceiver-Lv193× lentivector (iGene Biotechnology
Co., Ltd.) using FastDigest KpnI (cat. no. FD0524) and
XhoI (cat. no. FD0694) restriction endonucleases (both
Thermo Fisher Scientific, Inc.) to produce plasmids termed
KRT17-shRNA-1, KRT17-shRNA-2 and Scr-shRNA. A lentivector kit
(Lenti-Pac™; iGene Biotechnology Co., Ltd.) was used to produce
KRT17 or scrambled shRNA. 293T cells were co-infected with
KRT17-shRNA-1, KRT17-shRNA-2 or Scr-shRNA plasmids to produce
KRT17-shRNA-1, KRT17-shRNA-2 or Scr-shRNA lentiviruses. On day 3
post-transfection, the lentivectors were collected and resuspended
in PBS. PANC-1 cells were cultured in 6-well plates
(5×104 cells/well) and infected with KRT17-shRNA-1,
KRT17-shRNA-2 or Scr-shRNA lentivectors in PBS containing 5 µg/ml
polybrene (Merck KGaA). Cells were filtered with 2 µg/ml puromycin
(Sangon Biotech) until they stably expressed KRT17-shRNA or
Scr-shRNA. The same Scr-shRNA sequences were used and performed
simultaneously with the corresponding KRT17-shRNA-1 or KRT-shRNA-2
target sequences, but the two experiments were not performed at the
same time.
Efficiency of KRT17-shRNA
knockdown
Human 293T cells or pancreatic cancer cells were
plated into 6-well plates (5×104 cells/well) and
infected with KRT17-shRNA or Scr-shRNA lentiviruses. Cells were
incubated at 37°C in a 5% CO2 atmosphere for 48 h
(multiplicity of infection, 5), and a suspension of lentivectors (5
µl/well) was added to the RPMI-1640 medium. After infection for 3
days, eGFP expression in the cells was quantified using an inverted
fluorescence microscope (BX63; Olympus Corporation) at ×200
magnification to ascertain infection efficiency. The evaluation
formula of infection efficiency is: Number of cells expressing
EGFP/total number of cells ×100%. Cells with >75% infection
efficiency were collected for further analysis, as described
below.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA from each PAC sample or cultured cells was
extracted using TRIzol® reagent (Sangon Biotech)
according to the manufacturer's instructions and quantified using a
SpectraMax QuickDrop (Molecular Devices, LLC.). Reverse
transcription was performed using a SYBR® Green
Quantitative RT-PCR kit (cat. no. QR0100-1KT; Sigma-Aldrich; Merck
KGaA) according to the manufacturer's instructions. First, 50 ng
RNA template was mixed with 10 µl 2X SYBR Green quantitative RT-PCR
buffer, 1.125 µl each primer (10 µmol/l) and 0.125 µl MMLV RT
Enzyme, and ddH2O was added to a total volume of 25 µl.
The mixture was incubated at 43°C for 30 min for first strand
synthesis. PCR was conducted using the following thermocycling
conditions: Denaturation at 94°C for 30 sec, followed by 40 cycles
of 95°C for 5 sec, 55°C for 15 sec and 72°C for 10 sec. The primer
sequences were designed and synthesized (iGene Biotech) as follows:
KRT17 forward, 5′-AGGTGCGTACCATTGTGGAA-3′ and reverse,
5′-ATCAGGCAAGGAAGCATGGG-3′; GAPDH (internal control) forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′. Other primer sequences are listed in
Table I. Each sample was processed
in triplicate. Data were analyzed using the 2−ΔΔCq
method (38) and normalized against
GAPDH.
 | Table I.Primer sequences used for
quantitative PCR. |
Table I.
Primer sequences used for
quantitative PCR.
| Gene | Primer sequence
(5′-3′) |
|---|
| ERK1/2 |
|
|
Forward |
TTACTGCGCTTCAGACATGAGA |
|
Reverse |
ATCTGTTTCCATGAGGTCCTGT |
| Akt |
|
|
Forward |
CGGGCACATCAAGATAACGGAC |
|
Reverse |
TCGTTGTCCTCCAGCACCTCAG |
| Bad |
|
|
Forward |
CCCAGAGTTTGAGCCGAGTG |
|
Reverse |
GCTGTGCTGCCCAGAGGTT |
| p38 MAPK |
|
|
Forward |
GCTGAAGATTCTGGATTTTG |
|
Reverse |
GTTCTTCCAGTCAACAGCTC |
| SAPK/JNK |
|
|
Forward |
CCGCTGTCCCTTTGTCTTG |
|
Reverse |
GCACATCTATTCTGTTCCATACTACC |
| CAPS3 |
|
|
Forward |
CTGGAAAACCCAAACTTTTCATTA |
|
Reverse |
GCCAGGAAAAGTAACCAGGTGC |
| CAPS7 |
|
|
Forward |
CGGTCCTCGTTTGTACCGTC |
|
Reverse |
CGCCCATACCTGTCACTTTATCA |
| GAPDH |
|
|
Forward |
TGACTTCAACAGCGACACCCA |
|
Reverse |
CACCCTGTTGCTGTAGCCAAA |
Western blotting
Total protein from each sample was extracted using
RIPA lysis buffer (Sigma-Aldrich; Merck KGaA). Protein quantitation
was performed using a QuantiPro™ BCA Assay kit (Sigma-Aldrich;
Merck KGaA). Equal amounts (~10 µg) of total protein from each
sample were loaded onto a 15% gel, resolved using SDS-PAGE and
subsequently transferred to Immobilon®-P PVDF membranes
(EMD Millipore). The membranes were blocked with 1% bovine serum
albumin (BSA; cat. no. SRE0096; Sigma-Aldrich; Merck KGaA)
dissolved in TBS + 0.1% (v/v) Tween-20 (TBST) for 60 min at 37°C
and incubated with the appropriate primary monoclonal antibody
overnight at 4°C. The following mouse primary monoclonal antibodies
were used: Mouse anti-KRT17 (1:1,000; cat. no. WH0003872M1), rabbit
anti-Phospho-p38 MAPK (1:1,000; cat. no. 4511), rabbit anti-p38
MAPK (1:1,000; cat. no. 2387), rabbit anti-Phospho-Akt (1:1,000;
cat. no. 4060), rabbit anti-Akt (1:1,000; cat. no. 4691), or mouse
anti-GAPDH (1:5,000; cat. no. G8795) all from Sigma-Aldrich (Merck
KGaA). After rinsing 3–5 times with TBST, horseradish
peroxidase-conjugated goat anti-mouse IgG (H+L) antibody (1:10,000;
cat. no. AP308P; Sigma-Aldrich; Merck KGaA) was added as the
secondary antibody, incubated at 37°C for 1 h, and then quantified
using an ECL substrate kit (cat. no. ab133406; Abcam). The
densitometry values of the immune response bands were quantified
using ImageJ software (version 1.50; National Institutes of
Health).
MTT assay
Logarithmic phase cells (2,000 cells/well)
transfected with lentiviral KRT17-shRNA-1, KRT17-shRNA-2 or
Scr-shRNA were plated into the wells of 96-well plates and
maintained at 37°C in a 5% CO2 incubator. Cell
proliferation was measured each day for 5 days using a Cell
Proliferation Kit I (Roche Diagnostics) in accordance with the
manufacturer's instructions. Finally, the absorbance at a
wavelength of 490/570 nm in each well of the plate was measured
using a microplate reader (Norgen Biotek Corp.).
Detection of apoptosis and cell
cycle
Apoptosis was quantified using an Annexin V-FITC
Apoptosis Detection kit (Sigma-Aldrich; Merck KGaA) according to
the manufacturer's protocol. After culturing for 4 days, PANC-1
cells that had been infected with lentiviral KRT17-shRNA-1,
KRT17-shRNA-2 or Scr-shRNA were harvested and stained with a
propidium iodide solution (Sigma-Aldrich; Merck KGaA) according to
the manufacturer's instructions to achieve a final density of
1×106 cells/ml. KRT17-shRNA-1-, KRT17-shRNA-2- and
Scr-shRNA-infected PANC-1 cells were added to the wells of a 6-well
plate (1×106 cells/well) and fixed in 70% ethanol for 1
h on ice. A 5 µl aliquot of Annexin V-FITC was added to 100 µl cell
suspension, mixed and incubated for 10 min at room temperature. A
FACSCalibur™ flow cytometer (BD Biosciences) was used to measure
the percentage of apoptotic cells according to the manufacturer's
protocol.
Colony-formation assay
Logarithmic phase cells infected with lentiviral
KRT17-shRNA-1, KRT17-shRNA-2 or Scr-shRNA were seeded into 6-well
plates (800 cells/well) and maintained at 37°C in a 5%
CO2 incubator for 14 days. After discarding the cultured
RPMI-1640 medium, 4% paraformaldehyde (Sigma-Aldrich; Merck KGaA)
was added to each plate (1 ml/well) and incubated for 60 min at
room temperature to fix the cells. After removing the supernatant,
cellular clones were stained with 1 ml/well staining solution from
the Giemsa staining kit (Thermo Fisher Scientific, Inc.) for 1 min
and examined the number of colonies containing >50 cells under a
light microscope (magnification, ×200). Three wells per group were
stained and examined as biological replicates.
Transwell migration assay
A Transwell migration assay was performed to
evaluate cell migration. The wells were coated with 80 µl collagen
I (Thermo Fisher Scientific, Inc.), and 50 µl serum-free RPMI-1640
medium (Thermo Fisher Scientific, Inc.) was added. A total of 220
µl cell suspension (1×105 cells/ml) transfected with
either KRT17-shRNA or Scr-shRNA lentivector were then added to the
upper wells, and RPMI-1640 media with 30% FBS (Sigma-Aldrich; Merck
KGaA) was added to the lower wells. After culture at 37°C for 24 h,
cells in the lower wells were fixed with a 4% formaldehyde solution
(Sigma-Aldrich; Merck KGaA) for 15 min and stained with 1% crystal
violet (Sigma-Aldrich; Merck KGaA) at room temperature for 30 min.
Cells were counted using a light microscope and each experiment in
each group was performed in triplicate.
Tumorigenesis analysis
A total of 20 nude mice (18-22 g) were purchased
from the Institute of Comparative Medicine of Yangzhou University
(Yangzhou, China). The animal experiments were approved by The
First Affiliated Hospital of Wannan Medical College & Yijishan
Hospital Animal Experimental Ethics Committee (Wuhu, China). All
animals were kept in an SPF animal room at 37°C and 60–70%
humidity, with a 12-h light/dark cycle and free access to food and
water. Tumorigenesis was assessed using nude mice as previously
described (39). PANC-1 cells
(1×105 cells in 200 µl PBS per mouse) transfected with
KRT17-shRNA-1 or Scr-shRNA lentivector were injected subcutaneously
or below the armpit into female BALB/c mice (4-weeks old). Five
weeks after the initial injection, the dimensions of the tumors
were measured every three days and volumes were calculated using
the following formula: Tumor volume=(π/6) × L × W × W, where L
represented the longest diameter and W was the shortest diameter.
At the time of sacrifice, the mean weight of each excised tumors
was calculated.
Statistical analysis
Three separate experiments were conducted for each
condition. Data are presented as means ± SD. Statistical tests were
performed using GraphPad Prism software for Windows (version 8.0.1;
GraphPad Software, Inc.). Overall survival (OS) or disease free
survival (DFS) was performed by Log-rank test. Multiple group
comparisons were performed using least-significant difference (LSD)
as a post-hoc method after one-way analysis of variance, and
comparisons between two groups in all figures were performed using
student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
KRT17 expression in human pancreatic
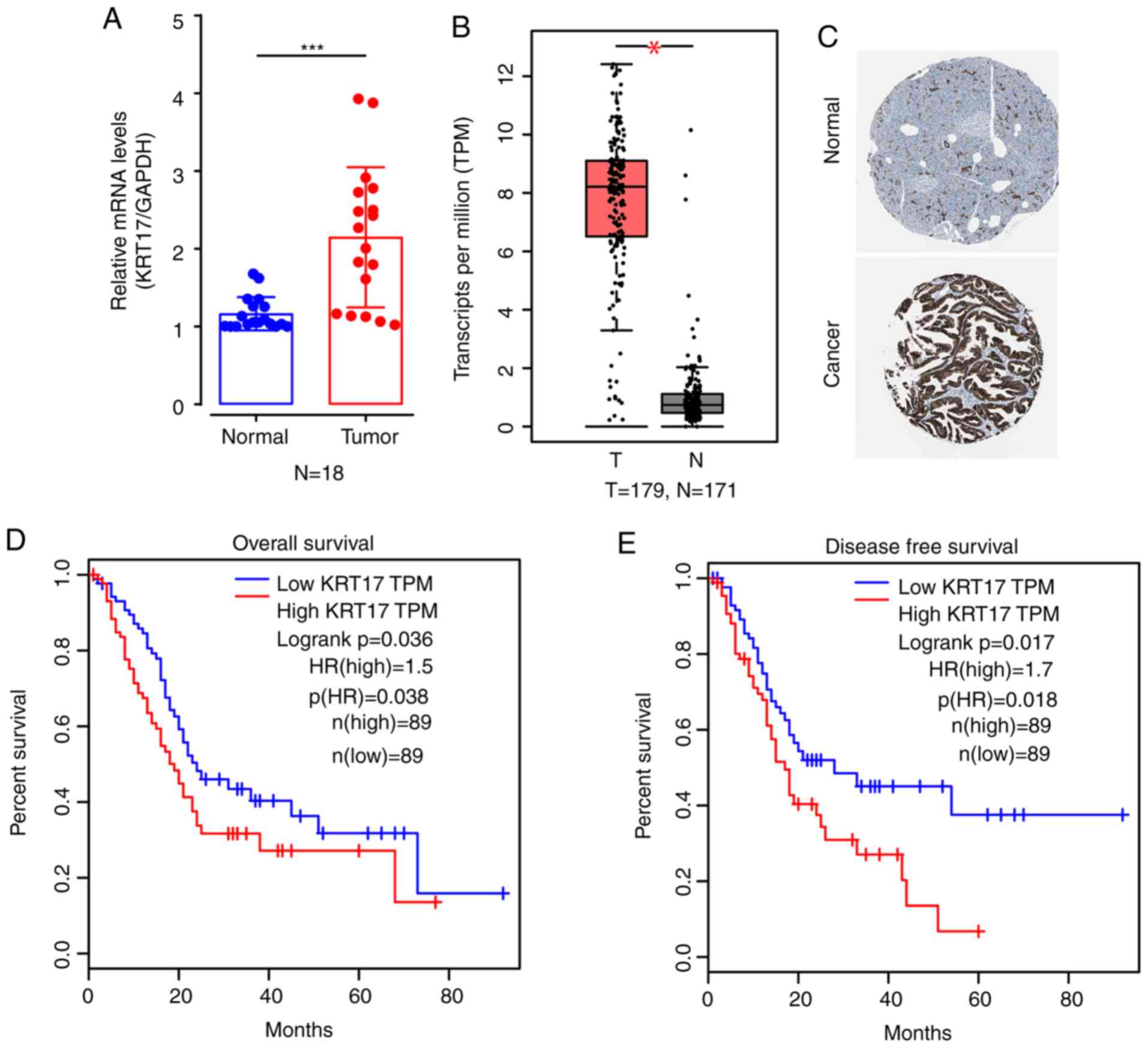
cancer samples
To investigate KRT17 expression in human PAC
tissue and matched adjacent non-tumor (normal) tissue, 18 samples
were randomly selected from a total of 18 human PAC specimens for
qPCR. The results indicated that the average expression levels of
KRT17 mRNA in PAC tissues were significantly elevated compared with
normal tissues (P<0.001; Fig.
1A). Bioinformatics results from historical samples in The
Human Protein Atlas also suggested that, compared with normal
tissues, KRT17 mRNA levels in PAC tissues were higher (P<0.05;
Fig. 1B), and the protein expression
levels of KRT17 were higher in PAC compared with normal tissues
based on the immunohistochemical staining results from The Human
Protein Atlas website (Fig. 1C). In
addition, bioinformatics analysis suggested that high expression of
KRT17 in PAC tissues was negatively associated with overall
survival (log-rank P=0.036; Fig. 1D)
and disease-free survival (log-rank P=0.017; Fig. 1E).
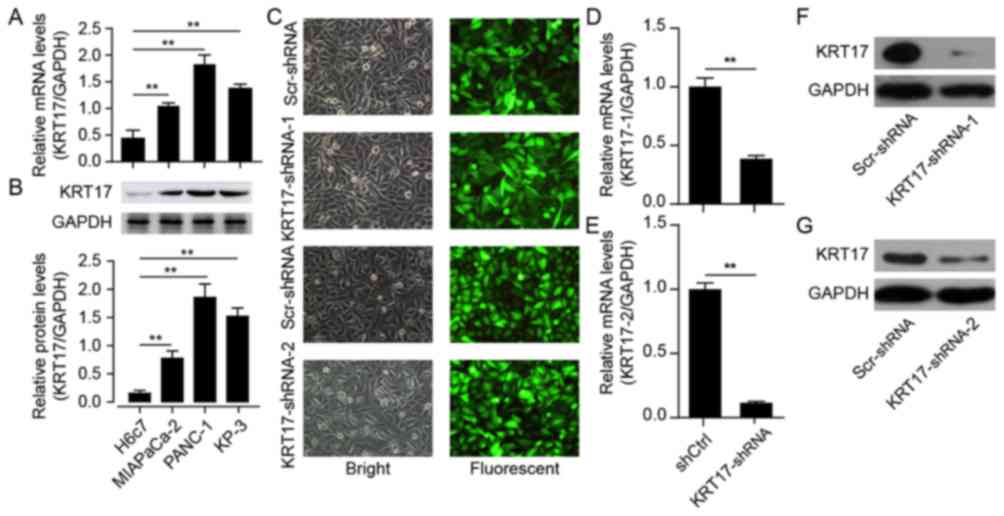
Identification of lentivirus-mediated
shRNA for human KRT17 silencing
To explore the functional role of KRT17 in PAC both
in vitro and in vivo, the expression levels of KRT17
mRNA were initially examined in three human pancreatic cancer cell
lines: MIA PaCa-2, PANC-1 and KP-3, in addition to its expression
levels in the pancreatic duct epithelial cell line H6c7. The
results demonstrated that KRT17 mRNA levels were elevated in the
human PAC cell lines MIA PaCa-2, PANC-1 and KP-3 compared with H6c7
cells (Fig. 2A) and confirmed by
western blotting (Fig. 2B).
Subsequently, PANC-1 cells were selected for follow-up studies, and
a lentivirus designed to silence KRT17 expression in PANC-1
cells was constructed for functional analysis. The results
demonstrated that GFP expression levels in PANC-1 cells transfected
with KRT17-shRNA-1-, KRT17-shRNA-2- or Scr-shRNA-lentivirus
indicated high rates of infection efficiency, >90% after 48 h
(Fig. 2C). qPCR results indicated
that the silencing efficiency of transfection of KRT17-shRNA in
PANC-1 cells was ~60–80% (Fig. 2D and
E), further confirmed by western blot analysis (Fig. 2F and G).
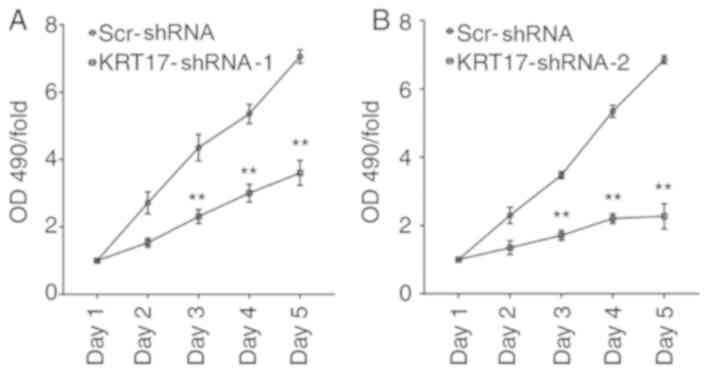
Knockdown of KRT17 significantly
reduces cell proliferation of PANC-1 cells
To demonstrate the effect of KRT17-shRNA in
vitro, PANC-1 cells were infected with lentiviral
KRT17-shRNA-1, KRT17-shRNA-2 or Scr-shRNA. Cell viability was
monitored every day for five days using an MTT assay. The results
demonstrated that KRT17-shRNA significantly decreased cell
viability 72 h post-transfection with KRT17-shRNA-1 or
KRT17-shRNA-2 lentivirus and that cell viability decreased over the
following two days compared with cells in the Scr-shRNA group
(P<0.01; Fig. 3A and B). Taken
together, these results suggested that KRT17 promoted pancreatic
cancer cell progression via accelerated cell proliferation.
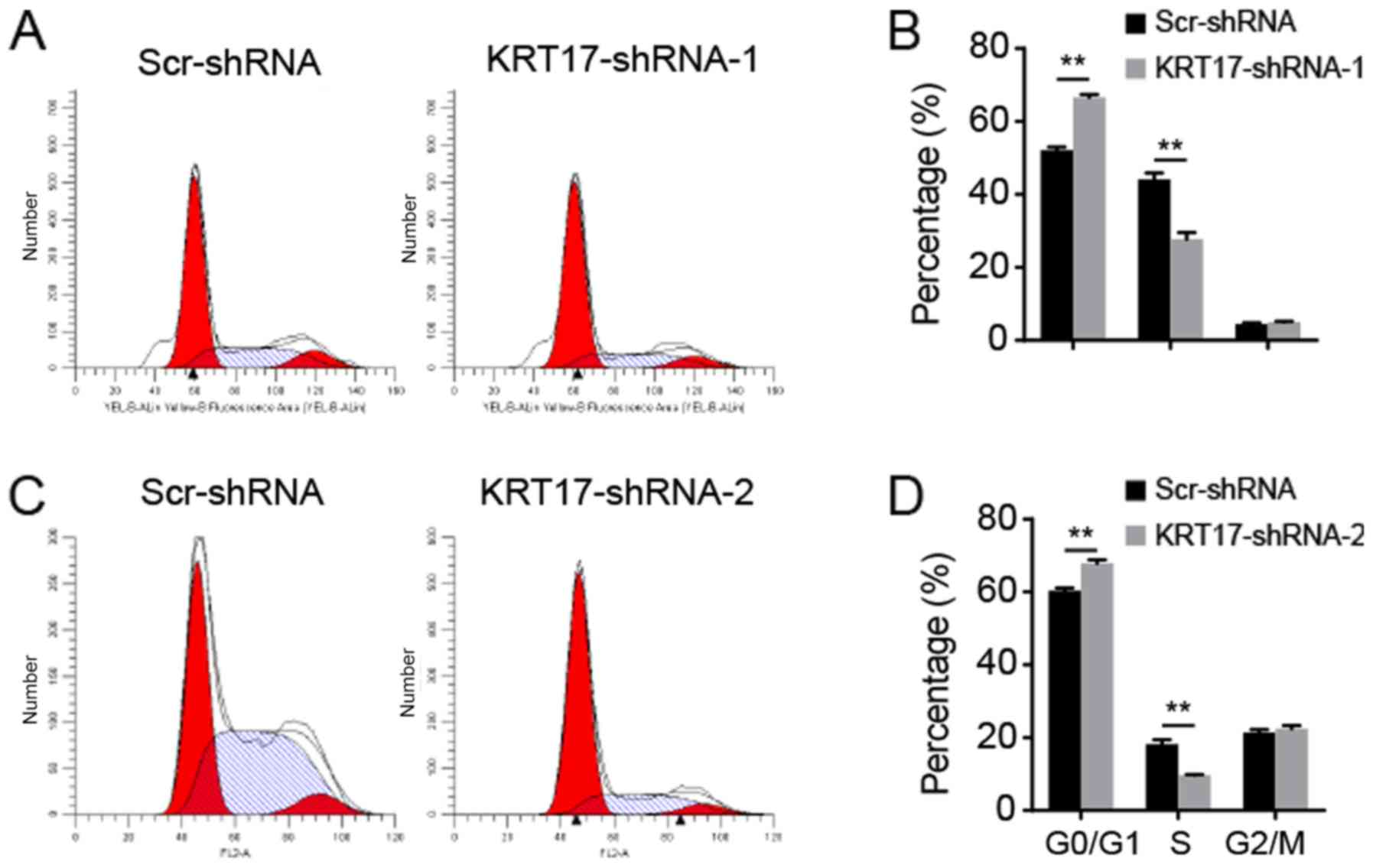
KRT17 knockdown induces cell cycle
arrest in PANC-1 cells
As KRT17-shRNA significantly inhibited PANC-1 cell
proliferation, the underlying molecular mechanism was investigated
using flow cytometry to analyze the cell cycle. The results
indicated that following transfection with the KRT17-shRNA-1
lentivirus, 65.58±2.02, 27.71±1.44 and 5.21±0.54% of cells were in
the G0/G1, S and G2/M phase,
respectively [Fig. 4A (upper panel) and
B]. Following transfection with the Scr-shRNA lentivirus,
52.28±0.99, 44.56±3.05 and 5.27±0.95% of cells were in the
G0/G1, S and G2/M phase,
respectively (Fig. 4B).
Additionally, following transfection with the KRT17-shRNA-2
lentivirus, 67.92±1.61, 9.52±0.93 and 22.8±1.08% of cells were in
the G0/G1, S and G2/M phase,
respectively [Fig. 4A (bottom panel) and
C]. Following transfection with the Scr-shRNA lentivirus,
60.78±1.23, 18.77±1.63 and 21.21±1.63% of the cells transfected
with KRT17-shRNA-2 lentivirus were in the
G0/G1, S and G2/M phase,
respectively (Fig. 4C). These
results suggested that inhibition of KRT17 expression resulted in
cell cycle arrest in the G0/G1 phase compared
with the Scr-shRNA group cells (all P<0.01; Fig. 4).
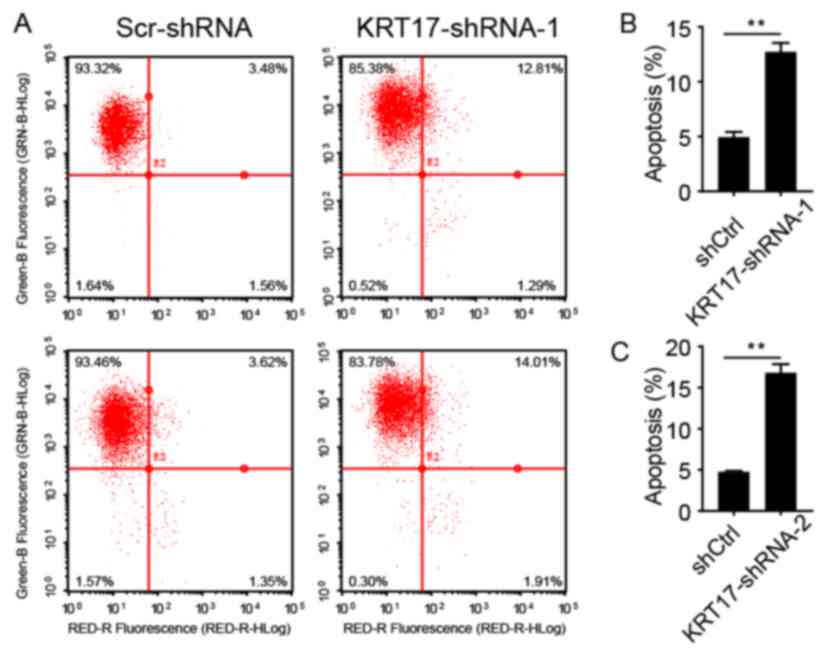
KRT17 silencing increases PANC-1 cell
apoptosis
To analyze whether KRT17 silencing was associated
with apoptosis in PANC-1 cells, infected PANC-1 cells were sorted
using flow cytometry after treatment with FITC-conjugated Annexin
V. Cells were infected with lentiviral KRT17-shRNA-1, KRT17-shRNA-2
or Scr-shRNA and sorted using flow cytometry after 5 days shown in
a representative picture in Fig. 5A.
As presented in Fig. 5B and C,
5.26±0.09 and 4.70±0.23% of cells infected with Scr-shRNA
lentivirus were apoptotic, but in PANC-1 cells infected with
KRT17-shRNA-1 or KRT17-shRNA-2 lentivirus, the proportion of
apoptotic was increased to 12.75±0.90 and 16.79±1.00%,
respectively, compared with the control cells (P<0.01). These
results demonstrated that KRT17 knockdown increased apoptosis in
human PANC-1 cells.
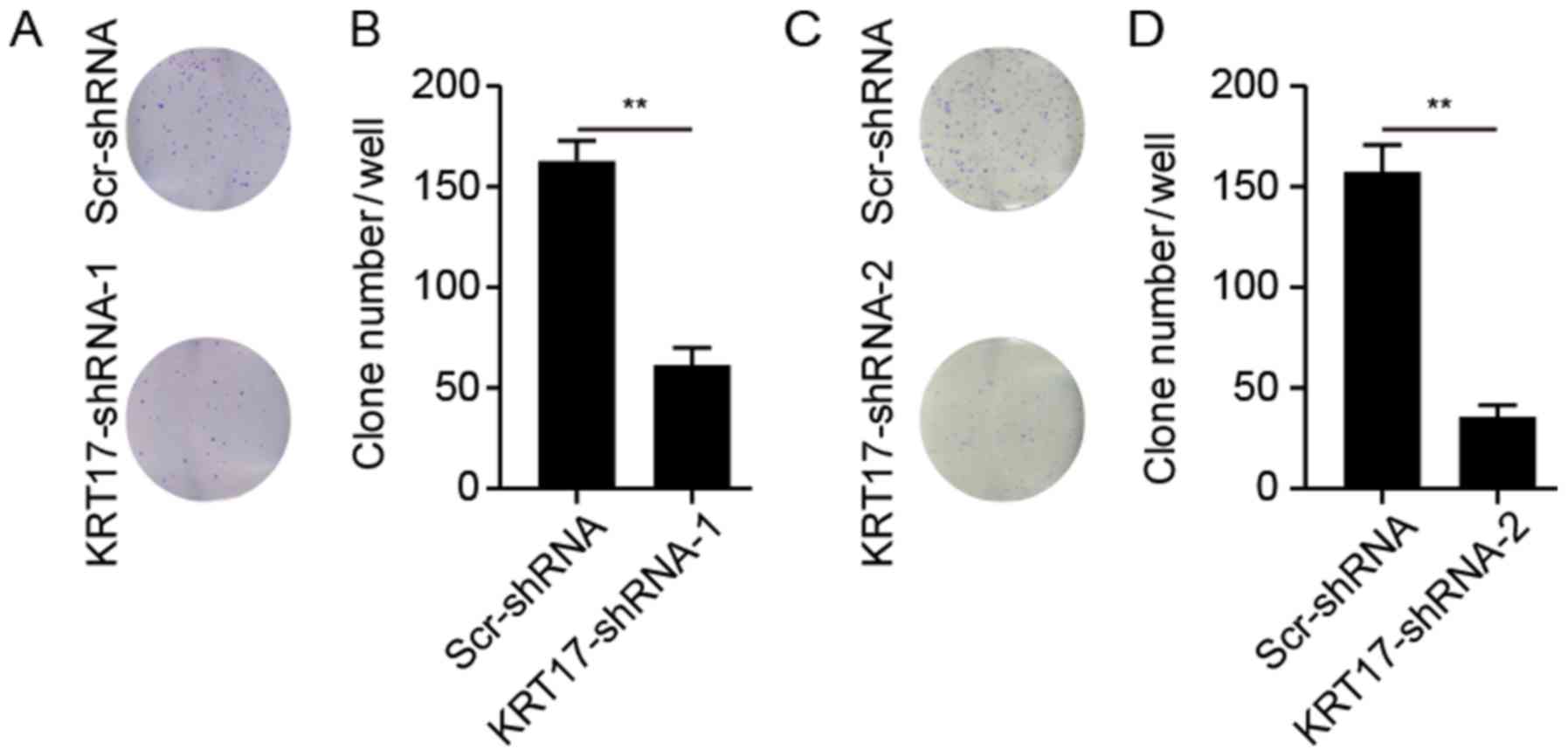
Knockdown of KRT17-shRNA inhibits
colony formation capability
The ability of PANC-1 cells to form colonies
following treatment with KRT17-shRNA or Scr-shRNA lentivirus was
explored. As presented in Fig. 6,
KRT17 silencing significantly reduced the number of colonies (62±6
colonies/well in PANC-1 cells transfected with KRT17-shRNA-1; 37±4
colonies/well in PANC-1 cells transfected with KRT17-shRNA-2)
compared with 164±8 and 157±18 colonies/well in PANC-1 cells in the
Scr-shRNA groups (P<0.01).
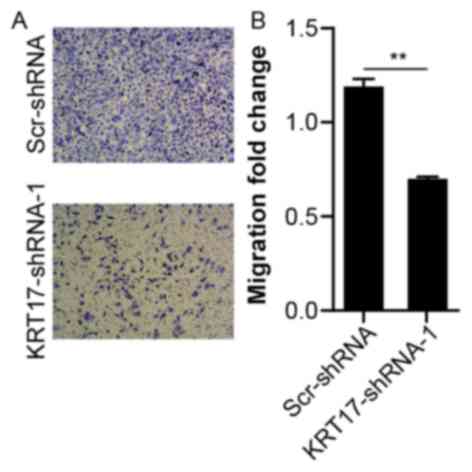
Knockdown of KRT17 inhibits the
migration of PANC-1 cells
To investigate the effects of KRT17 knockdown on the
migration of human PANC-1 cells, the migration capability of PANC-1
cells transfected with KRT17-shRNA-1 or Scr-shRNA lentivirus was
measured using a Transwell assay. Compared with the Scr-shRNA
group, the number of migratory cells in the KRT17-shRNA-1 group was
significantly lower (P<0.01, Fig. 7A
and B), suggesting that knockdown of KRT17 resulted in reduced
PANC-1 cell migration. This finding further emphasized the function
of KRT17 in the migration of PANC-1 cells.
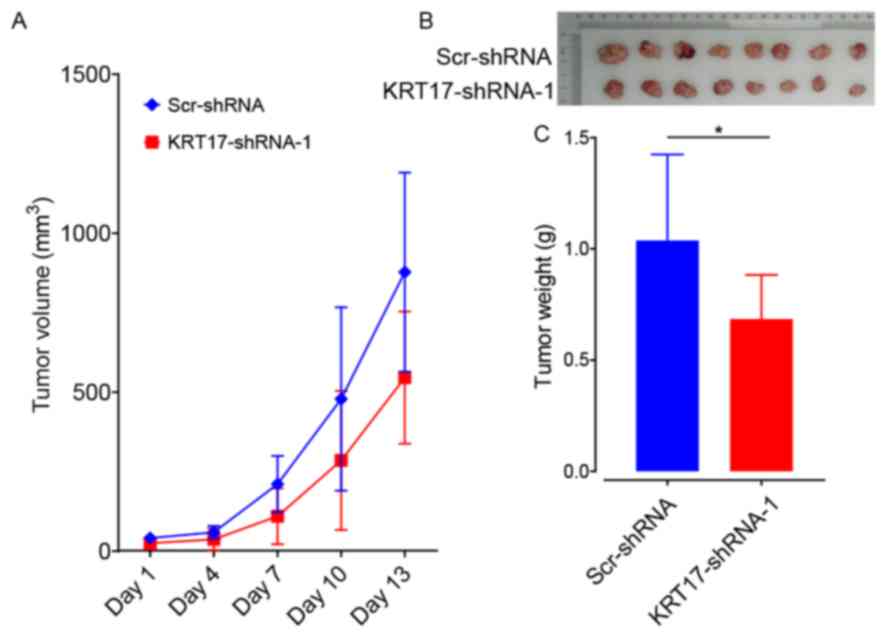
KRT17 silencing suppresses tumor
growth in vivo
To ascertain the role of KRT17-shRNA as a potential
target for the treatment of PAC, BALB/c nude mice were
subcutaneously injected with PANC-1 cells transfected with either
KRT17-shRNA-1 or Scr-shRNA lentivector. The results demonstrated
that, compared with the Scr-shRNA group, tumors grew more slowly in
the KRT17-shRNA group (Fig. 8A). In
addition, tumors from mice in the KRT17-shRNA group were also
smaller compared with the Scr-shRNA-group (Fig. 8B and C). This result showed that
KRT17 silencing inhibited PAC tumorigenesis in vivo. Taken
together, these data indicate that targeting KRT17 with
lentiviral-KRT17-shRNA could have an inhibitory effect in
vivo on pancreatic cancers in which KRT17 is overexpressed.
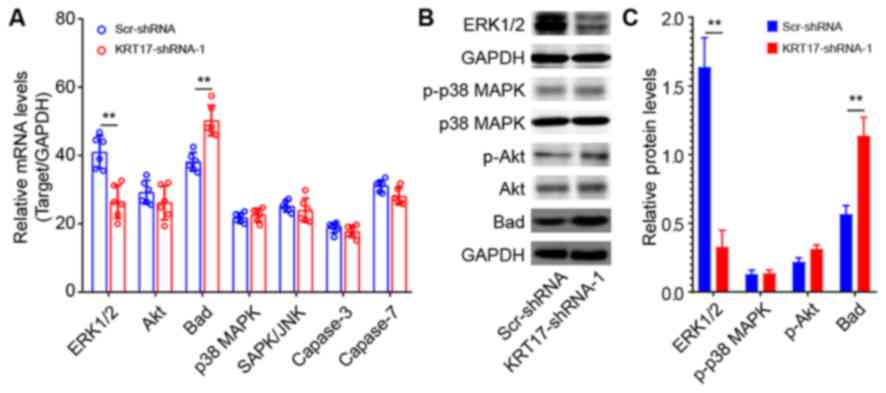
KRT17 silencing in PANC-1 cells
affects ERK1/2/Bad signaling
To investigate the underlying signaling mechanisms
of the effect of KRT17 silencing in PANC-1 cells, the expression
levels of signaling molecules relevant to cell proliferation and
apoptosis were analyzed using qPCR, including ERK1/2, Akt, Bad, p38
MAPK, SAPK/JNK, caspase-3 and caspase-7. The results demonstrated
that ERK1/2 was significantly downregulated in KRT17-shRNA-treated
PANC-1 cells compared with Scr-shRNA-treated cells (P<0.01;
Fig. 9A). By contrast, Bad was
significantly upregulated compared with the Scr-shRNA group
(P<0.01). Western blot analysis further confirmed that ERK1/2
expression levels were decreased, whereas Bad expression levels
were increased in the KRT17-shRNA group compared with the Scr-shRNA
group. In addition, KRT17 knockdown exhibited no effects on the
expression of p38 MAPK, Akt and phospho-Akt.
Discussion
The results of the present study demonstrated that
KRT17 expression levels were elevated in human PAC samples compared
with normal tissues. Bioinformatics analysis also demonstrated high
expression levels of KRT17 mRNA and protein in PAC samples compared
with normal tissues. In addition, elevated expression levels of
KRT17 were associated with the risk of PAC progression, including
less favorable overall and disease-free survival.
Using qPCR and western blotting, the present study
demonstrated that KRT17 expression levels were elevated in three
PAC cell lines: MIA PaCa-2, PANC-1 and KP-3. Cell proliferation and
colony formation of PANC-1 cells were significantly diminished in
KRT17-shRNA-transfected cells, whereas the percentage of those
cells that had undergone apoptosis was significantly higher
compared with the proportion of Scr-shRNA-transfected cells. In
addition, KRT17 silencing significantly reduced tumor growth in
vivo. These data suggested that elevated KRT17 expression
enhanced the development of PAC. Of note, the results of the
present study revealed that KRT17 silencing arrested the cell cycle
at the G0/G1 phase, suggesting that KRT17
augmented human PAC cell growth and colony formation by inhibiting
the cell cycle. However, the underlying mechanism of the cell cycle
arrest in KRT17-knockdown PANC-1 cells requires additional
investigation in future studies.
KRT17 is primarily expressed in the epidermis and
its appendages (40). Previous
studies have demonstrated that abnormal expression levels of KRT17
are associated with cell proliferation and tumor development. For
example, high expression levels of KRT17 promote cell proliferation
and tumorigenesis in skin tumors (41), oral squamous cell carcinoma (22,26,42),
cervical squamous cell carcinoma (20,21,43,44) and
gastric cancer (25). However,
reports on the functional role of KRT17 in pancreatic cancer are
limited. A recent study has confirmed that high expression of KRT17
is positively associated with less favorable prognosis in patients
with pancreatic cancer (45).
However, the underlying mechanism of this has not been reported.
The present study also revealed aberrantly increased expression
levels of KRT17 in PAC samples, which indicated that KRT17 may
serve a role in PAC tumorigenesis.
In the present study, the role of KRT17 in human PAC
was analyzed using transfection of PANC-1 cells with KRT17-shRNA.
The results demonstrated that KRT17 silencing suppressed cell
proliferation and facilitated apoptosis in human PANC-1 cells. In
addition, KRT17 knockdown decreased the migratory ability of PANC-1
cells. The results indicated that abnormally high expression levels
of KRT17 were implicated in human PAC progression and may be
associated with advanced tumor stage and distant metastasis of
pancreatic cancer, as observed in the previous results of this
study.
To explore the effects of KRT17 knockdown in human
PANC-1 cells, the changes in the expression levels of a number of
stress and apoptosis molecules were analyzed using qPCR and/or
western blot analysis. The results demonstrated that ERK1/2 was
significantly downregulated following lentiviral
transfection-mediated KRT17 knockdown, and the expression levels of
the Bad protein were significantly increased. Accumulating evidence
suggests that ERK1/2 activation contributes to the
epithelial-mesenchymal transition (EMT) in intrahepatic
cholangiocarcinoma (46), although
inhibition of the ERK1/2 pathway has failed to prevent transforming
growth factor β-induced EMT and migration. The present study
demonstrated that KRT17 knockdown decreased the level of ERK1/2
phosphorylation, suggesting that upregulation of KRT17 participated
in EMT and migration in PAC tumorigenesis. In the majority of
cancer types, ERK1/2 signaling promotes cell survival by inhibiting
the function of pro-death proteins, including Bad (47). In the present study, Bad
phosphorylation increased following KRT17 silencing using
KRT17-shRNA lentivirus transfection. These results suggested that
KRT17 may increase PAC tumorigenesis via ERK1/2/Bad signaling.
The present study demonstrated that KRT17 expression
levels were elevated in PAC tissues and pancreatic cancer cell
lines, MIA PaCa-2, PANC-1 and KP-3 cells. Lentivirus-mediated KRT17
knockdown inhibited cell proliferation and clone formation by
arresting the cells in the G0/G1 phase and by
accelerating apoptosis. The role of KRT17 may be to augment
pancreatic cancer tumorigenesis via upregulation of ERK1/2 and
downregulation of Bad expression. These results suggested that
KRT17 may serve a role in PAC tumorigenesis and may be useful as a
potential therapeutic target for the treatment of PAC.
Acknowledgements
Not applicable.
Funding
This study was supported by the Academic Funding
Project for Top Talents of Discipline (Professional) in Anhui
Province (grant no. gxbjZD17), Anhui Provincial Centralized Local
Science and Technology Development Special Project (grant no.
YDZX20183400004899) and Anhui Science and Technology Research Fund
Project (grant no. 1501041156).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GW and XW designed the study. PC, ZS and XF
performed the cytology experiments. PC and SX conducted the animal
experiments. XF, GW and JW analyzed the data. PC and XF wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The First
Affiliated Hospital of Wannan Medical College and Yijishan Hospital
Medical Ethics Committee (Wuhu, China). Written informed consent
was provided by all patients. Animal experiments were approved by
The First Affiliated Hospital of Wannan Medical College &
Yijishan Hospital Animal Experimental Ethics Committee (Wuhu,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Saad AM, Turk T, Al-Husseini MJ and
Abdel-Rahman O: Trends in pancreatic adenocarcinoma incidence and
mortality in the United States in the last four decades; a
SEER-based study. BMC Cancer. 18:6882018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hu D, Ansari D, Zhou Q, Sasor A, Said
Hilmersson K and Andersson R: Galectin 4 is a biomarker for early
recurrence and death after surgical resection for pancreatic ductal
adenocarcinoma. Scand J Gastroenterol. 54:95–100. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Falasca M, Kim M and Casari I: Pancreatic
cancer: Current research and future directions. Biochim Biophys
Acta. 1865:123–132. 2016.PubMed/NCBI
|
|
5
|
van den Hurk R, Dijkstra G, van Mil FN,
Hulshof SC and van den Ingh TS: Distribution of the intermediate
filament proteins vimentin, keratin, and desmin in the bovine
ovary. Mol Reprod Dev. 41:459–467. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Somji S, Cao L, Mehus A, Zhou XD, Sens MA,
Dunlevy JR, Garrett SH, Zheng Y, Larson JL and Sens DA: Comparison
of expression patterns of keratin 6, 7, 16, 17, and 19 within
multiple independent isolates of As(+3)- and Cd (+2)-induced
bladder cancer: Keratin 6, 7, 16, 17, and 19 in bladder cancer.
Cell Biol Toxicol. 27:381–396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jacob JT, Coulombe PA, Kwan R and Omary
MB: Types I and II keratin intermediate filaments. Cold Spring Harb
Perspect Biol. 10(pii): a0182752018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Herrmann H and Aebi U: Intermediate
filaments: Molecular structure, assembly mechanism, and integration
into functionally distinct intracellular Scaffolds. Annu Rev
Biochem. 73:749–789. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Herrmann H and Aebi U: Intermediate
filaments: Structure and assembly. Cold Spring Harb Perspect Biol.
8(pii): a0182422016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kurokawa I, Takahashi K, Moll I and Moll
R: Expression of keratins in cutaneous epithelial tumors and
related disorders-distribution and clinical significance. Exp
Dermatol. 20:217–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim S, Wong P and Coulombe PA: A keratin
cytoskeletal protein regulates protein synthesis and epithelial
cell growth. Nature. 441:362–365. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Proby CM, Churchill L, Purkis PE, Glover
MT, Sexton CJ and Leigh IM: Keratin 17 expression as a marker for
epithelial transformation in viral warts. Am J Pathol.
143:1667–1678. 1993.PubMed/NCBI
|
|
13
|
Shi X, Jin L, Dang E, Chang T, Feng Z, Liu
Y and Wang G: IL-17A upregulates keratin 17 expression in
keratinocytes through STAT1- and STAT3-dependent mechanisms. J
Invest Dermatol. 131:2401–2408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang W, Dang E, Shi X, Jin L, Feng Z, Hu
L, Wu Y and Wang G: The pro-inflammatory cytokine IL-22
up-regulates keratin 17 expression in keratinocytes via STAT3 and
ERK1/2. PLoS One. 7:e407972012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang L, Zhang S and Wang G: Keratin 17 in
disease pathogenesis: From cancer to dermatoses. J Pathol.
247:158–165. 2019.PubMed/NCBI
|
|
16
|
Ghazawi FM, Hassani-Ardakani K, Henriques
L and Jafarian F: Identification of a novel substitution mutation
(R103C) in the rod domain of the keratin 17 gene associated with
pachyonychia congenita type 2. Int J Dermatol. 58:233–236. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Agarwala M, Salphale P, Peter D, Wilson
NJ, Pulimood S, Schwartz ME and Smith FJD: Keratin 17 mutations in
four families from India with pachyonychia congenita. Indian J
Dermatol. 62:422–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ofaiche J, Duchatelet S, Fraitag S, Nassif
A, Nougue J and Hovnanian A: Familial pachyonychia congenita with
steatocystoma multiplex and multiple abscesses of the scalp due to
the p.Asn92Ser mutation in keratin 17. Br J Dermatol.
171:1565–1567. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zang D, Zhou C, He M, Ma X and Zhang J: A
novel mutation (p.Arg94Gly) of keratin 17 in a Chinese family with
steatocystoma multiplex. Eur J Dermatol. 21:142–144.
2011.PubMed/NCBI
|
|
20
|
Mockler D, Escobar-Hoyos LF, Akalin A,
Romeiser J, Shroyer AL and Shroyer KR: Keratin 17 is a prognostic
biomarker in endocervical glandular neoplasia. Am J Clin Pathol.
148:264–273. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Escobar-Hoyos LF, Yang J, Zhu J, Cavallo
JA, Zhai H, Burke S, Koller A, Chen E and Shroyer KR: Keratin 17 in
premalignant and malignant squamous lesions of the cervix:
Proteomic discovery and immunohistochemical validation as a
diagnostic and prognostic biomarker. Mod Pathol. 27:621–630. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mikami Y, Fujii S, Nagata K, Wada H,
Hasegawa K, Abe M, Yoshimoto RU, Kawano S, Nakamura S and Kiyoshima
T: GLI-mediated keratin 17 expression promotes tumor cell growth
through the anti-apoptotic function in oral squamous cell
carcinomas. J Cancer Res Clin Oncol. 143:1381–1393. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kirjushkina MS, Tchipysheva TA, Ermilova
VD and Guelstein VI: Breast tumor diagnosis in cytologic aspirates
using monoclonal antibodies to keratin 8 and 17. Acta Cytol.
41:307–312. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Merkin RD, Vanner EA, Romeiser JL, Shroyer
ALW, Escobar-Hoyos LF, Li J, Powers RS, Burke S and Shroyer KR:
Keratin 17 is overexpressed and predicts poor survival in estrogen
receptor-negative/human epidermal growth factor receptor-2-negative
breast cancer. Hum Pathol. 62:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ide M, Kato T, Ogata K, Mochiki E, Kuwano
H and Oyama T: Keratin 17 expression correlates with tumor
progression and poor prognosis in gastric adenocarcinoma. Ann Surg
Oncol. 19:3506–3514. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Khanom R, Nguyen CT, Kayamori K, Zhao X,
Morita K, Miki Y, Katsube K, Yamaguchi A and Sakamoto K: Keratin 17
is induced in oral cancer and facilitates tumor growth. PLoS One.
11:e01611632016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mikami T, Maruyama S, Abe T, Kobayashi T,
Yamazaki M, Funayama A, Shingaki S, Kobayashi T, Jun C and Saku T:
Keratin 17 is co-expressed with 14-3-3 sigma in oral carcinoma in
situ and squamous cell carcinoma and modulates cell proliferation
and size but not cell migration. Virchows Arch. 466:559–569. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Regenbogen E, Mo M, Romeiser J, Shroyer
ALW, Escobar-Hoyos LF, Burke S and Shroyer KR: Elevated expression
of keratin 17 in oropharyngeal squamous cell carcinoma is
associated with decreased survival. Head Neck. 40:1788–1798.
2018.PubMed/NCBI
|
|
29
|
Liu J, Liu L, Cao L and Wen Q: Keratin 17
promotes lung adenocarcinoma progression by enhancing cell
proliferation and invasion. Med Sci Monit. 24:4782–4790. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Takenami T, Maeda S, Karasawa H, Suzuki T,
Furukawa T, Morikawa T, Takadate T, Hayashi H, Nakagawa K, Motoi F,
et al: Novel biomarkers distinguishing pancreatic head Cancer from
distal cholangiocarcinoma based on proteomic analysis. BMC Cancer.
19:3182019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Jong EM, van Vlijmen IM, van Erp PE,
Ramaekers FC, Troyanovski SM and van de Kerkhof PC: Keratin 17: A
useful marker in anti-psoriatic therapies. Arch Dermatol Res.
283:480–482. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Babu S, Mockler DC, Roa-Pena L,
Szygalowicz A, Kim NW, Jahanfard S, Gholami SS, Moffitt R,
Fitzgerald JP, Escobar-Hoyos LF and Shroyer KR: Keratin 17 is a
sensitive and specific biomarker of urothelial neoplasia. Mod
Pathol. 32:717–724. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Karia PS, Morgan FC, Califano JA and
Schmults CD: Comparison of tumor classifications for cutaneous
squamous cell carcinoma of the head and neck in the 7th vs. 8th
edition of the AJCC cancer staging manual. JAMA Dermatol.
154:175–181. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45((W1)):
W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thul PJ and Lindskog C: The human protein
atlas: A spatial map of the human proteome. Protein Sci.
27:233–244. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lindskog C: The human protein atlas-an
important resource for basic and clinical research. Expert Rev
Proteomics. 13:627–629. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ponten F, Jirstrom K and Uhlen M: The
human protein atlas-a tool for pathology. J Pathol. 216:387–393.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sun Q, Zhang Y, Su J, Li T and Jiang Y:
Role of hydroxysteroid dehydrogenase-like 2 (HSDL2) in human
ovarian cancer. Med Sci Monit. 24:3997–4008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Troyanovsky SM, Leube RE and Franke WW:
Characterization of the human gene encoding cytokeratin 17 and its
expression pattern. Eur J Cell Biol. 59:127–137. 1992.PubMed/NCBI
|
|
41
|
Depianto D, Kerns ML, Dlugosz AA and
Coulombe PA: Keratin 17 promotes epithelial proliferation and tumor
growth by polarizing the immune response in skin. Nat Genet.
42:910–914. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kolokythas A, Schwartz JL, Pytynia KB,
Panda S, Yao M, Homann B, Sroussi HY, Epstein JB, Gordon SC and
Adami GR: Analysis of RNA from brush cytology detects changes in
B2M, CYP1B1 and KRT17 levels with OSCC in tobacco users. Oral
Oncol. 47:532–536. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Escobar-Hoyos LF, Shah R, Roa-Pena L,
Vanner EA, Najafian N, Banach A, Nielsen E, Al-Khalil R, Akalin A,
Talmage D and Shroyer KR: Keratin-17 promotes p27KIP1 nuclear
export and degradation and offers potential prognostic utility.
Cancer Res. 75:3650–3662. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hobbs RP, Batazzi AS, Han MC and Coulombe
PA: Loss of keratin 17 induces tissue-specific cytokine
polarization and cellular differentiation in HPV16-driven cervical
tumorigenesis in vivo. Oncogene. 35:5653–5662. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Roa-Pena L, Leiton CV, Babu S, Pan CH,
Vanner EA, Akalin A, Bandovic J, Moffitt RA, Shroyer KR and
Escobar-Hoyos LF: Keratin 17 identifies the most lethal molecular
subtype of pancreatic cancer. Sci Rep. 9:112392019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sritananuwat P, Sueangoen N, Thummarati P,
Islam K and Suthiphongchai T: Blocking ERK1/2 signaling impairs
TGF-β1 tumor promoting function but enhances its tumor suppressing
role in intrahepatic cholangiocarcinoma cells. Cancer Cell Int.
17:852017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cook SJ, Stuart K, Gilley R and Sale MJ:
Control of cell death and mitochondrial fission by ERK1/2 MAP
kinase signalling. FEBS J. 284:4177–4195. 2017. View Article : Google Scholar : PubMed/NCBI
|