Introduction
Cervical cancer is the fourth most prevalent type of
cancer in females globally and a significant cause of mortality in
developing countries (1). Cervical
cancer may develop due to persistent high-risk human papillomavirus
(HPV) infection (2). Progress has
been made in understanding the HPV genome replication during the
viral life cycle; however, accurate surveillance strategies and
targeted therapies are still required to eradicate this disease
(3).
Tumor necrosis factor-related apoptosis inducing
ligand (TRAIL) is a powerful cancer cell apoptosis-inducing factor
that can induce apoptosis in a p53-independent manner (4). However, TRAIL resistance in cancer
cells is a significant barrier to improve TRAIL-based clinical
therapies (5). Therefore, agents are
required to either improve the impact of TRAIL or overcome
resistance to it (6,7). Several studies have demonstrated that
the cervical cancer HeLa cell line is resistant to TRAIL-induced
apoptosis, and the mechanism of resistance is not fully understood
(8,9). Various mechanisms have been proposed
for TRAIL resistance (10,11). Upregulation of CASP8- and FADD-like
apoptosis regulator (cFLIP) and downregulation of death receptors
(DR4 and DR5) and the Bcl-2 family (Bcl-2, Bcl-xL and MCL1
apoptosis regulator, BCL2 family member) are the most commonly
proposed TRAIL resistance mechanisms (12).
Survivin is an anti-apoptotic molecule that is
generally overexpressed in malignant cells (13). Survivin is a member of the inhibitor
of apoptosis gene family, which regulates apoptosis and the cell
cycle (13,14). Survivin has been reported to interact
with and inhibit caspases to decrease apoptosis (15). Survivin is expressed in various types
of cancer, including breast cancer (16), cervical cancer (17), non-small cell lung cancer (18) and osteosarcoma (19). Its upregulation is associated with
survival and resistance to TRAIL-based therapy (20,21).
Small molecule inhibitor YM155, a survivin
suppressor, inhibits cell proliferation and mediates apoptosis in
breast (22), gastric (23), liver (24), and pancreatic (25) cancer models. Other than suppressing
the levels of survivin in various types of cancer, YM155 has also
been reported to have strong apoptogenic properties (26,27).
Additionally, it has been reported that YM155 has an anticancer
effect on breast (22), esophageal
(28), colon (29) and bone marrow (30) cancer and sensitizes different types
of cancer cells to TRAIL-mediated apoptosis (31). However, a growing body of evidence
has reported that survivin has a strong inhibitory effect on TRAIL
therapy (32).
The present study investigated the impact of a low
dosage of YM155 on TRAIL-mediated apoptosis and molecular
mechanisms underlying TRAIL sensitization in human cervical cancer
cells.
Materials and methods
Cell culture
HeLa cells were procured from the Korean Cell Line
Bank; Korean Cell Line Research Foundation. Cells were maintained
in DMEM (PAN-Biotech GmbH) with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.). A total of 100 µg/ml penicillin-streptomycin
(HyClone; Cytiva) was added to make complete media. Cells were
maintained in a CO2 cell culture incubator (Thermo
Fisher Scientific, Inc.) at 37°C with 5% CO2.
Reagents and antibodies
YM155 and TRAIL were purchased from Selleck
Chemicals and PeproTech, Inc., respectively. Western blot and
immunofluorescence analyses were performed using antibodies against
cleaved-poly (ADP-ribose) polymerase (PARP; dilution, 1:1,000; cat.
no. 5625S) from Cell Signaling Technology, Inc., cFLIP (dilution,
1:1,000; cat. no. 10394-1-AP) and DR5 (dilution, 1:1,000; cat. no.
15497-1-AP) from ProteinTech Group, Inc., γH2A histone family
member X (γH2AX; dilution, 1:1,000 for western blot and 1:100 for
immunofluorescence; cat. no. 05-636) from EMD Millipore, cleaved
caspase-3 (dilution, 1:100 for immunofluorescence; cat. no. 9661S)
from Cell Signaling Technology, Inc., GAPDH (dilution, 1:2,000;
cat. no. sc-32233) and β-tubulin (1:1,000; cat. no. sc-5274) from
Santa Cruz Biotechnology, Inc., and HRP-conjugated secondary
anti-mouse (1:10,000; cat. no. 31430) and anti-rabbit (1:10,000;
cat. no. 31460) antibodies from Thermo Fisher Scientific, Inc., and
fluorescence-conjugated secondary antibodies goat anti-mouse IgG
(H+L) Alexa Fluor 488 (1:250; cat. no. A32723) and goat anti-rabbit
IgG (H+L) Alexa Fluor 488 (1:250; cat. no. A27034) from Invitrogen,
Thermo Fisher Scientific, Inc.
Western blotting
HeLa cells were treated with 25 ng/ml TRAIL, 25 nM
YM155 or a combination of both for 24 h and subjected to western
blot analysis. Cells were collected after 24-h incubation at 37°C
and 5% CO2 and lysed with a protein extraction buffer
(20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM Na2EDTA, 1 mM
EGTA, 1% NP-40, 1% sodium deoxycholate, 2.5 mM sodium
pyrophosphate, 1 mM b-glycerophosphate, 1 mM
Na3VO4, 1 µg/ml leupeptin) with a PMSF
protease inhibitor (Cell Signaling Technology, Inc.). The protein
concentration was determined by Bradford assay (Bio-Rad
Laboratories, Inc.). Then, 50 µg protein was loaded on 8 or 12%
SDS-PAGE and transferred to PVDF membrane (EMD Millipore) activated
by methanol. Following transfer, non-specific binding was blocked
with 5% skim milk and incubated at room temperature for 1 h. The
membranes were probed with primary antibodies and incubated at 4°C
for overnight. Subsequently, the membranes were incubated with a
HRP-conjugated secondary antibody for 1 h at room temperature and
washed thrice for 5 min with TBS-Tween 20 (0.05%) to remove unbound
probes. The membranes were subjected to chemiluminescence-based
detection (Pierce ECL Plus; Thermo Fisher Scientific Inc.). Imaging
was performed using a ChemiDoc system with Image Lab software
version 5.2 (Bio-Rad Laboratories, Inc.).
Flow cytometry
Apoptosis was assessed using propidium iodide (PI)
staining kit (BD Biosciences) according to the manufacturer's
protocol. Briefly, HeLa cells were seeded at a density of
2.5×105 per well and treated with 25 ng/ml TRAIL and 25
nM YM155 for 24 h. Subsequently, 5×104 cells were
collected and washed with ice-cold PBS twice in the presence of 10%
FBS, and 70% ethanol was added into the cell suspensions. A total
of 2 mg/ml RNaseA was added and incubated for 15 min at 4°C.
Finally, cells were stained with 10 µl PI (50 mg/ml) at room
temperature for 10 min. DNA content was measured within 1 h using
flow cytometry (FACS CANTO II; BD Biosciences). Data were analyzed
using FACSDiva software (version 8; BD Biosciences).
Immunofluorescence
Immunofluorescence was used to assess nuclear
morphology and DNA damage by γH2AX foci formation and cleaved
caspase-3 activation assays. A total of 2×104 HeLa cells
were seeded into a 4-well dish containing cover slips and incubated
overnight. Subsequently, HeLa cells were treated with 25 ng/ml
TRAIL and/or 25 nM YM155 and incubated for 24 h. The reaction was
immediately stopped and cells were fixed with 4% paraformaldehyde
at room temperature (Wako Chemicals USA, Inc.) for 20 min. Fixed
cells were washed using 0.03% Triton-X 100 (Sigma-Aldrich; Merck
KGaA) three times. To eliminate non-specific binding, 5% BSA
(Bovogen Biologicals Pty Ltd.) in 1X PBS was used as a blocking
reagent. After 1 h of blocking at room temperature, primary
antibody was added, followed by incubation at 4°C overnight. The
next day, cells were washed using PBS and incubated with relevant
Alexa Fluor 48-8 and 594-conjugated secondary antibodies at room
temperature for 1 h. DAPI (1 µg/ml; Invitrogen; Thermo Fisher
Scientific, Inc.) was added to the cells and incubated at room
temperature for 15 min to stain the nuclei, and cells were mounted
on glass slides. Cells were imaged at ×40 magnification using a
confocal microscope (Leica TCS SP5; Leica Microsystems GmbH).
Reverse transcription-quantitative
(RT-q)PCR
HeLa cells were treated with 25 ng/ml TRAIL and/or
25 nM YM155 for 24 h and harvested for RNA extraction.
TRIzol® reagent (Favorgen Biotech Corporation) was used
to extract the RNA. A total of 40 ml RNase-free water was added to
the extracted vial to reconstitute the RNA. Total RNA concentration
was quantified using a Nanodrop (Thermo Fisher Scientific, Inc.).
The SuperScript III First-Strand Synthesis System (Thermo Fisher
Scientific, Inc.) was used to synthesize the cDNA according to the
manufacturer's protocol. Briefly, 1 µg of total RNA was reverse
transcribed using the SuperScript III kit with 25 pmol oligo-dT
primer and 50 pmol random hexamer. Then, the cDNA was diluted at
1:10 with dilution buffer provided by the manufacturer. QPCR was
performed using a real-time PCR system (Thermo Fisher Scientific,
Inc.) and Fast SYBR® Green Master Mix (Thermo Fisher
Scientific, Inc.) with the following primers: DR5 forward,
5′-CCCAACAAGACCTAGCTCCC-3′ and reverse, 5′-GACCTCCTTTTCTGCTTGCG-3′;
cFLIP forward, 5′-GCCGAGGCAAGATAAGCAAG-3′ and reverse,
5′-AGTCTGTTCAAGGAGCAGGG-3′; survivin forward,
5′-AGAACTGGCCCTTCTTGGAGG-3′ and reverse,
5′-CTTTTTATGTTCCTCTATGGGGTC-3′; and GAPDH forward,
5′-CTGACTTCAACAGCGACACC-3′ and reverse,
5′-TAGCCAAATTCGTTGTCATACC-3′. The relative quantification of gene
expression was determined using the 2−ΔΔCq method
(33). The thermocycling conditions
were as follows: Pre-incubation at 95°C for 3 min, followed by 45
cycles of 95°C for 15 sec and 60°C for 60 sec. The final
amplification cycle was followed by a melt curve analysis for the
specificity of the RT-qPCR.
Cell viability assay
The cytotoxicity of TRAIL and YM155 was assessed
using a Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies,
Inc.). Following drug treatment, 10 µl CCK-8 reagent was added, and
the mixture was incubated at 37°C for 2 h according to the
manufacturer's instructions. Subsequently, absorbance was measured
at 540 nm using a microtiter plate reader (Bio-Rad Laboratories,
Inc.).
Matrigel invasion assay
HeLa cells were seeded, and cell invasion was
evaluated using 0.8-µm Transwell chambers coated with Matrigel at
37°C for 1 h (Corning Inc.) according to the manufacturer's
protocol. In brief, HeLa cells were suspended at a density of
2.5×104 cells in 500 µl serum-free DMEM and placed in
the upper chamber. Subsequently, 700 µl complete DMEM was added to
the lower chamber, followed by incubation at 37°C for 24 h. Cells
on the surface of the insert were removed, and the lower surface
was fixed with ice-cold methanol at room temperature for 15 min.
After fixing, cells were stained with crystal violet (0.1%, diluted
in methanol) for 5 min at room temperature. Cells were counted
using bright field microscopy and compared among the DMSO, YM155,
TRAIL and combination groups. The numbers of invaded cells were
represented graphically.
In vitro scratch assay
An in vitro scratch assay was performed to
assess the migratory activity of HeLa cells treated with 25 ng/ml
TRAIL and/or 25 nM YM155. After 24-h treatment, the cells were
harvested and re-seeded at 90% confluency and incubated at 37°C
overnight. Using a sterile pipette tip, scratches were made in the
monolayers of HeLa cells. The scratched monolayer was washed with
PBS and incubated at 37°C with serum-free 1X DMEM. The wounded
region was captured under a light microscope (IX71; Olympus
Corporation) at 0, 24, 48 and 72 h, and ImageJ 1.51j8 software
(National Institutes of Health) was used to calculate the
percentage of wound closure as previously described (34). Data were obtained from three
independent experiments.
Soft agar assay
DMEM (1X) complete medium and 1% agarose were mixed
at an equal ratio of 1:1 (v/v) and plated onto 35-mm dishes. The
next day, cells were seeded at a density of 1×104 per
well and suspended in 0.7% agarose/1X DMEM mixture. Subsequently,
1% DMSO, 5 nM YM155, 5 ng/ml TRAIL and YM155/TRAIL combination were
added to DMEM. Medium containing the aforementioned drug
concentrations was added every other day for 14 days. Crystal
violet (0.01% diluted in 20% methanol) was added at room
temperature for 5 min to stain anchorage-independent colonies.
Finally, cells were observed under a light microscope at ×4
magnification.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8 (GraphPad Software, Inc.) and data are presented as the
mean ± standard deviation of three independent experiments. Data
were analyzed by one-way ANOVA. Multiple comparisons among the
groups were performed by Tukey's post hoc test. For in vitro
scratch assay, two-way ANOVA followed by Tukey's post hoc test was
performed. P<0.05 was considered to indicate a statistically
significant difference.
Results
YM155 sensitizes HeLa cells to
TRAIL-mediated apoptosis
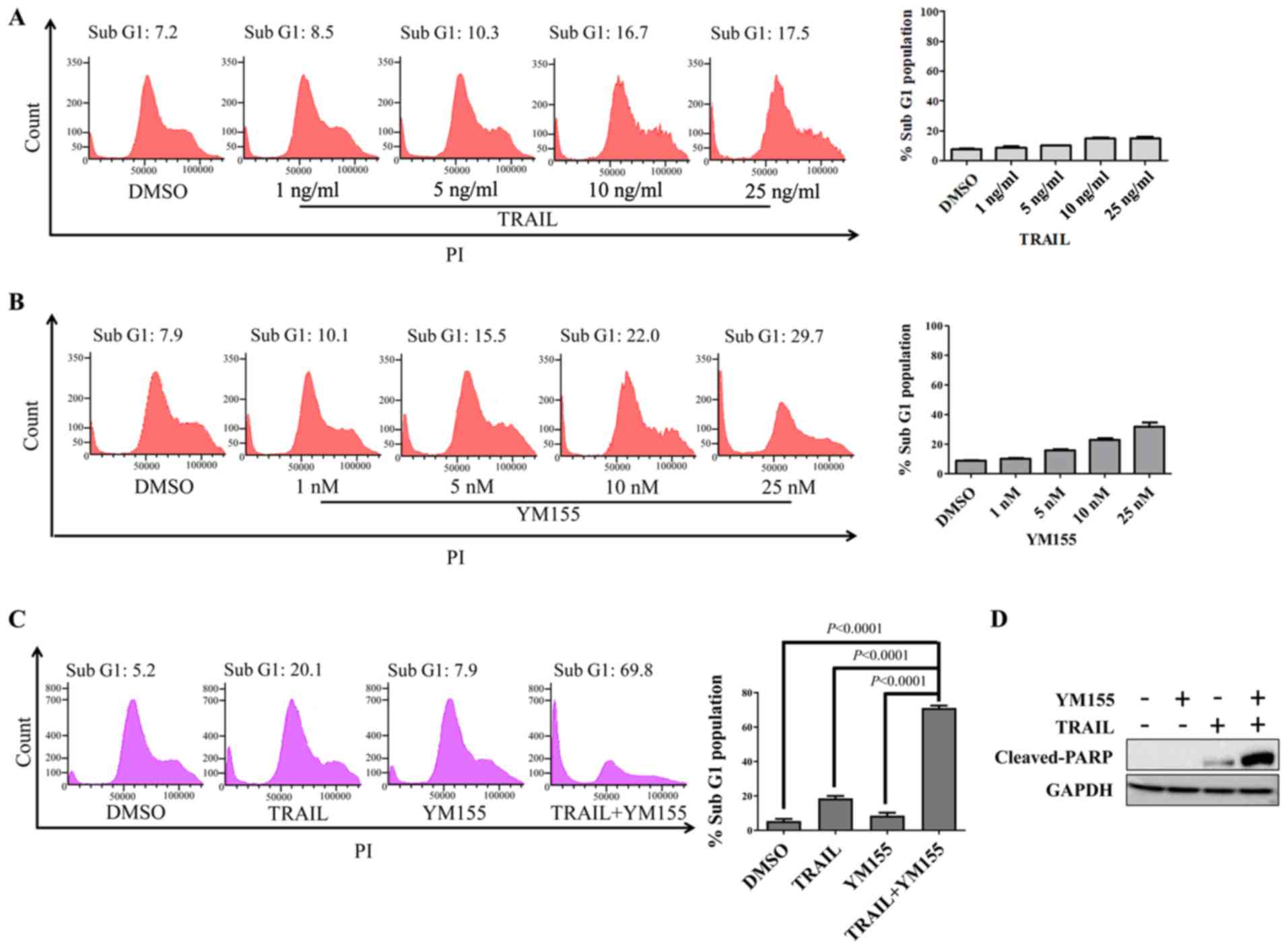
First, HeLa cells were treated with 1% DMSO, 25
ng/ml TRAIL or 25 nM YM155 for 24 h. PI staining was used to
analyze apoptotic cells by flow cytometry (Fig. 1A and B). TRAIL or YM155 alone did not
induce cell death in HeLa cells (Fig. 1A
and B), whereas the combination of TRAIL and YM155
significantly increased the sub-G1 population compared
with the DMSO control group in HeLa cells (Fig. 1C). To further validate the
combinatorial effect of TRAIL-YM155, western blotting was performed
to assess cleaved-PARP levels. Consistent with the sub-G1
population experimental results, combination treatment induced
greater cleaved-PARP expression than individual treatment (Fig. 1D). Overall, YM155 enriched
TRAIL-mediated apoptosis via an extrinsic signaling pathway in
human cervical cancer cells.
Combination of TRAIL and YM155 induces
DNA damage toxicity in HeLa cells
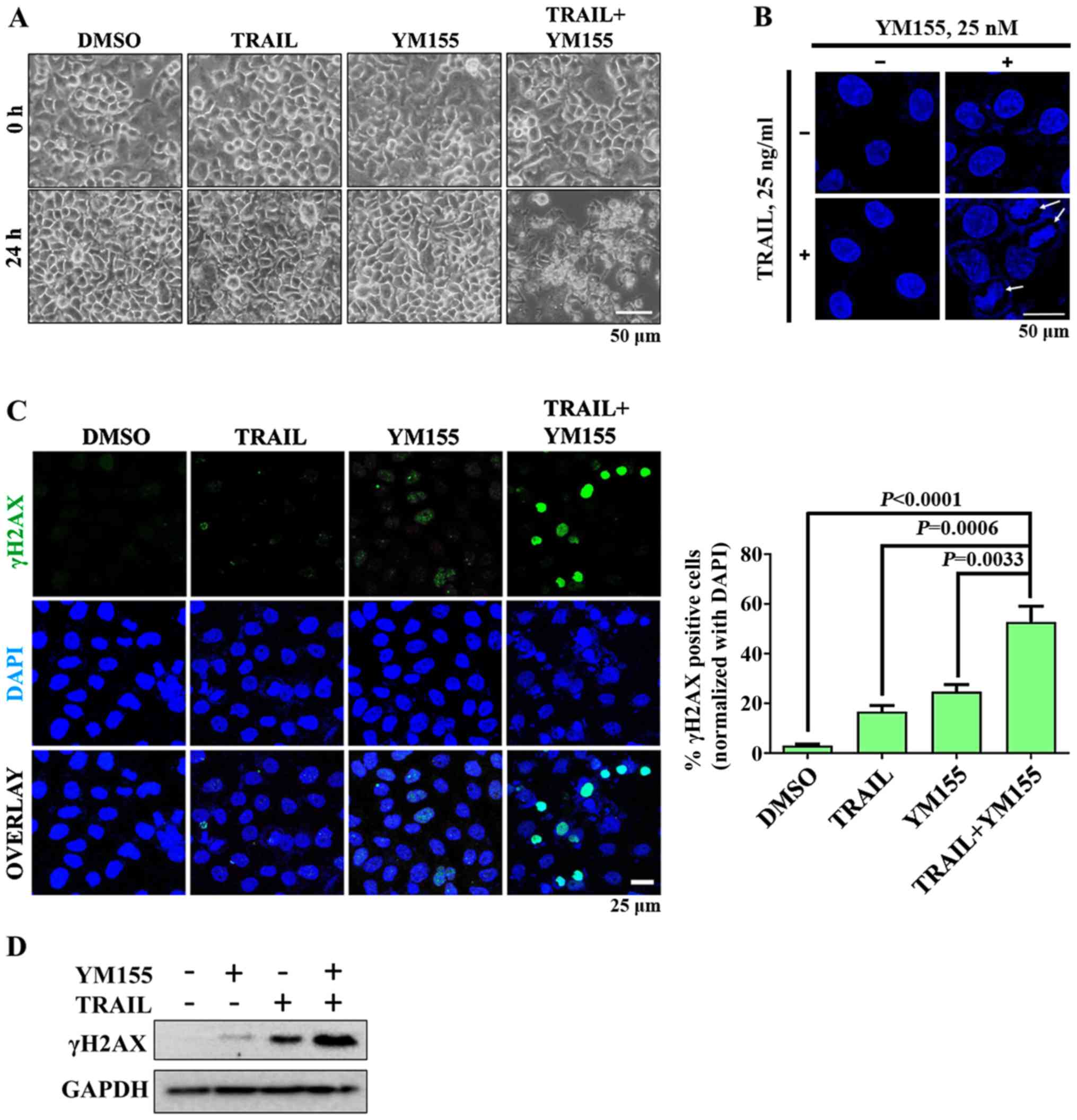
In order to investigate whether combination
treatment of TRAIL and YM155 lead to DNA damage, cells were treated
with 25 ng/ml TRAIL or 25 nM YM155 alone and in combination.
Bright-field microscopy images revealed that cells in the
combination treatment group exhibited apoptotic morphology, whereas
cells in the single agent treatment groups did not (Fig. 2A). Subsequently, the shape of the
nucleus of HeLa cells was assessed by staining with DAPI. As
presented in Fig. 2B, chromatin
condensation was observed following combination treatment but not
when cells were treated with TRAIL or YM155 alone. To further
confirm that combination treatment induces DNA damage in HeLa
cells, γH2AX foci formation was assessed using immunofluorescence.
As presented in Fig. 2C, robust H2AX
phosphorylation was observed in HeLa cells treated with the
combination of TRAIL and YM155 compared with in the single agent
treatment and DMSO control groups. Consistent with the
aforementioned results, combination treatment of TRAIL and YM155
resulted in higher protein expression levels of γH2AX than
individual treatment (Fig. 2D).
Overall, the combination of TRAIL and YM155 induced DNA damage in
HeLa cells but individual treatment did not.
YM155 increases TRAIL-induced
apoptosis via downregulation of cFLIP and survivin levels
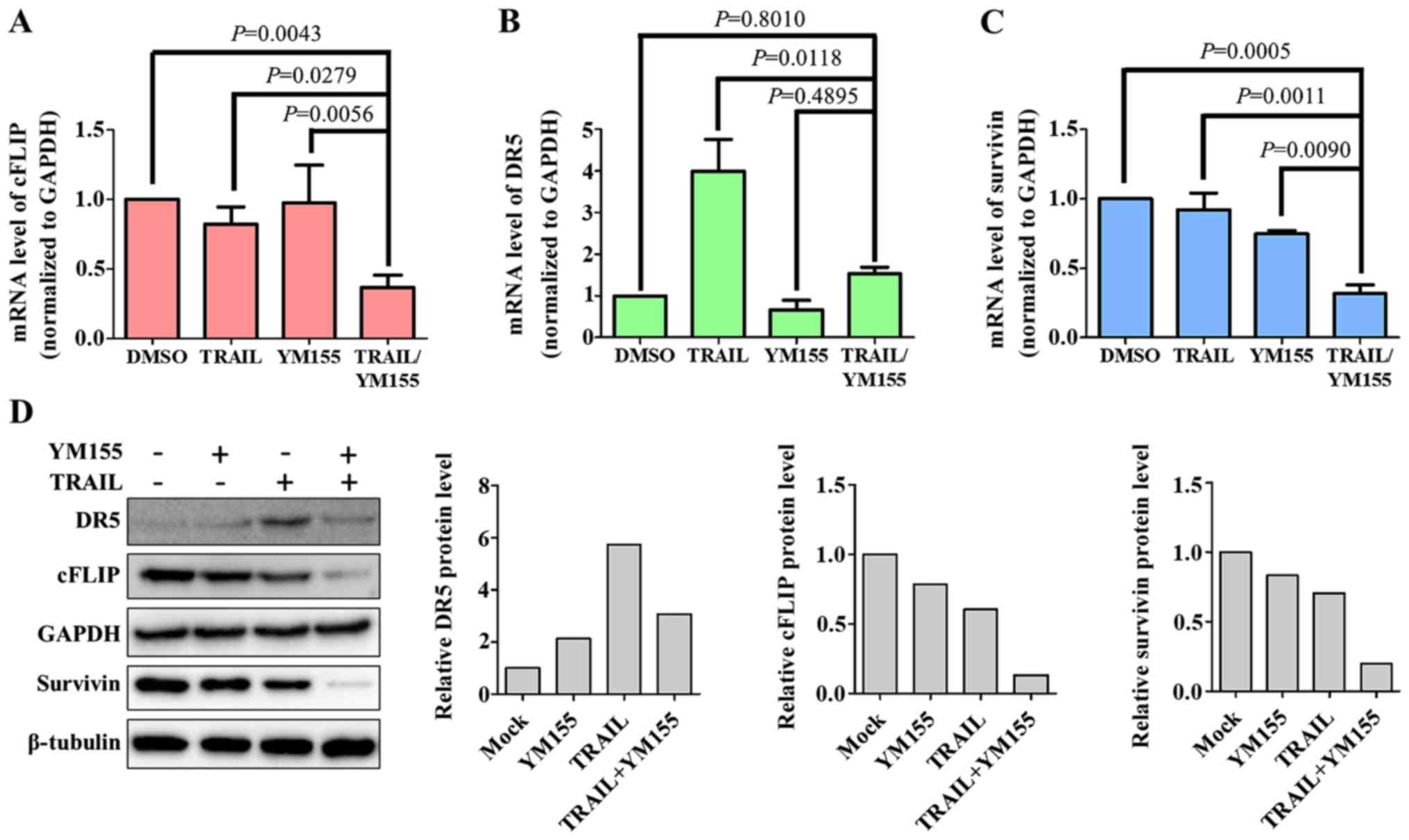
In order to determine the mechanisms underlying
YM155-mediated TRAIL sensitivity in HeLa cells,
apoptosis-associated proteins, including DR5 (pro-apoptotic), cFLIP
(anti-apoptotic) and survivin (anti-apoptotic) were examined in the
present study. HeLa cells were treated with 1% DMSO, 25 ng/ml TRAIL
or 25 nM YM155 alone and in combination for 24 h and then subjected
to RT-qPCR analysis. As presented in Fig. 3, HeLa cells treated with both TRAIL
and YM155 exhibited downregulation of cFLIP and survivin at the
mRNA and protein levels (Fig. 3A, C and
D). DR5, a pro-apoptotic protein involved in TRAIL sensitivity
(35), exhibited no changes
following combination treatment (Fig. 3B
and D). Notably, treatment with TRAIL alone induced a
significant increase in the mRNA levels of DR5 (Fig. 3B). Similarly, the protein level of
DR5 was also upregulated by TRAIL treatment (Fig. 3D). This suggests that DR5
upregulation was specific to TRAIL treatment alone and not in
combination. The aforementioned data indicate that downregulation
of cFLIP and survivin serves a major role in combination treatment
of YM155 and TRAIL-mediated apoptosis.
Combination treatment decreases cancer
progression in vitro
The effect of TRAIL and YM155 combination treatment
in HeLa cells was next assessed using a cell viability assay. As
presented in Fig. 4A, combination
treatment of TRAIL and YM155 significantly decreased the viability
of HeLa cells (2.08-fold) compared with the DMSO control. To test
the hypothesis that the combination of TRAIL and YM155 decreases
cancer progression, the effect of the combination treatment on HeLa
cell migration and invasion was assessed using an in vitro
scratch assay and Matrigel-coated chamber invasion assay,
respectively. HeLa cells treated with both TRAIL and YM155
exhibited a significant decrease in invasive capability compared
with the DMSO control and single agent treatment groups (Fig. 4B). Consistently, combination
treatment also resulted in a delayed wound closure rate compared
with the DMSO control and single agent treatment groups (Fig. 4C). It has been reported that
caspase-3 is a cysteine-aspartic acid protease that will be cleaved
by TRAIL and executes apoptosis in cancer cells (36,37).
Therefore, the present study monitored the levels of cleaved
caspase-3 in HeLa cells. To achieve this, HeLa cells were treated
with the 25 ng/ml TRAIL or 25 nM YM155 alone or in combination for
24 h. Subsequently, cleaved caspase-3 levels in samples were
evaluated by immunofluorescence. It was observed that the
combination treatment of TRAIL and YM155 significantly increased
the cleaved caspase-3 levels compared with DMSO and TRAIL or YM155
alone treated samples (Fig. 4D).
Subsequently, cell proliferation was assessed using an
anchorage-independent colony formation assay. HeLa cells were
selected with the combination of 5 ng/ml of TRAIL and 5 nM of YM155
or individual treatment, and colony formation was analyzed. The
number of colonies that appeared in cells treated with both TRAIL
and YM155 was significantly lower compared with the DMSO control
and single agent treatment groups (Fig.
4E). Overall, the results of the present study suggest that
combination treatment of TRAIL and YM155 decreased the viability,
invasion and migration of HeLa cells. The synergistic behavior of
TRAIL/YM155 combination further induced DNA damage and apoptosis
and may be an effective strategy to treat human cervical
cancer.
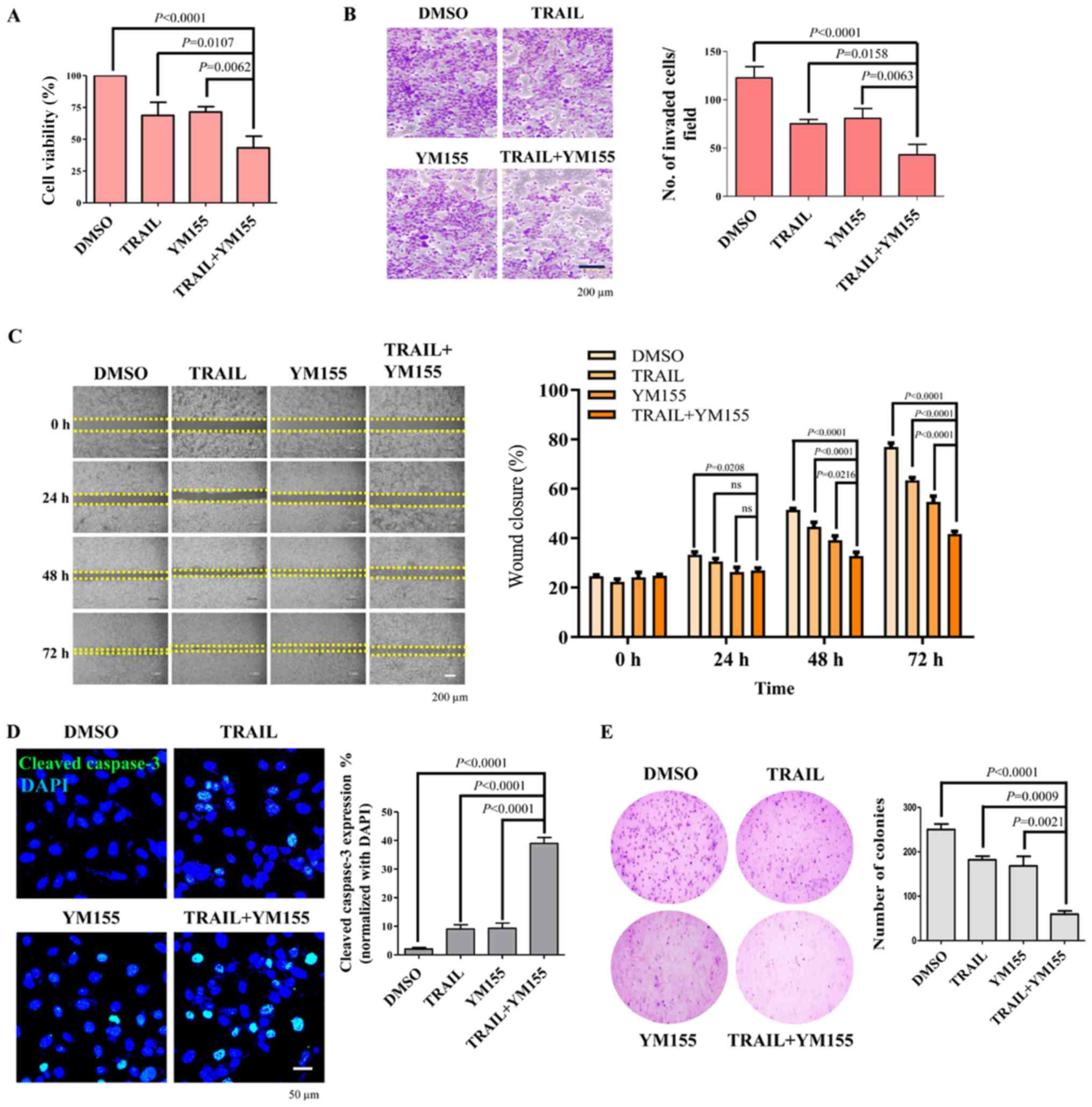 | Figure 4.Effect of combination treatment on
cell viability, cellular invasion and migration of HeLa cells. (A)
HeLa cells were treated with 25 ng/ml TRAIL and 25 nM YM155 for 24
h. Cell Counting Kit-8 reagent was added to the treated cells to
assess cell viability. (B) Matrigel cell invasion assay in HeLa
cells. Cells were treated with 25 ng/ml TRAIL and 25 nM YM155 for
24 h. Cells were seeded in the upper chamber of the plate. After 24
h of incubation, cells on the bottom of the chamber membrane were
fixed, stained and invasive cells were counted for graphical
representation. Scale bar, 200 µm. (C) HeLa cells were treated with
25 ng/ml TRAIL and 25 nM YM155 for 24 h and assessed for migratory
potential using a wound healing assay. Images were captured at 0,
24, 48 and 72 h. Scale bar, 200 µm. Right panel indicates the
percentage of wound closure. Data are presented as the mean and
standard deviation of three independent experiments. Two-way ANOVA
was used and P-values are as indicated. (D) HeLa cells were treated
with either TRAIL or YM155 or the combination at 25 ng/ml or 25 nM,
respectively, for 24 h. An immunofluorescence assay was performed
to monitor the activation of cleaved caspase-3 (green). Nuclei were
stained with DAPI (blue). Scale bar, 50 µm. Right panel is the
quantified data presented as the percentage of cleaved caspase-3
expression. (E) HeLa cells were selected with 5 ng/ml of TRAIL
and/or 5 nM of YM155. After 14 days of selection with 5 ng/ml TRAIL
and 5 nM YM155, colonies were stained with 0.01% crystal violet,
and images were captured at ×4 magnification. The assay was
performed in triplicate. The right panel is the quantified data of
the soft agar assay presented as colony numbers. Data are presented
as the mean and standard deviation of three independent
experiments. One-way ANOVA followed by Tukey's post hoc test was
used, and P-values are as indicated. TRAIL, tumor necrosis
factor-related apoptosis inducing ligand. |
Discussion
Despite the use of aggressive and toxic treatments,
patients with advanced stage cervical cancer have an unacceptably
high mortality rate; an estimated 13,800 new cervical cancer will
be diagnosed and 4,290 deaths will occur in the United States in
2020 (38). Epidemiological,
clinical and cellular studies have reported that persistent HPV
infections are involved in the development of cervical cancer
(38), although cervical cancer can
also develop without HPV infection. The poor survival percentage
demands research into more effective novel treatment approaches. In
the present study, the sensitivity of TRAIL improved with the
addition of YM155 in the treatment of human cervical cancer cells.
The therapeutic potential of TRAIL and YM155 combination treatment
was analyzed in HeLa cells. Previous research has demonstrated that
HeLa cells are less sensitive to TRAIL-induced apoptosis (39,40). To
sensitize the HeLa cells to TRAIL-mediated apoptosis, YM155 was
used as a combinatorial drug partner. YM155 has been reported as a
survivin inhibitor developed for cancer treatment, and its effects
have been demonstrated in multiple preclinical studies and phase
I/II clinical trials (41–44).
Several reports have suggested that YM155 sensitizes
cells to TRAIL-induced apoptosis in human renal carcinoma Caki
cells (31), triple negative breast
cancer (45) and a number of other
types of cancer. However, to the best of our knowledge, there are
currently no data on the combination of TRAIL and YM155 in human
cervical cancer. Therefore, the effect of YM155 on the induction of
TRAIL-mediated apoptosis in HeLa cells was assessed in the present
study. It was revealed that HeLa cells were sensitized by the
addition of a low concentration of YM155, but not in individually
treated groups. Previous studies have suggested that YM155 induces
DNA damage in several types of cancer (46,47).
Therefore, phosphorylation of H2AX, which causes double strand
breaks, was assessed by immunofluorescence and western blotting
assays. YM155-treated cells expressed γH2AX. Notably, treatment
with TRAIL alone did not induce DNA damage, whereas the combination
treatment of TRAIL and YM155 was associated with marked expression
of γH2AX in HeLa cells and eventually led to apoptosis. A previous
report has suggested that the combination of YM155 and TRAIL in
SiHa cells induced a 48% higher apoptotic rate compared with that
of the DMSO group (48), whereas the
present combinatorial approach demonstrated an ~71% increase in
apoptosis relative to that of the DMSO group by activating cleaved
caspase-3 in HeLa cells. The main difference is that the cells were
first treated with YM155 alone for 24 h and then TRAIL treatment
was applied for another 15 h in the previous study whereas the
present study induced apoptosis via synergistic action of YM155 and
TRAIL together in cervical cancer cells. This explains the
importance of the synergistic behavior of YM155/TRAIL in a cellular
environment to hinder the progress of cervical cancer growth. It is
important to note that the use of antibiotics in drug response
studies can alter gene expression (49). Both genetic and phenotypical
characterizations of YM155 and TRAIL have been studied extensively
(50–53). Throughout the experiments in the
present study, both the control and experimental groups received
equal amounts of antibiotics, and thus, their effects on the
results were negligible.
Previous data revealed that DR5 is the major
determinant of TRAIL-induced apoptosis (54,55), and
cFLIP and survivin confer resistance to TRAIL therapy for different
cancer types (56,57). Several reports have demonstrated that
DR5 serves a greater role in apoptosis compared with that of DR4
(58,59). DR5 activates the extrinsic apoptotic
signaling pathway by binding of the death ligand (TRAIL) (60,61).
However, the action of TRAIL-mediated apoptosis is mainly
associated with Fas-associated via death domain (FADD) recruitment
to the DR5/TRAIL complex. cFLIP inhibits the action of FADD and
prevents it from binding to the DR5/TRAIL complex (62,63).
High cFLIP expression in human cervical cancer confers resistance
to TRAIL therapy (64).
Additionally, it has been noted that DR4 serves an antagonistic
role to DR5 in TRAIL-mediated apoptosis (59). Furthermore, DR4 is responsible for
the sensitization of cancer cells during the combination treatment
involving irradiation (39).
Therefore, regulating cFLIP is more crucial than the other genes
involved in DR5/TRAIL-mediated apoptotic signaling pathway.
However, elucidating the role of DR4 in TRAIL-mediated apoptosis is
essential for future investigations. In our previous study, cFLIP
was successfully downregulated in HeLa cells using the CRISPR-Cas9
system and its effectiveness in TRAIL-mediated apoptosis was
examined (8). On the other hand,
consistent with our previous study (8), the mRNA and protein expression levels
of cFLIP were decreased when HeLa cells were treated with TRAIL and
YM155 together. However, this combinatorial approach did not alter
endogenous DR5 expression.
Both the mRNA and protein expression levels of
survivin were not significantly decreased when the low dose of
YM155 (25 nM) alone was applied. Similarly, a low concentration of
YM155 (20 nM) did not exhibit any inhibitory effect on survivin
expression in human renal cell carcinoma cell lines treated for 48
h; however, high concentrations of YM155 (>40 nM) decrease
survivin protein levels in renal cell carcinoma cells (27). It was hypothesized that a low
concentration of YM155 does not block the transcription of
baculoviral IAP repeat containing 5 (BIRC5), a gene that
encodes the survivin protein, and thus it has no inhibitory effect
on survivin in these cell lines (27). However, further research is required
at the transcriptional level to elucidate the interaction of the
low dosage of YM155 and the BIRC5 gene. In the present
study, the low dosage of YM155 sensitized HeLa cells to
TRAIL-mediated apoptosis and decreased the mRNA and protein levels
of survivin. Therefore, the combination therapy holds promise as a
method to overcome the problem of side effects due to high doses
and could be safe for use in patients. However, further
investigations are required to validate the combination treatment
in different human cervical cancer cell lines and its potential
outcome in mouse xenograft models.
As available treatments have limited efficacy in
human cervical cancer (64), the
identification of novel therapeutic approaches to enhance patient
outcomes is urgently required. The present study suggests
combination treatment using YM155 and TRAIL as a therapeutic
approach. Specifically, YM155 enhanced TRAIL-mediated apoptosis in
HeLa cells. The results demonstrated that YM155 sensitizes human
cervical cancer cells to TRAIL-induced cell death by activating the
intrinsic pathway of apoptosis, and YM155/TRAIL combination
downregulated anti-apoptotic proteins, such as cFLIP and survivin,
in HeLa cells.
Acknowledgements
The authors would like to thank Dr Bharathi Suresh
(Hanyang University, Seoul, Republic of Korea) for her assistance
with statistical analysis.
Funding
The present study was supported by the Korea Health
Technology R&D Project through the Korea Health Industry
Development Institute, funded by the Ministry of Health &
Welfare, Republic of Korea (grant no. HI18C2383) and the National
Research Foundation of Korea, which is funded by the Ministry of
Education (grant no. 2018M3A9H3022412).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SJO and SR were responsible for the study design.
APC and NP performed the main experiments related to apoptosis and
oncogenic based assays, and drafted the manuscript. NRK performed
cell viability assays and performed statistical analysis. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Burd EM: Human papillomavirus and cervical
cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Soto D, Song C and McLaughlin-Drubin ME:
Epigenetic alterations in human papillomavirus-associated cancers.
Viruses. 9:248–265. 2017. View
Article : Google Scholar
|
|
4
|
Willms A, Schittek H, Rahn S, Sosna J,
Mert U, Adam D and Trauzold A: Impact of p53 status on
TRAIL-mediated apoptotic and non-apoptotic signaling in cancer
cells. PLoS One. 14:e02148472019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang S and El-Deiry WS: TRAIL and
apoptosis induction by TNF-family death receptors. Oncogene.
22:8628–8633. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang L and Fang B: Mechanisms of
resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther.
12:228–2237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pitti RM, Marsters SA, Ruppert S, Donahue
CJ, Moore A and Ashkenazi A: Induction of apoptosis by Apo-2
ligand, a new member of the tumor necrosis factor cytokine family.
J Biol Chem. 271:12687–12690. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Poondla N, Chandrasekaran AP, Heese K, Kim
KS and Ramakrishna S: CRISPR-mediated upregulation of DR5 and
downregulation of cFLIP synergistically sensitize HeLa cells to
TRAIL-mediated apoptosis. Biochem Biophys Res Commun. 512:60–65.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seo OW, Kim JH, Lee KS, Lee KS, Kim JH,
Won MH, Ha KS, Kwon YG and Kim YM: Kurarinone promotes
TRAIL-induced apoptosis by inhibiting NF-κB-dependent cFLIP
expression in HeLa cells. Exp Mol Med. 44:653–664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang A, Wilson NS and Ashkenazi A:
Proapoptotic DR4 and DR5 signaling in cancer cells: Toward clinical
translation. Curr Opin Cell Biol. 22:837–844. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Russo M, Mupo A, Spagnuolo C and Russo GL:
Exploring death receptor pathways as selective targets in cancer
therapy. Biochem Pharmacol. 80:674–682. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Malhi H and Gores GJ: TRAIL resistance
results in cancer progression: A TRAIL to perdition? Oncogene.
25:7333–7335. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mobahat M, Narendran A and Riabowol K:
Survivin as a preferential target for cancer therapy. Int J Mol
Sci. 15:2494–2516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fukuda S and Pelus LM: Survivin, a cancer
target with an emerging role in normal adult tissues. Mol Cancer
Ther. 5:1087–1098. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shin S, Sung BJ, Cho YS, Kim HJ, Ha NC,
Hwang JI, Chung CW, Jung JK and Oh BH: An Anti-apoptotic protein
human survivin is a direct inhibitor of caspase-3 and −7.
Biochemistry. 40:1117–1123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lv YG, Yu F, Yao Q, Chen JH and Wang L:
The role of survivin in diagnosis, prognosis and treatment of
breast cancer. J Thorac Dis. 2:100–110. 2010.PubMed/NCBI
|
|
17
|
Cao XQ, Lu HS, Zhang L, Chen LL and Gan
MF: MEKK3 and survivin expression in cervical cancer: Association
with clinicopathological factors and prognosis. Asian Pacific J
Cancer Prev. 15:5271–5276. 2014. View Article : Google Scholar
|
|
18
|
Falleni M, Pellegrini C, Marchetti A,
Oprandi B, Buttitta F, Barassi F, Santambrogio L, Coggi G and
Bosari S: Survivin gene expression in early-stage non-small cell
lung cancer. J Pathol. 200:620–626. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Trieb K, Lehner R, Stulnig T, Sulzbacher I
and Shroyer KR: Survivin expression in human osteosarcoma is a
marker for survival. Eur J Surg Oncol. 29:379–82. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park SH, Park SJ, Kim JO, Shin J-H, Kim
ES, Jo YK, Kim JS, Park SJ, Jin DH, Hwang JJ, et al:
Down-regulation of survivin by nemadipine-a sensitizes cancer cells
to TRAIL-induced apoptosis. Biomol Ther (Seoul). 21:29–34. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hwang JS, Lee HC, Oh SC, Lee DH and Kwon
KH: Shogaol overcomes TRAIL resistance in colon cancer cells via
inhibiting of survivin. Tumor Biol. 36:8819–8829. 2015. View Article : Google Scholar
|
|
22
|
Véquaud E, Séveno C, Loussouarn D,
Engelhart L, Campone M, Juin P and Barillé-Nion S: YM155 potently
triggers cell death in breast cancer cells through an
autophagy-NF-kB network. Oncotarget. 6:13476–13486. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng XJ, Lin JC, Ding YF, Zhu L, Ye J and
Tu SP: Survivin inhibitor YM155 suppresses gastric cancer xenograft
growth in mice without affecting normal tissues. Oncotarget.
7:7096–7109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang C, Cao x, Gei Y, Wang Y, Liu G,
Cheng G and Liu Q: Silencing of survivin by YM155 induces apoptosis
and growth arrest in hepatocellular carcinoma cells. Oncol Lett.
10:1627–1631. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao X, Puszyk WM, Lu Z, Ostrov DA, George
TJ, Robertson KD and Liu C: Small molecule inhibitor YM155-mediated
activation of death receptor 5 is crucial for chemotherapy-induced
apoptosis in pancreatic carcinoma. Mol Cancer Ther. 14:80–89. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jane EP, Premkumar DR, Sutera PA, Cavaleri
JM and Pollack IF: Survivin inhibitor YM155 induces mitochondrial
dysfunction, autophagy, DNA damage and apoptosis in Bcl-xL silenced
glioma cell lines. Mol Carcinog. 56:1251–1265. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sim MY, Huynh H, Go ML and Yuen JSP:
Action of YM155 on clear cell renal cell carcinoma does not depend
on survivin expression levels. PLoS One. 12:e01781682017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao N, Mao Y, Han G, Ju Q, Zhou L, Liu F,
Xu Y and Zhao X: YM155, a survivin suppressant, triggers
PARP-dependent cell death (parthanatos) and inhibits esophageal
squamous-cell carcinoma xenografts in mice. Oncotarget.
6:18445–18459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li WL, Lee MR and Cho MY: The small
molecule survivin inhibitor YM155 may be an effective treatment
modality for colon cancer through increasing apoptosis. Biochem
Biophys Res Commun. 471:309–314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng W, Yoshida A and Ueda T: YM155
induces caspase-8 dependent apoptosis through downregulation of
survivin and Mcl-1 in human leukemia cells. Biochem Biophys Res
Commun. 435:52–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Woo SM, Min K, Seo BR and Kwon TK: YM155
sensitizes TRAIL-induced apoptosis through cathepsin S-dependent
down-regulation of Mcl-1 and NF-κB-mediated down-regulation of
c-FLIP expression in human renal carcinoma Caki cells. Oncotarget.
7:61520–61532. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Azuhata T, Scott D, Griffith TS, Miller M
and Sandler AD: Survivin inhibits apoptosis induced by TRAIL, and
the ratio between survivin and TRAIL receptors is predictive of
recurrent disease in neuroblastoma. J Pediatr Surg. 41:1431–1440.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Latifi-Pupovci H, Kuçi Z, Wehner S, Bönig
H, Lieberz R, Klingebiel T, Bader P and Kuçi S: In vitro migration
and proliferation (‘wound healing’) potential of mesenchymal
stromal cells generated from human CD271+ bone marrow mononuclear
cells. J Transl Med. 13:315–323. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuang AA, Diehl GE, Zhang J and Winoto A:
FADD is required for DR4- and DR5-mediated apoptosis. J Biol Chem.
275:25065–25068. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ponder KG and Boise LH: The prodomain of
caspase-3 regulates its own removal and caspase activation. Cell
Death Discov. 5:562019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Salvesen GS: Caspases: Opening the boxes
and interpreting the arrows. Cell Death Differ. 9:3–5. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Maduro JH, de Vries EGE, Meersma GJ,
Hougardy BMT, van der Zee AGJ and de Jong S: Targeting
pro-apoptotic TRAIL receptors sensitizes HeLa cervical cancer cells
to irradiation-induced apoptosis. Int J Radiat Oncol Biol Phys.
72:543–552. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nakahara T, Kita A, Yamanaka K, Mori M,
Amino N, Takeuchi M, Tominaga F, Hatakeyama S, Kinoyama I,
Matsuhisa A, et al: YM155, a novel small-molecule survivin
suppressant, induces regression of established human
hormone-refractory prostate tumor xenografts. Cancer Res.
67:8014–8021. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rauch A, Hennig D, Schäfer C, Wirth M,
Marx C, Heinzel T, Schneider G and Krämer OH: Survivin and YM155:
How faithful is the liaison? Biochim Biophys Acta. 1845:202–220.
2014.PubMed/NCBI
|
|
42
|
Kelly RJ, Thomas A, Rajan A, Chun G,
Lopez-Chavez A, Szabo E, Spencer S, Carter CA, Guha U, Khozin S, et
al: A phase I/II study of sepantronium bromide (YM155, survivin
suppressor) with paclitaxel and carboplatin in patients with
advanced non-small-cell lung cancer. Ann Oncol. 24:2601–2606. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Giaccone G, Zatloukal P, Roubec J, Floor
K, Musil J, Kuta M, van Klaveren RJ, Chaudhary R, Gunther A and
Shamsili S: Multicenter phase II trial of YM155, a small-molecule
suppressor of survivin, in patients with advanced, refractory,
non-small-cell lung cancer. J Clin Oncol. 27:4481–4486. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pennati M, Sbarra S, Cesare MD, Lopergolo
A, Locatelli SL, Campi E, Daidone MG, Carlo-Stella C, Gianni AM and
Zaffaroni N: YM155 sensitizes triple-negative breast cancer to
membrane-bound TRAIL through p38 MAPK- and CHOP-mediated DR5
upregulation. Int J Cancer. 136:299–309. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Iwasa T, Okamoto I, Suzuki M, Nakahara T,
Satoh T, Fukuoka M, Ono K and Nakagawa K: Radiosensitizing effect
of YM155, a novel Small-molecule survivin suppressant, in Non-small
cell lung cancer cell lines. Clin Cancer Res. 14:6496–6504. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hong M, Ren MQ, Silva J, Paul A, Wilson
WD, Schroeder C, Weinberger P, Janik J and Hao Z: YM155 inhibits
topoisomerase function. Anticancer Drugs. 28:142–152. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nakamura H, Taguchi A, Kawana K, Baba S,
Kawata A, Yoshida M, Fujimoto A, Ogishima J, Sato M, Inoue T, et
al: Therapeutic significance of targeting survivin in cervical
cancer and possibility of combination therapy with TRAIL.
Oncotarget. 9:13451–13461. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ryu AH, Eckalbar WL, Kreimer A, Yosef N
and Ahituv N: Use antibiotics in cell culture with caution:
Genome-wide identification of antibiotic-induced changes in gene
expression and regulation. Sci Rep. 7:75332017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Winter GE, Radic B, Mayor-Ruiz C, Blomen
VA, Trefzer C, Kandasamy RK, Huber KVM, Gridling M, Chen D, Klampfl
T, et al: The solute carrier SLC35F2 enables YM155-mediated DNA
damage toxicity. Nat Chem Biol. 10:768–773. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Griffith T, Stokes B, Kucaba T, Earel J
Jr, van Oosten R, Brincks E and Norian L: TRAIL Gene therapy: From
preclinical development to clinical application. Curr Gene Ther.
9:9–19. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yamanaka K, Nakahara T, Yamauchi T, Kita
A, Takeuchi M, Kiyonaga F, Kaneko N and Sasamata M: Antitumor
activity of YM155, a selective Small-molecule survivin suppressant,
alone and in combination with docetaxel in human malignant melanoma
models. Clin Cancer Res. 17:5423–5431. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wiley SR, Schooley K, Smolak PJ, Din WS,
Huang CP, Nicholl JK, Sutherland GR, Smith TD, Rauch C, Smith CA,
et al: Identification and characterization of a new member of the
TNF family that induces apoptosis. Immunity. 3:673–682. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kim K, Fisher MJ, Xu SQ and El-Deiry WS:
Molecular determinants of response to TRAIL in killing of normal
and cancer cells. Clin Cancer Res. 6:335–346. 2000.PubMed/NCBI
|
|
54
|
Kang Z, Sun SY and Cao L: Activating Death
receptor DR5 as a therapeutic strategy for rhabdomyosarcoma. ISRN
Oncol. 2012:3959522012.PubMed/NCBI
|
|
55
|
Day TW, Najafi F, Wu CH and Safa AR:
Cellular FLICE-like inhibitory protein (c-FLIP): A novel target for
Taxol-induced apoptosis. Biochem Pharmacol. 71:1551–1561. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zong H, Yin B, Chen J, Ma B, Cai D and He
X: Over-Expression of c-FLIP confers the resistance to
TRAIL-induced apoptosis on gallbladder carcinoma. Tohoku J Exp Med.
217:203–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Surget S, Chiron D, Gomez-Bougie P,
Descamps G, Ménoret E, Bataille R, Moreau P, Gouill SL, Amiot M and
Pellat-Deceunynck C: Cell death via DR5, but not DR4, is regulated
by p53 in myeloma cells. Cancer Res. 72:4562–4573. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen S, Fu L, Raja SM, Yue P, Khuri FR and
Sun SY: Dissecting the roles of DR4, DR5 and c-FLIP in the
regulation of Geranylgeranyltransferase I inhibition-mediated
augmentation of TRAIL-induced apoptosis. Mol Cancer. 9:232010.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Refaat A, Abd-Rabou A and Reda A: TRAIL
combinations: The new ‘trail’ for cancer therapy (Review). Oncol
Lett. 7:1327–1332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gasparian ME, Chernyak BV, Dolgikh DA,
Yagolovich AV, Popova EN, Sycheva AM, Moshkovskii SA and
Kirpichnikov MP: Generation of new TRAIL mutants DR5-A and DR5-B
with improved selectivity to death receptor 5. Apoptosis.
14:778–787. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kischkel FC, Lawrence DA, Chuntharapai A,
Schow P, Kim KJ and Ashkenazi A: Apo2L/TRAIL-dependent recruitment
of endogenous FADD and caspase-8 to death receptors 4 and 5.
Immunity. 12:611–620. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Thomas LR, Henson A, Reed JC, Salsbury FR
and Thorburn A: Direct binding of Fas-associated death domain
(FADD) to the tumor necrosis Factor-related Apoptosis-inducing
ligand receptor DR5 is regulated by the death effector domain of
FADD. J Biol Chem. 279:32780–32785. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang W, Wang S, Song X, Sima N, Xu X, Luo
A, Chen G, Deng D, Xu Q, Meng L, et al: The relationship between
c-FLIP expression and human papillomavirus E2 gene disruption in
cervical carcinogenesis. Gynecol Oncol. 105:571–577. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hu Z and Ma D: The precision prevention
and therapy of HPV-related cervical cancer: New concepts and
clinical implications. Cancer Med. 7:5217–5236. 2018. View Article : Google Scholar : PubMed/NCBI
|