Introduction
In 2015, osteosarcoma (OS) was reported as the most
common type of malignancy of the bone, accounting for 5% of all
pediatric tumors and 8.9% of cancer-associated mortality in
children worldwide (1,2). OS is predominantly caused by metaphysis
of the long bones (3). In 2015, the
5-year survival rate of patients with OS was reported to be 60–70%
worldwide, due to improvements in chemotherapy and surgical
resection (1–4). However, the prognosis of OS patients
with distant metastasis remains poor, with a reported 5-year
survival rate of only 20% worldwide in 2015 (1–4).
Therefore, there is an urgent requirement to investigate the
molecular mechanisms underlying OS progression, which may assist
with the development of novel treatment strategies for OS.
The small GTPase proteins of the Rab family have
been demonstrated to serve key roles in the development and
malignant progression of numerous types of human cancer (5,6).
Ras-related protein Rab-31 (RAB31), a member of the Rab5 subfamily,
is a lipid-anchored protein consisting of 194 amino acids, which
localizes to the cytoplasmic side of the cell membrane and serves
essential roles in vesicle and granule targeting (7). Previous studies have reported that
RAB31 serves an oncogenic role in human cancer types (8–10). For
example, Pan et al (8) used
glioblastoma and cervical cancer cell lines to investigate the
function of RAB31 in cancer progression, which revealed that RAB31
promotes cancer cell proliferation, migration and
epithelial-mesenchymal transition, and inhibit cell apoptosis.
Knockdown of RAB31 was identified to suppress tumor growth in vivo.
Furthermore, Sui et al (9)
demonstrated that Rab31 promotes hepatocellular carcinoma
progression by inhibiting cell apoptosis induced by the
PI3K/AKT/Bcl-2/Bax pathway. Notably, Grismayer et al (10) identified that overexpression of RAB31
in breast cancer cells causes a switch from an invasive to a
proliferative phenotype. These findings suggest that the different
functions of RAB31 may be tumor-specific. However, to the best of
our knowledge, the expression and function of RAB31 in OS has not
previously been reported.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs that contain 22–25 nucleotides and regulate gene expression by
binding to a complementary sequence within the 3′-untranslated
region (3′UTR) of their target mRNA, which leads to translation
repression or RNA degradation (11,12). By
affecting the expression of their target genes, a large number of
miRNAs have been demonstrated to serve key roles in various
biological processes, including cell survival, differentiation,
proliferation, cell cycle, apoptosis, migration and invasion
(13–15). Recently, numerous miRNAs have been
identified to be dysregulated in OS and to serve a promoting or
suppressive role (16–19). For example, miR-195-5p inhibits OS
cell proliferation and invasion by suppressing the expression of
naked cuticle homolog 1 (19).
Previously, miR-26b has been reported to function as
a tumor suppressor (20–23). Li et al (22) reported that miR-26b inhibits the
metastasis of colorectal cancer by inhibiting fucosyltransferase 4.
miR-26b has also been identified to inhibit proliferation and
invasion of neuroglioma cells by downregulating Bcl-2 expression
and enhancing caspase-3 activity (23). In OS, miR-26b has been reported to
suppress cancer cell proliferation, migration and invasion, and
induce OS cell apoptosis by inhibiting glycolysis (20). Furthermore, miR-26b can inhibit OS
cell migration and invasion by targeting
6-phosphofructo-2-kinase/fructose-2-6-biphosphatase (PFKFB3),
connective tissue growth factor (CTGF) and SMAD family member 1
(Smad1) (21,24). However, to the best of our knowledge,
the association between miR-26b and RAB31 has not previously been
studied. Therefore, the present study aimed to evaluate the
clinical significance of RAB31 expression in OS. In addition, the
regulatory role of RAB31 in the malignant phenotype of OS cells was
investigated.
Materials and methods
Clinical tissue samples
The present study was approved by the Ethics
Committee of The Second Affiliated Hospital of Nanchang University
(Nanchang, China) and written informed consent was obtained from
all patients. OS tissue samples and matched adjacent non-tumor
tissue samples were collected from 54 patients with OS at the
Second Affiliated Hospital of Nanchang University between May 2010
and May 2013. The patients included 33 males and 21 females, with a
mean age of 18.8 years (range, 10–26 years). The clinical
characteristics of the patients are summarized in Table I. No patients received radiotherapy
or chemotherapy prior to surgery. All tissues samples were stored
at −80°C prior to use.
 | Table I.Association between RAB31 expression
and clinicopathological characteristics of patients with
osteosarcoma. |
Table I.
Association between RAB31 expression
and clinicopathological characteristics of patients with
osteosarcoma.
| Characteristic | Cases (n=54) | Low RAB31 expression
(n=33) | High RAB31 expression
(n=21) | P-value |
|---|
| Age, years |
|
|
| 0.784 |
|
<20 | 22 | 14 | 8 |
|
| ≥20 | 32 | 19 | 13 |
|
| Sex |
|
|
| 0.575 |
| Male | 33 | 19 | 14 |
|
|
Female | 21 | 14 | 7 |
|
| Tumor size, cm |
|
|
| 0.092 |
|
<8 | 24 | 18 | 6 |
|
| ≥8 | 30 | 15 | 15 |
|
| Location |
|
|
| 0.772 |
| Femur
or tibia | 37 | 22 | 15 |
|
|
Other | 17 | 11 | 6 |
|
| Lung
metastasis |
|
|
| 0.001a |
| No | 38 | 29 | 9 |
|
|
Yes | 16 | 4 | 12 |
|
| TNM stage |
|
|
| 0.018a |
|
I/IIA | 19 | 16 | 3 |
|
|
IIB/III | 35 | 17 | 18 |
|
| Serum lactate
dehydrogenase |
|
|
| 0.151 |
|
Normal | 20 | 15 | 5 |
|
|
Elevated | 34 | 18 | 16 |
|
| Serum alkaline
phosphatase |
|
|
| 0.168 |
|
Normal | 22 | 16 | 6 |
|
|
Elevated | 32 | 17 | 15 |
|
Cell culture
The normal human osteoblastic cell line hFOB 1.19
and the human OS cell lines SAOS2, U2OS, MG63 and HOS were
purchased from The Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China). hFOB 1.19 cells were
cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 (Gibco;
Thermo Fisher Scientific, Inc.) containing 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2. OS cells were maintained in DMEM with 10% FBS, 100
U/ml penicillin and 100 U/ml streptomycin at 37°C with 5%
CO2.
Cell transfection
Cells were cultured at 37°C with 5% CO2
in a 6-well plate at a density of ~1×106 cells/well.
When a confluency of 70–80% was reached, the cells were transfected
with 100 nM negative control (NC) small interfering (si)RNA (cat.
no. AM4611; Thermo Fisher Scientific, Inc.), RAB31 siRNA (cat. no.
AM16708; Thermo Fisher Scientific, Inc.), miR-NC (cat. no. 4464058;
Thermo Fisher Scientific, Inc.), miR-26b mimic (cat. no. 4464066;
Thermo Fisher Scientific, Inc.), NC inhibitor (cat. no. 4464077;
Thermo Fisher Scientific, Inc.) or miR-26b inhibitor (cat. no.
4464084; Thermo Fisher Scientific, Inc.) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Following incubation for 48 h at 37°C, subsequent experiments were
performed.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from tissue samples and
cells (1×106) using TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.). RT was performed using miScript
Reverse Transcription kit (Qiagen GmbH), according to the
manufacturer's protocol. qPCR was conducted using QuantiTect
SYBR-Green RT-PCR kit (Qiagen GmbH) with an ABI 7500 fast real-time
PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The thermocycling conditions for PCR were as follows: 95°C for 1
min, followed by 40 cycles of 95°C for 15 sec and 60°C for 30 sec.
GAPDH or U6 was used as the internal reference for mRNA and miRNA,
respectively. Data were quantified using the 2−ΔΔCq
method (25). The primer sequences
were as follows: RAB31 forward, 5′-GGGGTTGGGAAATCAAGCATC-3′ and
reverse, 5′-GCCAATGAATGAAACCGTTCCT-3′; GAPDH forward,
5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse,
5′-GCCATCACGCCACAGTTTC-3′. Primers for U6 (cat. no. HmiRQP9001) and
miR-16 (cat. no. HmiRQP0356) were obtained from Guangzhou FulenGen,
Co., Ltd.
Western blot analysis
Tissue samples and cells (1×107) were
solubilized with radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology). The protein concentration
was measured using a BCA protein assay kit (Beyotime Institute of
Biotechnology). Total protein (50 µg) was separated by 10% SDS-PAGE
and then transferred to polyvinylidene difluoride membranes (EMD
Millipore). The membranes were blocked with 5% non-fat milk in 0.1%
TBS/Tween-20 at room temperature for 1 h, followed by incubation
with rabbit anti-human RAB31 (dilution, 1:100; cat. no. ab230881;
Abcam), or rabbit anti-human GAPDH (dilution, 1:200; cat. no.
ab181602; Abcam) primary antibodies at room temperature for 3 h.
Subsequently, the membranes were incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody
(dilution, 1:5,000; cat. no. ab6721; Abcam) at room temperature for
1 h. Enhanced chemiluminescent reagent (Pierce; Thermo Fisher
Scientific, Inc.) was used to detect the signals on the membranes.
ImageJ software (v1.46; National Institutes of Health) was used for
densitometry analysis.
Cell proliferation assay
Cell proliferation was investigated using Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.),
according to the manufacturer's protocol. A total of 48 h after
transfection, cells (3,000 cells/well) were seeded into 96-well
plates and cultured for 0, 24, 48 or 72 h. The optical density
value at 450 nM was measured using a microplate reader.
Cell apoptosis assay
The transfected cells were harvested and washed
twice with ice-cold PBS. Subsequently, 1×106 cells were
resuspended with 300 µl binding buffer. Following staining with
Annexin V-fluorescein isothiocyanate and propidium iodide (BD
Biosciences), according to the manufacturer's protocol, apoptotic
cells were analyzed using a flow cytometer and Accuri C6 Software
(version 1.0; BD Biosciences).
Wound healing assay
The transfected cells (1×106 cells per
well) were cultured in 6-well plates for 24 h. Subsequently, an
artificial wound was created using a sterile 10 µl pipette tip in
the confluent cell monolayer. Photographs were captured at 0 and 24
h using an inverted light microscope (magnification, ×40).
Cell invasion assay
Matrigel (BD Biosciences) was diluted with
serum-free DMEM and added to the upper chamber of a 24-well
Transwell™ chamber (Corning Inc.), which was then incubated at 37°C
for 1 h. Cells were diluted to a density of 2×105
cells/ml with serum-free DMEM. Subsequently, 100 µl cell suspension
was added to the upper chamber and 700 µl DMEM containing 10% FBS
was added to the bottom chamber. Following incubation at 37°C for
24 h, the chamber was washed with PBS. The cells were then fixed
with 4% paraformaldehyde at room temperature for 30 min, stained
with crystal violet at room temperature for 30 min and photographs
were obtained using an inverted light microscope (magnification,
×400).
Bioinformatics analysis and luciferase
reporter assay
TargetScan software version 7.1 was used for
bioinformatics analysis (26). The
wild-type (WT) or mutant type (MUT) 3′-UTR of RAB31 was inserted
downstream of the luciferase reporter gene in the pMIR-REPORT
vector (Thermo Fisher Scientific, Inc.), generating WT-RAB31 and
MUT-RAB31. MG63 and U2OS cells were co-transfected with miR-26b
mimics/miR-NC, WT-RAB31/MUT-RAB31 and pRL-SV40 (Promega
Corporation) expressing Renilla luciferase. Following 48 h
incubation at 37°C with 5% CO2, the luciferase activity
was measured using the Dual-Luciferase® Reporter Assay
System (Promega Corporation).
Statistical analysis
All data are presented as the mean ± standard
deviation from a minimum of three independent experiments.
Statistical analysis was performed using SPSS 20.0 (IBM Corp.).
Student's t-test was used for comparisons between two groups.
Multiple comparisons were performed using one-way analysis of
variance followed by Tukey's test. The patients with OS were
divided into a high RAB31 expression group and low RAB31 expression
group using the median expression level of RAB31 (1.91) as the
cut-off, and χ2 test was used to perform statistical
analysis in Table I. Correlations
between the mRNA expression levels of miR-26b and RAB3 were
analyzed using Pearson's correlation coefficient. P<0.05 was
considered to indicate a statistically significant difference.
Results
Increased expression levels of RAB31
are associated with OS progression
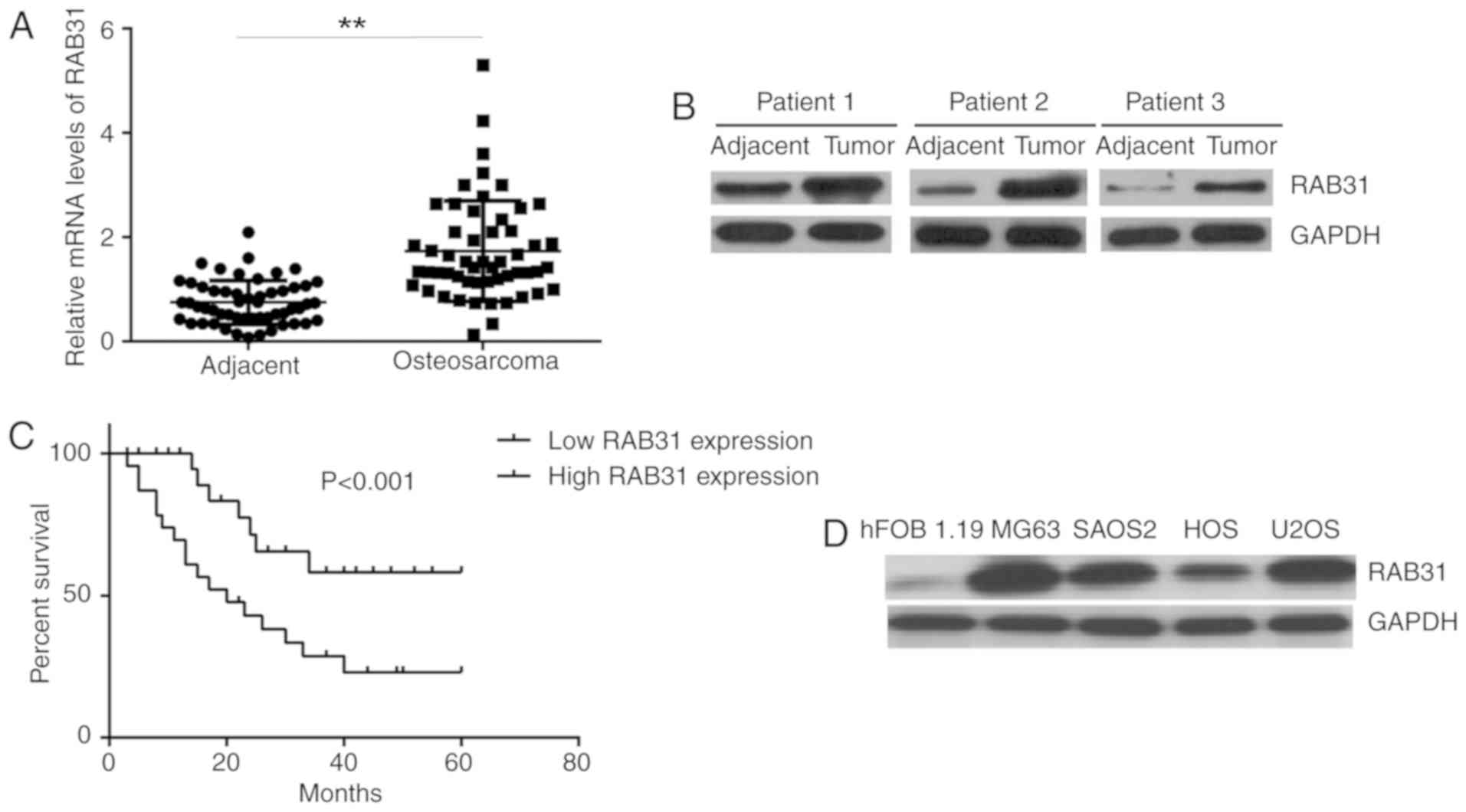
First, the mRNA and protein expression levels of
RAB31 were examined in tissue samples from OS patients. RT-qPCR and
western blot analysis demonstrated that the expression levels of
RAB31 were significantly higher in OS tissues compared with
adjacent non-tumor tissues (Fig. 1A and
B). The patients with OS were then divided into a high RAB31
expression group and low RAB31 expression group using the median
expression level of RAB31 (1.91) as the cut-off. As presented in
Table I, high expression of RAB31
was identified to be significantly associated with lung metastasis
and advanced clinical stage. Furthermore, the patients with high
RAB31 expression exhibited a significantly shorter survival time
compared with those with low RAB31 expression (Fig. 1C), which suggested that RAB31
upregulation was associated with a poor prognosis for patients with
OS. In addition, the protein expression levels of RAB31 were
markedly higher in the OS cell lines SAOS2, U2OS, MG63 and HOS
compared with the normal human osteoblastic cell line hFOB 1.19
(Fig. 1D). MG63 and U2OS cells
exhibited the highest expression levels of RAB31 protein (Fig. 1D); these cell lines were therefore
selected for subsequent experiments.
Silencing of RAB31 inhibits OS cell
proliferation, cell cycle progression, migration and invasion, and
induces cell apoptosis
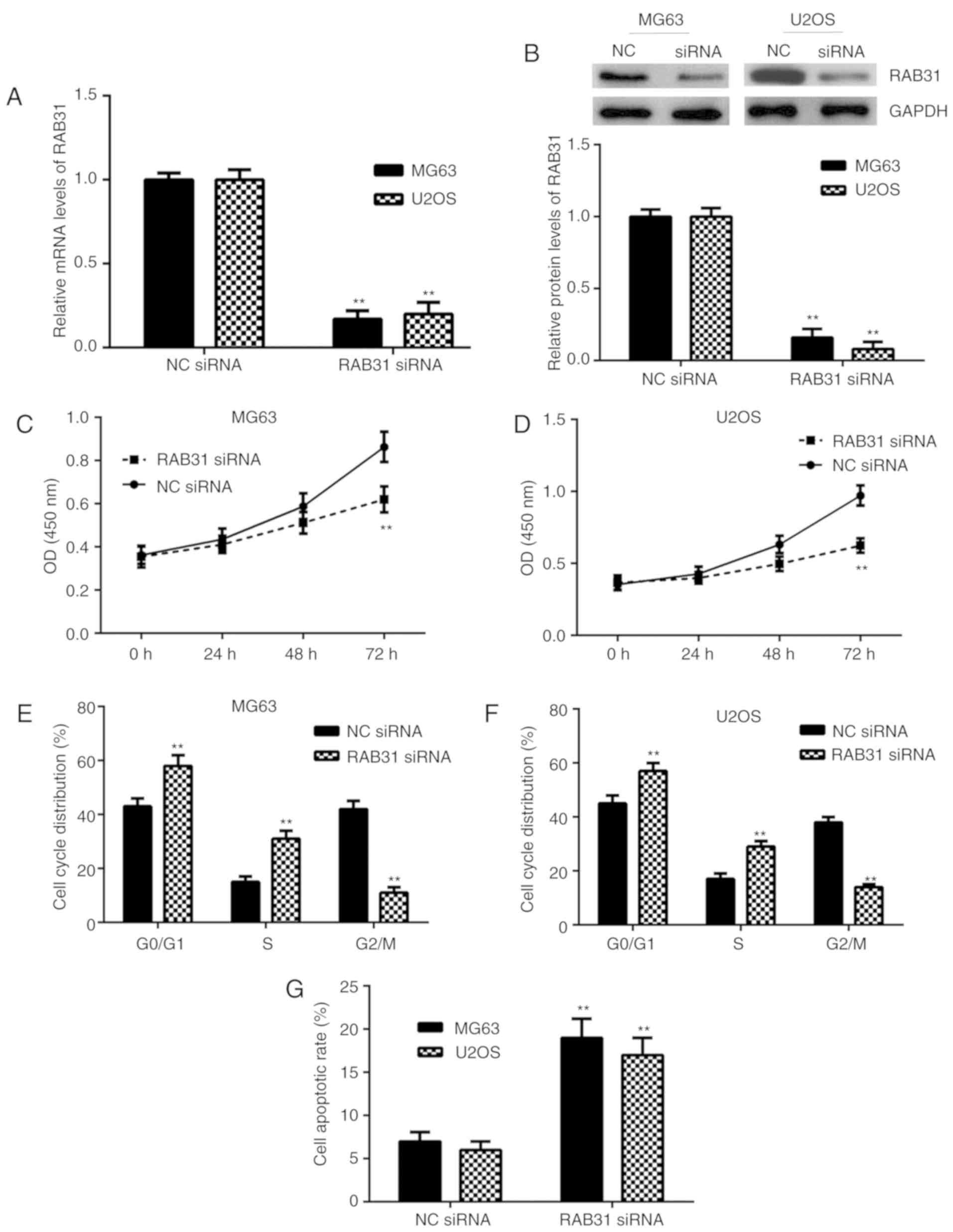
To inhibit the expression of RAB31, MG63 and U2OS
cells were transfected with RAB31 siRNA. As presented in Fig. 2A and B, the mRNA and protein
expression levels of RAB31 were significantly lower in MG63 and
U2OS cells transfected with RAB31 siRNA compared with the NC siRNA
group. A CCK-8 assay was then performed to examine cell
proliferation. As presented in Fig. 2C
and D, knockdown of RAB31 significantly inhibited OS cell
proliferation. The cell cycle distribution and cell apoptosis were
then examined by flow cytometry. The data from this experiment
indicated that silencing of RAB31 was significantly associated with
cell cycle arrest at the G1 stage and significantly promoted OS
cell apoptosis (Fig. 2E-H). The
aforementioned results suggested that RAB31 may serve a promoting
role in OS growth.
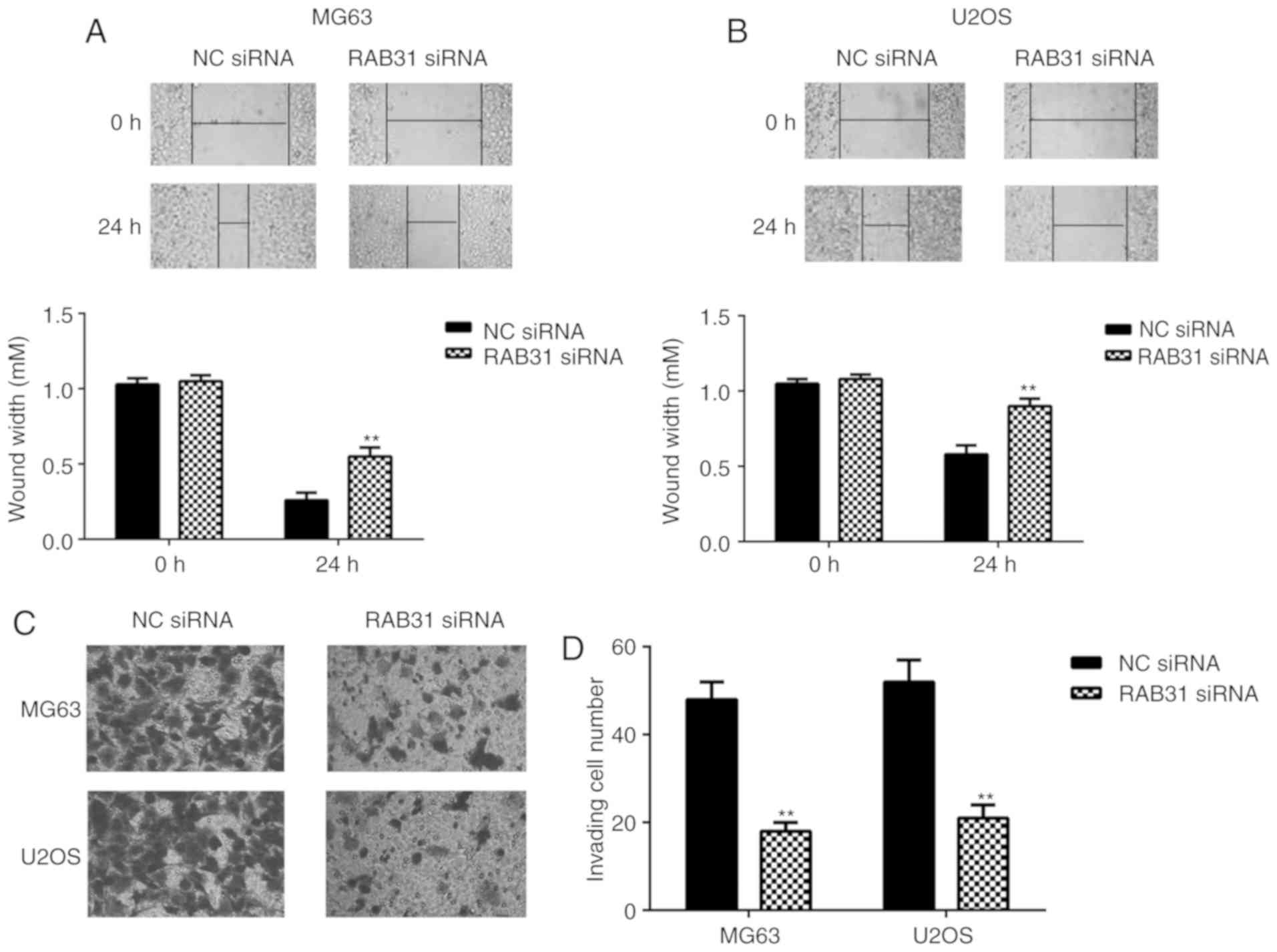
To study the potential role of RAB31 in OS
metastasis, a wound healing assay and Transwell assay were
performed to examine cell migration and invasion, respectively. As
presented in Fig. 3, silencing of
RAB31 expression significantly inhibited the migration and invasion
of MG63 and U2OS cells compared with the NC siRNA group. In
summary, RAB31 silencing inhibited OS cell proliferation, cell
cycle progression, migration and invasion, and induced cell
apoptosis.
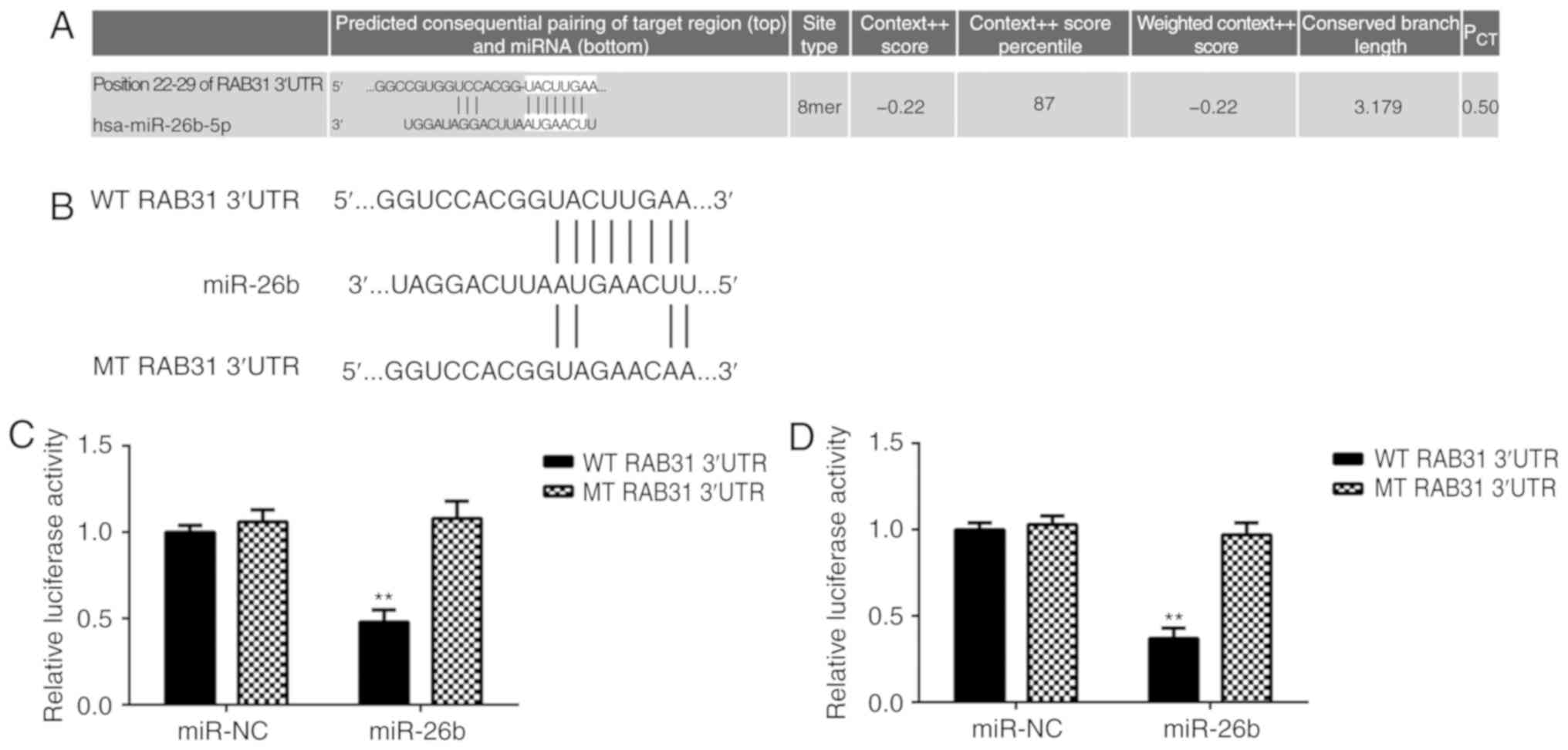
RAB31 is a target gene of miR-26b in
OS cells
TargetScan software data predicted that RAB31 is a
putative target gene of miR-26b (Fig.
4A). To confirm the association between RAB31 and miR-26b,
wild-type (WT) and mutant (MT) RAB31 luciferase reporter plasmids
were generated (Fig. 3B) and a
luciferase reporter gene assay was performed. The data demonstrated
that transfection with miR-26b mimic significantly inhibited the
luciferase activity in MG63 and U2OS cells transfected with WT
RAB31 3′UTR reporter plasmid; however, no effect was observed on
the luciferase activity in cells transfected with the MT RAB31
3′UTR reporter plasmid (Fig. 4C and
D). This indicates that RAB31 is a target gene of miR-26b in OS
cells.
RAB31 is negatively regulated by
miR-26b in OS cells
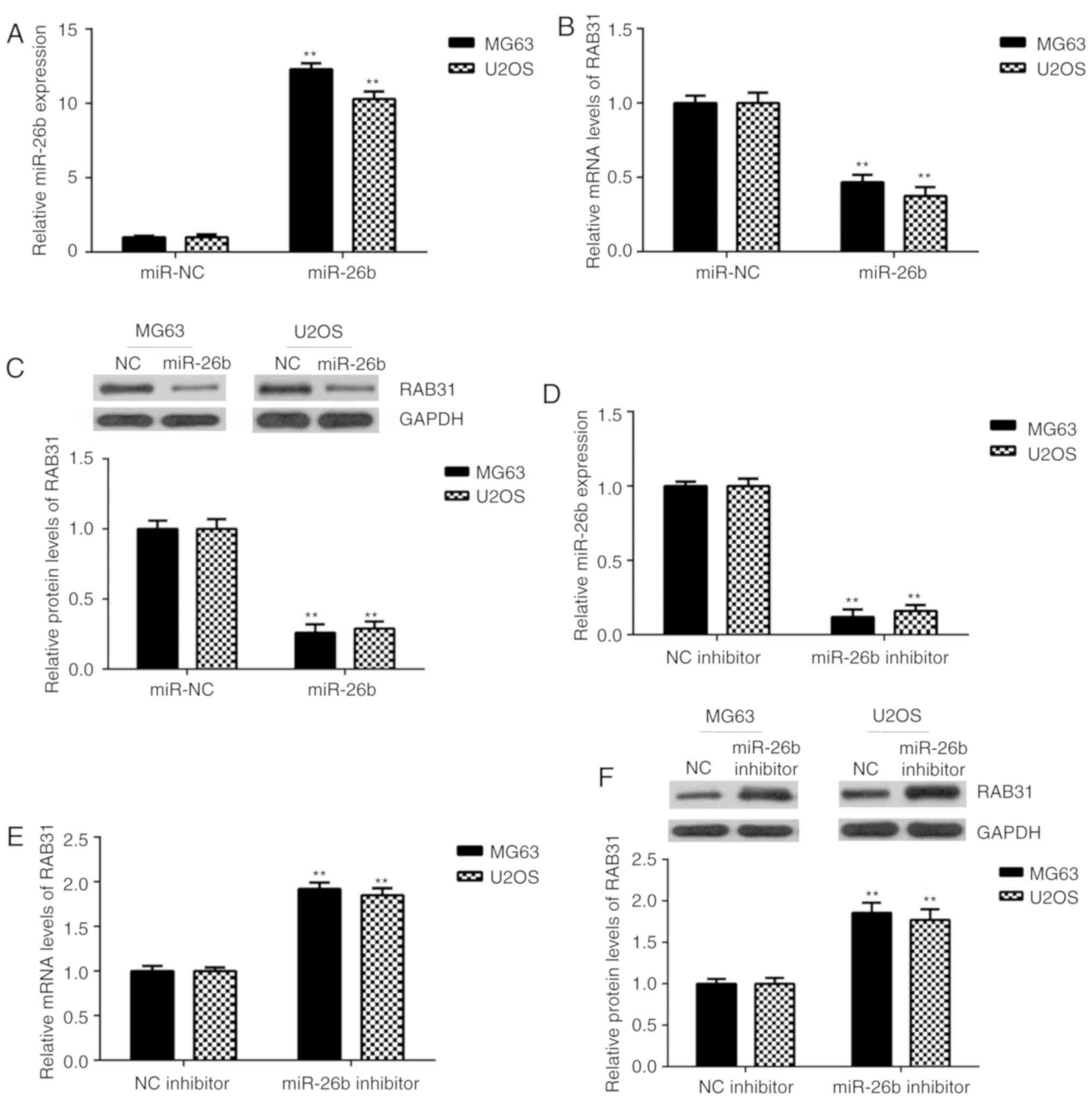
The effects of miR-26b on RAB31 expression were then
investigated in OS cells. MG63 and U2OS cells were transfected with
miR-26b mimic or miR-NC. Following transfection, miR-26b expression
was significantly upregulated in the miR-26b mimic group compared
with the miR-NC group (Fig. 5A).
Further experiments revealed that the mRNA and protein expression
levels of RAB31 were significantly decreased following miR-26b
overexpression (Fig. 5B and C). To
further confirm these findings, MG63 and U2OS cells were
transfected with miR-26b inhibitor or NC inhibitor. RT-qPCR
analysis revealed that miR-26b was significantly downregulated in
the miR-26b inhibitor-transfected group compared with the NC
inhibitor-transfected group (Fig.
5D). As presented in Fig. 5E and
F, the mRNA and protein expression levels of RAB31 were
significantly increased in the miR-26b inhibitor-transfected group
compared with the NC inhibitor-transfected group. These data
indicated that RAB31 was negatively regulated by miR-26b in OS
cells.
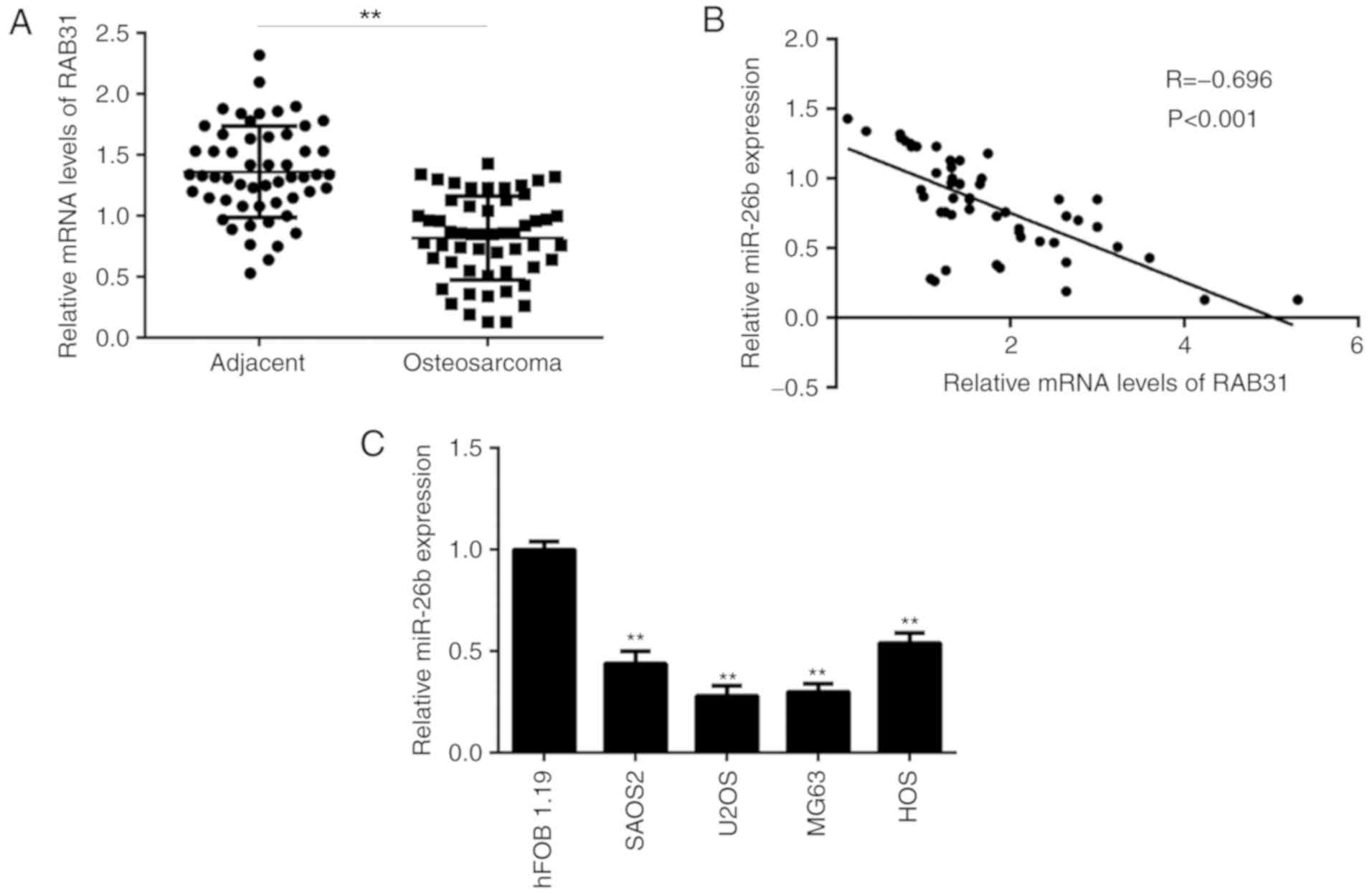
miR-26b expression is inversely
correlated with RAB31 expression in OS tissues
RT-qPCR analysis identified that miR-26b levels were
significantly lower in OS tissues compared with matched adjacent
non-tumor tissues (Fig. 6A).
Additionally, an inverse correlation was identified between the
miR-26b and RAB31 expression levels in OS tissues (Fig. 6B), which suggested that the increased
expression levels of RAB31 in OS tissues may be due to
downregulation of miR-26b. Furthermore, miR-26b levels were
demonstrated to be decreased in OS cell lines compared with hFOB
1.19 cells (Fig. 6C).
Discussion
To the best of our knowledge, the function of RAB31
in OS has previously been unclear. The present study revealed that
RAB31 was upregulated in OS tissues and cell lines, and that high
RAB31 expression levels were associated with malignant progression
and a poor prognosis for patients with OS. Silencing of RAB31
expression significantly inhibited OS cell proliferation, cell
cycle progression, migration and invasion, and induced cell
apoptosis. RAB31 was then identified as a direct target gene of
miR-26b. Additionally, the expression of RAB31 was negatively
regulated by miR-26b in OS cells. Furthermore, miR-26b was
downregulated in OS tissues and reversely correlated to the
expression levels of RAB31.
Initially, RAB31 was reported to be upregulated in
breast cancer and high expression of RAB31 is significantly
associated with worse outcome for patients (27). Furthermore, the mucin 1 C-terminal
subunit (MUC1-C) oncoprotein and RAB31 have been demonstrated to
function in an autoinductive loop that contributes to
overexpression of MUC1-C in breast cancer cells (28). Grismayer et al (10) reported that RAB31 overexpression
increases breast cancer cell proliferation, reduces adhesion and
invasion in vitro, and results in decreased lung metastasis
formation in vivo. In addition, RAB31 has been revealed to promote
hepatocellular carcinoma progression by inhibiting tumor cell
apoptosis induced by the PI3K/AKT/Bcl-2/Bax pathway (9). However, no previous study has focused
on the function of RAB31 in OS.
The present study first examined the expression
levels of RAB31 in OS tissues and cell lines. The data demonstrated
that RAB31 was significantly upregulated in OS tissues compared
with adjacent non-tumor tissues. Furthermore, it was identified
that high expression of RAB31 was significantly associated with
lung metastasis and advanced clinical stage in patient with OS,
which suggested that an upregulation of RAB31 may contribute to OS
progression. Additionally, patients with OS exhibiting high RAB31
expression had shorter survival times compared with patients
exhibiting low RAB31 expression, which indicates that RAB31 may be
used as a novel predicator for the prognosis of patients with OS.
Consistent with the clinical data, RAB31 was also significantly
upregulated in OS cell lines compared with the normal human
osteoblastic hFOB 1.19 cell line. To further clarify the function
of RAB31 in OS, two common OS cell lines MG63 and U2OS were used to
perform in vitro experiments. The results demonstrated that
silencing of RAB31 expression significantly reduced the
proliferation, cell cycle progression, migration and invasion of OS
cells, and induced cell apoptosis. Therefore, the current results
suggested that RAB31 may have a promoting role in regulating the
malignant phenotype of OS cells. These findings indicate that
targeting RAB31 may be a promising strategy for the treatment of
OS.
The mechanism underlying RAB31 expression in OS was
further investigated in the current study. As a large number of
genes are regulated by miRNAs (11,12),
bioinformatics analysis was used to predict the putative miRNAs
that may directly target RAB31. Among the predicted miRNAs, miR-26b
was selected as it has been reported to function as a tumor
suppressor in OS (24). Duan et al
(24) identified that miR-26b
inhibits OS metastasis by targeting CTGF and Smad1. Zheng et al
(21) reported that miR-26b
represses OS cell migration and invasion by inhibiting the
expression of PFKFB3. In addition, miR-26b has been demonstrated to
inhibit OS cell proliferation, migration and invasion, and induce
cell apoptosis by inhibiting PFKFB3-driven glycolysis (20). To further clarify the association
between miR-26b and RAB31, the present study performed a luciferase
reporter gene assay, which confirmed that RAB31 was a target gene
of miR-26b. In addition, it was identified that the expression of
RAB31 was negatively regulated by miR-26b in OS cells and that
miR-26b levels were inversely correlated with the expression levels
of RAB31 in OS tissues. These findings suggested that
downregulation of miR-26b may contribute to the upregulation of
RAB31 in OS. A limitation of the current study is that the effects
of miR-26b on proliferation, apoptosis, migration and invasion were
not investigated; this should be performed in future studies.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that RAB31 was
upregulated in OS, was significantly associated with malignant
progression and a poor prognosis for patients with OS, and that
RAB31 silencing inhibited the malignant phenotype of OS cells.
Furthermore, a novel association between RAB31 and miR-26b, a tumor
suppressor in OS, was identified. Therefore, the present results
suggested that RAB31 may be a promising therapeutic target for
OS.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QW designed the study and wrote the manuscript. QF
collected sample tissues and performed statistical analysis. YX and
XL performed the experiments. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Second Affiliated Hospital of Nanchang University
(Nanchang, China). Written informed consent was obtained from all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Saraf AJ, Fenger JM and Roberts RD:
Osteosarcoma: Accelerating Progress Makes for a Hopeful Future.
Front Oncol. 8:42018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harrison DJ, Geller DS, Gill JD, Lewis VO
and Gorlick R: Current and future therapeutic approaches for
osteosarcoma. Expert Rev Anticancer Ther. 18:39–50. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shaughnessy R and Echard A: Rab35 GTPase
and cancer: Linking membrane trafficking to tumorigenesis. Traffic.
19:247–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang S, Hu C, Wu F and He S: Rab25 GTPase:
Functional roles in cancer. Oncotarget. 8:64591–64599. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bao X, Faris AE, Jang EK and Haslam RJ:
Molecular cloning, bacterial expression and properties of Rab31 and
Rab32. Eur J Biochem. 269:259–271. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan Y, Zhang Y, Chen L, Liu Y, Feng Y and
Yan J: The Critical role of Rab31 in cell proliferation and
apoptosis in cancer progression. Mol Neurobiol. 53:4431–4437. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sui Y, Zheng X and Zhao D: Rab31 promoted
hepatocellular carcinoma (HCC) progression via inhibition of cell
apoptosis induced by PI3K/AKT/Bcl-2/BAX pathway. Tumour Biol.
36:8661–8670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grismayer B, Sölch S, Seubert B, Kirchner
T, Schäfer S, Baretton G, Schmitt M, Luther T, Krüger A, Kotzsch M,
et al: Rab31 expression levels modulate tumor-relevant
characteristics of breast cancer cells. Mol Cancer. 11:622012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moss EG: MicroRNAs: Hidden in the genome.
Curr Biol. 12:R138–R140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L, Xu J, Yang G, Li H and Guo X:
miR-202 inhibits cell proliferation, migration, and invasion by
targeting EGFR in human bladder cancer. Oncol Res. 26:949–957.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang C, Wu K, Wang S and Wei G: Long
non-coding RNA XIST promotes osteosarcoma progression by targeting
YAP via miR-195-5p. J Cell Biochem. 119:5646–5656. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou Y, Yang C, Wang K, Liu X and Liu Q:
MicroRNA-33b inhibits the proliferation and migration of
osteosarcoma cells via targeting hypoxia-inducible factor-1α. Oncol
Res. 25:397–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang R and Xia T: Long non-coding RNA
XIST regulates PDCD4 expression by interacting with miR-21-5p and
inhibits osteosarcoma cell growth and metastasis. Int J Oncol.
51:1460–1470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qu Q, Chu X and Wang P: MicroRNA-195-5p
suppresses osteosarcoma cell proliferation and invasion by
suppressing naked cuticle homolog 1. Cell Biol Int. 41:287–295.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Du JY, Wang LF, Wang Q and Yu LD: miR-26b
inhibits proliferation, migration, invasion and apoptosis induction
via the downregulation of
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-3 driven
glycolysis in osteosarcoma cells. Oncol Rep. 33:1890–1898. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng WD, Zhou FL and Lin N: MicroRNA-26b
inhibits osteosarcoma cell migration and invasion by
down-regulating PFKFB3 expression. Genet Mol Res. 14:16872–16879.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Sun Z, Liu B, Shan Y, Zhao L and Jia
L: Tumor-suppressive miR-26a and miR-26b inhibit cell
aggressiveness by regulating FUT4 in colorectal cancer. Cell Death
Dis. 8:e28922017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li YP, Dai WM, Huang Q, Jie YQ, Yu GF, Fan
XF, Wu A and Mao DD: Effects of microRNA-26b on proliferation and
invasion of glioma cells and related mechanisms. Mol Med Rep.
16:4165–4170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duan G, Ren C, Zhang Y and Feng S:
MicroRNA-26b inhibits metastasis of osteosarcoma via targeting CTGF
and Smad1. Tumour Biol. 36:6201–6209. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:42015. View Article : Google Scholar
|
|
27
|
Kotzsch M, Sieuwerts AM, Grosser M, Meye
A, Fuessel S, Meijer-van Gelder ME, Smid M, Schmitt M, Baretton G,
Luther T, et al: Urokinase receptor splice variant
uPAR-del4/5-associated gene expression in breast cancer:
Identification of rab31 as an independent prognostic factor. Breast
Cancer Res Treat. 111:229–240. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jin C, Rajabi H, Pitroda S, Li A,
Kharbanda A, Weichselbaum R and Kufe D: Cooperative interaction
between the MUC1-C oncoprotein and the Rab31 GTPase in estrogen
receptor-positive breast cancer cells. PLoS One. 7:e394322012.
View Article : Google Scholar : PubMed/NCBI
|