Introduction
Chronic myeloid leukemia (CML) is a
myeloproliferative disorder that stems from myeloid
CD34+/CD38–/CD44+ progenitors in
the bone marrow (BM) (1). The
incomplete differentiation of hematopoietic stem cells in CML
results in immature leucocytes in both the BM and peripheral blood
(PB) (2). Translocation of the
abelson (ABL1) gene at chromosome 9 produces the so called
‘Philadelphia chromosome,’ which is considered the main cause of
CML (3). This mutation activates the
BCR-ABL1 fusion oncoprotein, which modulates a number of signaling
pathways and enables stem cells to avoid cell death. Furthermore,
this potent tyrosine kinase (TK) activates different signaling
pathways, including the Wnt/β-catenin, Bcl-2 and JAK/STAT MAPK/JNK
signaling pathways, that are responsible for the pathogenesis of
CML (3–5). Thus, BCR-ABL1 is considered a
therapeutic chemotherapy target for CML, using the imatinib class
of drugs. However, the main drawback of therapeutic TK inhibitors
is the severe side effects exhibited in patients with CML (6), including cardiovascular, pulmonary,
gastrointestinal and endocrine toxicities (7), as well as development of resistance
(8). Thus, it is essential to
investigate alternative agents to minimize the side effects.
Complementary and alternative medicine, extensively
used for the treatment of disease (9), may be an alternative approach in
decreasing the side effects of patients with cancer undergoing
chemotherapy, which compromises patients quality of life (10). Natural compounds are well recognized
for the management of several diseases, including CML (11). Mushroom-derived compounds have the
ability to act as anti-leukemia, antiviral, antioxidant and
anti-inflammatory agents, and exert tumor apoptosis-inducing
properties (12). Additionally, it
has been reported that chemical substances derived from
microorganisms, plants and animals possess anticancer potential
(13). Thus, it is of great interest
to investigate whether there is anti-CML potential associated with
different mushrooms and determine the underlying molecular
mechanism of action.
Agaricus blazei Murill (AbM) has been
extensively applied in alternative medicine in Brazil since 1960
(11). It has been reported that the
putative active functions of AbM lie in relieving atopic dermatitis
and decreasing glycosuria (9).
Although several polysaccharides contain AbM, including dextran,
mannan and heteropolysaccharides (14,15), the
molecular mechanism underlying such agents in anticancer activity
is yet to be investigated. The most useful compound derived from
AbM for anticancer activity is a proteoglucan, acid RNA protein
complex, FA-2-b-β, which functions in regulating natural killer
cell and macrophage activity (16).
In addition, AbM-derived linoleic acid is a bactericidal agent that
regulates host immunity, enhancing helper T cell expansion and
natural killer cell activation, and helping to decrease the
percentage of body fat and visceral fat, as well as blood
cholesterol and glucose levels in human volunteers of clinical
research (14,17). This evidence strongly supports the
use of AbM as a health-promoting supplement for immunomodulation
(14,17). Steroids have also been identified as
an effective component in AbM against acute monocytic leukemia
(18).
The Wnt/β-catenin signaling pathway is involved in
embryogenesis, cell development, proliferation, differentiation and
self-renewal, and homeostasis of multiple organ systems (19). Thus, dysregulation of the
Wnt/β-catenin signaling pathway is closely associated with
tumorigenesis (20). For example,
upregulation of β-catenin is observed in CML, with the expansion of
the BCR/ABL gene (21). Conversely,
inhibition of β-catenin expression decreases drug resistance in
CML, with delayed entry into the blast phase (22).
However, the ability of AbM to inhibit human
leukemia in vitro remains unknown. Thus, the present study
aimed to investigate the effect of FA-2-b-β, derived from AbM, in
the induction of apoptosis in human leukemia cells in vitro,
and considered whether such a potential effect targeted the
Wnt/β-catenin signaling pathway. The results of the present study
provide insight on naturally-derived compounds in the development
of a novel therapeutic target in the management of CML.
Materials and methods
Cell culture and reagents
The CML K562 cells and primary CML BM cells from
four female patients and one male patient with CML (mean age, 48
years; age range, 40–56 years) were collected. CML K562 cells were
purchased from Lanzhou University Medical School (Lanzhou, China),
and primary CML BM cells were purchased from Gansu Provincial
People's Hospital (Lanzhou, China). The BM samples from patients
with chronic leukemia were extracted and isolated by density
gradient centrifugation at 1,000 × g (1,000 r/min) at 4°C for 3 h,
using a sterile operating table, after obtaining written informed
consent from the patients. The cell density was adjusted to
5×106/ml. Either the K562 or the primary CML BM cells
were maintained in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc), at 37°C in an atmosphere with 5%
CO2. Sub-culturing of the cells was performed when 80%
confluence was reached.
Details of the primary antibodies are as follows:
Anti-c-myc antibody (1:1,000; cat. no. 5605; Cell Signaling
Technology, Inc.), anti-cyclin D1 antibody (1:1,000; cat. no. 2978;
Cell Signaling Technology, Inc.), anti-CD44 antibody (1:1,000; cat.
no. 3570; Cell Signaling Technology, Inc.), anti-c-Jun antibody
(1:1,000; cat. no. 9165; Cell Signaling Technology, Inc.), anti-MMP
antibody (1:1,000; cat. no. 3801; Cell Signaling Technology, Inc.),
anti-Lef/Tcf antibody (1:1,000; cat. no. 2203; Cell Signaling
Technology, Inc.), anti-Bcl-2 antibody (1:1,000; cat. no.
12789-1-AP; ProteinTech Group, Inc.), anti-Bax antibody (1:1,000;
cat. no. ab32503; Abcam), anti-β-actin antibody (1:1,000; cat. no.
GB-12001; Beijing Biosynthesis Biotechnology Co., Ltd.),
anti-β-catenin antibody (1:1,000; cat. no. ab22656; Abcam). The
HRP-linked goat anti-rabbit IgG (1:4,000; cat. no. 7074) or goat
anti-mouse IgG (1:1,000; cat. no. 7076) were purchased from Cell
Signaling Technology, Inc.
AbM was supplied by the Edible Mushroom Center of
Shanxi Agricultural University (Shanxi, China) (23). The RNA-protein complex, FA-2-b-β, was
extracted in the School of Chemistry and Chemical Engineering,
Lanzhou University (Lanzhou, China), as previously described
(24). The purified compounds were
obtained from Lanzhou University using ethanol precipitation, gel
filtration, DEAE-cellulose and Sephadex G-200 column chromatography
(15). The fraction FA-2-b-β was
dissolved in distilled water for subsequent experimentation.
Cell proliferation assay
K562 cells or primary CML BM cells (10,000) were
seeded into 96-well plates in a total of 100 µl and cultured with
FA-2-b-β. FA-2-b-β at the concentrations of 1.2, 1.5, 1.8, 2.1 and
2.4 mg/ml or vehicle only (RPMI-1640) were cultured at 37°C for 24,
48 and 72 h. To prevent medium evaporation, 100 μl PBS was added
into each well. Cell proliferation was assessed via the Cell
Counting Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.)
according to the manufacturers protocol (25). The solution (10 µl/ml) was added and
incubated for 1 h at 37°C. The solution in the wells was aspirated
gently following centrifugation at 500 × g for 5 min at 4°C.
Following mixing for 10 min, the absorbance was measured at 450 nm
using an ELX800 absorbance microplate reader (BioTek Instruments,
Inc.). Each assay was repeated five times.
Apoptosis and cell cycle assay
For the apoptosis assay, K562 cells or primary CML
BM cells (100,000) were seeded into 6-well plates with different
concentrations of FA-2-b-β, at a dose range of 0, 1.2, 1.5, 1.8 and
2.4 mg/ml and time periods of 24 and 48 h. These cells were
harvested after 24 or 48 h of treatment and permeabilized using
Triton-X, washed and resuspended in binding buffer (50 mM HEPES,
700 mM NaCl, 12.5 mM CaCl2 pH 7.4 in 100 µl) with the
addition of 2.5 µl Annexin V (cat. no. R8021) and 5 µl PI (cat. no.
EZ2811E231) from an Annexin V-FITC Apoptosis Detection kit (BD
Biosciences). Briefly, 100,000 cells were washed with ice-cold PBS,
resuspended in 195 µl PBS and stained for 15 min at room
temperature with 5 µl FITC conjugated anti-Annexin V antibody.
Unbound Annexin V antibody was removed by washing with binding
buffer. The percentage of apoptotic K562 cells and primary CML BM
cells (Annexin V-positive) was determined by flow cytometry
analysis. Apoptotic cells were pre-treated at 37°C in the presence
or absence of 3 mmol/l N-acetylcysteine (NAC; BD Biosciences) for
cell cycle analysis for 2 h, followed by treatment with different
concentrations of FA-2-b-β for 24 or 48 h at 37°C. Subsequently,
the supernatant was removed, and the treated cells were washed
three times with PBS and fixed using 70% ethanol overnight at 4°C.
The cells were washed with PBS buffer, incubated with 5 µl RNase
(0.25 mg/ml; BD Biosciences) for 30 min at 37°C. The pelleted cells
were resuspended in PBS-diluted (1:100) Annexin V-FITC Apoptosis
reagent (50 µg/ml) and incubated for 15 min at room temperature in
the dark for flow cytometry (BD FACSuite v1.0.538.41; BD
Biosciences).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA from K562 and primary CML BM cells was
extracted using TRIzol® reagent (Takara Bio, Inc).
RNase-free DNase I (Promega Corporation) was used to remove the
genomic DNA. RT was performed at 70°C for 5 min using the
PrimeScript RT MasterMix Reverse transcriptase cDNA synthesis kit
(cat. no. SD2193; Takara Bio, Inc.). The Real Time PCR reaction was
performed with SYBRGreen (cat. no. SD3034; Takara Bio, Inc.) using
the qPCR Applied Biosystems 7500 Real-time System StepOne PLUS with
the following thermocycling conditions: Initial denaturation at
95°C for 4 min, then 40 cycles at 95°C for 15 sec and 60°C for 45
sec. β-actin was used as the internal control for AbM inhibition of
apoptosis determining Bcl-2, Bax and β-catenin expression. Primers
were designed by Takara Bio, Inc, and are presented in Table I. The expression levels of the
aforementioned genes were determined using the 2−ΔΔCq
method (26).
 | Table I.PCR primer sequences. |
Table I.
PCR primer sequences.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| Actin |
TGGCACAAAGCACAATGAA |
CTAAGTCATAGTCCGCCTAGAAGC |
| β-catenin |
GTGCATCTACACCGACAACTCC |
GTTCCACTTGAGCTTGTTCACC |
| Bcl-2 |
GGATTGTGGCCTTCTTTGAG |
TACCCAGCCTCCGTTATCCT |
| Bax |
CCGATTCATCTACCCTGCTG |
TGAGCCAATTCCAGAGGCAGT |
Western blot analysis
K562 and primary CML BM cells were seeded into a
6-well plate and incubated with FA-2-b-β for 24/48 h at 37°C. Cells
were lysed with SDS lysis buffer (cat. no. P1200; Beijing Solarbio
Science & Technology Co., Ltd.) and total protein was obtained.
Protein concentration was measured using a bicinchoninic acid
protein assay. After protein concentration was measured, equal
amounts of protein (10 µg/lane) were separated via 8–12% SDS-PAGE
and transferred onto polyvinylidene fluoride membranes (EMD
Millipore). Membranes were blocked with 5% skimmed dried milk for 1
h at room temperature, and then incubated with the appropriate
primary antibodies overnight at 4°C. After primary antibody
incubation, the membranes were washed three times with TBS-Tween
(0.05% Tween-20; Boster Biological Technology), and then the
corresponding HRP-linked secondary antibody was added for 2 h at
room temperature. Apoptosis protein bands were detected using
enhanced chemiluminescence (ECL kit; cat. no. P10010; Molecular
Biotech Co., Ltd.) according to the manufacturers protocol and
analyzed using Image Lab™ software v6.0 (Bio-Rad Laboratories,
Inc.).
Statistical analysis
Statistical analysis was performed using SPSS
software (version 18; SPSS, Inc.). All quantitative data were
analyzed using one-way ANOVA, followed by Dunnett's post hoc
multiple comparison test to compare all columns with the control
column. Data shown are representative of three independent
experiments with cells. Data are presented as the mean ± SD.
P<0.05 and P<0.001 were considered to indicate statistically
significant differences.
Results
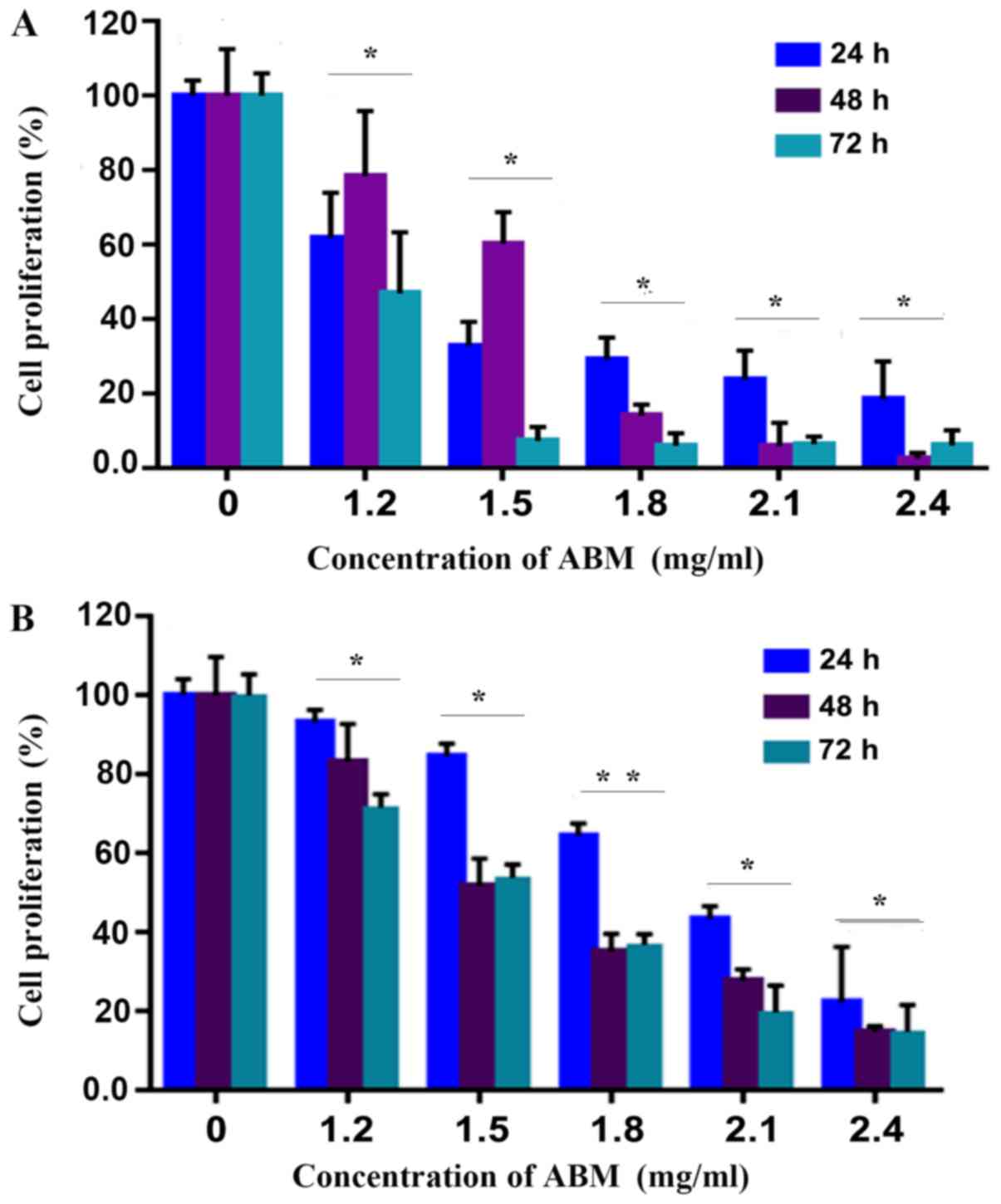
FA-2-b-β inhibits CML cell
proliferation
The CCK-8 assay was employed in order to determine
if FA-2-b-β was able to inhibit the cell proliferation of CML.
FA-2-b-β significantly inhibited proliferation of K562 cells or
primary CML BM cells (P<0.05), in a concentration dependent
manner (Fig. 1A and B). This effect
was enhanced in 1.2 mg/ml of FA-2-b-β over the entire culture
period.
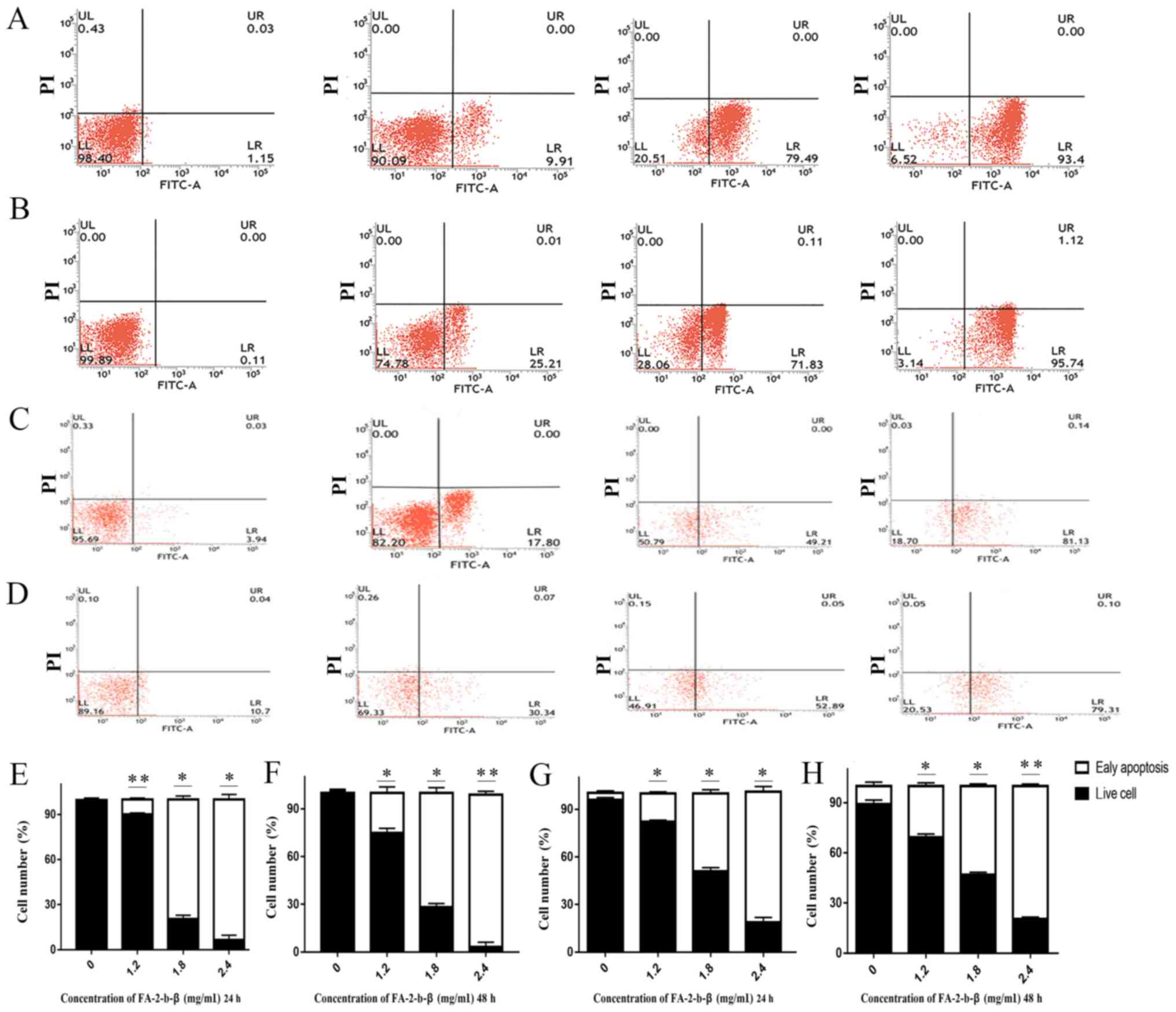
FA-2-b-β induces CML apoptosis
Some of the K562 or primary CML BM cells were
completely damaged or inhibited by FA-2-b-β at the dosage used
(Fig. 1A and B), but it was unclear
if other residual cells were still viable. The apoptosis of the
FA-2-b-β treated K562 cells was further determined using flow
cytometry. Annexin V-FITC/PI double staining assay was used in
order to determine the percentage of cells at different survival
status in the entire culture (Fig.
2A-H). The proportion of apoptotic cells increased following
treatment with FA-2-b-β. The total apoptotic K562 cells percentages
of the control group were 1.15 and 0.11% at 24 and 48 h,
respectively. This apoptotic rate was increased to 9.91, 79.49 and
93.40% (P<0.05; Fig. 2E), and
25.21, 71.83 and 95.74% (Fig. 2B)
following treatment with FA-2-b-β (at 1.2, 1.8 and 2.4 mg/ml,
respectively) for 24 and 48 h, respectively (Fig. 2E-F). The total apoptotic primary CML
BM cell percentages of the control group were 3.94 and 10.70% at 24
and 48 h mock treatment, respectively; whereas apoptosis in the
primary cells increased to 17.80 and 30.34%, 49.21 and 52.89%, and
81.13 and 79.31% following treatment with FA-2-b-β (at 1.2, 1.8 and
2.4 mg/ml, respectively) for 24 and 48 h, respectively (P<0.05;
Fig. 2G and H).
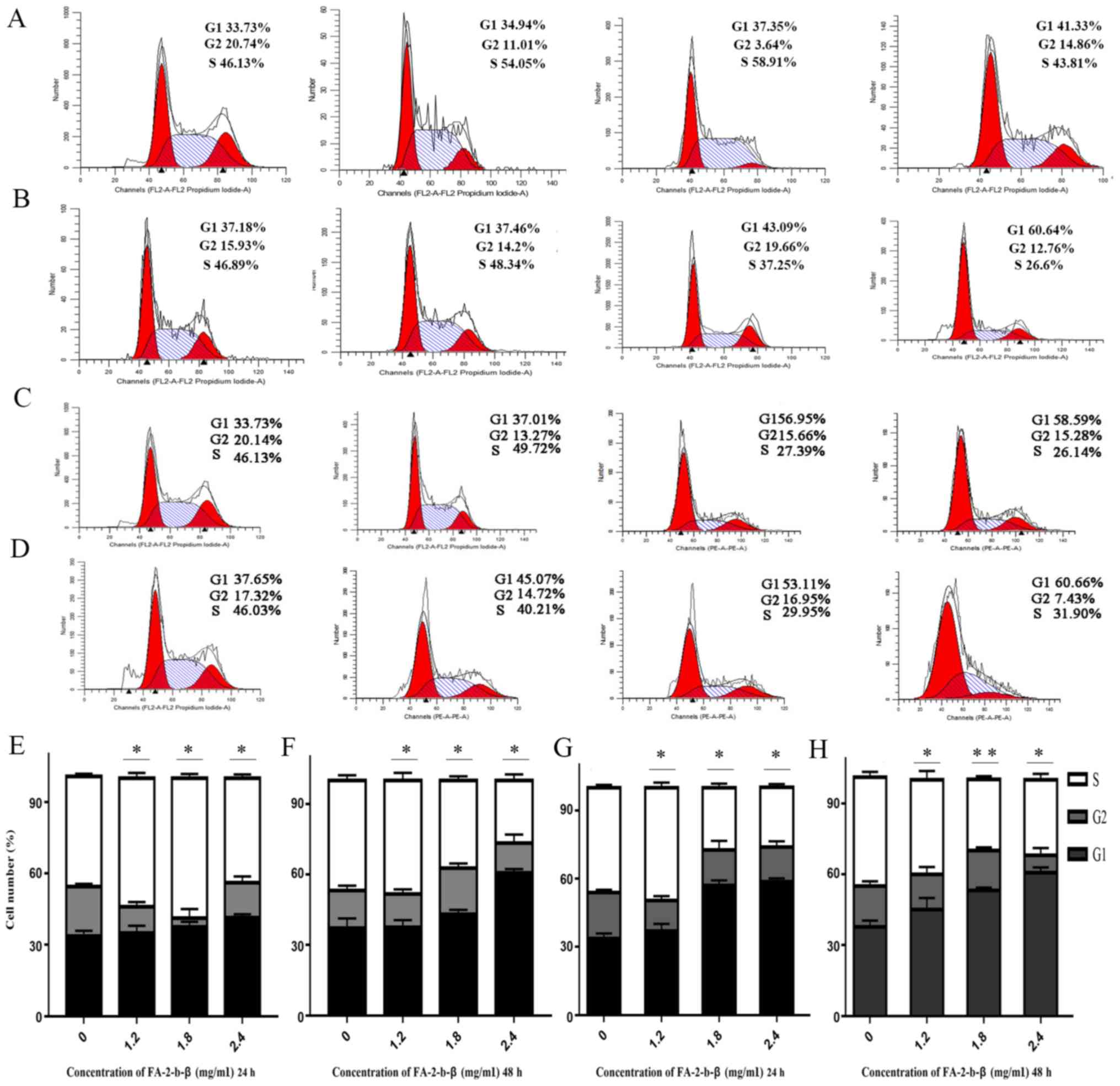
FA-2-b-β induced CML cells apoptosis
through inhibited G1 phase
Cell cycle distribution of the K562 cells or primary
CML BM cells was evaluated using flow cytometry, in order to
determine the mechanisms underlying the cytotoxic activity of
FA-2-b-β on the aforementioned cell types. The association between
the DNA content and the intensity of the PI was calculated
following treatment with FA-2-b-β at 0, 1.2, 1.8 and 2.4 mg/ml, for
24 and 48 h (Fig. 3A-H). There was
an increase in the G1 phase, but a partial decrease in
the S phase in a dose-dependent manner. The proportion of K562
cells at G1 or S phases was 33.73 and 33.73%, and 46.13
and 46.13%, respectively, with mock treatment for 24 and 48 h,
respectively. The proportion of cells in G1 phase
increased to 34.94, 37.35 and 41.33% using 1.2, 1.8 and 2.4 mg/ml
of FA-2-b-β for 24 h (Fig. 3A), and
to 37.01, 56.95 and 58.59% using 1.2, 1.8 and 2.4 mg/ml of FA-2-b-β
for 48 h (Fig. 3C); whereas the
proportion of cells in S phase partially decreased to 54.05, 58.91
and 43.81% at 24 h (Fig. 3A), and
49.72, 27.39 and 26.14% at 48 h (Fig.
3C). In the primary CML BM cells, the proportion of cells in
G1 and S phases were 37.18 and 37.65%, and 46.89 and
46.03%, respectively, with mock treatment for 24 and 48 h,
respectively. The proportion of cells in G1 phase
increased to 37.46, 43.09 and 60.64% using 1.2, 1.8 and 2.4 mg/ml
of FA-2-b-β for 24 h (Fig. 3B), and
to 45.07, 53.11 and 60.66% using 1.2, 1.8 and 2.4 mg/ml of FA-2-b-β
for 48 h (Fig. 3D). The proportion
of cells in S phase decreased to 48.34, 37.25 and 26.6% at 24 h
(Fig. 3B), whereas the proportion of
cells in S phase partially decreased to 40.21, 29.95 and 31.90% at
48 h (Fig. 3D) in the presence of
1.2, 1.8 and 2.4 mg/ml FA-2-b-β, respectively (Fig. 3A-H).
Cyclin D1 is a key cell cycle regulator that is
essential for the G1 phase, whereby its expression is
closely associated with the development and prognosis of numerous
types of cancer. Following FA-2-b-β treatment, the expression of
cyclin D1 decreased in a concentration-dependent manner, confirming
that G1 phase arrest occurred after FA-2-b-β treatment
in K562 cells or primary CML BM cells (Fig. 4A and B or Fig. 5A and B). Thus, FA-2-b-β induced both
cell cycle arrest and apoptosis in K562 cells and primary CML BM
cells, demonstrating similar findings to those that occur in the
patients undergoing chemotherapy (27).
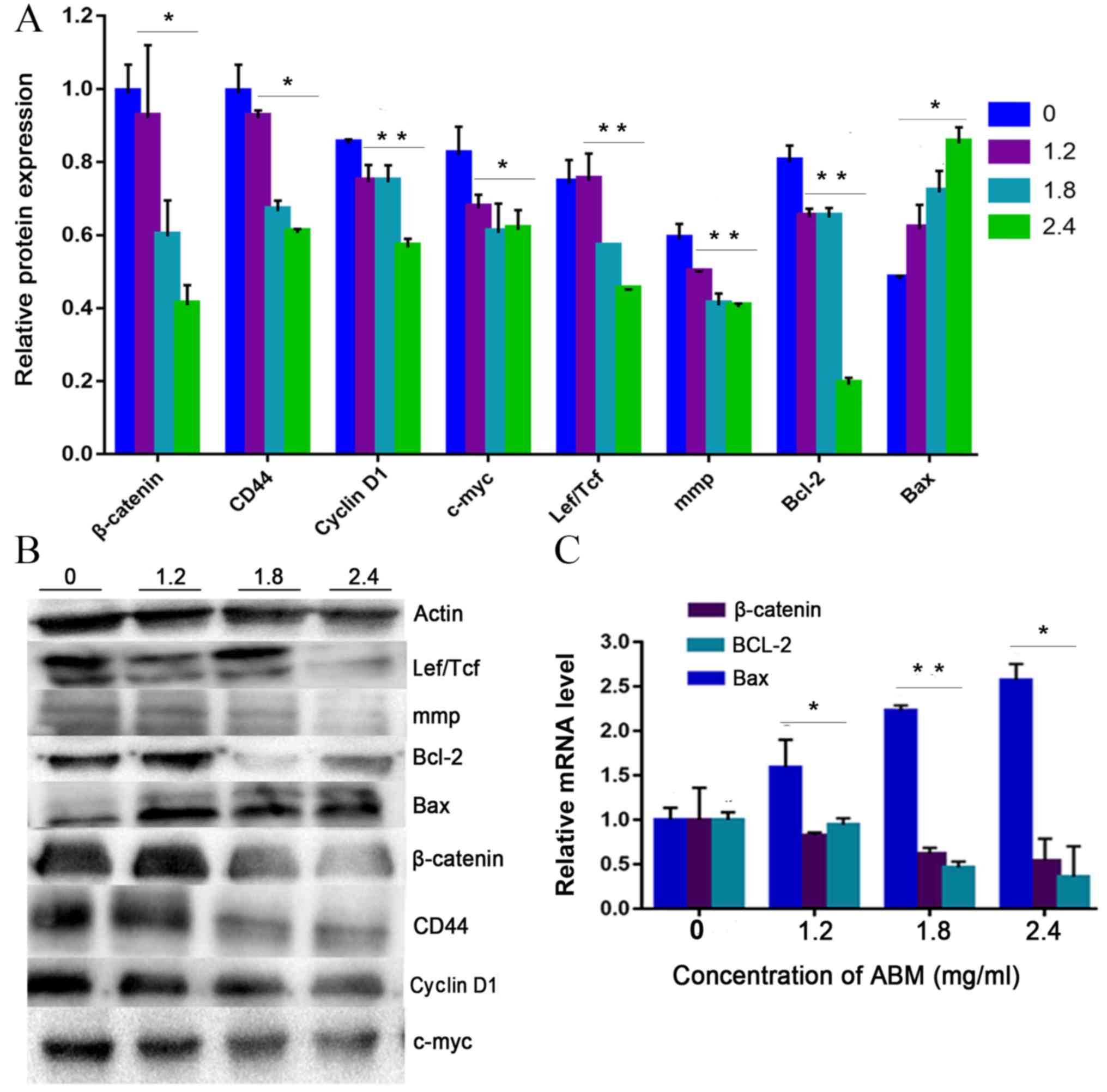
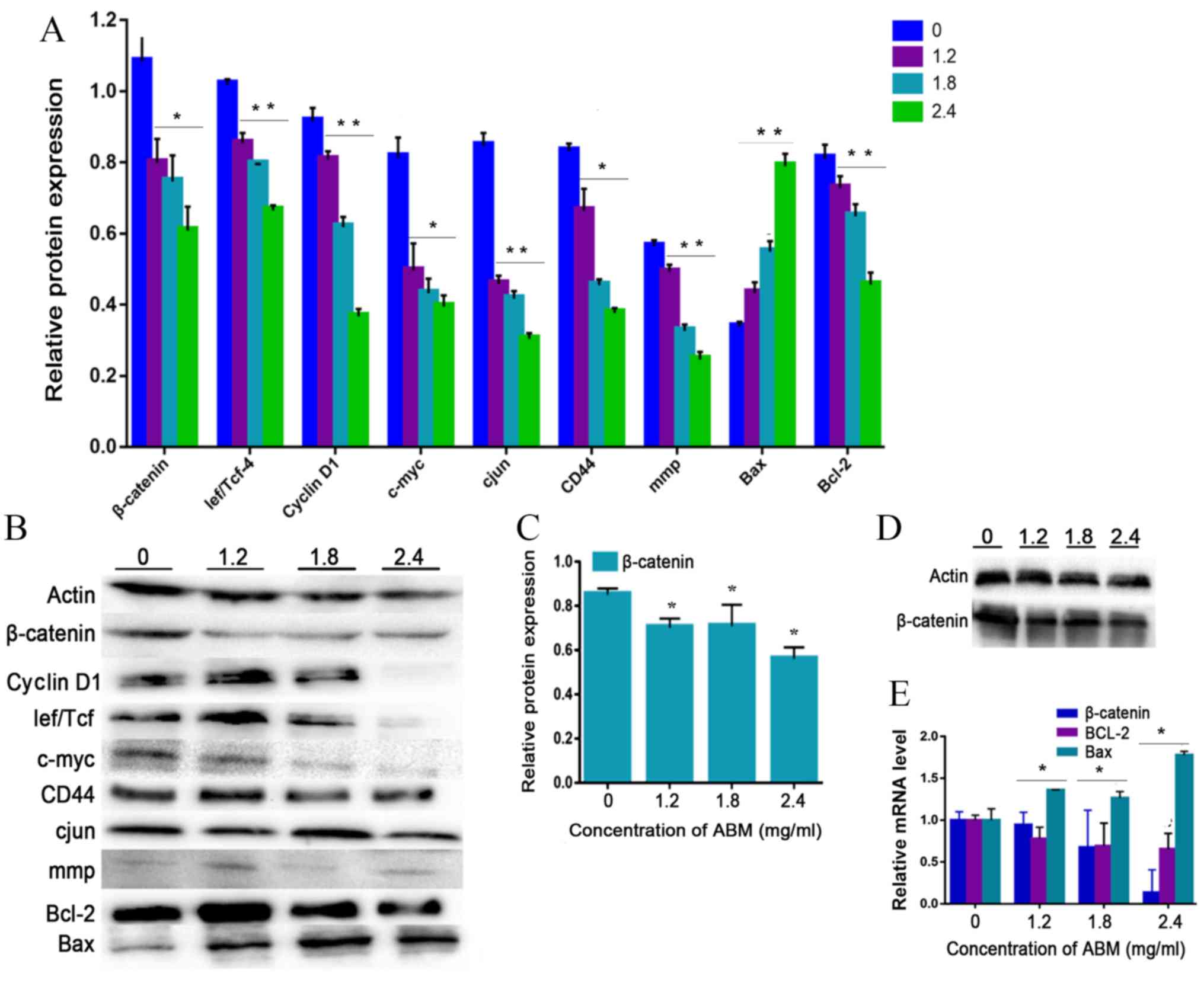 | Figure 4.AbM induces apoptosis of K562 cells.
K562 cells were treated with different concentrations of AbM (0,
1.2, 1.8 and 2.4 mg/ml) for 48 h and the expression of
apoptosis-associated genes and proteins was determined using
western blot analysis for (A and B) 24 h (*P<0.05 and
**P<0.001) or (C and D) 48 h and (E) reverse
transcription-quantitative PCR. The relative expression levels of β
catenin, Bax and Bcl-2 genes in K562 cells were quantified
according to the gene expression levels of actin. The relative
expression levels of cyclin D1, Lef/Tcf, c-myc, β catenin, CD44,
c-Jun, mmp, Bax and Bcl-2 proteins in K562 cells were quantified
according to the protein expression levels of actin. Data are
representative of three independent experiments and are expressed
as the mean ± SD. *P<0.05 vs. 0 mg/ml. |
 | Figure 5.AbM induces apoptosis of CML cells.
Primary CML BM cells were treated with different concentrations of
AbM (0, 1.2, 1.8 and 2.4 mg/ml) for 24 h and the expression of
apoptosis-related genes and proteins was determined using (A and B)
western-blot analysis (*P<0.05 and **P<0.001) and (C) reverse
transcription-quantitative PCR. The relative expression levels of β
catenin, Bax and Bcl-2 genes in primary CML BM cells were
quantified according to the expression levels of actin. The
relative expression levels of cyclin D1, Lef/Tcf, c-myc, β catenin,
CD44, c-Jun, mmp, Bax and Bcl-2 proteins in primary CML BM cells
were quantified according to the expression levels of Actin. Data
are representative of three independent experiments and are
expressed as the mean ± SD. *P<0.05, **P<0.001. |
The present study further tested the hypothesis that
FA-2-b-β may exert its effect on CML cells through negative
modulation of Wnt/β-catenin signaling. The K562 cells or primary
CML BM cells were dose dependently incubated with FA-2-b-β,
following which RT-qPCR and western blot analysis were performed to
assess the level of Wnt/β-catenin-related apoptosis gene and
protein expression at 24 h. The data are presented as the mean of
three independent experiments (P<0.05). β-catenin-associated
apoptosis protein and gene expression in K562 cells or primary CML
BM cells, exposed to FA-2-b-β at 1.2, 1.8 and 2.4 mg/ml for 24 h
(Fig. 4A, B and E, and Fig. 5) or 48 h (Fig. 4C and D), was determined. β-catenin
expression decreased at 24/48 h in a dose-dependent manner.
Comparison was performed in the protein extracts between the
drug-treated cells and controls in order to determine if there was
an influence of spontaneous fluctuations in the associated
apoptosis genes and proteins at each concentration. Actin was also
evaluated to allow normalized protein and gene loadings. Bcl-2 and
Bax protein and gene expression has been compared with the control
cells. Bcl-2 expression decreased whilst Bax expression increased,
in a dose-dependent manner. Based on our current study, it was
observed that FA-2-b-β decreases cyclin D1 protein expression for
binding to the promoter region and decreases β-catenin protein and
gene expression for nuclear localization. The present study
revealed ~50 or 25% of downregulated β-catenin mRNA in CML cells
treated with FA-2-b-β 1.8 or 2.4 mg/ml for 24 h (Figs. 4E and 5C). Thus, FA-2-b-β may stimulate β-catenin
via Lef/Tcf trans-inhibition activity, subsequently downregulating
cyclin D1 in K562 cells as well as in the primary cells (P<0.05;
Figs. 4A and 5A).
Discussion
Chemo-resistance remains a major challenge for
clinicians in cancer therapy. Anti-cancer therapy is usually
utilized to induce apoptosis or repair defects in cancer cells.
Chemo-resistance is partially due to cancer-induced mutations in
those genes responsible for apoptotic pathways (28). Thus, inhibition of cell proliferation
and/or promotion of cell death are strategies in the management of
cancer (29). It is necessary to
develop novel therapeutic strategies for the treatment of CML. A
significant advantage of drug repositioning over the conventional
drug discovery and development process is the rapid implementation
of anti-leukemia activity. As natural compounds derived from plants
are easily accessible, such materials are routinely used for
anti-cancer treatment in China, Japan, Brazil and the United States
(30).
AbM has been used for the treatment of numerous
types of diseases, including; Diabetes, atherosclerosis, hepatitis,
hypercholesterolemia and heart disease (31). Immune-enhancing effects have been
demonstrated to occur with AbM, including anti-oxidant,
anti-mutagenic and anti-tumor effects (32). The positive mechanism of action of
various medicinal plants in acute myeloid leukemia occurs via
polysaccharides, which is primarily associated with; inhibition of
cell proliferation, invasion and further induction of cancer cell
apoptosis (33). Apoptosis is a form
of programmed cell death without the accompanied inflammation, a
phenomenon that has been a popular topic in anti-cancer research
(34). The development of disrupted
apoptotic pathways has been demonstrated to be associated with
chemo-resistance of leukemia (35).
This is consistent with the results of the present study which
demonstrated that decreased viabilities of K562 cells or primary
CML BM cells occurs in response to treatment with FA-2-b-β.
Additionally, the present study observed that FA-2-b-β-treated K562
cells or primary CML BM cells presented morphological
characteristics of apoptosis that are associated with
downregulation of cyclin Dl and CD44. Cyclin Dl is a target of the
Wnt/β-catenin pathway, which is associated with a number of steps
of CML development (36).
Conversely, cyclin Dl and CD44 are believed to be key genes of the
Wnt/β-catenin pathway (36,37). Thus, the results of the present study
suggest that FA-2-b-β-induced apoptosis in CML cells utilizes the
Wnt/β-catenin pathway. β-catenin is another key mediator in cancer
progression, which is involved in the regulation of cell cycle
transition and cell apoptosis (38).
This is consistent with the results of a previous study which
demonstrated that AbM extracts induce apoptotic effects on leukemia
cells (39).
c-Jun, c-myc, cyclin D1, CD44 and mmp all contribute
to cancer cell proliferation and invasion (40). c-Jun activates β-catenin/Tcf
transcription activity via the Wnt/β-catenin pathway, which is
reported to be induced in CML (41).
Furthermore, β-catenin interacts directly with c-Jun to activate
the cyclin D1 and c-myc genes. Currently, a number of β-catenin
signaling pathway inhibitors are under investigation, with the aim
to disrupt β-catenin activity and its interaction with the
transcription factors (42). The
present study observed that the effect of FA-2-b-β for anti-cancer
activity is via targeting c-myc (Wnt genes). FA-2-b-β decreased
β-catenin and c-Jun production, particularly at the G1-S
transition which resulted in cell cycle arrest and cell apoptosis
at the G0/G1 phase. The results of the
present study are supported by a previous study (43), which demonstrated that accumulated
β-catenin activates the Wnt signaling pathway. Furthermore,
knockout of Wnt signaling decreases β-catenin via the degradation
of cytoplasmic β-catenin (44).
Kim et al (45), demonstrated that inhibition of
β-catenin reverses the transformed properties in cancer, suggesting
that β-catenin plays a key role in oncogenesis. These results may
shed light on the development of pharmacological therapeutic
targets against tumors. The results of the present study are
consistent with others in demonstrating that β-catenin is important
in the pathogenesis and progression of CML (46). β-catenin knockout decreases the risk
of development of CML (47).
Conversely, enhanced Wnt/β-catenin activity is associated with poor
clinical outcomes with chemotherapy during ‘blast crisis’ in CML
(48), and a meta-analysis revealed
that high-dose imatinib achieved a major molecular response at 12
months of therapy (49–51). The present study demonstrated that
FA-2-b-β inhibited the proliferation of K562 cells or primary CML
BM cells by inducing apoptosis and G1 arrest.
Furthermore, the present study revealed that FA-2-b-β suppressed
the expression of β-catenin as well as the protein products of Wnt
target genes; c-myc, c-Jun and cyclin D1 in the G1
transition. Subsequently, FA-2-b-β treatment contributed to cell
cycle arrest during the G1 phase and this observation is
supported by a previous study (52).
In the absence of Wnt signaling, β-catenin remains low through the
degradation of cytoplasmic β-catenin, which prevents induction of
the transcription of a number of proliferation-associated genes,
including; cyclin D1, c-myc and fibronectin. A decrease in
transcription, which would normally cause malignant transformation
(53), was detected following
downregulation of the protein, β-catenin after treatment with
FA-2-b-β. This ultimately inhibits the Wnt signaling pathway,
accompanied by decreased cyclin D1, c-myc and CD44 in K562 cells or
primary CML BM cells, in a dose-dependent manner. On the other
hand, β-catenin is also capable of decreasing cell apoptosis via
Bcl-2 or Bax targeting (36). Thus,
the results of the present study suggest that the anti-tumor
activity of AbM may be due to decreased cellular proliferation, but
also the induction of cell apoptosis (54). This is supported by other studies
that demonstrate that anti-cancer agents regulate Bcl-2 family
members (including Bax or Bcl-2) (55–57).
This evidence strongly supports the results of the present study,
which demonstrated that FA-2-b-β induced leukemia cell apoptosis
in vitro, accompanied by elevated Bax expression and
decreased level of Bcl-2. These results suggest that cyclin D1,
c-myc, c-Jun and CD44 can drive proliferation in CML as a
consequence of β-catenin activity. These data further confirm that
the role of β-catenin is indispensable for self-renewal of CML line
cells.
The present study presents a number of limitations.
First, it would provide more insight to perform the experiment
evaluating TK inhibitor-resistant cells and normal BM cells.
Secondly, future studies should include the experiment in
vivo in order to determine the direct effect of AbM in CML.
FA-2-b-β exhibits anti-cancer potential through
modulating different nodes of Wnt/β-catenin signaling pathways,
which subsequently affects apoptosis, as well as cell cycling.
Therefore, FA-2-b-β has potential for future treatment approaches
for CML. The mushroom extract has minimal toxicity in animal models
(58) and, to the best of our
knowledge is considered safe for human consumption. As Agaricus
blazei extract contains natural compounds derived from plants,
which can be obtained at relatively low costs for the patients, it
is routinely used in a number of clinics throughout China as an
anti-cancer therapy.
Acknowledgements
Not applicable.
Funding
The present study was funded by Gansu Provincial
Department of Science and Technology (grant no. GZK-2013-8) and the
Natural Science Foundation of China (grant no. 8156140621).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors contributions
MC and YS managed the design and coordination of the
present study. YS provided professional writing. YS, QZ and KY
carried out acid RNA protein complex (FA-2-b-β) extraction from
Agaricus blazei Murill. LD, ZM, SY and KB performed the in vitro
experiments. PY and XZ analyzed the results. All authors read and
approved the final published version of this article.
Ethics approval and consent to
participate
The present study was approved by the Human Ethics
Committee of Gansu Provincial Hospital (approval no. 2019-157). All
patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AbM
|
Agaricus blazei Murill
|
|
CML
|
chronic myeloid leukemia
|
|
BM
|
bone marrow
|
|
PB
|
peripheral blood
|
|
ABL1
|
abelson
|
|
BCR
|
breakpoint cluster region
|
|
TK
|
tyrosine kinase
|
References
|
1
|
Bamodu OA, Kuo KT, Yuanc LP, et al: HDAC
inhibitor suppresses proliferation and tumorigenicity of
drug-resistant chronic myeloid leukemia stem cells through
regulation of hsa-miR-196a targeting BCR/ABL1. Exp Cell Res.
370:519–530. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou H, Mak PY, Mu H, Mak DH, Zeng Z,
Cortes J, Liu Q, Andreeff M and Carter BZ: Combined inhibition of
β-catenin and Bcr-Abl synergistically targets tyrosine kinase
inhibitor-resistant blast crisis chronic myeloid leukemia blasts
and progenitors in vitro and in vivo. Leukemia. 31:2065–2074. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiao X, Liu P, Li D, Xia Z, Wang P, Zhang
X, Liu M, Liao L, Jiao B and Ren R: Combination therapy of
BCR-ABL-positive B cell acute lymphoblastic leukemia by tyrosine
kinase inhibitor dasatinib and c-JUN N-terminal kinase inhibition.
J Hematol Oncol. 13:802020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deshpande A and Buske C: Knocking the Wnt
out of the sails of leukemia stem cell development. Cell Stem Cell.
1:597–598. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zovko A, Yektaei-Karin E, Daniel S,
Nilsson A, Wallvik J and Stenke L: Montelukast, a cysteinyl
leukotriene receptor antagonist, inhibits the growth of chronic
myeloid leukemia cells through apoptosis. Oncol Rep. 40:902–908.
2018.PubMed/NCBI
|
|
6
|
Braun TP, Eide CA and Druker BJ: Response
and resistance to BCR-ABL1-targeted therapies. Cancer Cell.
37:530–542. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vetrie D, Helgason GV and Copland M: The
leukaemia stem cell: Similarities, differences and clinical
prospects in CML and AML. Nat Rev Cancer. 20:158–173. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumar R, Pereira RS, Zanetti C, et al:
Specific, targetable interactions with the microenvironment
influence imatinib-resistant chronic myeloid leukemia. Leukemia.
34:2087–2101. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cronan TA, Kaplan RM, Posner L, Blumberg E
and Kozin F: Prevalence of the use of unconventional remedies for
arthritis in a metropolitan community. Arthritis Rheum.
32:1604–1607. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
AlGhamdi KM, Khurrum H, Al-Natour SH, et
al: Use of complementary and alternative medicine among dermatology
outpatients: Results from a national survey. J Cutan Med Surg.
19:570–579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu L, Yu J, Chen R, Liu Y, Lou L, Wu Y,
Huang L, Fan Y, Gao P, Huang M, et al: Dual inhibition of Bcr-Abl
and Hsp90 by C086 potently inhibits the proliferation of
imatinib-resistant CML cells. Clin Cancer Res. 21:833–843. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ryan NM, Vertigan AE and Birring SS: An
update and systematic review on drug therapies for the treatment of
refractory chronic cough. Expert Opin Pharmacother. 19:687–711.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai WJ, Yang SC, Huang YL, Chen CC,
Chuang KA and Kuo YC: 4-Hydroxy-17-methylincisterol from Agaricus
blazei decreased cytokine production and cell proliferation in
human peripheral blood mononuclear cells via inhibition of NF-AT
and NF-κB activation. Evid Based Complement Alternat Med.
2013:4359162013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hetland G, Tangen JM, Mahmood F, et al:
Antitumor, anti-inflammatory and antiallergic effects of
Agaricus blazei mushroom Extract and the related medicinal
Basidiomycetes mushrooms, Hericium erinaceus and Grifola
frondosa: A review of preclinical and clinical studies.
Nutrients. 12:13392020. View Article : Google Scholar
|
|
15
|
Li Y, Sheng Y, Lu X, Guo X, Xu G, Han X,
An L and Du P: Isolation and purification of acidic polysaccharides
from Agaricus blazei Murill and evaluation of their lipid-lowering
mechanism. Int J Biol Macromol. 157:276–287. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun YQ, Guo TK, Xi YM, Chen C, Wang J and
Wang ZR: Effects of AZT and RNA-protein complex (FA-2-b-β)
extracted from Liang Jin mushroom on apoptosis of gastric cancer
cells. World J Gastroenterol. 13:4185–4191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin MH, Lee KM, Hsu CY, et al:
Immunopathological effects of Agaricus blazei Murill
polysaccharides against Schistosoma mansoni infection by Th1 and
NK1 cells differentiation. Int Immunopharmacol. 73:502–514. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ziliotto L, Pinheiro F, Barbisan LF and
Rodrigues MAM: Screening for in vitro and in vivo antitumor
activities of the mushroom Agaricus Blazei. Nutr Cancer.
61:245–250. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pak S, Park S, Kim Y, et al: Correction
to: The small molecule WNT/β-catenin inhibitor CWP232291 blocks the
growth of castration-resistant prostate cancer by activating the
endoplasmic reticulum stress pathway. J Exp Clin Cancer Res.
38:4402019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin HJ, Huang Y, Huang WL, Feng XL, Peng
Y, Zhang XJ, Jin Z and Fan XM: Regulatory mechanism of microRNA and
Wnt/β-catenin signaling pathway in tumorigenesis and metastasis.
Sci Sin. 45:5–14. 2018.
|
|
21
|
Li ZJ, Qiu LG, Li X, et al: Expression of
β-catenin gene in CML and its relationship with bcr/abl. J Exp
Hematol. 15:931–935. 2007.(In Chinese).
|
|
22
|
Hu Y, Yu K, Wang G, Zhang D, Shi C, Ding
Y, Hong D, Zhang D, He H, Sun L, et al: Lanatoside C inhibits cell
proliferation and induces apoptosis through attenuating
Wnt/β-catenin/c-Myc signaling pathway in human gastric cancer cell.
Biochem Pharmacol. 150:280–292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mizuno T, Inagaki R, Kanao T, Hagiwara T,
Nakamura T, Ito H, Shimura K, Sumiya T and Asakura A: Antitumor
activity and some properties of water-insoluble hetero-glycans from
‘Himematsutake’, the fruiting body of Agaricus blazei
Murill. Agr Biol Chem. 54:2897–2905. 1990. View Article : Google Scholar
|
|
24
|
Dong S, Furutani Y, Suto Y, Furutani M,
Zhu Y, Yoneyama M, Kato T, Itabe H, Nishikawa T, Tomimatsu H, et
al: Estrogen-like activity and dual roles in cell signaling of an
Agaricus blazei Murrill mycelia-dikaryon extract. Microbiol Res.
167:231–237. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He B, Wang Q, Liu X, et al: A novel HDAC
inhibitor chidamide combined with imatinib synergistically targets
tyrosine kinase inhibitor resistant chronic myeloid leukemia cells.
Biomed Pharmacother. 129:1103902020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tang K and Lightner DV: Detection and
quantification of infectious hypodermal and hematopoietic necrosis
virus in penaeid shrimp by real-time PCR. Dis Aquat Organ.
44:79–85. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen SH, Hsieh YY, Tzeng HE, et al: ABL
genomic editing sufficiently abolishes oncogenesis of human chronic
myeloid leukemia cells in vitro and in vivo. Cancers (Basel).
12:13992020. View Article : Google Scholar
|
|
28
|
Ryu H, Nam KY, Kim JS, Hwang SG, Song JY
and Ahn J: The small molecule AU14022 promotes colorectal cancer
cell death via p53-mediated G2/M-phase arrest and
mitochondria-mediated apoptosis. J Cell Physiol. 233:4666–4676.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun S, Zhang C, Gao J, Qin Q, Zhang Y, Zhu
H, Yang X, Yang D and Yan HT: Benzoquinone induces ROS-dependent
mitochondria-mediated apoptosis in HL-60 cells. Toxicol Ind Health.
34:270–281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Taofiq O, Rodrigues F, Barros L, et al:
Agaricus blazei Murrill from Brazil: an ingredient for
nutraceutical and cosmeceutical applications. Food Funct.
10:565–572. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang H, Fu Z and Han C: The medicinal
values of culinary-medicinal royal sun mushroom (Agaricus blazei
Murrill). Evid Based Complement Alternat Med. 2013:8426192013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu S, Li F, Jia S, et al: Drying effects
on the antioxidant properties of polysaccharides obtained from
Agaricus blazei Murrill. Carbohydr Polym. 103:414–417. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang F, Song X, Li L, Wang J, Lin L, Li
C, Li H, Lv Y, Jin Y, Liu Y, et al: Polygala tenuifolia
polysaccharide PTP induced apoptosis in ovarian cancer cells via a
mitochondrial pathway. Tumor Biol. 36:2913–2919. 2015. View Article : Google Scholar
|
|
34
|
Schultz DR and Harringto WJ: Apoptosis:
Programmed cell death at a molecular level. Semin Arthritis Rheum.
32:345–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mondal A, Banerjee D, Majumder R, Maity TK
and Khowala S: Evaluation of in vitro antioxidant, anticancer and
in vivo antitumour activity of Termitomyces clypeatus MTCC 5091.
Pharm Biol. 54:2536–2546. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hou YC, Chao YJ, Tung HL, Wang HC and Shan
YS: Coexpression of CD44-positive/CD133-positive cancer stem cells
and CD204-positive tumor-associated macrophages is a predictor of
survival in pancreatic ductal adenocarcinoma. Cancer.
120:2766–2777. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu Q, Yang Y, Wu S, et al: Evaluation of
the correlation of KAI1/CD82, CD44, MMP7 and β-catenin in the
prediction of prognosis and metastasis in colorectal carcinoma.
Diagn Pathol. 10:1762015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao H, Ashihara E and Maekawa T: Targeting
the Wnt/β-catenin signaling pathway in human cancers. Expert Opin
Ther Targets. 15:873–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Collin-Hansen C, Pedersen SA, Andersen RA
and Steinnes E: First report of phytochelatins in a mushroom:
induction of phytochelatins by metal exposure in Boletus edulis.
Mycologia. 99:161–174. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Saadeddin A, Babaei-Jadidi R, Spencer-Dene
B and Nateri A: The links between Transcription, -catenin/JNK
signaling, and carcinogenesis. Mol Cancer Res. 7:1189–1196. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gan X, Wang J, Xi Y, Wu Z, Li Y and Li L:
Nuclear Dvl, c-Jun, beta-catenin, and TCF form a complex leading to
stabilization of beta-catenin-TCF interaction. J Cell Biol.
180:1087–1100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lepourcelet M, Chen YNP, France DS, et al:
Small-molecule antagonists of the oncogenic Tcf/beta-catenin
protein complex. Cancer Cell. 5:91–102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Silverberg E and Lubera J: Cancer
statistics, 1987. CA Cancer J Clin. 37:2–19. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chairoungdua A, Smith DL, Pochard P, Hull
M and Caplan MJ: Exosome release of β-catenin: A novel mechanism
that antagonizes Wnt signaling. J Cell Biol. 190:1079–1091. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim JS, Crooks H, Foxworth A and Waldman
T: Proof-of-principle: Oncogenic β-catenin is a valid molecular
target for the development of pharmacological inhibitors 1. Mol
Cancer Ther. 1:1355–1359. 2002.PubMed/NCBI
|
|
46
|
Liu N, Zang S, Liu Y, et al: FZD7
regulates BMSCs-mediated protection of CML cells. Oncotarget.
7:6175–6187. 2015. View Article : Google Scholar
|
|
47
|
Iwasaki H and Akashi K: Identification and
biology of CML stem cells. Springer; Japan: 2016,
doi:10.1007/978-4-431-55714-2_1. View Article : Google Scholar
|
|
48
|
Sun J, Li B, Jia Z, Zhang A, Wang G, Chen
Z, Shang Z, Zhang C, Jian C and Yang W: RUNX3 inhibits glioma
survival and invasion via suppression of the β-catenin/TCF-4
signaling pathway. J Neurooncol. 140:15–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hoffmann VS, Hasford J, Deininger M,
Cortes J, Baccarani M and Hehlmann R: Systematic review and
meta-analysis of standard-dose imatinib vs. high-dose imatinib and
second generation tyrosine kinase inhibitors for chronic myeloid
leukemia. J Cancer Res Clin Oncol. 143:1311–1318. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ge L, Tian JH, Li YN, Pan JX, Li G, Wei D,
Xing X, Pan B, Chen YL, Song FJ, et al: Association between
prospective registration and overall reporting and methodological
quality of systematic reviews: A meta-epidemiological study. J Clin
Epidemiol. 93:45–55. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tian J, Zhang J, Ge L, Yang K and Song F:
The methodological and reporting quality of systematic reviews from
China and the USA are similar. J Clin Epidemiol. 85:50–58. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sun J, Li B, Jia Z, Zhang A, Wang G, Chen
Z, Shang Z, Zhang C, Cui J and Yang W: RUNX3 inhibits glioma
survival and invasion via suppression of the β-catenin/TCF-4
signaling pathway. J Neurooncol. 140:15–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu ZL, Hu J, Huang ZL, Li H, Liu Y and
Feng WL: Influence of AKT inhibitor on Wnt/β-catenin pathway in
chronic myeloid leukemia K562 cells. Medical Journal of Chinese
People's Liberation Army. 40:710–715. 2015.
|
|
54
|
Zhang L, Xu J, Zhang X, Zhang Y, Wang L,
Huang X and Xu Z: The role of tumoral FOXP3 on cell proliferation,
migration, and invasion in gastric cancer. Cell Physiol Biochem.
42:1739–1754. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Srivastava R, Cao Z, Nedeva C, et al:
BCL-2 family protein BOK is a positive regulator of uridine
metabolism in mammals. Proc Natl Acad Sci USA. 116:15469–15474.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tajadura V, Hansen MH, Smith J, et al:
β-catenin promotes endothelial survival by regulating eNOS activity
and flow-dependent anti-apoptotic gene expression. Cell Death Dis.
11:4932020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liang J, Wang W, Wei L, Gao S and Wang Y:
Oridonin inhibits growth and induces apoptosis of human neurocytoma
cells via the Wnt/β-catenin pathway. Oncol Lett. 16:3333–3340.
2018.PubMed/NCBI
|
|
58
|
Sumiya T, Ikeda Y, Broadmeadow A, May K,
Pritchard L, Horne C and Burlinson B: Himematsutake (Iwade Strain
101) extract (ABM-FD): Genetic toxicology and a 3-month dietary
toxicity study in rats. Food Chem Toxicol. 46:1949–1959. 2008.
View Article : Google Scholar : PubMed/NCBI
|