Introduction
Lung cancer is a leading cause of cancer-associated
mortality. There are >2 million newly diagnosed lung cancer
cases and 1.8 million mortalities from lung cancer each year
worldwide (1). Non-small cell lung
cancer (NSCLC) accounts for 85% of all lung cancer cases (2). Lung cancer treatments include surgery,
radiotherapy, chemotherapy, targeted therapies and immunotherapies
(3). However, despite the
technological advances in recent years, the 5-year survival rate of
patients with NSCLC remains low (14%) (4). Poor responsiveness to therapies for
NSCLC is associated with complex underlying mechanisms (5). Thus, understanding the molecular
mechanism of NSCLC invasion and metastasis is important for
identifying novel therapeutic targets and prognostic biomarkers to
improve treatments of patients with NSCLC.
Leukemia inhibitory factor (LIF) is a
multifunctional cytokine that belongs to the interleukin-6 family
of cytokines; it is expressed in numerous types of tissues and
cells, such as embryonic stem cells and monocytes (6). Binding of LIF to its receptor activates
critical signaling pathways that regulate cell proliferation,
survival and differentiation. These pathways include the Janus
tyrosine kinase (JAK)/signal transducer and activator of
transcription 3 (STAT3), extracellular signal-regulated protein
kinase (ERK) and phosphoinositide 3-kinase (PI3K) signaling
pathways (7–9). LIF can play opposing roles in different
types of cancer. For example, LIF can promote tumor growth in
rhabdomyosarcoma (10),
nasopharyngeal carcinoma (11),
colorectal cancer (12) and oral
squamous cell carcinoma (13),
whereas it acts as a tumor suppressor in breast cancer (14,15),
melanoma (16) and hepatocellular
carcinoma (17). However, the
expression and role of LIF in NSCLC are largely unknown; thus,
further studies are needed.
The present study determined the expression of LIF
and the activation of its downstream signaling molecule STAT3 in
tissues derived from patients with NSCLC. The association between
LIF expression and the clinicopathological features of patients
with NSCLC was analyzed to evaluate the role of LIF in NSCLC
development, the effects of LIF treatment on cell proliferation,
migration and invasiveness were measured in a NSCLC-derived cell
line.
Materials and methods
Tissue specimen collection
Paraffin-embedded tissues from 105 patients with
adenocarcinoma or squamous cell carcinoma, as confirmed by
pathological biopsy at People's Hospital Affiliated to Ningbo
University (Ningbo, China) were collected in the present study and
underwent immunohistochemical analysis. Patient ages ranged from 38
to 80 years old (mean age, 60.79 years). Informed consent was
provided by all patients based on the established protocol approved
by the Ethics Committee of Yinzhou People's Hospital.
Specimens of tumor, adjacent and normal tissues from
33 patients with NSCLC were collected from January 2018 to January
2019 at Yinzhou People's Hospital. Tissue 2 cm away from the tumor
was considered adjacent, while tissue >5 cm away from the tumor
was considered normal tissue. The specimens were flash frozen in
liquid nitrogen immediately after surgical resection and stored at
−80°C. All specimens were pathologically confirmed as one of two
subtypes of NSCLC: Adenocarcinoma or squamous cell carcinoma.
Frozen specimens were used to detect LIF mRNA and protein
expression levels, as well as the expression levels of
phosphorylated (p)-STAT3.
Immunohistochemistry (IHC)
IHC for LIF and p-STAT3 was performed as previously
described (18). Briefly,
paraffin-embedded tissue sections were baked at 62°C for 30 min,
deparaffinized in xylene (Sinopharm Chemical Reagent Co., Ltd.) and
rehydrated in ethanol prior to pretreatment with 3% hydrogen
peroxide/methanol solution (Sinopharm Chemical Reagent Co., Ltd.)
for 15 min to block endogenous peroxidase activity. After blocking
with 10% normal goat serum (cat. no. ST023; Beyotime Institute of
Biotechnology, Inc.) for 30 min at room temperature, the tissue
sections were incubated with anti-LIF (1:400 dilution; NBP2-27406;
Novus Biologicals, Ltd.) and anti-p-STAT3 (Tyr705; 1:400 dilution;
CST9145; Cell Signaling Technology, Inc.) primary antibodies
overnight at 4°C. The tissue sections were then treated with a
biotinylated goat anti-rabbit (1:400 dilution; cat. no. BA1000;
Vector Laboratories, Inc.) or goat anti-rat (1:400 dilution; cat.
no. BA9400; Vector Laboratories, Inc.) secondary antibodies at 37°C
for 30 min. Immunoreactivity was visualized with a VECTASTAIN Elite
ABC kit (Vector Laboratories, Inc.). The results of IHC were
analyzed by two pathologists independently.
For evaluation of LIF expression, two pathologists
randomly selected five areas of each slice under a microscope (Axio
Lab.A1; Zeiss AG) with a 10X objective according to the
distribution and intensity of LIF staining (0 for no expression, 1
for weak expression, 2 for moderate expression and 3 for high
expression). The mean value of the five areas was used as the final
score for each slice.
The expression of p-STAT3 was analyzed using ImageJ
software (v.1.8.0.112; National Institutes of Health). Five areas
of each slice were selected under a microscope (Axio Lab A1; Zeiss
AG) with a 10X objective, and cells with positive staining in the
nuclei were counted. The score was determined according to the
percentage of positive cells (0 for no staining, 1 for <10%
staining, 2 for 11–50% staining and 3 for >50% staining) and
averaged. A mean value of 0–0.4 was considered as non-expression,
0.5–1.4 as weak expression, 1.5–2.4 as moderate expression group
and 2.5–3 as high expression (13,19).
Western blot analysis
A standard western blot technique was used to detect
protein expression in the tumor, adjacent and normal lung tissues,
as well as in A549 cells. The tissues and A549 cells were collected
in ice-cold RIPA lysis buffer (cat. no. P0013B; Beyotime Institute
of Biotechnology, Inc.) containing protease inhibitor and
phosphatase inhibitors (cat. no. A32961; Thermo Fisher Scientific,
Inc.), and then centrifuged at 13,000 × g for 30 min at 4°C. The
concentration of homogenate protein was determined using a BCA
protein assay kit (cat. no. P0012; Beyotime Institute of
Biotechnology, Inc.). Samples (30 µg) were loaded on 10% gels for
SDS-PAGE and separated proteins were transferred to polyvinylidene
fluoride membranes (0.45 µm). The membranes were blocked with 5%
non-fat milk (cat. no. LP0031; Solarbio Life Sciences, Inc.) in
TBST (1% Tween 20) at room temperature for 1 h, and incubated with
overnight with primary antibodies against anti-LIF (1:2,000
dilution; cat. no. NBP2-27406; Novus Biologicals, Ltd.),
anti-p-STAT3 (Tyr705; 1:2,000 dilution; cat. no. CST9145; Cell
Signaling Technology, Inc.), anti-STAT3 (1:2,000 dilution; cat. no.
C-20; Santa Cruz Biotechnology, Inc.) and anti-β-actin (1:2,000
dilution; cat. no. A5441; Sigma-Aldrich; Merck KGaA) at 4°C.
Membranes were washed three times in TBST with 1% Tween-20 and then
incubated with secondary goat anti-rabbit (1:2,000 dilution; cat.
no. ab97051; Abcam, Inc.) or goat anti-rat (1:2,000 dilution; cat.
no. ab97057; Abcam, Inc.) antibodies at room temperature for 1 h.
Blots were developed using enhanced chemiluminescence according to
the manufacturer's instruction (cat. no. 1705060; Bio-Rad, Inc.).
The intensity of each band was analyzed using Image J software
(v.1.8.0.112; National Institutes of Health). β-actin was used as a
loading control.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from the tissues was purified using the
RNeasy Kit (Qiagen, Inc.) according to the manufacturer's protocol,
and it was reverse transcribed into cDNA using the reverse
transcriptase and random hexamers found in the TaqMan™ Reverse
Transcription Reagents kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol (10 min
at 25°C, 120 min at 37°C, 5 min at 85°C and 4°C thereafter). Human
LIF and β-actin cDNA levels were then quantified using the
StepOnePlus™ Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The TaqMan™ probes of LIF and β-actin
were purchased from Applied Biosystems (Thermo Fisher Scientific,
Inc; cat. no. for kit, 4351370). Each sample was measured in
triplicate by RT-qPCR using the TaqMan™ PCR Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.; cat. no. 4369016). The
thermocycling conditions for qPCR were as follows: 95°C for 10 min
(enzyme activation), followed by 40 cycles consisting of 95°C for
15 sec (denaturing) and 60°C for 1 min (annealing/extension). The
expression level of the LIF gene was normalized to that of the
internal reference gene β-actin (20). Relative gene expression was
calculated using the 2−∆∆Cq method. Primer sequences
were as follows: LIF forward, 5′-CCAACGTGACGGACTTCCC-3′ and
reverse, 5′-TACACGACTATGCGGTACAGC-3′; and β-actin forward,
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′ and reverse,
5′-TGGCACCCAGCACAATGAA-3′.
Cell line and culture
The A549 cell line was purchased from the American
Type Culture Collection. Cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), penicillin (100 IU/ml) and streptomycin sulfate
(100 µg/ml), and incubated at 37°C in a humidified incubator with
5% CO2. Culture medium was replaced every 2–3 days and
cells were harvested following treatment with 0.05% trypsin-EDTA
solution (Gibco Thermo Fisher Scientific, Inc.) when 80–90%
confluence was reached.
Cell proliferation and invasion
assays
Cell proliferation was determined by Cell Counting
Kit-8 (CCK-8; cat. no. HY-K0301; MedChemExpress, Inc.) assay
following the manufacturer's protocol. A total of 1,000 cells in a
volume of 100 µl per well in a 96-well plate, in medium containing
10% FBS, were incubated for 24 h at 37°C and with 5%
CO2. Next, the different concentrations of LIF (100, 200
and 400 ng/ml) (cat. no. 7734-LF-02; R&D Systems, Inc.) were
added on the following day and incubated for 48 h. Untreated A549
cells were used as the control. At the end of incubation, the CCK-8
reagent (10 µl) was added to 90 µl DMEM to generate a working
solution, and 100 µl per well was used for incubation for 2 h, with
absorbance measured at 450 nm. Cell invasion assays were performed
in 24-well Matrigel-coated Transwell invasion chambers with 8-µm
membrane pores (cat. no. 354480; Corning, Inc.). For the cell
invasion assays, a cell suspension (1×105 cells) in
serum-free medium with different concentrations of LIF (100, 200
and 400 ng/ml) was applied to the upper chamber. Untreated A549
cells were used as the control. The lower chamber contained DMEM
supplemented with 30% FBS. After 24 h of incubation, the invaded
cells on the underside of the invasion chamber were stained with
0.5% crystal violet (cat. no. E607309; Shenggong, Inc.). The
stained cells were counted under a microscope (TE2000; Nikon
Corporation) and imaged from 10 different view fields for each
membrane. This value represented the number of migrated cells
through the membrane. Data were presented as the mean ± SD of
triplicate assays for each experimental condition.
Wound healing assay
A549 cells grown to 90% confluency in DMEM with 10%
FBS were placed in medium containing 2% FBS and incubated overnight
at 37°C. The cell layer was then scratched with a 200-µl pipette
tip and rinsed twice with PBS to remove floating cells and debris.
The wound gap created by the scratch was measured with a microscope
(TE2000; Nikon Corporation) immediately after the scratch and at 48
h after incubation with different concentrations of LIF (100, 200
and 400 ng/ml) (7734-LF-02; R&D Systems, Inc.) in DMEM with 2%
FBS. Untreated A549 cells were used as the control. In the STAT3
blocking study, Stattic, a specific STAT3 inhibitor, was employed
to block STAT3. A total of 200 ng/ml LIF and 20 µM Stattic
(Sigma-Aldrich; Merck KGaA) were added to the A549 cells for
incubation in DMEM with 2% FBS for 48 h. All experiments were
repeated three times.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 7 (GraphPad Software). Data are expressed as the mean ±
standard deviation (SD). All expriments were independently repeated
at last three times. An unpaired, two-tailed Student's t-test was
used to statistically compare LIF and p-STAT3 expression between
the adenocarcinoma and squamous carcinoma tissues. One-way ANOVA
and Tukey's post hoc test were used for statistical comparisons in
in vitro experiments, and repeated measures ANOVA with
Bonferroni correction was used for the protein and mRNA expression
comparisons among tumor, adjacent and normal samples. The
correlation between the LIF and p-STAT3 IHC scores was evaluated by
Pearson's correlation analysis. The association between LIF
expression and the clinicopathological parameters of patients with
NSCLC was analyzed using the χ2 test, except for lymph
node metastasis, which was analyzed using Fisher's exact test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
LIF is overexpressed in NSCLC,
specifically in the adenocarcinoma subtype
The mRNA level of LIF in NSCLC was measured. Using
qPCR, the LIF mRNA levels in the tumor tissues were compared to
those in the corresponding adjacent tissue and normal lung samples.
As shown in Fig. 1A, the LIF mRNA
level was significantly higher in the tumor tissues compared with
that in the adjacent and normal tissues, and as expected, the LIF
mRNA level in the adjacent tissue was higher compared with that in
the normal tissue. LIF levels were also determined in NSCLS
subtypes. The LIF mRNA level was significantly higher in
adenocarcinoma tissues compared with that in squamous cell
carcinoma tissues (Fig. 1B). Next,
LIF protein expression in tumor, corresponding adjacent, and normal
lung tissues were determined using western blot analysis, and
corresponding protein bands derived from the same membrane. It was
found that LIF expression was significantly higher in tumor tissues
compared with that in the corresponding adjacent and normal
tissues, which was consistent with the mRNA levels (Fig. 1C and D). Semi-quantitative analysis
of LIF IHC staining revealed that LIF expression in NSCLC tumor
tissues was significantly higher than that in the corresponding
adjacent and normal tissues, and LIF staining score in adjacent
tissues was also significantly higher than that in normal tissues
(Fig. 1E and F). In addition, LIF
protein expression was significantly higher in adenocarcinoma
tissues compared with that in squamous cell carcinoma tissues
(Fig. 1G). Thus, the present data
indicate that LIF expression is elevated in NSCLC, particularly in
the adenocarcinoma subtype.
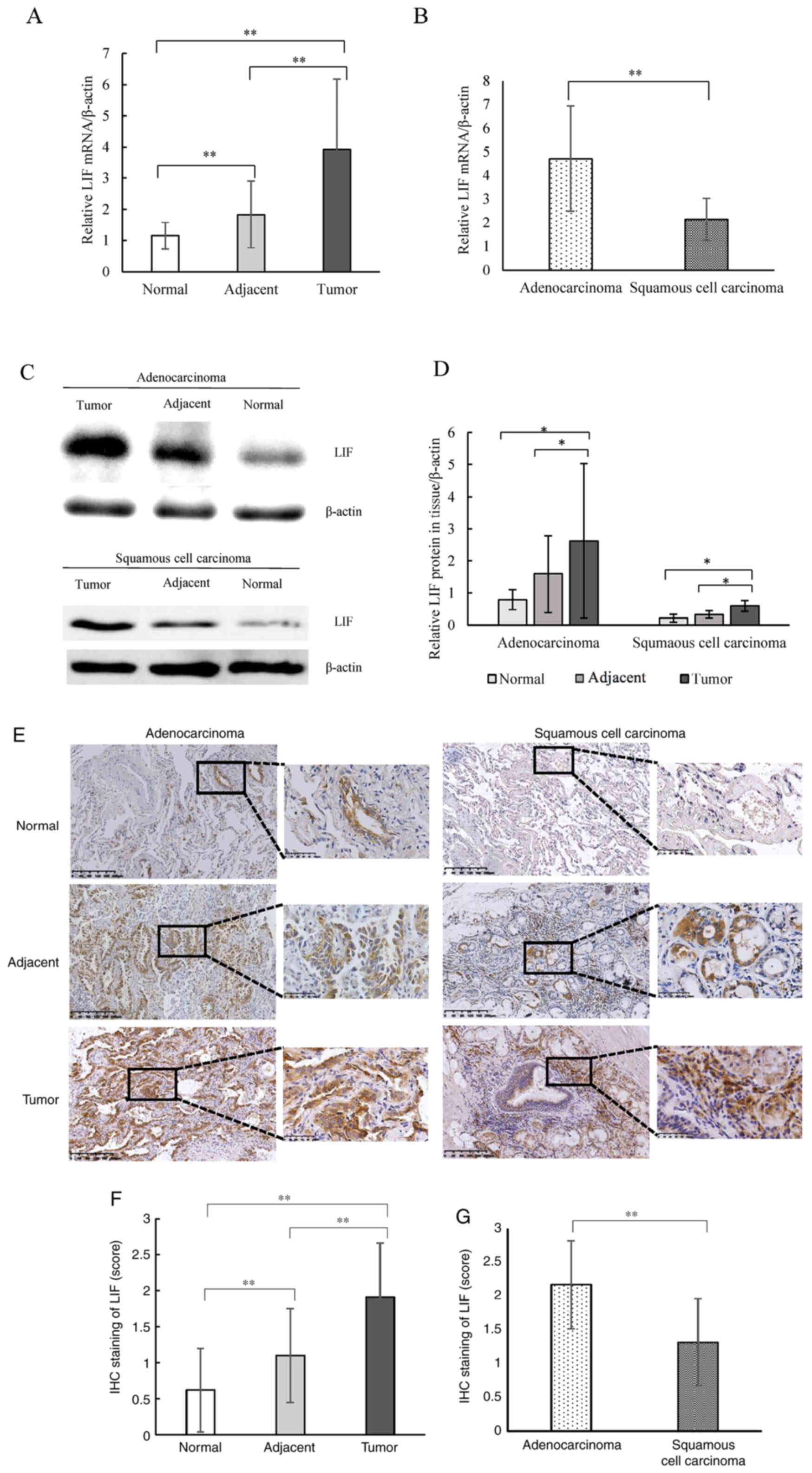 | Figure 1.LIF expression in tissues derived
from patients with NSCLC. (A) Relative mRNA expression of
LIF/β-actin in NSCLC tumor, adjacent and normal tissues. (B)
Relative mRNA expression of LIF/β-actin in adenocarcinoma and
squamous cell carcinoma subtypes. (C) Western blot analysis of LIF
expression in NSCLC tumor, adjacent and normal tissues. (D)
Quantitative analysis of the western blot results for relative
protein expression of LIF/β-actin in tumor, adjacent and normal
tissues in adenocarcinoma and squamous cell carcinoma subtypes. (E)
IHC staining of LIF in tumor, adjacent and normal tissues of
adenocarcinoma and squamous cell carcinoma subtypes (scale bar, 200
and 50 µm). (F) Quantitative analysis of IHC staining scores of LIF
in tumor, adjacent and normal tissues. (G) Quantitative analysis of
IHC staining scores of LIF in adenocarcinoma and squamous cell
carcinoma subtypes. *P<0.05, **P<0.01. NSCLC, non-small cell
lung cancer; LIF, leukemia inhibitory factor; IHC,
immunohistochemistry. |
LIF/STAT3 signaling is involved in the
progression of NSCLC
Overexpression and activation of STAT3 are
associated with the malignant behavior of carcinomas. To further
explore the mechanism of LIF in NSCLC progression, western blot
analysis was used to determine the activation of STAT3 in tissues
with different LIF expression levels. It was found that the
expression of p-STAT3 relative to that of total STAT3 (T-STAT3) in
the tumor tissues from both adenocarcinoma and squamous cell
carcinoma subtypes was significantly higher compared with that of
the corresponding adjacent and normal tissues (Fig. 2A and B). IHC staining also showed
that p-STAT3 expression was higher in the tumor tissues compared
with that in the matched adjacent and normal tissues (Fig. 2C). Semi-quantitative analysis
revealed that p-STAT3 staining in tumor tissues of NSCLC was
significantly higher compared with that in the corresponding normal
tissues and adjacent tissues (Fig.
2D). p-STAT3 expression in adenocarcinoma tissues was higher
than that in squamous cell carcinoma tissues (Fig. 2E). Correlation analysis showed that
the IHC scores for p-STAT3 expression were positively correlated
with LIF expression (r=0.346; P<0.01) (Fig. 2F). These results indicate that STAT3
is activated in NSCLC, particularly in adenocarcinoma tissues, and
its activation is associated with LIF expression.
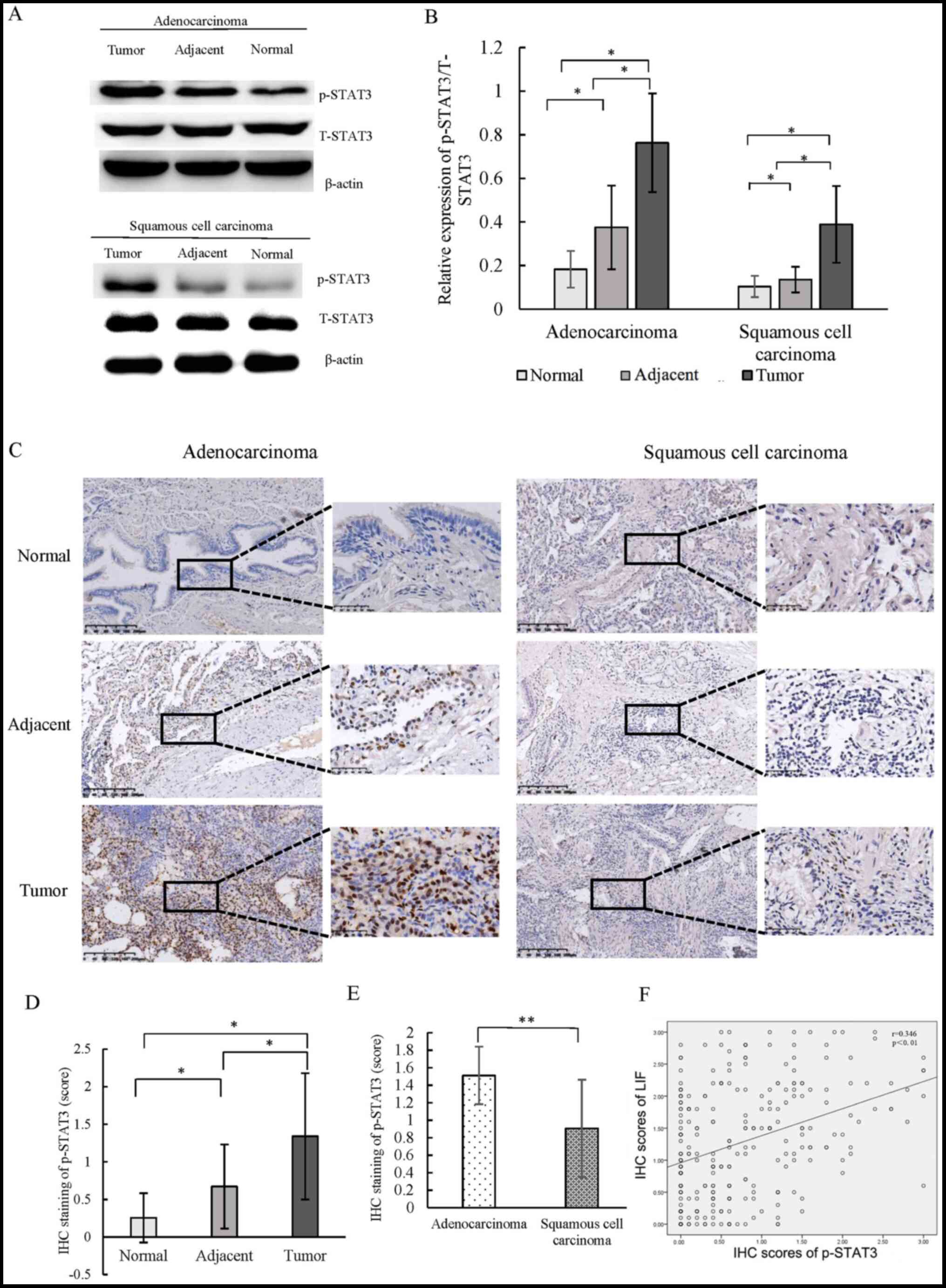 | Figure 2.Expression of p-STAT3 expression in
tissues derived from patients with NSCLC. (A) Western blot analysis
of p-STAT3 and T-STAT3 expression in two subtypes of NSCLC. (B)
Quantitative analysis of the p-STAT3/T-STAT3 expression ratio in
tumor, adjacent and normal tissues of adenocarcinoma and squamous
cell carcinoma subtypes. (C) IHC staining of p-STAT3 in tumor,
adjacent and normal tissues of adenocarcinoma and squamous cell
carcinoma subtypes (scale bar, 200 and 50 µm); (D) Quantitative
analysis of IHC staining scores of p-STAT3 in tumor, adjacent and
normal tissues. (E) Quantitative analysis of IHC staining scores of
p-STAT3 in adenocarcinoma and squamous cell carcinoma subtypes. (F)
Correlation analysis for IHC scores of p-STAT3 and LIF expression
in tumor, adjacent and normal tissues. *P<0.05, **P<0.01.
STAT3, signal transducer and activator of transcription 3; p-,
phosphorylated; T-, total; NSCLC, non-small cell lung cancer; IHC,
immunohistochemistry. |
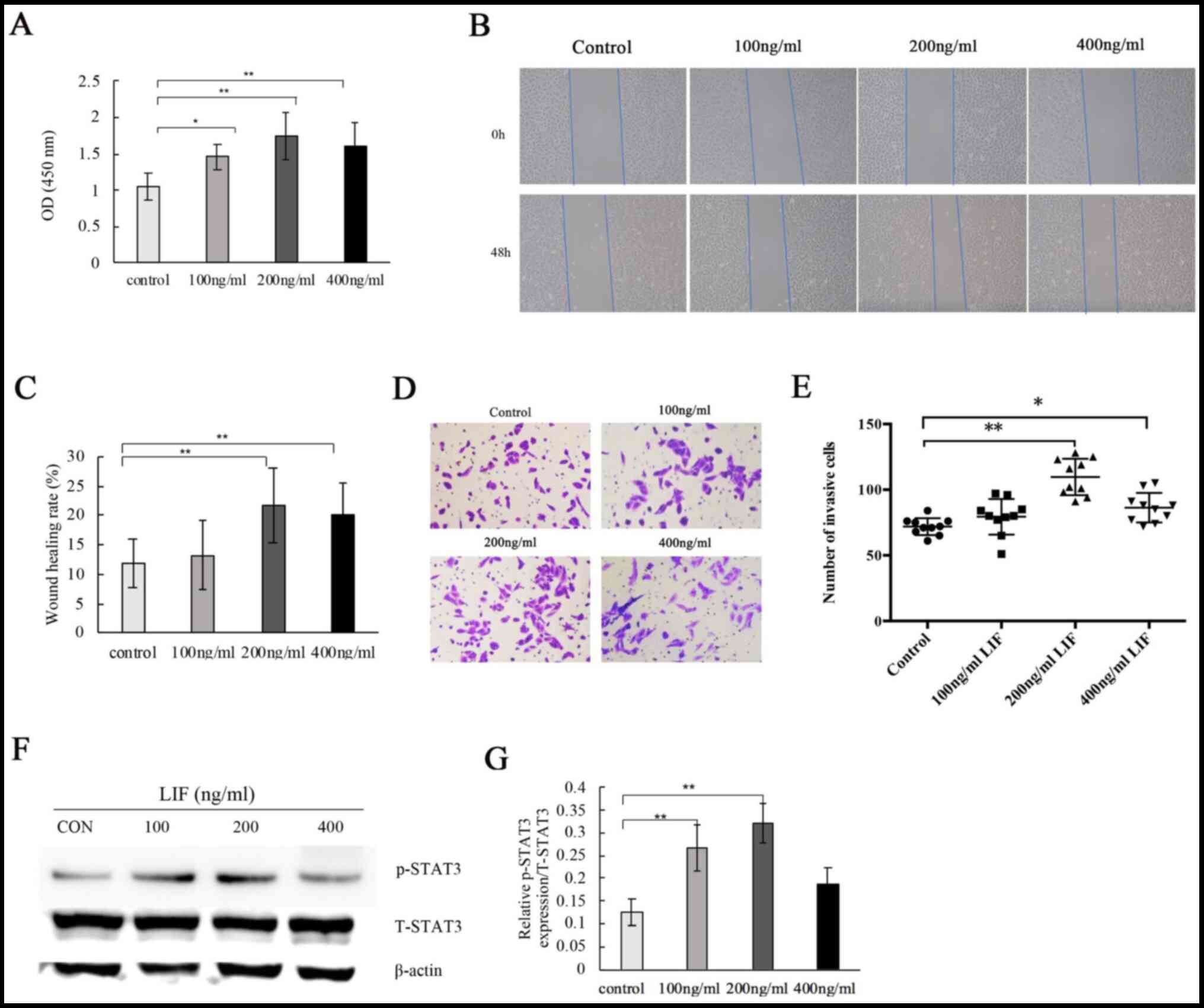
LIF induces adenocarcinoma cell
proliferation, invasion and migration, which is dependent on STAT3
activation
To further evaluate the effects of LIF on cell
proliferation, invasion and migration, NSCLC-derived A549
adenocarcinoma cells were treated with LIF. It was found that LIF
treatment significantly stimulated A549 cell proliferation
(Fig. 3A). Wound healing assay
revealed that high concentrations of LIF (200 and 400 ng/ml)
significantly increased the invasion ability of A549 cells compared
with that of the untreated control and low LIF concentration (100
ng/ml) groups. No significant difference was found for 100 ng/ml
(Fig. 3B and C). Transwell assays
showed that high concentrations of LIF promoted invasion of A549
cells (Fig. 3D). Quantitative
analysis revealed that the number of invasive cells significantly
increased with LIF treatment (200 and 400 ng/ml) compared with that
of the untreated control (Fig.
3E).
To explore the association of STAT3 activation and
LIF overexpression in tissues derived from patients with NSCLC, the
effects of LIF treatment on p-STAT3 expression relative to T-STAT3
expression were measured in A549 cells. Western blot analysis
showed that LIF treatment (100 and 200 ng/ml) significantly
increased the p-STAT3/T-STAT3 ratio compared with that of the
untreated control. Higher concentrations (400 ng/ml) had a tendency
to activate p-STAT3 expression (P>0.05), but the activity was
the highest at 200 ng/ml (Fig. 3F and
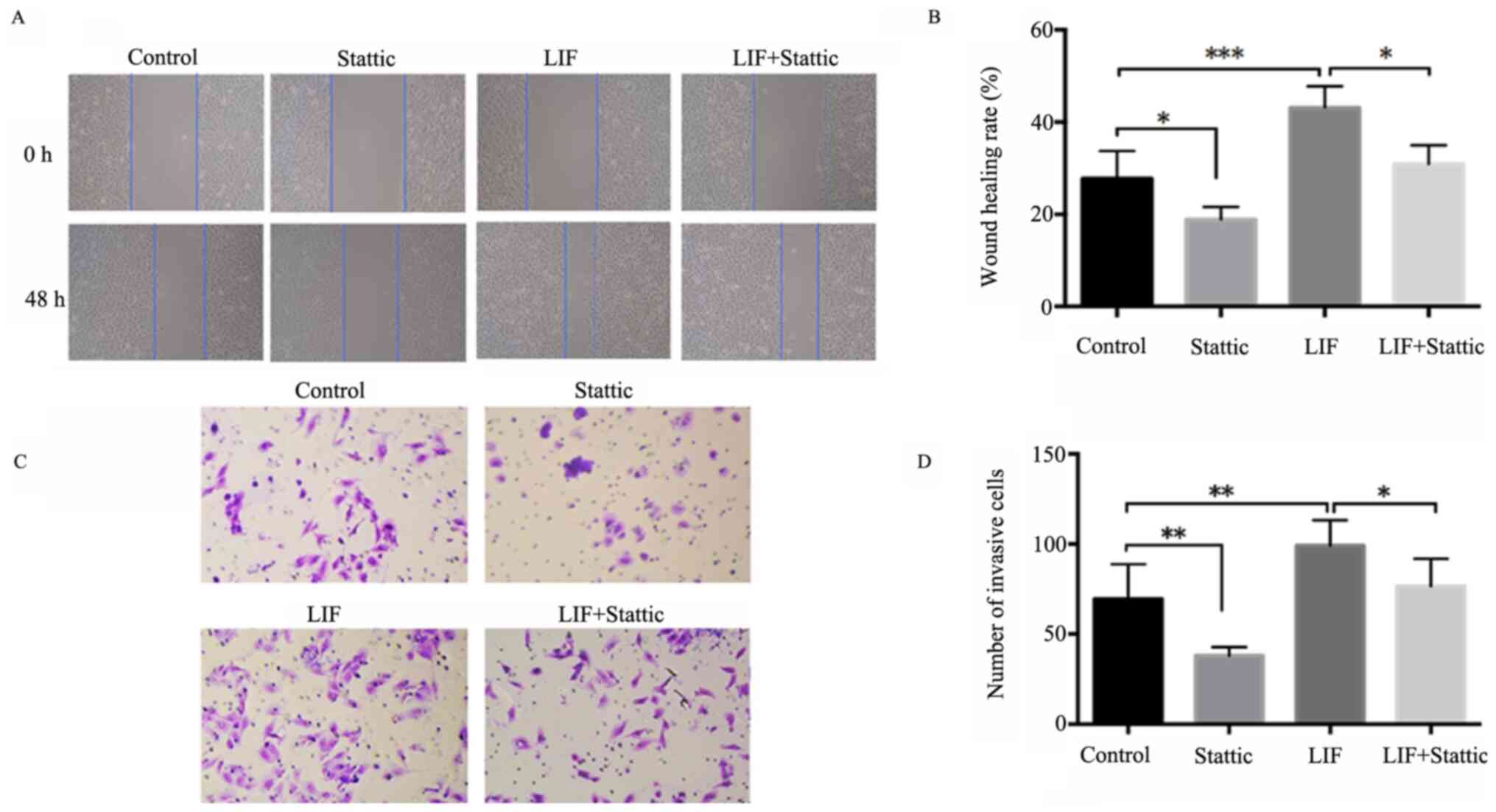
G). Stattic, a specific STAT3 inhibitor, prevents activation,
dimerization and nuclear translocation of STAT3 by interacting with
the SH2 domain (21). It was also
used to determine the effects of blocking STAT3 following LIF
treatment (200 ng/ml) on cell invasion and migration. Wound healing
assays showed that Stattic decreased A549 cell migration both in
the control group treated only with Stattic and in the group
treated with LIF and Stattic (Fig. 4A
and B). Transwell assays showed that Stattic significantly
attenuated A549 cell invasion. There was no significant difference
between the control and LIF + Stattic groups (Fig. 4C and D). Thus, LIF plays a role in
adenocarcinoma cell proliferation, invasion and migration through
activating the LIF/STAT3 signaling pathway.
Association between LIF expression and
clinical features of patients with NSCLC
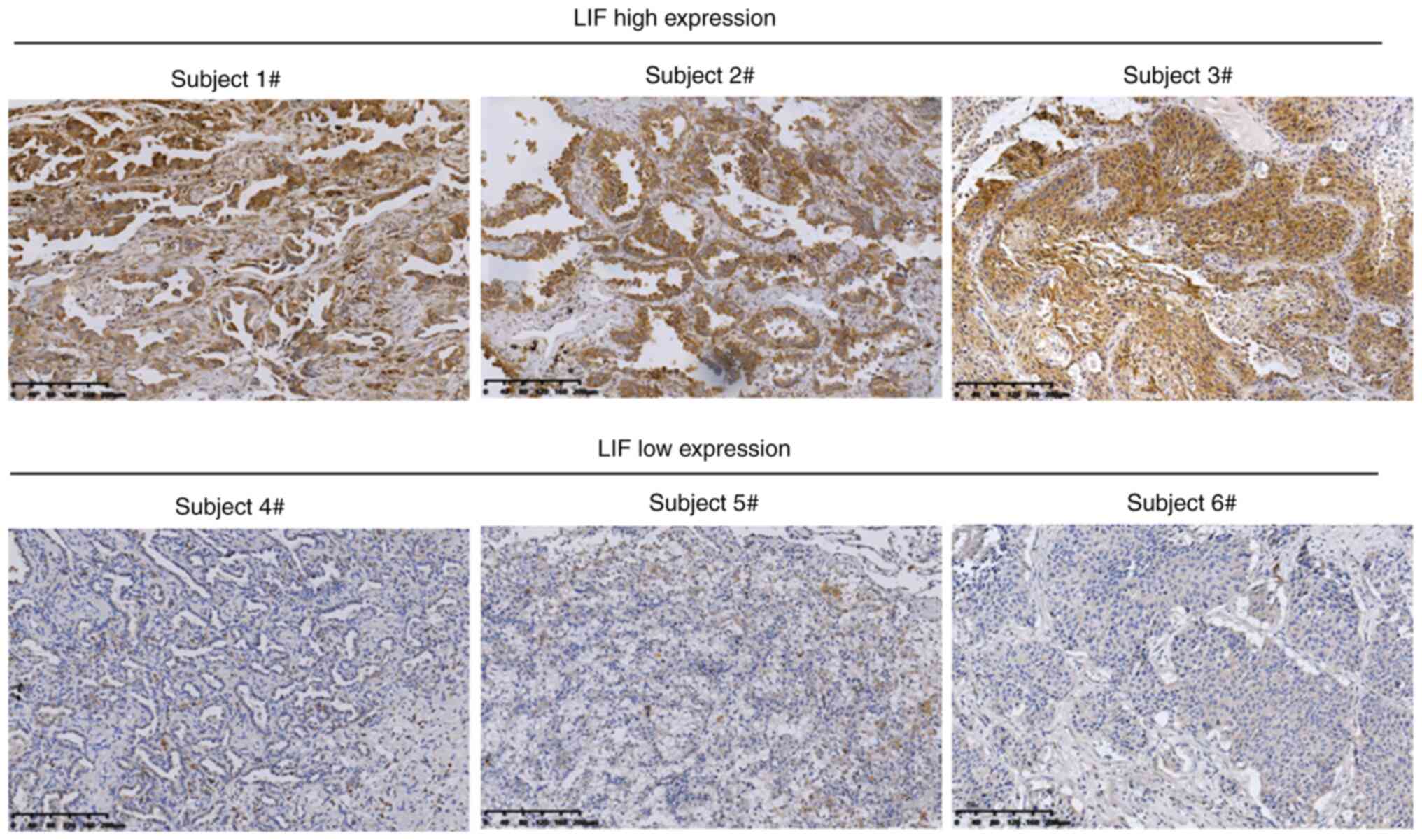
The present study explored the association between
LIF expression in tumor tissues and the clinical features of
patients with NSCLC. Potentially due to the stage and histological
type, LIF expression varied greatly among NSCLC tissues (Fig. 5). Based on the IHC score for LIF
expression, the cohort of 105 patients with NSCLC was divided into
LIFhigh and LIFlow groups (low, 0–1.49; high,
1.50–3.00). As shown in Table I,
high LIF expression significantly correlated with histological type
(P<0.0001) and aggressive tumor characteristics, including lymph
node metastasis (P=0.012) and advanced tumor stage (P=0.043). These
data suggest that LIF expression is negatively correlated with
prognosis in patients with NSCLC.
 | Table I.Association between LIF expression
and clinicopathological characteristics in 105 non-small cell lung
cancer tissues. |
Table I.
Association between LIF expression
and clinicopathological characteristics in 105 non-small cell lung
cancer tissues.
|
|
| LIF expression
level (score) |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Patients, n | Lowa (n=36) | Highb (n=69) | P-value |
|---|
| Age, years |
|
|
|
|
|
<60 | 40 | 9 | 31 | 0.058 |
|
≥60 | 65 | 27 | 38 |
|
| Sex |
|
|
|
|
|
Male | 45 | 18 | 27 | 0.285 |
|
Female | 60 | 18 | 42 |
|
| Smoking status |
|
|
|
|
|
Smoker | 60 | 20 | 40 | 0.812 |
|
Non-smoker | 45 | 16 | 29 |
|
| Histological
type |
|
|
|
|
|
Squamous cell carcinoma | 36 | 28 | 8 |
<0.0001c |
|
Adenocarcinoma | 69 | 8 | 61 |
|
| Tumor stage |
|
|
|
|
|
I–II | 86 | 33 | 53 | 0.047c |
|
III–IV | 19 | 3 | 16 |
|
| Lymph node
metastasis |
|
|
|
|
|
Yes | 10 | 0 | 10 | 0.012c,d |
| No | 95 | 36 | 59 |
|
| Tumor size, cm |
|
|
|
|
|
<2 | 83 | 26 | 57 | 0.214 |
| ≥2 | 22 | 10 | 12 |
|
|
Differentiation |
|
|
|
|
|
Well/moderate | 88 | 30 | 58 | 0.924 |
|
Poor | 17 | 6 | 11 |
|
Discussion
LIF was originally shown to regulate the
differentiation of myeloid leukemia cells (22), and was thus named ‘leukemia
inhibitory factor’. However, other studies have since demonstrated
that LIF can also facilitate the development and progression of a
variety of solid tumors (11,12,23).
In fact, LIF overexpression has been found in breast, colorectal,
head and neck, and ovarian cancer, as well as in melanoma,
nasopharyngeal carcinoma and pancreatic adenocarcinoma. High
expression of LIF in tumors is linked to poor clinical outcomes in
patients with cancer (10–12,23–26).
Increased LIF expression also contributes to tumor resistance to
chemotherapy and radiotherapy (11,12).
These lines of evidence strongly suggest that LIF can promote
tumorigenesis, particularly in solid tumors. However, the role of
LIF in lung cancer remained unknown.
The present study provides evidence that
overexpression of LIF in NSCLC tissue is associated with cell
metastasis to the lymph node; patients who underwent surgery had
earlier stages and fewer lymph node metastases, and a statistically
significant correlation was noted between LIF expression and lymph
node metastasis in enrolled patients. These results indicate that
LIF plays an important role in the tumorigenesis of NSCLC, similar
to its previously reported role in other solid tumors, as
aforementioned. Since LIF overexpression can increase the
proliferation rate of cultured cancer cells, the growth rate of
xenograft tumors and the metastasis of multiple human tumor types
(10–12,23,24). The
present study also evaluated the effects of LIF on a human lung
cancer cell line in vitro. It was found that LIF stimulation
promoted cancer cell proliferation, migration and invasion, further
suggesting that LIF induces metastasis of lung cancer. These in
vitro results are consistent with findings from clinical
specimens, which showed that LIF is overexpressed in tumor tissues
along with activation of the STAT3 pathway. Together, these
findings demonstrate that LIF is important for lung cancer
progression. The present findings also provide a useful guide in
clinical practice, since high LIF expression could be used a marker
of regional lymph node metastasis in patients with NSCLC.
The molecular mechanisms of LIF's role in promoting
tumor metastasis are not fully understood. LIF functions in an
autocrine and/or paracrine manner through binding to the LIF
receptor (LIF-R) complex (which is composed of LIF-R and gp-130),
which in turn activates certain signaling pathways, including the
PI3K/AKT and JAK/STAT3 pathways (27–29).
STAT3 is a critical downstream effector of LIF signaling in
numerous types of cells and tissues, such as embryonic stem cells
and monocytes (27,30). LIF expression in colorectal cancer
cells can be induced by hypoxia and transcription factor
hypoxia-inducible factor-2α (18).
In addition, LIF can negatively regulate p53 in colorectal cancer
cells, which is associated with its effect of increasing resistance
of cancer cells to chemotherapy and radiotherapy (31). Yang et al (32) showed that STAT5B and STAT6 could be
effective prognostic biomarkers of survival in patients with NSCLC,
and that STAT2 may be a promising therapeutic target for the
treatment of NSCLC, and in particular lung adenocarcinoma. STAT3 is
frequently activated in multiple types of human cancer, and is
crucial to the survival and growth of tumor cells (33–36).
Jiang et al (37) reported
that immunoreactivity of p-STAT3 was significantly increased in
lung cancer tissue compared with that of normal tissue. The present
study showed a high level of p-STAT3 expression in tumor tissues
compared with that of the corresponding adjacent and normal lung
tissues. Furthermore, the expression of p-STAT3 was positively
correlated with LIF expression in NSCLC tissue samples. LIF (200
and 400 ng/ml) stimulation also induced STAT3 phosphorylation in
A549 cells, which is correlated with LIF expression levels in NSCLC
tissues. The STAT3-specific inhibitor Stattic inhibited cell
migration and invasion. These findings suggest that LIF promotes
the development and progression of NSCLC through activating the
LIF/STAT3 signaling pathway, although other pathways could also be
involved.
Although there are significant differences between
lung adenocarcinoma and lung squamous cell carcinoma in regards to
clinical features, gene mutations, immune checkpoints, cytokines
and non-coding RNA expression (38–41), the
comparison of LIF expression between these two subtypes of NSCLC
has not been reported to date. The present results show that LIF
and p-STAT3 expression levels are higher in lung adenocarcinoma
compared with those of lung squamous cell carcinoma. However, the
detailed molecular mechanisms underlying these differences need to
be further studied.
In conclusion, the present results demonstrate that
LIF overexpression can promote NSCLC development through activating
the LIF/STAT3 signaling pathway. Therefore, LIF may serve as a
potential prognostic marker for patients with NSCLC, and LIF
signaling pathways could be potential targets for anticancer drug
discovery for NSCLC.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Nature
Scientific Foundation of Ningbo City (grant no. 2019A610229), and
the Major Project of Diseases Prevention and Treatment of
Traditional Chinese Medicine of Zhejiang Province (grant no.
2018ZY010). The funding bodies had no role in the design of the
study; in the collection, analysis or interpretation of the data;
or in the writing of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HW performed the IHC staining, participated in
patient enrollment and follow-up, performed the statistical
analysis, and drafted the manuscript. SS performed the RT-qPCR,
western blotting and IHC staining. MJ and KH participated in
patient enrollment, clinical data analysis and IHC staining. LC
performed the cell culture and in vitro experiments,
including cell proliferation and invasion assays, and the
wound-healing assay. WY conceived the study, participated in its
design and coordination, and helped to draft the manuscript. All
authors read and approved the final manuscript. HW, SS, LC and WY
confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Yinzhou People's Hospital (approval no. 2018003).
Written informed consent to participate was obtained from all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
LIF
|
leukemia inhibitory factor
|
|
NSCLC
|
non-small cell lung cancer
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
JAK
|
Janus tyrosine kinase
|
|
ERK
|
extracellular signal-regulated protein
kinase
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
IHC
|
immunohistochemistry
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
FBS
|
fetal bovine serum
|
|
CCK-8
|
Cell Counting Kit-8
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xiong D, Zhu SQ, Wu YB, Jin C, Jiang JH,
Liao YF, Long X, Wu HB, Xu JJ, Li JJ and Ding JY: Ring finger
protein 38 promote non-small cell lung cancer progression by
endowing cell EMT phenotype. J Cancer. 9:841–850. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pirker R: Conquering lung cancer: Current
status and prospects for the future. Pulmonology. 26:283–290. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scrima M, Zito Marino F, Oliveira DM,
Marinaro C, La Mantia E, Rocco G, De Marco C, Malanga D, De Rosa N,
Rizzuto A, et al: Aberrant signaling through the HER2-ERK1/2
pathway is predictive of reduced disease-free and overall survival
in early stage non-small cell lung cancer (NSCLC) Patients. J
Cancer. 8:227–239. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rotow J and Bivona TG: Understanding and
targeting resistance mechanisms in NSCLC. Nat Rev Cancer.
17:637–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nicola NA and Babon JJ: Leukemia
inhibitory factor (LIF). Cytokine Growth Factor Rev. 26:533–544.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dahéron L, Opitz SL, Zaehres H, Lensch MW,
Andrews PW, Itskovitz-Eldor J and Daley GQ: LIF/STAT3 signaling
fails to maintain self-renewal of human embryonic stem cells. Stem
Cells. 22:770–778. 2004. View Article : Google Scholar
|
|
8
|
Niwa H, Ogawa K, Shimosato D and Adachi K:
A parallel circuit of LIF signalling pathways maintains
pluripotency of mouse ES cells. Nature. 460:118–122. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pera MF and Tam PP: Extrinsic regulation
of pluripotent stem cells. Nature. 465:713–720. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wysoczynski M, Miekus K, Jankowski K,
Wanzeck J, Bertolone S, Janowska-Wieczorek A, Ratajczak J and
Ratajczak MZ: Leukemia inhibitory factor: A newly identified
metastatic factor in rhabdomyosarcomas. Cancer Res. 67:2131–2140.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu SC, Tsang NM, Chiang WC, Chang KP,
Hsueh C, Liang Y, Juang JL, Chow KP and Chang YS: Leukemia
inhibitory factor promotes nasopharyngeal carcinoma progression and
radioresistance. J Clin Invest. 123:5269–5283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu H, Yue X, Zhao Y, Li X, Wu L, Zhang C,
Liu Z, Lin K, Xu-Monette ZY, Young KH, et al: LIF negatively
regulates tumour-suppressor p53 through Stat3/ID1/MDM2 in
colorectal cancers. Nat Commun. 5:52182014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin TA, Wu TS, Li YJ, Yang CN, Illescas
Ralda MM and Chang HH: Role and mechanism of LIF in oral squamous
cell carcinoma progression. J Clin Med. 9:2952020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hergovich A: YAP-Hippo signalling
downstream of leukemia inhibitory factor receptor: Implications for
breast cancer. Breast Cancer Res. 14:3262012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen D, Sun Y, Wei Y, Zhang P, Rezaeian
AH, Teruya-Feldstein J, Gupta S, Liang H, Lin HK, Hung MC and Ma L:
LIFR is a breast cancer metastasis suppressor upstream of the
Hippo-YAP pathway and a prognostic marker. Nat Med. 18:1511–1517.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Humbert L, Ghozlan M, Canaff L, Tian J and
Lebrun JJ: The leukemia inhibitory factor (LIF) and p21 mediate the
TGFβ tumor suppressive effects in human cutaneous melanoma. BMC
Cancer. 15:2002015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo Q, Wang C, Jin G, Gu D, Wang N, Song
J, Jin H, Hu F, Zhang Y, Ge T, et al: LIFR functions as a
metastasis suppressor in hepatocellular carcinoma by negatively
regulating phosphoinositide 3-kinase/AKT pathway. Carcinogenesis.
36:1201–1212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu L, Yu H, Zhao Y, Zhang C, Wang J, Yue
X, Yang Q and Hu W: HIF-2α mediates hypoxia-induced LIF expression
in human colorectal cancer cells. Oncotarget. 6:4406–4417. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shiao YH, Palli D, Caporaso NE, Alvord WG,
Amorosi A, Nesi G, Saieva C, Masala G, Fraumeni JF Jr and Rice JM:
Genetic and immunohistochemical analyses of p53 independently
predict regional metastasis of gastric cancers. Cancer Epidemiol
Biomarkers Prev. 9:631–633. 2000.PubMed/NCBI
|
|
20
|
Morse DL, Carroll D, Weberg L, Borgstrom
MC, Ranger-Moore J and Gillies RJ: Determining suitable internal
standards for mRNA quantification of increasing cancer progression
in human breast cells by real-time reverse transcriptase polymerase
chain reaction. Anal Biochem. 342:69–77. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guha P, Gardell J, Darpolor J, Cunetta M,
Lima M, Miller G, Espat NJ, Junghans RP and Katz SC: STAT3
inhibition induces Bax-dependent apoptosis in liver tumor
myeloid-derived suppressor cells. Oncogene. 38:533–548. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moreau JF, Donaldson DD, Bennett F,
Witek-Giannotti J, Clark SC and Wong GG: Leukaemia inhibitory
factor is identical to the myeloid growth factor human interleukin
for DA cells. Nature. 336:690–692. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li X, Yang Q, Yu H, Wu L, Zhao Y, Zhang C,
Yue X, Liu Z, Wu H, Haffty BG, et al: LIF promotes tumorigenesis
and metastasis of breast cancer through the AKT-mTOR pathway.
Oncotarget. 5:788–801. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuphal S, Wallner S and Bosserhoff AK:
Impact of LIF (leukemia inhibitory factor) expression in malignant
melanoma. Exp Mol Pathol. 95:156–165. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang D, Liu K, Yang Y, Wang T, Rao Q, Guo
W and Zhang Z: Prognostic value of leukemia inhibitory factor and
its receptor in pancreatic adenocarcinoma. Future Oncol.
16:4461–4473. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McLean K, Tan L, Bolland DE, Coffman LG,
Peterson LF, Talpaz M, Neamati N and Buckanovich RJ: Leukemia
inhibitory factor functions in parallel with interleukin-6 to
promote ovarian cancer growth. Oncogene. 38:1576–1584. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takahashi Y, Takahashi M, Carpino N, Jou
ST, Chao JR, Tanaka S, Shigeyoshi Y, Parganas E and Ihle JN:
Leukemia inhibitory factor regulates trophoblast giant cell
differentiation via Janus kinase 1-signal transducer and activator
of transcription 3-suppressor of cytokine signaling 3 pathway. Mol
Endocrinol. 22:1673–1681. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heinrich PC, Behrmann I, Haan S, Hermanns
HM, Müller-Newen G and Schaper F: Principles of interleukin
(IL)-6-type cytokine signalling and its regulation. Biochem J.
374:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Watanabe S, Umehara H, Murayama K, Okabe
M, Kimura T and Nakano T: Activation of Akt signaling is sufficient
to maintain pluripotency in mouse and primate embryonic stem cells.
Oncogene. 25:2697–2707. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Metcalf D: The unsolved enigmas of
leukemia inhibitory factor. Stem Cells. 21:5–14. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yue X, Zhao Y, Zhang C, Li J, Liu Z, Liu J
and Hu W: Leukemia inhibitory factor promotes EMT through
STAT3-dependent miR-21 induction. Oncotarget. 7:3777–3790. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang M, Chen H, Zhou L, Chen K and Su F:
Expression profile and prognostic values of STAT family members in
non-small cell lung cancer. Am J Transl Res. 11:4866–4880.
2019.PubMed/NCBI
|
|
33
|
Lassmann S, Schuster I, Walch A, Göbel H,
Jütting U, Makowiec F, Hopt U and Werner M: STAT3 mRNA and protein
expression in colorectal cancer: Effects on STAT3-inducible targets
linked to cell survival and proliferation. J Clin Pathol.
60:173–179. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin L, Liu A, Peng Z, Lin HJ, Li PK, Li C
and Lin J: STAT3 is necessary for proliferation and survival in
colon cancer-initiating cells. Cancer Res. 71:7226–7237. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L and Karin M: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bollrath J, Phesse TJ, von Burstin VA,
Putoczki T, Bennecke M, Bateman T, Nebelsiek T, Lundgren-May T,
Canli O, Schwitalla S, et al: gp130-mediated Stat3 activation in
enterocytes regulates cell survival and cell-cycle progression
during colitis-associated tumorigenesis. Cancer Cell. 15:91–102.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang R, Jin Z, Liu Z, Sun L, Wang L and
Li K: Correlation of activated STAT3 expression with
clinicopathologic features in lung adenocarcinoma and squamous cell
carcinoma. Mol Diagn Ther. 15:347–352. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tian Y, Yu M, Sun L, Liu L, Wang J, Hui K,
Nan Q, Nie X, Ren Y and Ren X: Distinct Patterns of mRNA and lncRNA
expression differences between lung squamous cell carcinoma and
adenocarcinoma. J Comput Biol. 27:1067–1078. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang XC, Wang J, Shao GG, Wang Q, Qu X,
Wang B, Moy C, Fan Y, Albertyn Z, Huang X, et al: Comprehensive
genomic and immunological characterization of Chinese non-small
cell lung cancer patients. Nat Commun. 10:17722019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim H, Kwon HJ, Park SY, Park Y, Park E
and Chung JH: Clinicopathological analysis and prognostic
significance of programmed cell death-ligand 1 protein and mRNA
expression in non-small cell lung cancer. PLoS One.
13:e01986342018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu S, Wang X, Qin W, Genchev GZ and Lu H:
Transcription factors contribute to differential expression in
cellular pathways in lung adenocarcinoma and lung squamous cell
carcinoma. Interdiscip Sci. 10:836–847. 2018. View Article : Google Scholar : PubMed/NCBI
|