Introduction
Ovarian cancer ranks the most lethal gynecologic
malignancy and is now the second leading cause of death in women
around the world (1,2). It has been reported that <50% of
patients survive for >5 years after diagnosis (3). Ovarian cancer affects women of all
ages but is most commonly diagnosed after menopause (4). Since early-stage disease is typically
asymptomatic and symptoms of late-stage disease are nonspecific,
>75% of affected women are diagnosed at an advanced stage
(5–7). Therefore, it is important to explore
the risk factors at the early stage.
Ferroptosis is a unique iron-dependent nonapoptotic
cell death that is driven by depletion of glutathione (GSH) and
accumulation of lipid reactive oxygen species (ROS) (8). Recently, elevated expression of
nuclear factor erythroid 2-related factor 2 (Nrf2) was identified
as an antioxidant transcription factor that defends malignant cells
from ferroptosis (9). There are a
number of different downstream effectors that are regulated by
Nrf2, including heme oxygenase-1 (HO-1), glutathione peroxidase 4
(GPX4) and the cystine/glutamate antiporter system X(c)(−) (xCT)
(10,11). HO-1 is associated with the
endogenous antioxidant system and is an important member of the
defense system (12). HO-1 can be
activated by Nrf2, which subsequently eliminates hydroxyl-free
radicals and excessive oxidation of lipids (12). xCT and GPX4 are two key regulators
of ferroptosis (13).
Downregulation of xCT and GPX4 can decrease intracellular cystine
concentrations and lipid peroxide degradation, thereby leading to
the accumulation of intracellular lipid peroxide and subsequent
ferroptosis (13,14). Therefore, targeting the Nrf2 system
may represent a potential therapeutic target for the treatment of
ovarian cancer.
Carboxymethylated pachyman (CMP) is a
carboxymethylated derivative of pachyman isolated from Poria
cocos (Chinese name: Fu Ling) (15). Studies have demonstrated that CMP
is characterized by immune regulatory, antitumor and antioxidant
activities (15,16). For instance, CMP can improve colon
injuries induced by 5-fluorouracil in CT26 tumor-bearing mice by
regulating the NF-κB, Nrf2-ARE and MAPK/P38 pathways (16). However, whether CMP could
contribute to the treatment of ovarian cancer remains to be
elucidated.
Materials and methods
Cell culture
The ovarian cancer cell lines Hey and SKOV3 were
purchased from Procell Life Science & Technology Co., Ltd. A
human normal ovarian epithelial cell line (IOSE80) was purchased
from BioVector NTCC, Inc. Cell line authentication was performed
using short tandem repeats. Hey and SKOV3 cells were maintained in
DMEM (HyClone; Cytiva) supplemented with 10% fetal bovine serum
(FBS; HyClone; Cytiva) and 1% antibiotics (penicillin and
streptomycin; HyClone; Cytiva). IOSE80 cells were cultured in 90%
RPMI-1640 medium (HyClone; Cytiva) and 10% FBS (HyClone; Cytiva).
All cells were maintained in an incubator and 5% CO2 at
37°C.
CCK-8 assay
In brief, SKOV3 and Hey cells were seeded in 96-well
plates at a density of 3,000 cells/well overnight at 37°C.
Thereafter, SKOV3 and Hey cells were incubated with 100, 200, 300,
400 and 500 µg/ml CMP for 24 h at 37°C. Then, 10 µl CCK-8 (Beijing
Solarbio Science & Technology Co., Ltd.) was added to each well
and incubated at 37°C for 4 h. The absorbance was determined at 450
nm using a microplate reader (Thermo Fisher Scientific, Inc.).
Annexin-PE/7-AAD assay
SKOV3 and Hey cells were seeded in 6-well plates at
a density of 10,000 cells/well overnight at 37°C. Then, SKOV3 and
Hey cells were treated with 100 and 200 µg/ml CMP for 24 h at 37°C.
Cell death was analyzed using an Annexin V-PE/7-AAD Apoptosis
Detection kit (cat. no. WE0328; Beijing Biolab Technology Co.,
Ltd.) according to the manufacturer's instructions. In brief, the
cells were centrifuged at 1,000 × g for 5 min at 37°C and
collected. Then, 250 µl binding buffer was added, and the cell
density was adjusted to 106 cells/ml. Next, the cells
were incubated with 5 µl Annexin V-PE and 10 µl 7-AAD for 15 min at
room temperature. Subsequently, 400 µl PBS was added, and the data
were analyzed using a CytoFLEX V2-B4-R2 flow cytometer (cat. no.
C02945; Beckman Coulter, Inc.). The data [early (Q3) + late (Q2)
apoptotic cells] were analyzed using FlowJo v10 (FlowJo LLC).
Reverse transcription-quantitative PCR
(RT-qPCR)
SKOV3 and Hey cells were seeded in 6-well plates at
a density of 10,000 cells/well overnight at 37°C. Then, SKOV3 and
Hey cells were treated with 100 and 200 µg/ml CMP for 24 h at 37°C
in the presence or absence of ferrostatin-1 (Fer-1, MCE). Total RNA
was isolated from SKOV3 and Hey cells using RNAVzol LS (Vigorous
Biotechnology Beijing Co., Ltd.) according to the manufacturer's
protocol. The concentration and purity of RNA samples were
determined by measuring the optical density (OD) 260/OD280.
RT-PCR was performed using a Takara PrimeScript One
Step RT-PCR kit (Takara Bio, Inc.) according to the manufacture's
protocols with three experiments replicated. The PCR amplifications
were performed in a 50-µl reaction system containing 20 µl RNase
Free ddH2O, 25 µl 2X Step Buffer, 2 µl PrimeScript 1
Step Enzyme Mix, 1 µl upstream primer (1 µM), and 1 µl downstream
primer (1 µM). The PCR was as follows: 50°C for 30 min; 94°C for 2
min; and 30 cycles of 94°C for 30 sec, 55°C for 30 sec, 72°C for 1
min. GAPDH was used as an internal control using the
2−∆∆Cq method (17).
The primers were designed using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi?LINK_LOC=BlastHome)
and the sequences were listed in Table
I.
 | Table I.Primers used in the present study. |
Table I.
Primers used in the present study.
| Primer name | Sequence (5′-3′) |
|---|
| CHAC1-F |
CCCCATCCTGGAACTTGACC |
| CHAC1-R |
CTATGGATGGCTGGGCTGAG |
| PTGS2-F |
GAGGGATCTGTGGATGCTTCG |
| PTGS2-R |
AAACCCACAGTGCTTGACAC |
| NRF2-F |
AAAGTGGCTGCTCAGAATTGC |
| NRF2-R |
TTGCCATCTCTTGTTTGCTGC |
| HO-1-F |
AGGGAATTCTCTTGGCTGGC |
| HO-1-R |
GCTGCCACATTAGGGTGTCT |
| GAPDH-F |
TTGCCCTCAACGACCACTTT |
| GAPDH-R |
TGGTCCAGGGGTCTTACTCC |
Western blotting
Total proteins were isolated from SKOV3 and Hey
cells using a total protein extraction kit (Beijing Solarbio
Science & Technology Co., Ltd.) and collected following
centrifugation at 12,000 × g for 30 min at 4°C. A BCA protein assay
kit (Pierce; Thermo Fisher Scientific, Inc.) was used to determine
the protein concentration. A total of 20 µg protein was separated
using 12% SDS-PAGE (F15012Gel; ACE Biotechnology Co., Ltd.),
transferred onto polyvinylidene difluoride membranes (Pierce;
Thermo Fisher Scientific, Inc.) and blocked with 5% fat-free milk
at room temperature for 2 h. The membrane was incubated with the
following primary antibodies: Nrf2 (cat. no. 12721; 1:1,000; Cell
Signaling Technology, Inc.), HO-1 (cat. no. Ab52947; 1:1,000;
Abcam), xCT (cat. no. 12691 for human, cat. no. 98051 for mouse;
1:1,000; Cell Signaling Technology, Inc.), GPX4 (cat. no. ab125066;
Abcam) and GAPDH (cat. no. 5174; Cell Signaling Technology, Inc.),
at 4°C overnight. Then, the membranes were washed with PBST thrice.
Next, the membrane was incubated with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG (both 1:5,000; cat. no.
ZB-2301; OriGene Technologies, Inc.) for 2 h at room temperature.
Enhanced chemiluminescence (MilliporeSigma) was used to determine
the protein concentrations according to the manufacturer's
protocol. Signals were detected using a Super ECL Plus kit (Nanjing
KeyGen Biotech Co., Ltd.) Quantitative analysis was performed using
UVP 7.0 software (UVP LLC). Relative protein expression was
normalized to GAPDH. All experiments were repeated thrice. ImageJ
v1.43b software (National Institutes of Health) was used for
densitometry analysis.
Quantification of Fe2+,
superoxide dismutase (SOD), glutathione (GSH) and malondialdehyde
(MDA)
The intracellular levels of Fe2+, MDA,
SOD and GSH were determined using colorimetric assay kits,
including an iron assay kit (cat. no. EC-BC-K304-S; Elabscience
Biotechnology, Inc.), Lipid Peroxidation MDA Assay kit (cat. no.
A003-1; Nanjing Jiancheng Biotechnology Co. Ltd.), superoxide
dismutase (SOD) Assay kit (cat. no. A001-3-1; Nanjing Jiancheng
Biotechnology Co. Ltd.) and Reduced Glutathione Assay kit (cat. no.
A005-1; Nanjing Jiancheng Biotechnology Co. Ltd.) according to the
manufacturer's instructions.
DCFH-DA
(2,7-Dichlorodi-hydrofluorescein diacetate) staining
SKOV3 and Hey cells were seeded in 6-well plates at
a density of 10,000 cells/well overnight. Then, SKOV3 and Hey cells
were treated with 100 and 200 µg/ml CMP for 24 h at 37°C. Then, the
cells were incubated with 1 ml DCFH-DA (1:1,000) at room
temperature for 20 min. Cells were washed with DMEM culture without
FBS thrice. Representative images were obtained under a
fluorescence microscope (magnification, ×20; CKX53; Olympus
Corporation).
In vivo assay
A total of eight nude female BALB/cA-nu mice (6
weeks old, weighing 20.1±1.8 g) were purchased from SPF (Beijing)
Biotechnology Co., Ltd. (n=4 per group). The mice were kept in a 12
h light/dark cycle with controlled humidity (50–70%) and
temperature (20–24°C) with free access to food and water. They were
randomly assigned to two experimental groups (n=4 per group). All
experiments were approved according to the Ethics Committee of
Tengzhou Central People's Hospital (Tengzhou, China; approval
number TZH2020AH6) and were performed according to the National
Institute of Health guidelines. SKOV3 cells (2×106 cells
in 100 µl PBS) were subcutaneously injected into the flanks of
6-week-old female nude mice to induce tumor formation. After 24 h,
mice in the control group were orally administered distilled water
(20 ml/kg, 1 time/day for 28 days). Mice in the therapy group were
orally administered CMP (50 mg/kg, 1 time/day for 28 days). All
mice were sacrificed 28 days after injection by resection of the
decapitation under deep isoflurane anesthesia (5%) (1). The successful induction of anesthesia
was confirmed by observation of the following parameters:
respiration decreased in frequency and increased in depth, eyelid
and cornea reflexes disappeared, muscle tension and the reflex
response reduced, and no response to pain or other stimulation was
exhibited. Tumor grafts were excised, weighed, and harvested for
further analysis. Tumor diameters were measured at regular
intervals, and the tumor volume was calculated using the following
formula: volume = length × width2/2.
Statistical analysis
All data were expressed as the mean ± standard
deviation. Unpaired Student's t-test was used to compare the
differences between the two groups. One-way analysis of variance
followed by Turkey analysis was used to analyze differences among
three or more groups. Statistical analysis was performed using
GraphPad Prism 8.0 Software (GraphPad Software, Inc.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Carboxymethylated pachyman (CMP)
induces ovarian cancer cell death
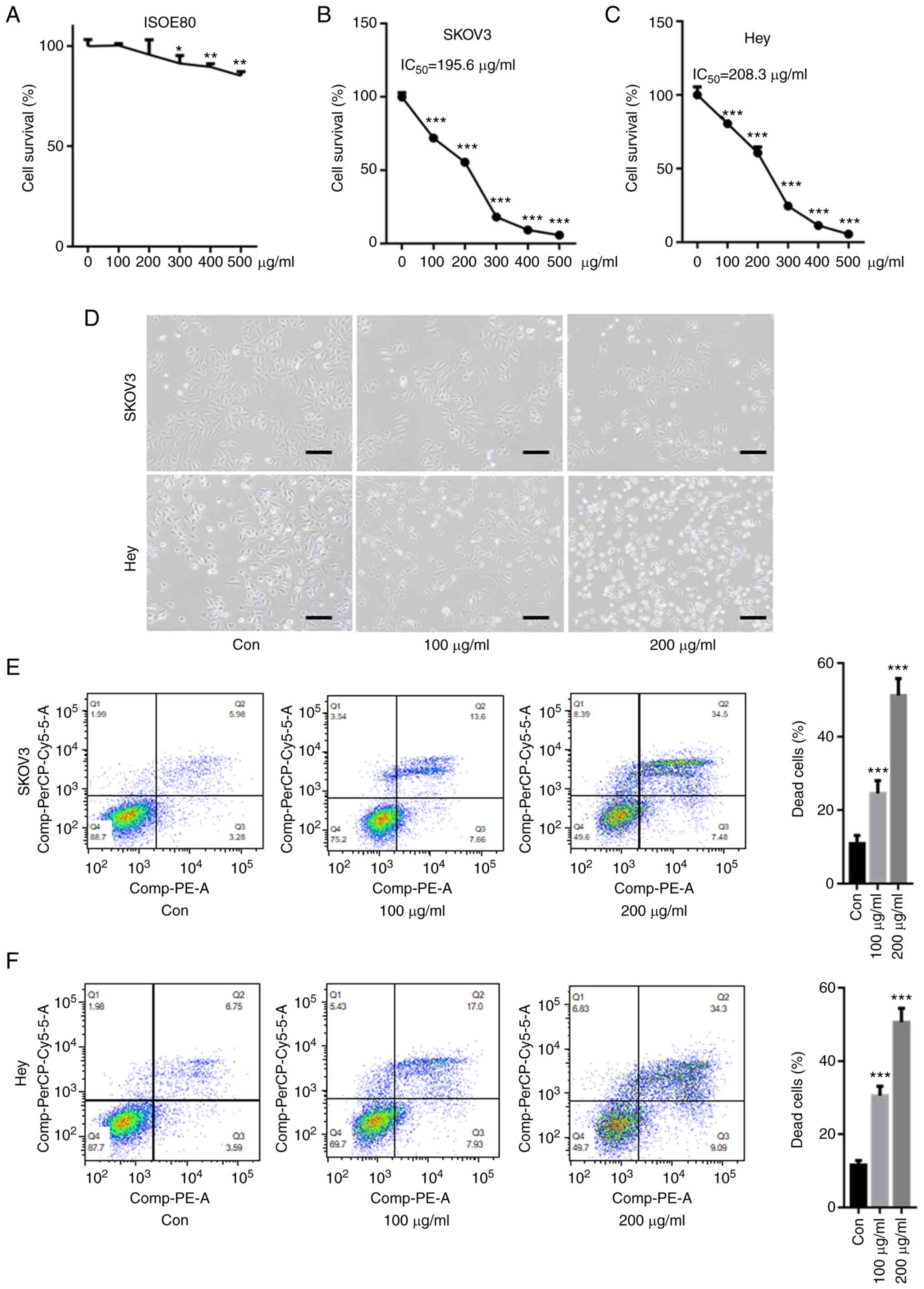
First, the cytotoxicity of CPM on a human normal
ovarian epithelial cell line, IOSE80 was tested. As shown in
Fig. 1A, 100 and 200 µg/ml CPM did
not significantly decreased cell survival of IOSE80, but 300, 400,
500 µg/ml CPM slightly reduced IOSE80 cell survival rate. The CCK-8
assay showed that CMP significantly reduced the cell survival rate
in SKOV3 and Hey cells in a dose-dependent manner (Fig. 1B and C). Compared with SKOV3 and
Hey cells, CMP did not result in too much cytotoxicity in IOSE80
cells, indicating the drug was not toxic to normal cells. The
IC50 values of CMP in SKOV3 and Hey cells were 195.6 and
208.3 µg/ml, respectively. Optical microscopy images demonstrated
that 100 and 200 µM CMP clearly reduced cell viability and
increased cell death in SKOV3 and Hey cells (Fig. 1D). Furthermore, flow cytometry
assays suggested that the cell death of both SKOV3 and Hey cells
after 100 and 200 µg/ml CMP treatment, respectively, was
significantly increased compared with that of the control (Fig. 1E and F).
CMP induces ferroptosis in SKOV3 and
Hey cells
SKOV3 and Hey cells were preincubated with different
inhibitors, including ferroptosis inhibitors (ferrostatin-1,
Fer-1), apoptosis inhibitors (Z-VAD-fluoromethylketone, Z-VAD-FMK),
autophagy inhibitors (3-methyladenine, 3-MA), and necrosis
inhibitors (necrostatin-1, Nec-1). As shown in Fig. 2A and B, CMP-induced cell death was
largely reversed by preincubation with Fer-1 and Z-VAD-FMK but was
not abolished by Nec-1 and 3-MA. Z-VAD-FMK decreased CMP-induced
cell death by ~6.4%, whereas Fer-1 reduced CMP-induced cell death
by ~26.1% (Fig. 2A and B). The
present study further evaluated the mRNA levels of PTGS2 and CHAC1,
two important ferroptosis markers, in SKOV3 and Hey cells treated
with CMP and Fer-1. RT-qPCR analysis indicated that CMP
significantly increased PTGS2 and CHAC1 mRNA levels, but
preincubation with Fer-1 obviously reduced PTGS2 and CHAC1 mRNA
levels in SKOV3 and Hey cells (Fig. 2C
and D). These data indicated that CMP could induce ferroptosis
in ovarian cancer cells.
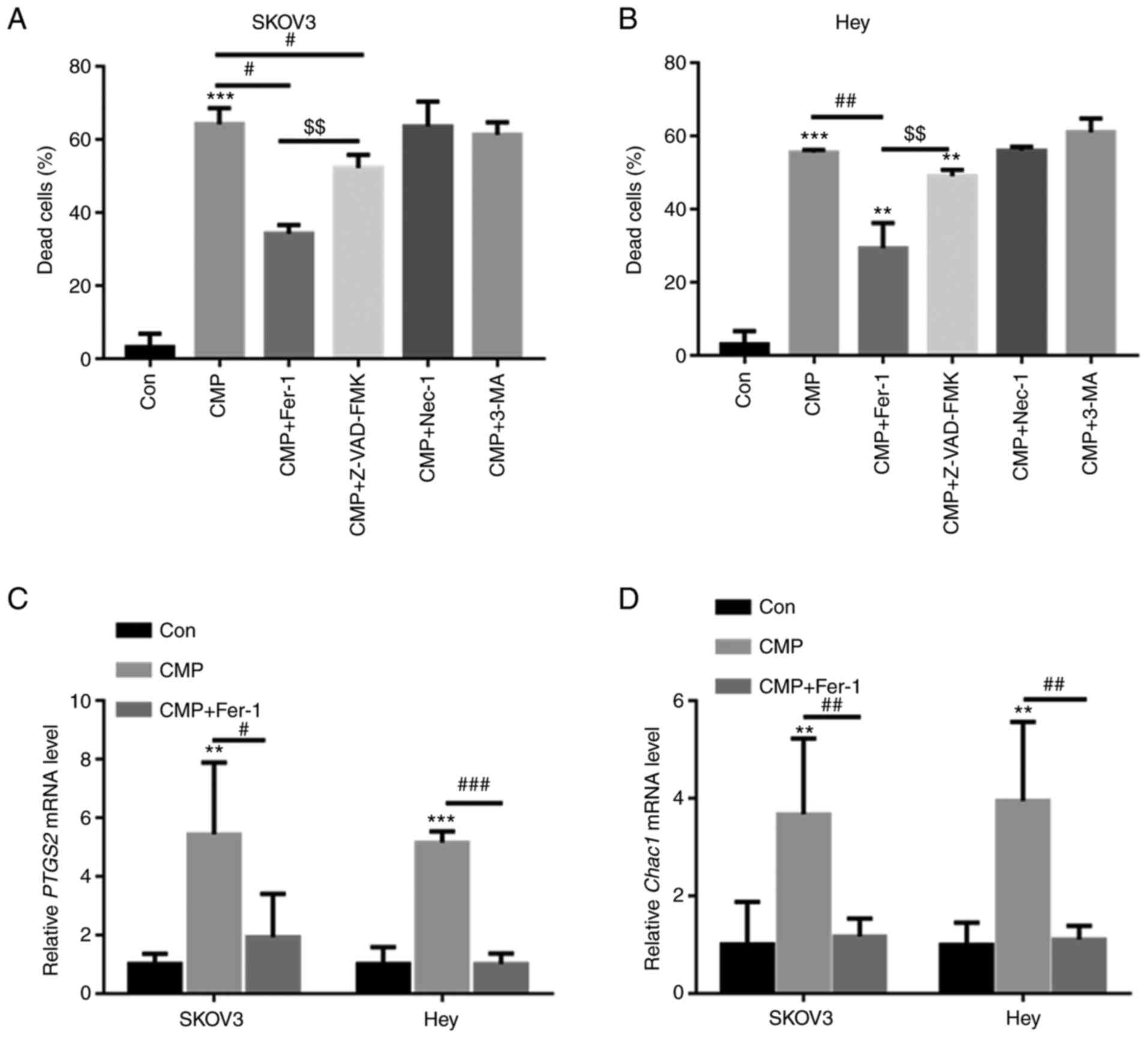 | Figure 2.CMP induces ferroptosis in SKOV3 and
Hey cells. SKOV3 and Hey cells were preincubated with 1 mM Fer-1,
10 mM Z-VAD-FMK, 10 mM 3-MA and 10 mM Nec-1 for 1 h. Then, the
cells were treated with 100 µg/ml CMP for 24 h. Cell death was
quantified using a CCK-8 assay in (A) SKOV3 and (B) Hey cells.
Quantitative PCR analysis indicated that CMP significantly
increased PTGS2 and CHAC1 mRNA levels, but preincubation with Fer-1
obviously reduced (C) PTGS2 and (D) CHAC1 mRNA levels in SKOV3 and
Hey cells. **P<0.01, ***P<0.001 vs. con;
#P<0.05, ##P<0.01,
###P<0.001 vs. CMP; $$P<0.01 vs.
CMP+ZVAD. CMP, carboxymethylated pachyman; Fer-1, ferrostatin-1;
Z-VAD, Z-VAD-fluoromethylketone; PTGS2, prostaglandin-endoperoxide
synthase 2; CHAC1, Chac glutathione specific
γ-glutamylcyclotransferase 1; Con, control. |
CMP upregulated intracellular SOD and
Fe2+ in ovarian cancer cells
DCFH-DA staining showed that both 100 and 200 µg/ml
CMP elevated the intracellular SOD contents compared with those of
the control in both SKOV3 and Hey cells (Fig. 3A and B). The intracellular levels
of SOD, GSH, MDA and Fe2+ were then quantified. The data
showed that 100 and 200 µg/ml CMP enhanced the production of SOD,
MDA and Fe2+ (Fig.
3C-E) but decreased GSH levels in SKOV3 and Hey cells (Fig. 3F).
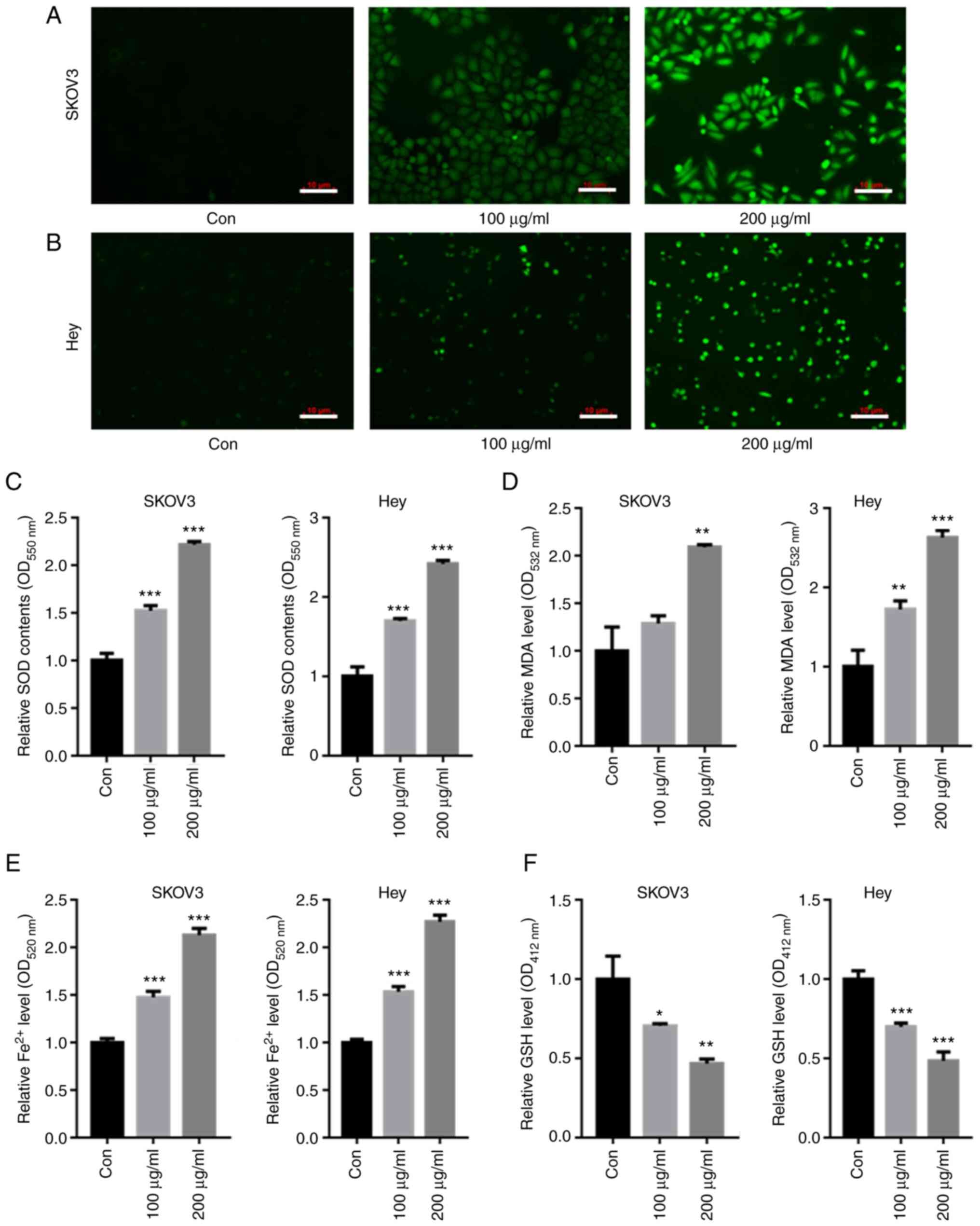 | Figure 3.CMP upregulated intracellular ROS and
Fe2+ in SKOV3 and Hey cells. SKOV3 and Hey cells were
treated with 100 and 200 µg/ml CMP for 24 h. DCFH-DA staining
showed that the intracellular ROS contents were enhanced in both
(A) SKOV3 and (B) Hey cells treated with CMP (scale bar=10 µm). CMP
enhanced the production of (C) SOD, (D) MDA and (E) Fe2+
but decreased the contents of (F) GSH in SKOV3 and Hey cells.
*P<0.05, **P<0.01, ***P<0.001 vs. con. CMP,
carboxymethylated pachyman; ROS, reactive oxygen species; DCFH-DA,
2,7-Dichlorodi-hydrofluorescein diacetate; SOD, superoxide
dismutase; MDA, malondialdehyde; GSH, glutathione; Con,
control. |
CMP induced ovarian cancer cell
ferroptosis by downregulating Nrf2
Nrf2-associated antioxidant stress plays a key role
in ferroptosis inhibition (18).
RT-qPCR analysis showed that CMP reduced Nrf2 and HO-1 mRNA levels
but increased PTGS2 mRNA levels (Fig.
4A-C). In addition, the effects of CMP on the expression of
Nrf2 and corresponding downstream target genes was tested. The data
showed that Nrf2, HO-1, xCT and GPX4 protein levels were decreased
in SKOV3 and Hey cells treated with CMP (Fig. 4D).
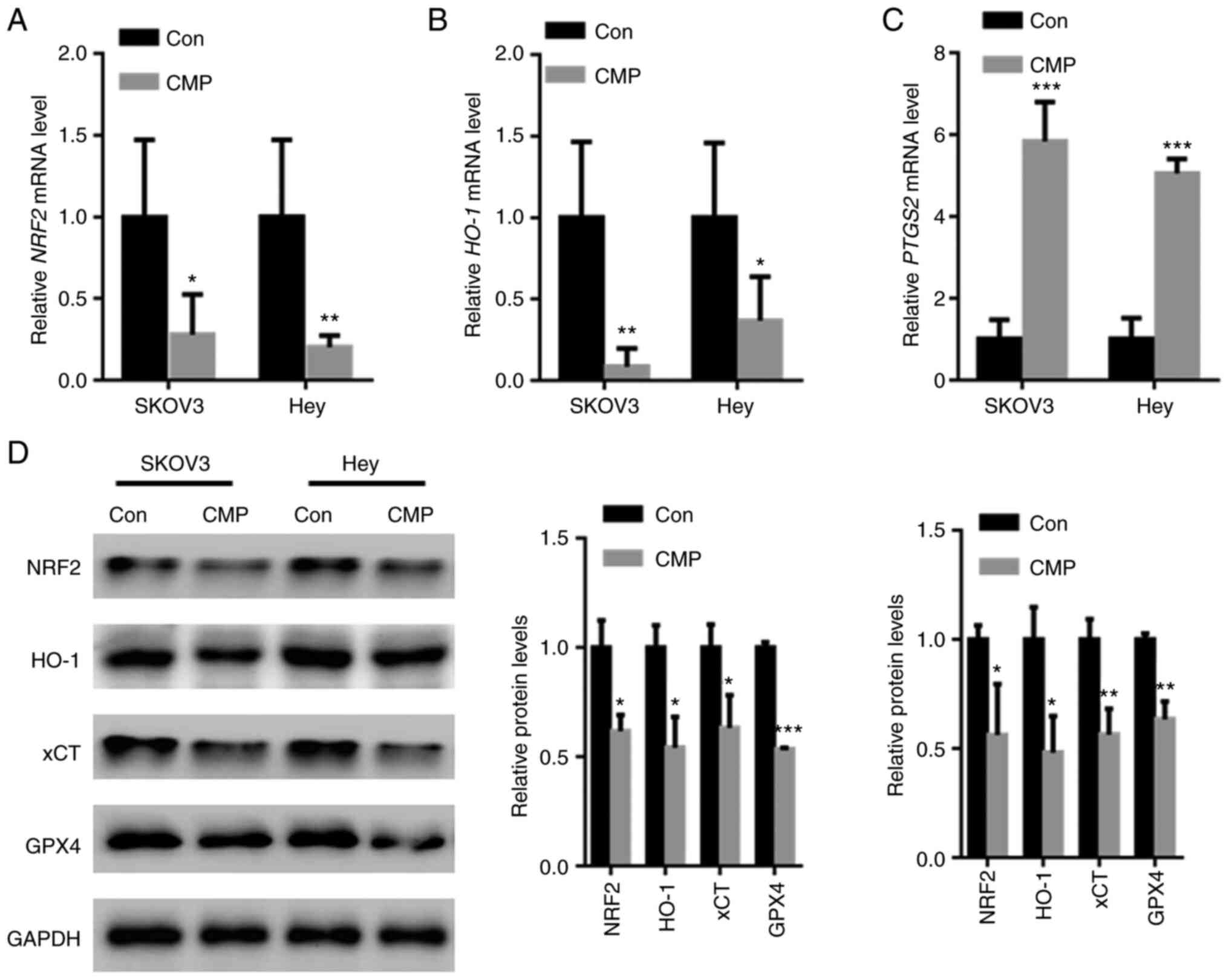 | Figure 4.CMP induced ovarian cancer cell
ferroptosis by downregulating Nrf2. SKOV3 and Hey cells were
treated with 100 µg/ml CMP for 24 h. RT-PCR analysis showed that
CMP reduced (A) Nrf2 and (B) HO-1 mRNA levels but increased the (C)
PTGS2 mRNA levels. (D) CMP decreased the expression of Nrf2, HO-1,
xCT and GPX4 in SKOV3 and Hey cells. *P<0.05, **P<0.01,
***P<0.001 vs. con. CMP, carboxymethylated pachyman; Nrf2,
nuclear factor erythroid 2-related factor 2; HO-1, heme
oxygenase-1; PTGS2, prostaglandin-endoperoxide synthase 2; xCT,
cystine/glutamate antiporter system X(c)(−); Con, control. |
CMP decreased in vivo tumor growth by
suppressing Nrf2-associated ferroptosis
In vivo assays showed that CMP significantly
suppressed tumor volume and weight compared with those of the
control (Fig. 5A-C). The
intracellular contents of Fe2+, MDA, SOD and GSH was
further quantified. The data showed that CMP significantly
increased the accumulation of Fe2+, MDA, and SOD but
reduced the levels of GSH (Fig.
5D-G). In addition, Nrf2, HO-1, xCT and GPX4 protein levels
were suppressed in nude mice treated with CMP compared with those
of the control (Fig. 5H).
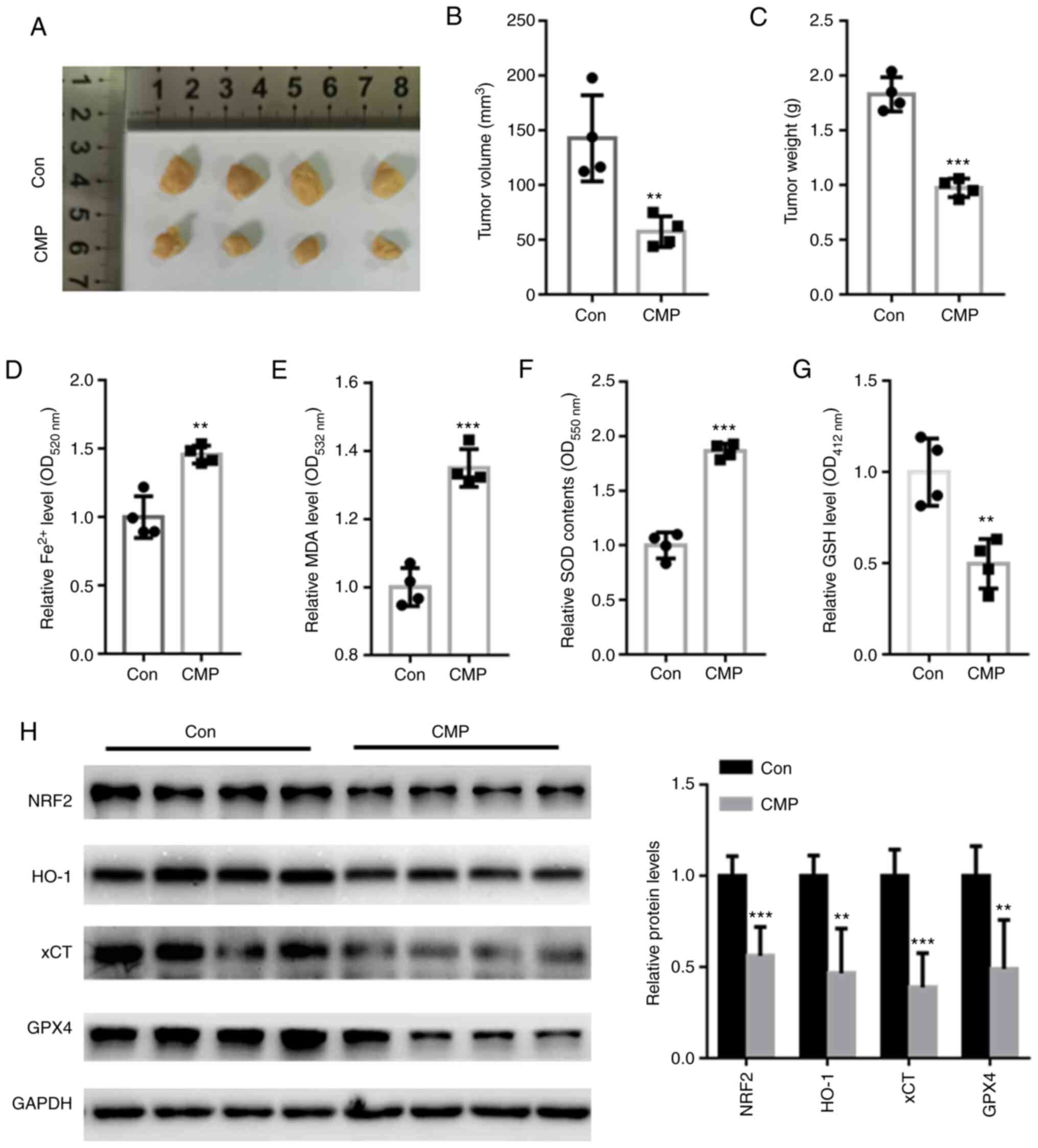 | Figure 5.CMP (50 mg/kg, 1 time/day for 28 days)
decreased in vivo tumor growth by suppressing
NRF2-associated ferroptosis. (A) Representative tumor images. CMP
significantly suppressed tumor (B) volume and (C) weight compared
with those of the control. CMP significantly increased the
accumulation of (D) Fe2+, (E) MDA and (F) SOD but
reduced the levels of (G) GSH. (H) Western blot assay showed that
Nrf2, HO-1, xCT and GPX4 protein levels were suppressed in nude
mice treated with CMP compared with those of the control.
**P<0.01, ***P<0.001 vs. con. CMP, carboxymethylated
pachyman; Nrf2, nuclear factor erythroid 2-related factor 2; MDA,
malondialdehyde; SOD, superoxide dismutase; GSH, glutathione; HO-1,
heme oxygenase-1; xCT, cystine/glutamate antiporter system X(c)(−);
Con, control. |
Discussion
As the leading cause of death from gynecological
malignancy worldwide, >75% of affected women are diagnosed at
advanced stages of ovarian cancer with vague and nonspecific
symptoms (7). Following diagnosis,
the 5-year survival rate of late-stage patients is reported to be
<33% (3). One novel promising
anticancer treatment method is ferroptosis, which is a
well-regulated cell death characterized by lipid peroxidation
(19).
The antitumor effects of CMP have been identified in
colon cancer and hepatocellular carcinoma (15,16).
Consistent with these findings, the present study showed novel data
that CMP significantly suppressed cell survival and induced cell
death in ovarian cancer. To further analyze which form of cell
death could be induced by CMP, ovarian cancer cells were
preincubated with different inhibitors, including apoptosis
inhibitors, ferroptosis inhibitors, necrosis inhibitors and
autophagy inhibitors. The data showed that CMP-induced cell death
could be significantly abolished by Fer-1, a ferroptosis inhibitor.
In addition, CMP-induced upregulation of PTGS2 and CHAC1, two
ferroptosis markers, was significantly decreased by preincubation
with Fer-1. These findings indicated that CMP exhibited potent
anticancer effects on ovarian cancer cells by inducing
ferroptosis.
Upregulation of oxidative stress (ROS) levels is
suggested to be a hallmark of ferroptosis (20). Hence, the effects of CMP on
intracellular ROS production in ovarian cancer cells was evaluated.
The data showed that CMP enhanced the production of ROS. CMP also
enhanced iron levels and MDA contents in SKOV3 and Hey cells. GSH
synthesis is mediated via xCT, which exchanges intracellular
glutamate for extracellular cystine (20). As a major endogenous antioxidant,
GPX4 protects cancer cells from ferroptosis by eliminating lipid
hydroperoxide (20). Consistently,
it was found that CMP decreased GSH in ovarian cancer cells,
indicating that ovarian cancer cells are more vulnerable to
ferroptosis under CMP treatment.
Oxidative damage induced via free radicals promotes
the pathogenesis of multiple diseases, including cancer (21). The transcription factor nuclear
factor Nrf2 renders cells resistant to cellular defense against
toxic and oxidative insults by regulating genes involved in drug
detoxification and antioxidant defense responses (22). For instance, Nrf2-mediated
activation of HO-1 is indicated to protect against oxidative stress
(23). xCT is an important gene
that participates in modulating ‘iron overload-ferroptosis’
(23). Decreased xCT expression
leads to significant oxygen elevation and a reduction in
intracellular antioxidant capacity (23). Silencing of Nrf2 has been shown to
reduce xCT and HO-1 expression, thereby facilitating lipid peroxide
production (23). In addition,
Nrf2 also serves a key role in regulating the antioxidant system by
involving iron metabolism and glutathione synthesis (24). Of note, GPX4, which suppresses the
canonical ferroptosis pathway, is a downstream target gene of Nrf2
(25). Consistent with these
findings, the present study found that CMP significantly suppressed
Nrf2, HO-1, xCT and GPX4 expression in ovarian cancer cells and
tumors. These observations suggested that CMP induced ferroptosis
in ovarian cancer cells by suppressing the Nrf2/HO-1-mediated
ferroptosis pathway.
However, there are limitations to the present study.
First, it will be rewarding to see the results to be verified in
human studies. Second, whether CMP suppresses the development of
ovarian cancer via other molecular mechanisms deserves further
study.
In conclusion, the present study produced novel data
that CMP could induce ferroptotic cell death in ovarian cancer
cells by suppressing Nrf2/HO-1/xCT/GPX4. All these findings
indicated that CMP may have great potential in anti-ovarian cancer
cell therapy by inducing ferroptosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from Tengzhou Central
People's Hospital (grant no. TZH-20190721).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TJ performed the experiments, analyzed the data and
wrote the paper. YG performed part of the RT-qPCR experiments. YW
designed the experiments, analyzed the data and gave final approval
of the version to be published. TJ and YW confirm the authenticity
of all the raw data All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Tengzhou Central People's Hospital (Tengzhou
City, China; approval no. TZH2020AH6).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang Y and Zhu Z: Oridonin inhibits
metastasis of human ovarian cancer cells by suppressing the mTOR
pathway. Arch Med Sci. 15:1017–1027. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang C, Xia BR, Zhang ZC, Zhang YJ, Lou G
and Jin WL: Immunotherapy for ovarian cancer: Adjuvant,
combination, and neoadjuvant. Front Immunol. 11:5778692020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Doubeni CA, Doubeni AR and Myers AE:
Diagnosis and management of ovarian cancer. Am Fam Physician.
93:937–944. 2016.PubMed/NCBI
|
|
4
|
Achatz MI, Caleffi M, Guindalini R,
Marques RM, Nogueira-Rodrigues A and Ashton-Prolla P:
Recommendations for advancing the diagnosis and management of
hereditary breast and ovarian cancer in Brazil. JCO Glob Oncol.
6:439–452. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jafari M, Hasanzadeh M, Solhi E,
Hassanpour S, Shadjou N, Mokhtarzadeh A, Jouyban A and Mahboob S:
Ultrasensitive bioassay of epitope of Mucin-16 protein (CA 125) in
human plasma samples using a novel immunoassay based on silver
conductive nano-ink: A new platform in early stage diagnosis of
ovarian cancer and efficient management. Int J Biol Macromol.
126:1255–1265. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Johnson C and Jazaeri AA: Diagnosis and
management of immune checkpoint inhibitor-related toxicities in
ovarian cancer: A series of case vignettes. Clin Ther. 40:389–394.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun Y, Chen P, Zhai B, Zhang M, Xiang Y,
Fang J, Xu S, Gao Y, Chen X, Sui X and Li G: The emerging role of
ferroptosis in inflammation. Biomed Pharmacother. 127:1101082020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dodson M, Castro-Portuguez R and Zhang DD:
NRF2 plays a critical role in mitigating lipid peroxidation and
ferroptosis. Redox Biol. 23:1011072019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang H, Yuan B, Huang H, Qu S, Yang S and
Zeng Z: Gastrodin induced HO-1 and Nrf2 up-regulation to alleviate
H2O2-induced oxidative stress in mouse liver
sinusoidal endothelial cells through p38 MAPK phosphorylation. Braz
J Med Biol Res. 51:e74392018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan Z, Wirth AK, Chen D, Wruck CJ, Rauh M,
Buchfelder M and Savaskan N: Nrf2-Keap1 pathway promotes cell
proliferation and diminishes ferroptosis. Oncogenesis. 6:e3712017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li B, Nasser MI, Masood M, Adlat S, Huang
Y, Yang B, Luo C and Jiang N: Efficiency of traditional Chinese
medicine targeting the Nrf2/HO-1 signaling pathway. Biomed
Pharmacother. 126:1100742020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen D, Fan Z, Rauh M, Buchfelder M,
Eyupoglu IY and Savaskan N: ATF4 promotes angiogenesis and neuronal
cell death and confers ferroptosis in a xCT-dependent manner.
Oncogene. 36:5593–5608. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee N, Carlisle AE, Peppers A, Park SJ,
Doshi MB, Spears ME and Kim D: xCT-Driven expression of GPX4
determines sensitivity of breast cancer cells to ferroptosis
inducers. Antioxidants (Basel). 10:3172021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang C, Huo X, Gao L, Sun G and Li C:
Hepatoprotective effect of carboxymethyl pachyman in
fluorouracil-treated CT26-bearing mice. Molecules. 22:7562017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Yang S, Gao L, Wang L and Cao L:
Carboxymethyl pachyman (CMP) reduces intestinal mucositis and
regulates the intestinal microflora in 5-fluorouracil-treated CT26
tumour-bearing mice. Food Funct. 9:2695–2704. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qiang Z, Dong H, Xia Y, Chai D, Hu R and
Jiang H: Nrf2 and STAT3 alleviates ferroptosis-mediated IIR-ALI by
regulating SLC7A11. Oxid Med Cell Longev. 2020:51469822020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carbone M and Melino G: Stearoyl Coa
desaturase regulates ferroptosis in ovarian cancer offering new
therapeutic perspectives. Cancer Res. 79:5149–5150. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu J, Xiong Y, Zhang Y, Wen J, Cai N,
Cheng K, Liang H and Zhang W: The molecular mechanisms of
regulating oxidative stress-induced ferroptosis and therapeutic
strategy in tumors. Oxid Med Cell Longev. 2020:88107852020.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oh YS and Jun HS: Effects of glucagon-like
peptide-1 on oxidative stress and Nrf2 Signaling. Int J Mol Sci.
19:262017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bellezza I, Giambanco I, Minelli A and
Donato R: Nrf2-Keap1 signaling in oxidative and reductive stress.
Biochim Biophys Acta Mol Cell Res. 1865:721–733. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong H, Qiang Z, Chai D, Peng J, Xia Y, Hu
R and Jiang H: Nrf2 inhibits ferroptosis and protects against acute
lung injury due to intestinal ischemia reperfusion via regulating
SLC7A11 and HO-1. Aging (Albany NY). 12:12943–12959. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song X and Long D: Nrf2 and Ferroptosis: A
new research direction for neurodegenerative diseases. Front
Neurosci. 14:2672020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deng HF, Yue LX, Wang NN, Zhou YQ, Zhou W,
Liu X, Ni YH, Huang CS, Qiu LZ, Liu H, et al: Mitochondrial iron
overload-mediated inhibition of Nrf2-HO-1/GPX4 Assisted ALI-induced
nephrotoxicity. Front Pharmacol. 11:6245292021. View Article : Google Scholar : PubMed/NCBI
|