Introduction
The 2020 Global Cancer Statistics report published
by the International Agency for Research on Cancer showed that in
2020, female breast cancer (BC) was the leading cause of
cancer-associated death in women worldwide, with 2,261,419 new
cases, accounting for 24.5% of all new cancer cases in women.
Additionally, there were 684,996 deaths, accounting for 15.5% of
all cancer-associated deaths in 2020 (1). Metastatic BC remains a major
contributor to cancer-associated mortality in women. Multiple
reports have shown that 20–30% of BC patients have distant
metastases when first diagnosed (2–4).
Gene expression profiling has had a considerable impact on our
understanding of breast cancer biology. In the past 15 years,
technological progress has revealed the emergence of at least five
different molecular subtypes (Luminal A, Luminal B, HER-2 enriched,
Basal-like and Claudin low) and normal breast like groups, which
are based on gene expression clustering (5). According to the biological
characteristics of the different subtypes, different targeted
therapeutics are used in the clinic. For example, trastuzumab, an
anti-HER2 drug, has significantly changed the therapeutic field of
BC management in the past 20 years (6,7).
However, its concomitant cardiotoxicity and the chances of acquired
drug resistance remain significant challenges that need to be
overcome. Trodelvy is a targeted drug for the treatment of
triple-negative BC, although it is associated with adverse
reactions such as neutropenia, a decrease in white blood cell count
and anaemia (8–11). Cell cycle arrest has been used in
the field of BC treatment with CDK inhibitors such as Palbociclib,
Ribociclip, and Abemaciclib (12).
Since the approval of Palbociclib by the FDA in 2015, CDK4/6
inhibitors have become a first-line treatment option for patients
with metastatic hormone receptor-positive
(HR+)/HER2-negative (HER-2−) BC (13,14).
The use of CDK4/6 inhibitors alone or in combination
with other drugs has also become a research hotspot in recent years
(15–18). Despite significant progress in the
field of chemotherapy-based treatments for BC, as well as
anti-angiogenic therapy, immunotherapy, targeted therapy, and other
emerging therapies, the clinical effects of these approaches remain
unsatisfactory, and the effective rate of immunotherapy is
generally low (19,20). Therefore, there is an urgent need
to identify novel treatment strategies for BC.
Phytochemicals are promising sources for the
development of novel cancer therapeutics, due to their potential
efficiency and low toxicity profiles (21,22),
and they have generally been shown to be promising for the
development of novel agents in the management of numerous diseases
(23,24). Plant phytochemicals represent an
exciting opportunity for improving an individual's general health
through a balanced and appropriate diet, and have also been
considered suitable options for identifying novel therapeutic
agents.
Traditional Chinese Medicines (TCMs) have been used
to treat numerous human diseases in China for thousands of years
(25,26), and have served as a remarkable
source for drug discovery (27).
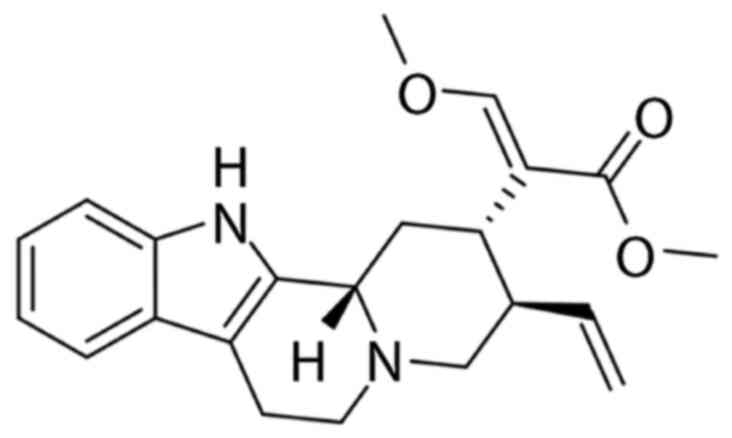
In the present study, the effect of hirsuteine (Fig. 1), an active compound extracted from
the traditional well-known Chinese herb Uncaria
rhynchophylla, on BC was assessed in vivo. Several
studies have shown that hirsuteine possesses a number of
therapeutically relevant properties, especially for treating
central nervous system and cardiovascular disorders, namely,
hypertension, epilepsy, dizziness, convulsion, preeclampsia, and
tremor, amongst others, initially confirming the efficacy and
safety (28–30).
However, the antiproliferative activity and the
underlying mechanisms by which hirsuteine reduces cancer
development and progression have not been determined. Cancer
progression is dependent on its unique ability to escape programmed
cell death (31). Apoptosis is
usually initiated by the death receptor (DR) (extrinsic) or
mitochondrial (intrinsic) axis, and it functions to eliminate
injured cells to maintain homeostasis (32). In both axes, active caspase 3 and 7
cleave poly-ADP ribose polymerase 1 (PARP1) following DNA
degeneration (33). The intrinsic
network is mitochondrial-based apoptosis, which involves cytochrome
c release and activation of caspase-9, which in turn activates
caspase-3. The extrinsic network is dependent on DR stimulation,
which in turn induces the FAS-related death domain (FADD) and
generates the death-induced signal complex, which regulates
downstream caspases-8, −7, −6, and −3 (34,35).
Research on the active ingredients of botanicals
have received increasing attention. Our research group has
primarily studied the TCM Uncaria rhynchophylla, and
hirsuteine is an alkaloid extracted from Uncaria
rhynchophylla, which has a significant inhibitory effect on
several cancer cell lines (36,37).
Our previous study demonstrated that hirsuteine is cytotoxic to
numerous tumor cells in vitro (38). Nevertheless, despite its valuable
properties, little is known regarding its antitumor capacity and
the possible mechanism of hirsuteine in BC. Therefore, determining
the underlying mechanisms by which hirsuteine affects BC cancer
development and progression was assessed in the present study.
The aim of the present study was to examine the
possible anticancer effects of hirsuteine and the molecular
mechanisms underlying its therapeutic efficacy. In particular,
hirsuteine-mediated regulation of MDA-MB-453 cell apoptosis was
assessed. The distribution of cells in the cell cycle and apoptosis
induction were examined in MDA-MB-453 cells.
Materials and methods
Chemicals and reagents
Hirsuteine (ST17300105; 5 mg/dose; purity ≥98%) was
acquired from Shanghai Standard Technology Co., Ltd. The CCK-8
Detection kit, BSA, and Annexin V-FITC Detection kit were acquired
from Beyotime Institute of Biotechnology. B-cell lymphoma-2 (Bcl-2;
ab32124; dilution, 1:1,000), Bcl-2-associated X protein (Bax;
ab32503; dilution, 1:1,000), Cyclin B1 (ab32053; dilution,
1:1,000), CDK1 (ab133327; dilution, 1:1,000), Apaf1 (ab234436;
dilution, 1:1,000), cytochrome C (ab133504; dilution, 1:1,000),
cleaved-caspase3 (ab32042; dilution, 1:1,000), cleaved-caspase9
(ab2324; dilution, 1:1,000), cleaved-PARP1 (ab32064; dilution,
1:1,000), β-actin (ab8227; dilution, 1:5,000) antibodies, goat
anti-rabbit IgG secondary antibody (ab6721; dilution, 1:20,000)
were purchased from Abcam. Other reagents were analytical reagent
grade and from commercial sources.
Cell culture
Human BC MDA-MB-453, MDA-MB-231, and MCF-7 cells
were acquired from American Type Culture Collection. Cells were
cultured using RPMI 1640 or DMEM supplemented with 10% FBS,
penicillin (100 U/ml), and streptomycin (100 µg/ml). Normal human
breast cells Hs 578Bst, human normal lung epithelial cell BEAS-2B,
and normal human hepatocyte THLE-2 cells were obtained from Nanjing
KeyGen Biotech Co., Ltd. and cultured in DMEM supplemented with 10%
FBS, penicillin (100 U/ml), and streptomycin (100 µg/ml) in a
humidified incubator supplied with 5% CO2 at 37°C.
CCK-8 assay
Cells were plated in 96-well culture plates
(1×104 cells/well) overnight and then treated with a
range of hirsuteine concentrations (0, 2.5, 5, 10, 20, 40, or 80
µM) for 24, 48 and 72 h, as described previously (38). For Hs 578Bst, THLE-2, and BEAS-2B
cells, assay protocols were as above, with cells being treated for
48 h with various concentrations of hirsuteine (0, 2.5, 5, 10, 20,
40, or 80 µM) prior to addition of the CCK-8 reagent. The 50%
growth inhibition (IC50) value was determined using a
hirsuteine survival concentration curve. All experiments were
performed at least three times.
Colony formation assay
Cells were plated in 6-well culture plates at
2–5×103 cells per well for 24 h. Cells were then exposed
to varying hirsuteine concentrations (0, 5, 10, or 25 µM) for 48 h,
after which the hirsuteine-containing media was removed, and fresh
media was added. The cells were then cultured for 14 days in
supplemented media. Finally, cells were fixed in methanol at room
temperature for 15 min, dyed with 1% crystal violet solution at
room temperature for 10 min, washed with PBS three times, dried in
the room and observed under a microscope, with images captured. The
number of colonies consisting of ≥50 cells was counted.
Apoptosis staining
Cells were exposed to varying concentrations of
hirsuteine (0, 5, 10, or 25 µM) for 48 h and apoptosis was measured
as described previously (38).
Cell cycle analysis
Briefly, a six-well plate was seeded with
3×105 cells per well. After incubation, the media was
replaced with supplemented media containing a range of hirsuteine
concentrations (0, 5, 10, or 25 µM) for an additional 48 h. Cells
were then detached using 0.25% trypsin, and centrifuged at 4°C for
5 min at 300 × g, washed once with PBS, and fixed overnight at 4°C
with 70% chilled ethanol. Subsequently, the cells were centrifuged
again as above, fully suspending the cells after treating with 100
µl RNase A (50 µg/ml) at 37°C in a water bath for 30 min, prior to
a 30 min staining at 4°C in the dark with 400 µl PI (50 µg/ml). All
samples were evaluated using a FACScan flow cytometer (BD
Biosciences). Data were analyzed using CellQuest Pro software,
version 5.1 (BD Biosciences). All experiments were performed three
times independently.
Western blotting
Following 48 h of treatment with hirsuteine (0, 5,
10, or 25 µM), chilled RIPA buffer was used for cell lysis for 30
min. Extracted proteins were resolved and transferred as described
previously (38). The membranes
were blocked for 1 h at 37°C with 5% milk in Tris-buffered saline
(TBS) containing 0.05% Tween-20 (TBST) and then incubated with
cleaved caspase-3, cleaved caspase-9, cleaved CARP, Bax, Bcl-2,
Apaf1, cytochrome c, CDK1, Cyclin B1 and β-actin (Abcam)
antibodies at 4°C overnight. Membranes were then washed using TBST
and incubated with the goat anti-rabbit IgG secondary antibody
(Abcam) at room temperature for 2 h. Immunoblotted proteins were
analyzed with the ChemiDoc XRS imaging system and QuantityOne
software (Version 4.6.9; Bio-Rad Laboratories, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Cells were treated with different concentrations of
hirsuteine for 48 h. Total RNA was isolated from cells using
TRIzol® reagent followed by reverse transcription to
cDNA using a QuantiTect Reverse Transcription kit, according to the
manufacturer's protocol. Subsequently, qPCR was performed using
ChamQ™ SYBR qPCR MasterMix in triplicate according to the
manufacturer's protocol. Each reaction was conducted in duplicate
using the following thermocycling conditions: Initial denaturation
for 10 min at 95°C, followed by 40 cycles of denaturation for 30
sec at 95°C, annealing for 30 sec at 60°C and extension for 20 sec
at 72°C. The sequences of the primers were: Bax forward,
5′-AAGAAGCTGAGCGAGTGTCT-3′ and reverse, 5′-GTTCTGATCAGTTCCGGCAC-3′;
Bcl-2 forward, 5′-GCCTTCTTTGAGTTCGGTGG-3′ and reverse,
5′-GAAATCAAACAGAGGCCGCA-3′; caspase-3 forward,
5′-ACTGGACTGTGGCATTGAGA-3′ and reverse, 5′-GCACAAAGCGACTGGATGAA-3′;
caspase-9 forward, 5′-ACATGCTGGCTTCGTTTCTG-3′ and reverse,
5′-TCTCAAGAGCACCGACATCA-3′; and GAPDH forward,
5′-TCAAGAAGGTGGTGAAGCAGG-3′ and reverse,
5′-TCAAAGGTGGAGGAGTGGGT-3′. GAPDH was used as the loading control.
Expression was quantified using the 2−∆∆Cq method
(39).
Statistical analysis
Data are presented as the mean ± SD from three
independent repeats. Statistical comparisons were performed by
one-way analysis of variance, followed by Bonferroni's test.
P<0.05 was considered to indicate a statistically significant
difference. Data were analyzed using GraphPad Prism version 6.0
(GraphPad Software Inc.).
Results
Cytotoxicity of hirsuteine against
human BC MDA-MB-453 cells
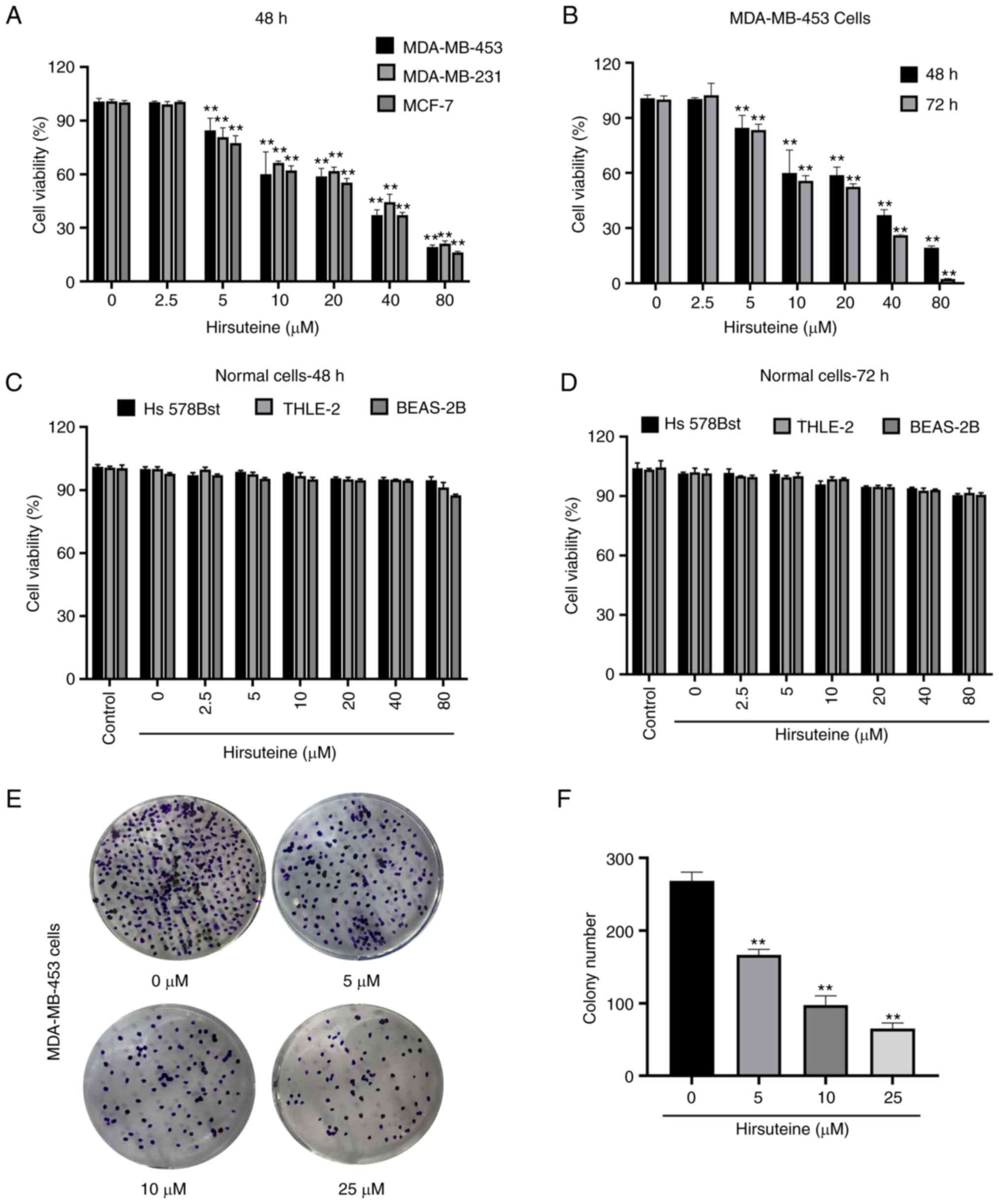
Hirsuteine-mediated regulation of MDA-MB-453,
MDA-MB-231, and MCF-7 cell viability was assessed using a CCK-8
assay (Fig. 2A). Hirsuteine
lowered MDA-MB-453 cell viability in a concentration and
time-dependent manner (Fig. 2B).
Next, we explored the impact of hirsuteine on the proliferation of
Hs 578Bst, BEAS-2B, and human hepatocyte THLE-2 cells using a CCK-8
assay. The results showed that the cell viability after hirsuteine
treatment in normal cells was >80%, which was considered
non-toxic for the non-cancerous cell lines after 48 and 72 h of
treatment at doses up to and including 80 µM (Fig. 2C and D). As indicated in Fig. 2E and F, hirsuteine markedly reduced
colony formation in a concentration-dependent manner. Based on the
results, hirsuteine inhibited MDA-MB-453 cell proliferation and
lowered MDA-MB-453 colony formation activity, whilst not exerting a
notable effect on healthy human cell lines at the same doses.
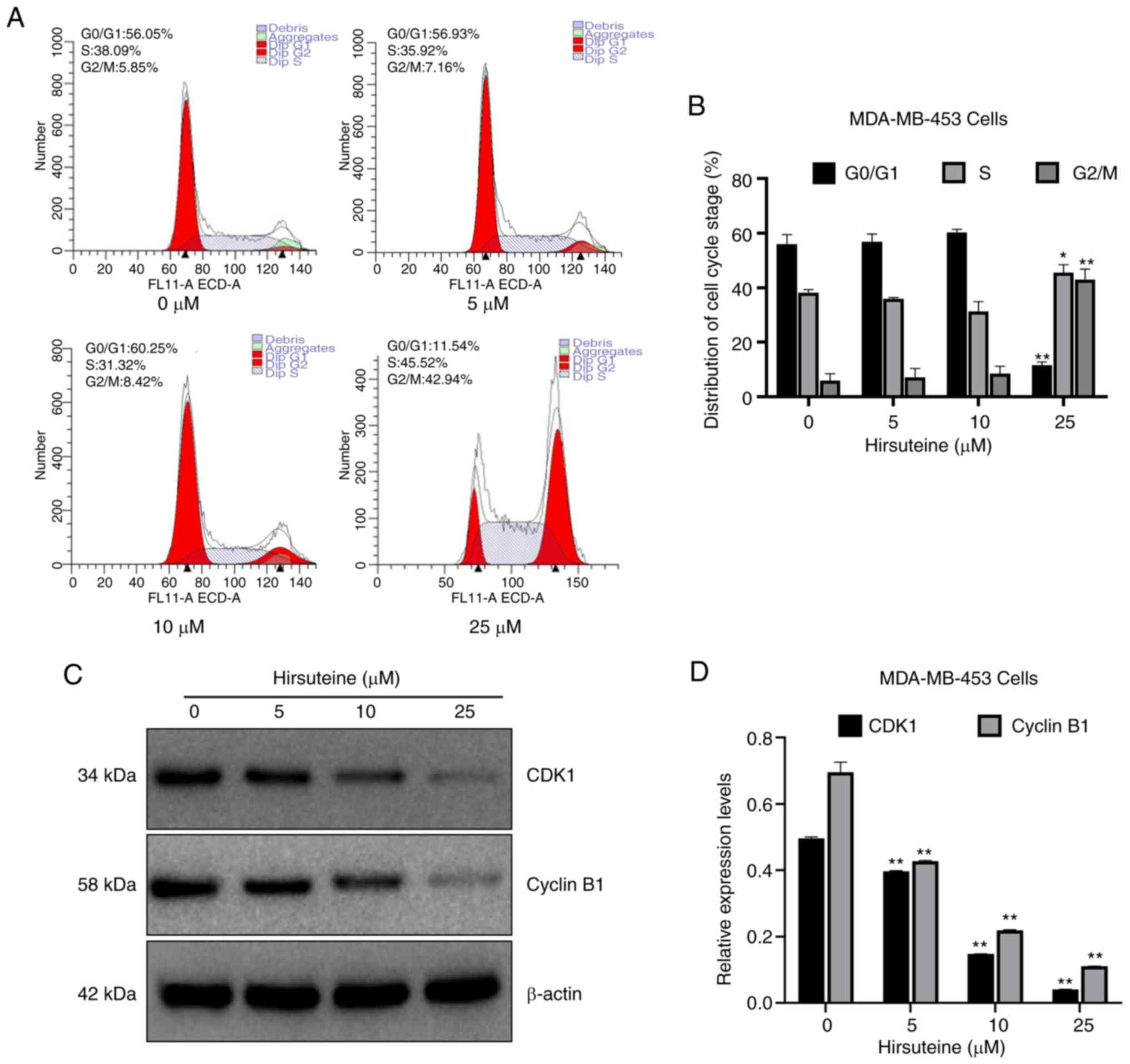
Hirsuteine inhibits cell cycle
progression
Cell-cycle distribution was assessed using flow
cytometry. Apoptosis can be characterized by DNA fragmentation and
damage. There are multiple phases and subphases in the cell cycle,
namely, G0/G1 (DNA pre-synthesis and stationary); S (DNA
synthesis); and G2/M (DNA post-synthesis and mitosis) (40–42).
To elucidate whether hirsuteine altered cell cycle distribution,
the number of cells in each phase of the cell cycle was evaluated
following hirsuteine treatment. Based on the results, the
proportion of cells in the G0/G1 phase was reduced from 56.1±3.4 to
11.5±1.3%, while the proportion of cells in the G2/M phase
increased from 5.9±2.6 to 42.9±3.9% significantly, and the
proportion of cells in the S phase increased from 38.1±1.2 to
45.5±2.9% following hirsuteine treatment (Fig. 3A). The proportion of cells in each
phase of the cycle were compared (Fig.
3B). To determine the potential mechanism by which cell cycle
arrest was induced by hirsuteine, western blotting was used to
determine the expression levels of cell cycle-associated proteins
in hirsuteine treated MDA-MB-453 cells. A marked decrease was
observed in CDK1 and Cyclin B1 protein levels, which may underlie
the G2/M phase arrest induced by hirsuteine (Fig. 3C and D). Thus, hirsuteine promoted
the progression of BC cells from the S phase to the G2/M phase,
prior to cell cycle arrest in G2/M phase, resulting in a reduction
in proliferative ability and cell viability.
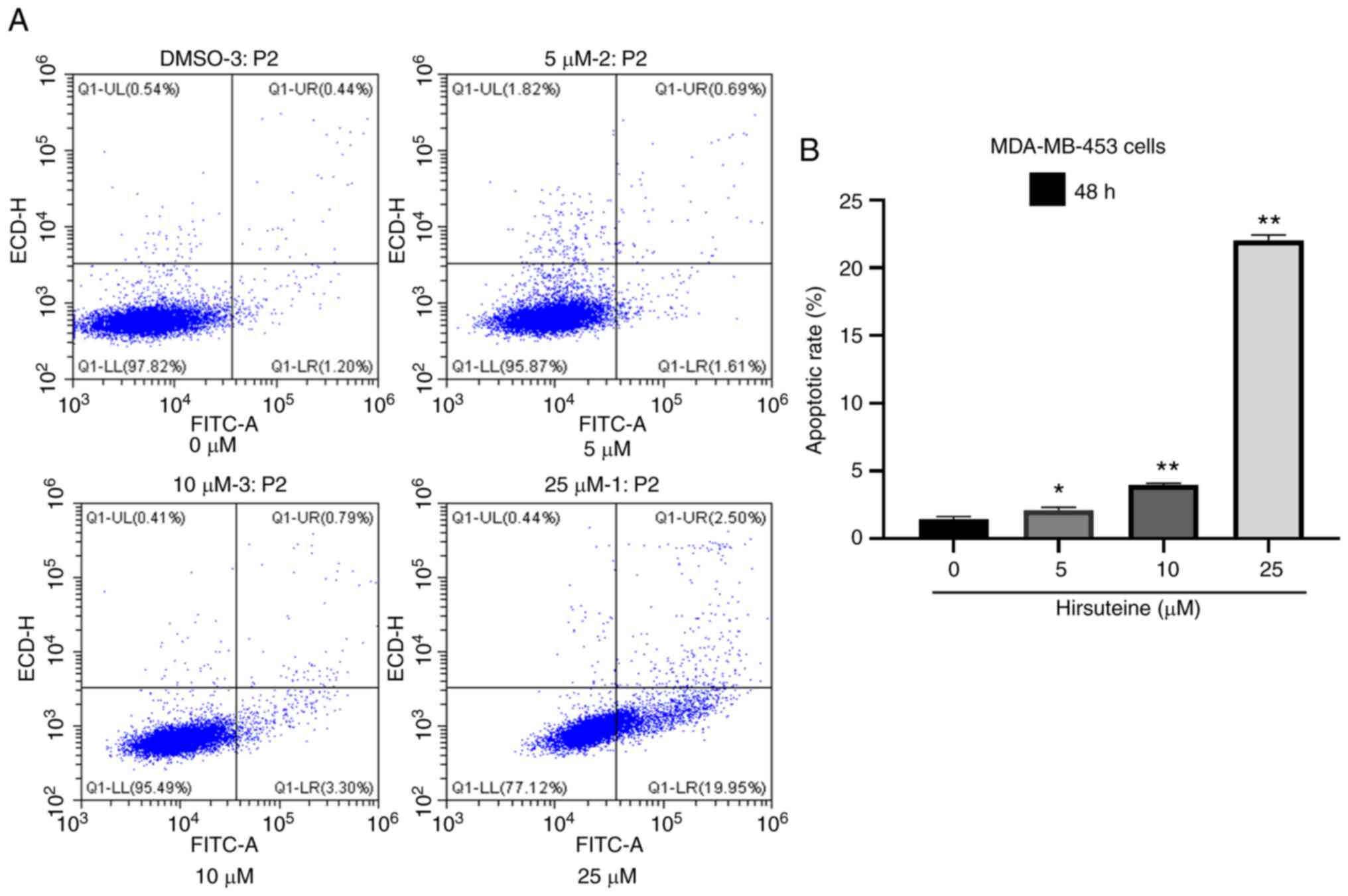
Hirsuteine induces MDA-MB-453 cell
apoptosis
Hirsuteine-treated (0, 5, 10, and 25 µM) MDA-MB-453
cell apoptosis was quantified using flow cytometry using Annexin V
labeling and PI exclusion staining. A four-quadrant schematic was
used to classify the following cell states: Mechanical, necrotic,
normal, early apoptotic, and late apoptotic. Relative to the
control cells, the quantity of healthy cells decreased from
97.7±0.2 to 77.5±0.4%, and the quantity of early and late apoptotic
cells increased from 1.4±0.2 to 22.1±0.4% with increasing
hirsuteine concentrations (Fig.
4A). We also recorded the number of apoptotic cells in the
control and treated groups (Fig.
4B). Thus, hirsuteine suppressed MDA-MB-453 cell proliferation
whilst inducing apoptosis.
Hirsuteine regulates apoptosis-related
proteins
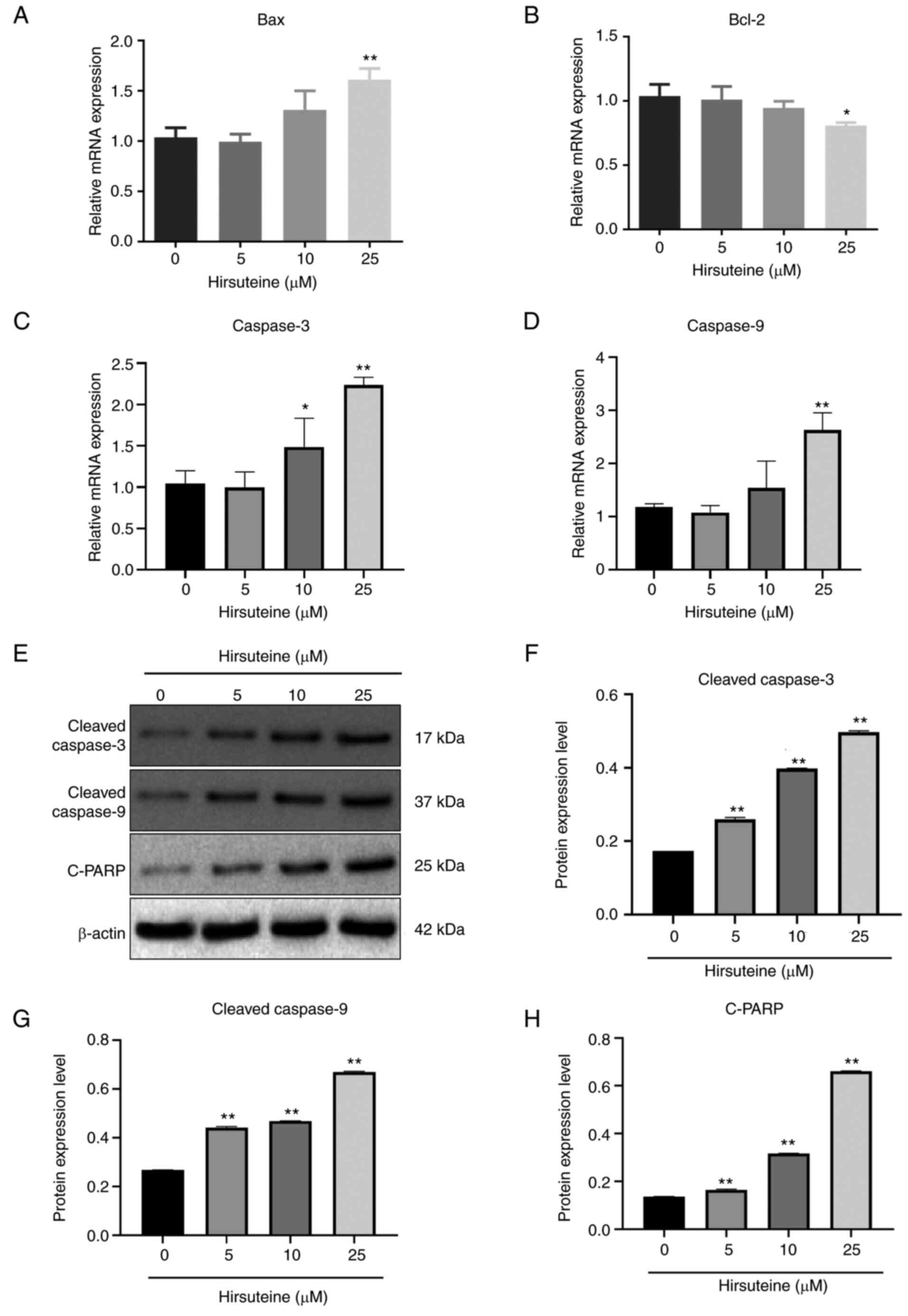
Bax, Bcl-2, cleaved caspase-3, and cleaved caspase-9
expression levels were assessed following 48 h of treatment with
hirsuteine using qPCR and western blotting. Following treatment
with different hirsuteine concentrations for 48 h, the RT-qPCR
results showed that in the MDA-MB-453 cells, Bax,
cleaved-caspase-3, and cleaved caspase-9 mRNA levels were promoted
and Bcl-2 expression was decreased (Fig. 5A-D), resulting in an increase in
pro-apoptotic/anti-apoptotic protein ratio. After 48 h of treatment
with different doses of hirsuteine, the western blot results showed
that hirsuteine increased the levels of cleaved caspase-3, cleaved
caspase-9, and cleaved-PARP levels in the MDA-MB-453 cells
(Fig. 5E). Additionally,
densitometry analysis of the protein expression levels was
performed, and the results are shown in Fig. 5F-H. These results showed that
hirsuteine induced apoptosis in MDA-MB-453 cells via the intrinsic
apoptotic pathway.
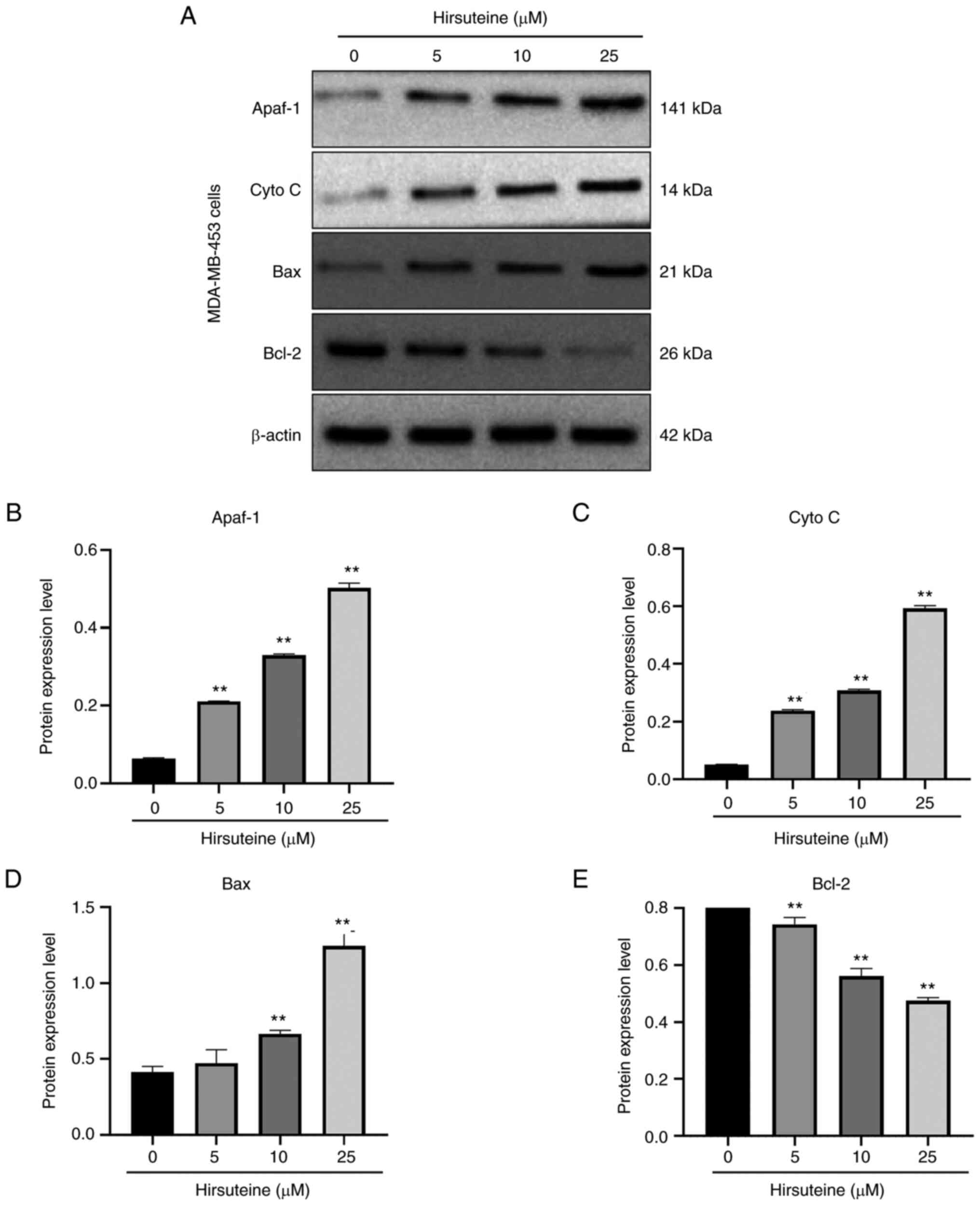
Hirsuteine induces apoptosis in
MDA-MB-453 cells via the Bcl-2/Bax axis
The Bcl-2 family members induce apoptosis via the
mitochondrial network (43). To
examine the hirsuteine-mediated regulation of cell apoptosis, we
assessed the expression levels of several apoptotic proteins. After
48 h of treatment, with hirsuteine (0, 5, 10, or 25 µM), increased
expression of the proapoptotic Bax, Apaf-1, and cytoplasmic
cytochrome-c levels, and reduced levels of Bcl-2 were observed,
resulting in an imbalance in the Bax/Bcl-2 ratio relative to
control cells in a dose-dependent manner (Fig. 6A). Furthermore, densitometry
analysis of the protein expression levels was performed, and the
results are shown in Fig. 6B-E.
Together, these results demonstrated that hirsuteine activated the
Bcl-2/Bax signaling pathway, resulting in apoptosis in MDA-MB-453
cells.
Discussion
At present, chemotherapy is the primary method of
treatment of BC, and it can lead to several side effects (44). The common adverse reactions of
traditional chemotherapeutic drugs include gastrointestinal
reactions, bone marrow toxicity, hepatorenal toxicity,
cardiotoxicity, neurotoxicity, and systemic reactions (45). The adverse reactions caused by
immune checkpoint inhibitors differ from traditional
chemotherapy-related adverse effects. Immune-related adverse events
(IRAEs) caused by immune checkpoint inhibitors usually include
endocrine dysfunction, skin toxicity, gastrointestinal adverse
reactions, and a small number of cases of liver toxicity (46,47).
The incidence of serious IRAEs is low, but if it exceeds
expectations and cannot be appropriately handled, it may endanger a
patient's life (46). Therefore,
increasing attention is being paid to the discovery of safe and
more effective novel compounds for the treatment of patients with
BC.
Phytochemicals are promising compounds that have
been used for several years to develop cancer medications owing to
their potential efficacy and reduced toxicity profiles (21,22).
Multiple reports have suggested that a majority of Uncaria
rhynchophylla-mediated biological properties can be attributed
to the alkaloid constituents (48,49).
Hirsuteine may serve as a novel and specific SPHK1 inhibitor,
exerting anti-leukemic activity by inhibiting the SPHK1/S1P/S1PR1
and BCR-ABL/PI3K/Akt pathways in CML cells (36). Similarly, we previously reported
that the oxindole alkaloid purified from Uncaria
rhynchophylla could effectively suppress the survival of Jurkat
Clone E6-1 cells (T-cell leukemia cells) (38).
In the present study, CCK-8 assays of the effect of
hirsuteine on MDA-MB-231 and MCF-7 breast cancer cells were used to
determine the safe concentration range of hirsuteine. For all
subsequent experiments, 25 µM was used as the upper limit as the
inhibition rate of hirsuteine on the proliferation of MDA-MB-453
cells was about 45% at this concentration. However, in future
studies, 50% IC50 dose, IC50 dose, and 2×
IC50 dose will be used. In this study, it was shown that
hirsuteine exhibited cytotoxicity against cancer cells in a
concentration-dependent manner (MDA-MB-453) without affecting
normal cells (Hs 578Bst, THLE-2 and BEAS-2B), which corroborates
earlier investigations which showed that Uncaria
rhynchophylla inhibited the proliferation of numerous tumor
cell lines (50–52). The results of flow cytometry also
revealed the effectiveness of hirsuteine on human MDA-MB-453 BC
cells, indicating the underlying mechanisms by which hirsuteine
exerts its anti-cancer-specific effects.
Cell cycle arrest is a promising approach for
inhibiting tumor development, particularly since multiple studies
demonstrated notable cell cycle dysregulation in several types of
cancer (53–56). Based on the results of the present
study, hirsuteine-treated MDA-MB-453 cells exhibited
G2/M phase cell cycle arrest and displayed enhanced
cleaved caspase 3 levels, indicative of an increase in the cell
cycle following hirsuteine treatment. It was previously reported
that CDK1 modulates cell cycle progression via interaction with
cyclins B1 and A2 (57).
There is increasing evidence that numerous viral
proteins can cause host G2/M cell cycle arrest. The
G2/M arrest induced by some viral proteins is linked to
the inhibition of cyclin B1-CDK1 kinase activity, inactivating the
cyclin B1-CDK1 complex, which in turn prevents premature entry into
the M phase (58,59). Hence, Cyclin B1 is responsible for
entry into the M phase (60,61).
Upon entry into prophase, Cyclin B1-CDK1 activity gradually
increases (62), which activates
the Cyclin B1-CDK1 complex, resulting in its translocation to the
nucleus to promote cell division (63). Based on the western blot analysis
of the aforementioned study, there was a strong association between
decreased CDK1 and cyclin B1 levels with suppressed MDA-MB-453 cell
proliferation following hirsuteine treatment, which eventually
resulted in cell cycle arrest in the G2/M phase.
Apoptosis activation occurs in one of two methods:
i) the intrinsic axis involves mitochondrial cytochrome c release,
which induces various downstream caspases, and ii) the extrinsic
axis which involves activation of the Fas death receptor via an
external stimulus (64). The
intrinsic apoptotic axis does not require the activation of a
membrane receptor. In fact, the signal is generated in the
mitochondria itself, and this axis is critical for drug-activated
apoptosis, whereas the extrinsic apoptotic axis involves
membrane-bound death receptors that belong to the TNF gene
superfamily (65,66). Caspase-9 (intrinsic axis) and
caspase-8 (extrinsic axis) activate caspase-3 (67–70).
Caspase-9 initiates the apoptotic process by promoting the
formation of the apoptosome complex within the mitochondrial
network. Caspase-3 cleaves both PARP and DNA, which are hallmarks
of the apoptotic process. Upon release of mitochondrial cytochrome
c, it interacts with the adaptor protein Apaf-1 (71). Herein, hirsuteine substantially
augmented Bax, PARP1, caspase-9, and caspase-3 expression,
indicating that the mitochondria-based apoptotic network
contributed to the hirsuteine-driven increase in MDA-MB-453 cell
apoptosis. Collectively, the results of the present study showed
that the intrinsic apoptotic axis was stimulated by hirsuteine in
MDA-MB-453 cells.
In conclusion, this study revealed that hirsuteine
strongly suppressed human BC cell development via activation of the
caspase-3-based apoptotic network. These results highlight a novel
target for the management of BC, as well as a promising agent for
further assessment.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BY, YY and JM designed the present study. JM and YY
performed the experiments. JM and YL analyzed the data. JM drafted
the initial manuscript. BY and YY revised the initial manuscript.
JM and BY confirm the authenticity of all the raw data. BY and YY
helped guide JM in the whole process. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kennecke H, Yerushalmi R, Woods R, Cheang
MC, Voduc D, Speers CH, Nielsen TO and Gelmon K: Metastatic
behavior of breast cancer subtypes. J Clin Oncol. 28:3271–3277.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eckhardt BL, Francis PA, Parker BS and
Anderson RL: Strategies for the discovery and development of
therapies for metastatic breast cancer. Nat Rev Drug Discov.
11:479–497. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Redig AJ and McAllister SS: Breast cancer
as a systemic disease: A view of metastasis. J Intern Med.
274:113–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Perou CM, Sorlie T, Eisen MB, van de Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tinoco G, Warsch S, Gluck S, Avancha K and
Montero AJ: Treating breast cancer in the 21st century: Emerging
biological therapies. J Cancer. 4:117–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pinto AC, Ades F, de Azambuja E and
Piccart-Gebhart M: Trastuzumab for patients with HER2 positive
breast cancer: Delivery, duration and combination therapies.
Breast. 22 (Suppl 2):S152–S155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bardia A, Hurvitz SA, Tolaney SM, Loirat
D, Punie K, Oliveira M, Brufsky A, Sardesai SD, Kalinsky K, Zelnak
AB, et al: Sacituzumab govitecan in metastatic triple-negative
breast cancer. N Engl J Med. 384:1529–1541. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dean AQ, Luo S, Twomey JD and Zhang B:
Targeting cancer with antibody-drug conjugates: Promises and
challenges. MAbs. 13:19514272021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zaman S, Jadid H, Denson AC and Gray JE:
Targeting Trop-2 in solid tumors: Future prospects. Onco Targets
Ther. 12:1781–1790. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Syed YY: Sacituzumab govitecan: First
approval. Drugs. 80:1019–1025. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
O'Leary B, Finn RS and Turner NC: Treating
cancer with selective CDK4/6 inhibitors. Nat Rev Clin Oncol.
13:417–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Piezzo M, Cocco S, Caputo R, Cianniello D,
Gioia GD, Lauro VD, Fusco G, Martinelli C, Nuzzo F, Pensabene M and
Laurentiis MD: Targeting cell cycle in breast cancer: CDK4/6
Inhibitors. Int J Mol Sci. 21:64792020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Spring LM, Wander SA, Zangardi M and
Bardia A: CDK 4/6 inhibitors in breast cancer: Current
controversies and future directions. Curr Oncol Rep. 21:252019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Spring LM, Wander SA, Andre F, Moy B,
Turner NC and Bardia A: Cyclin-dependent kinase 4 and 6 inhibitors
for hormone receptor-positive breast cancer: Past, present, and
future. Lancet. 395:817–827. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Finn RS, Crown JP, Lang I, Boer K,
Bondarenko IM, Kulyk SO, Ettl J, Patel R, Pinter T, Schmidt M, et
al: The cyclin-dependent kinase 4/6 inhibitor palbociclib in
combination with letrozole versus letrozole alone as first-line
treatment of oestrogen receptor-positive, HER2-negative, advanced
breast cancer (PALOMA-1/TRIO-18): A randomised phase 2 study.
Lancet Oncol. 16:25–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Corona SP and Generali D: Abemaciclib: A
CDK4/6 inhibitor for the treatment of
HR+/HER2− advanced breast cancer. Drug Des
Devel Ther. 12:321–330. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Johnston SRD, Harbeck N, Hegg R, Toi M,
Martin M, Shao ZM, Zhang QY, Rodriguez JLM, Campone M, Hamilton E,
et al: Abemaciclib combined with endocrine therapy for the adjuvant
treatment of HR+, HER2-, node-positive, high-risk, early breast
cancer (monarchE). J Clin Oncol. 38:3987–3998. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Richardson JL, Marks G and Levine A: The
influence of symptoms of disease and side effects of treatment on
compliance with cancer therapy. J Clin Oncol. 6:1746–1752. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Coates A, Abraham S, Kaye SB, Sowerbutts
T, Frewin C, Fox RM and Tattersall MH: On the receiving end-patient
perception of the side-effects of cancer chemotherapy. Eur J Cancer
Clin Oncol. 19:203–208. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chirumbolo S: Plant phytochemicals as new
potential drugs for immune disorders and cancer therapy: Really a
promising path? J Sci Food Agric. 92:1573–1577. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thakur VS, Deb G, Babcook MA and Gupta S:
Plant phytochemicals as epigenetic modulators: Role in cancer
chemoprevention. AAPS J. 16:151–163. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang YJ, Gan RY, Li S, Zhou Y, Li AN, Xu
DP and Li HB: Antioxidant phytochemicals for the prevention and
treatment of chronic diseases. Molecules. 20:21138–21156. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Howes MJ and Simmonds MS: The role of
phytochemicals as micronutrients in health and disease. Curr Opin
Clin Nutr Metab Care. 17:558–566. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Z, Feiyue Z and Gaofeng L: Traditional
chinese medicine and lung cancer-from theory to practice. Biomed
Pharmacother. 137:1113812021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li S, Wu Z and Le W: Traditional chinese
medicine for dementia. Alzheimers Dement. 17:1066–1071. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ndagijimana A, Wang X, Pan G, Zhang F,
Feng H and Olaleye O: A review on indole alkaloids isolated from
Uncaria rhynchophylla and their pharmacological studies.
Fitoterapia. 86:35–47. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakazawa T, Banba K, Hata K, Nihei Y,
Hoshikawa A and Ohsawa K: Metabolites of hirsuteine and hirsutine,
the major indole alkaloids of Uncaria rhynchophylla, in rats. Biol
Pharm Bull. 29:1671–1677. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Horie S, Yano S, Aimi N, Sakai S and
Watanabe K: Effects of hirsutine, an antihypertensive indole
alkaloid from Uncaria rhynchophylla, on intracellular calcium in
rat thoracic aorta. Life Sci. 50:491–498. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Reed JC: Dysregulation of apoptosis in
cancer. J Clin Oncol. 17:2941–2953. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sola S, Morgado AL and Rodrigues CM: Death
receptors and mitochondria: Two prime triggers of neural apoptosis
and differentiation. Biochim Biophys Acta. 1830:2160–2166. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Isabelle M, Moreel X, Gagne JP, Rouleau M,
Ethier C, Gagne P, Hendzel MJ and Poirier GG: Investigation of
PARP-1, PARP-2, and PARG interactomes by affinity-purification mass
spectrometry. Proteome Sci. 8:222010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Debatin KM: Apoptosis pathways in cancer
and cancer therapy. Cancer Immunol Immunother. 53:153–159. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao S, Guo T, Luo S, Zhang Y, Ren Z, Lang
X, Hu G, Zuo D, Jia W, Kong D, et al: Growth inhibitory and
pro-apoptotic effects of hirsuteine in chronic myeloid leukemia
cells through targeting sphingosine kinase 1. Biomol Ther (Seoul).
30:553–561. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang BY, Zeng Y, Li YJ, Huang XJ, Hu N,
Yao N, Chen MF, Yang ZG, Chen ZS, Zhang DM and Zeng CQ: Uncaria
alkaloids reverse ABCB1-mediated cancer multidrug resistance. Int J
Oncol. 51:257–268. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Meng J, Su R, Wang L, Yuan B and Li L:
Inhibitory effect and mechanism of action (MOA) of hirsutine on the
proliferation of T-cell leukemia Jurkat clone E6-1 cells. PeerJ.
9:e106922021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Otsuki L and Brand AH: Cell cycle
heterogeneity directs the timing of neural stem cell activation
from quiescence. Science. 360:99–102. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Poncelet L, Garigliany M, Ando K, Franssen
M, Desmecht D and Brion JP: Cell cycle S phase markers are
expressed in cerebral neuron nuclei of cats infected by the Feline
Panleukopenia virus. Cell Cycle. 15:3482–3489. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yamada T, Das Gupta TK and Beattie CW:
p28-mediated activation of p53 in G2-M phase of the cell cycle
enhances the efficacy of DNA damaging and antimitotic chemotherapy.
Cancer Res. 76:2354–2365. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Handrick R, Ganswindt U, Faltin H, Goecke
B, Daniel PT, Budach W, Belka C and Jendrossek V: Combined action
of celecoxib and ionizing radiation in prostate cancer cells is
independent of pro-apoptotic Bax. Radiother Oncol. 90:413–421.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Odle TG: Adverse effects of breast cancer
treatment. Radiol Technol. 85:297M–319M. 2014.PubMed/NCBI
|
|
45
|
Chopra D, Rehan HS, Sharma V and Mishra R:
Chemotherapy-induced adverse drug reactions in oncology patients: A
prospective observational survey. Indian J Med Paediatr Oncol.
37:42–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mina LA, Lim S, Bahadur SW and Firoz AT:
Immunotherapy for the treatment of breast cancer: Emerging new
data. Breast Cancer (Dove Med Press). 11:321–328. 2019.PubMed/NCBI
|
|
47
|
Brahmer JR, Lacchetti C, Schneider BJ,
Atkins MB, Brassil KJ, Caterino JM, Chau I, Ernstoff MS, Gardner
JM, Ginex P, et al: Management of immune-related adverse events in
patients treated with immune checkpoint inhibitor therapy: American
society of clinical oncology clinical practice guideline. J Clin
Oncol. 36:1714–1768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang W, Ip SP, Liu L, Xian YF and Lin ZX:
Uncaria rhynchophylla and its major constituents on central nervous
system: A review on their pharmacological actions. Curr Vasc
Pharmacol. 18:346–357. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lee J, Son D, Lee P, Kim SY, Kim H, Kim CJ
and Lim E: Alkaloid fraction of Uncaria rhynchophylla protects
against N-methyl-D-aspartate-induced apoptosis in rat hippocampal
slices. Neurosci Lett. 348:51–55. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen XX, Leung GP, Zhang ZJ, Xiao JB, Lao
LX, Feng F, Mak JCW, Wang Y, Sze SCW and Zhang KYB:
Proanthocyanidins from Uncaria rhynchophylla induced apoptosis in
MDA-MB-231 breast cancer cells while enhancing cytotoxic effects of
5-fluorouracil. Food Chem Toxicol. 107:248–260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lee JS, Kim J, Kim BY, Lee HS, Ahn JS and
Chang YS: Inhibition of phospholipase cgamma1 and cancer cell
proliferation by triterpene esters from Uncaria rhynchophylla. J
Nat Prod. 63:753–756. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang R, Li G, Zhang Q, Tang Q, Huang J,
Hu C, Liu Y, Wang Q, Liu W, Gao N and Zhou S: Hirsutine induces
mPTP-dependent apoptosis through ROCK1/PTEN/PI3K/GSK3β pathway in
human lung cancer cells. Cell Death Dis. 9:5982018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Molinari M: Cell cycle checkpoints and
their inactivation in human cancer. Cell Prolif. 33:261–274. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu G, Chen G, Zhou J, Zhu H, Chu J and
Zhang F: Liriodenine enhances radiosensitivity in esophageal cancer
ECA-109 cells by inducing apoptosis and G2/M arrest. Oncol Lett.
16:5020–5026. 2018.PubMed/NCBI
|
|
55
|
Maharjan S, Kwon YS, Lee MG, Lee KS and
Nam KS: Cell cycle arrest-mediated cell death by morin in
MDA-MB-231 triple-negative breast cancer cells. Pharmacol Rep.
73:1315–1327. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Han YH, Mun JG, Jeon HD, Kee JY and Hong
SH: Betulin inhibits lung metastasis by inducing cell cycle arrest,
autophagy, and apoptosis of metastatic colorectal cancer cells.
Nutrients. 12:662019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gong D and Ferrell JE Jr: The roles of
cyclin A2, B1, and B2 in early and late mitotic events. Mol Biol
Cell. 21:3149–3161. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kino T, Gragerov A, Valentin A,
Tsopanomihalou M, Ilyina-Gragerova G, Erwin-Cohen R, Chrousos GP
and Pavlakis GN: Vpr protein of human immunodeficiency virus type 1
binds to 14-3-3 proteins and facilitates complex formation with
Cdc25C: Implications for cell cycle arrest. J Virol. 79:2780–2787.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Knight GL, Turnell AS and Roberts S: Role
for Wee1 in inhibition of G2-to-M transition through the
cooperation of distinct human papillomavirus type 1 E4 proteins. J
Virol. 80:7416–7426. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jackman M, Lindon C, Nigg EA and Pines J:
Active cyclin B1-Cdk1 first appears on centrosomes in prophase. Nat
Cell Biol. 5:143–148. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kramer A, Mailand N, Lukas C, Syljuasen
RG, Wilkinson CJ, Nigg EA, Bartek J and Lukas J:
Centrosome-associated Chk1 prevents premature activation of
cyclin-B-Cdk1 kinase. Nat Cell Biol. 6:884–891. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gavet O and Pines J: Progressive
activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev Cell.
18:533–543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Gavet O and Pines J: Activation of cyclin
B1-Cdk1 synchronizes events in the nucleus and the cytoplasm at
mitosis. J Cell Biol. 189:247–259. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Strasser A, Harris AW, Jacks T and Cory S:
DNA damage can induce apoptosis in proliferating lymphoid cells via
p53-independent mechanisms inhibitable by Bcl-2. Cell. 79:329–339.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Schmitt CA, Rosenthal CT and Lowe SW:
Genetic analysis of chemoresistance in primary murine lymphomas.
Nat Med. 6:1029–1035. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
67
|
Thornberry NA and Lazebnik Y: Caspases:
Enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wolf BB and Green DR: Suicidal tendencies:
Apoptotic cell death by caspase family proteinases. J Biol Chem.
274:20049–20052. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Pal MK, Jaiswar SP, Srivastav AK, Goyal S,
Dwivedi A, Verma A, Singh J, Pathak AK, Sankhwar PL and Ray RS:
Synergistic effect of piperine and paclitaxel on cell fate via
cyt-c, Bax/Bcl-2-caspase-3 pathway in ovarian adenocarcinomas
SKOV-3 cells. Eur J Pharmacol. 791:751–762. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Qu Z, Jiang C, Wu J and Ding Y:
Lenalidomide induces apoptosis and inhibits angiogenesis via
caspase3 and VEGF in hepatocellular carcinoma cells. Mol Med Rep.
14:4781–4786. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View Article : Google Scholar : PubMed/NCBI
|