Introduction
Due to its malignant growth and high invasive
ability, colorectal cancer is one of the leading cancers and
recurrence and metastasis are usually the main causes of mortality
in affected patients (1–3). Unfortunately, a number of patients are
diagnosed with advanced-stage colon cancer, leading to a high rate
of mortality (4). At present, the
standard treatment for colorectal cancer includes surgery and
chemotherapy. Despite significant progress in the treatment of
colon cancer patients, the overall prognosis and long-term survival
rate of patients have not been significantly improved in the past
decade (5,6). Therefore, elucidating its pathological
process is of great significance in finding new targets for the
treatment of colon cancer.
Chaperone-mediated autophagy (CMA) is a selective
form of autophagy in which the substrate protein contains a peptide
sequence motif (KFERQ) (7,8), which can be recognized by the
molecular chaperone heat shock cognate 70 (HSC70), targeted to the
lysosome surface (9) and then
combined with lysosome-associated membrane protein type 2A (LAMP2A)
to enter lysosomes for degradation (10). The activity of CMA is determined by
the level of LAMP2A on the lysosomal membrane (11,12)
and the efficiency of LAMP2A assembly and disassembly (13). Studies have shown that CMA leads to
significant changes in glucose and lipid metabolism, as well as
whole body energy metabolism (14,15),
which is closely related to the occurrence and development of
tumors. CMA has been demonstrated to be abnormally activated in a
variety of tumor tissues (16–21),
which can promote the survival, migration and drug resistance of
tumor cells. It has been confirmed that downregulation of CMA
reduces cell proliferation by regulating glucose metabolism in lung
cancer cells and melanoma (17).
Our research also found that CMA can promote glycolysis in breast
cancer (22). A recent study found
that LAMP2A expression levels are elevated in colon cancer tissues
and mouse models and LAMP2A knockdown can inhibit the proliferation
of colon cancer cells (16).
However, the role of CMA in colorectal cancer cells remains largely
unknown and exploring the undefined relationship between CMA and
LAMP2A may provide valuable insight into the pathogenesis of
colorectal cancer. The present study hypothesized that LAMP2A can
regulate colorectal cancer growth via CMA.
Materials and methods
Tissue array and
immunohistochemistry
A human colon cancer tissue array (CO808) was
purchased from Alenabio Biotechnology and written informed consent
was obtained from all subjects before collecting the samples. All
procedures were approved by the Ethics Committee of the General
Hospital of Tibet Area Military Command with the ethical approval
number 2023-KD004-01. All experiments were performed in accordance
with the relevant guidelines and regulations of the Declaration of
Helsinki. Immunohistochemistry was conducted as described
previously (20). Briefly, the
slide was deparaffinized in xylene, rehydrated in alcohol and
water, antigen retrieved with an improved citrate antigen retrieval
solution (Beyotime Institute of Biotechnology) at 100°C for 20 min,
blocked using an immunohistochemical blocking buffer (Beyotime
Institute of Biotechnology) at 37°C for 60 min, incubated with
LAMP2A antibody (Abcam, 1 mg/ml; cat. no. ab18528; 1:100) at 4°C
overnight, treated with a horseradish peroxidase-labeled polymer at
room temperature 25°C for 60 min, then incubated with DAB (Beyotime
Institute of Biotechnology) and counterstained with hematoxylin
(Beyotime Institute of Biotechnology) at room temperature 25°C for
5 min. Each sample was assigned an immunoreactivity score that
calculated the sum of the intensity of positive tumor cells
(0=none; 1=weak; 2=intermediate, 3=strong) and the estimated
fraction of positive staining tumor cells (0=none, 1 ≤10%, 2=10-50%
and 3 ≥50%).
Cell culture and cell infection
SW480 and HT29 human colorectal cancer cell lines
were purchased from Cobioer Biosciences and cultured in DMEM
supplemented with 10% fetal bovine serum (all from Gibco; Thermo
Fisher Scientific, Inc.), incubated in an atmosphere of 5%
CO2 at 37°C and STR profiling given by Cobioer
Biosciences was used for authentication of the HT29 cell line. The
short interfering (sh)RNA sequences targeting the coding region of
LAMP2A gene and negative control were 5′-GCAGTGCAGATGACGACAA-3′
(1283) and 5′-TTCTCCGAACGTGTCACGT-3′, respectively. The full shRNA
sequences used for inserting into the BamHI and EcoRI
sites of the pGLV-EGFP vector (GenePharma) were synthesized as
follows: LAMP2A sense:
5′-GATCCGCAGTGCAGATGACGACAATTCAAGAGATTGTCGTCATCTGCACTGCTTTTTTG-3′,
LAMP2A antisense:
5′-AATTCAAAAAAGCAGTGCAGATGACGACAATCTCTTGAATTGTCGTCATCTGCACTGCG-3′;
negative shRNA control sense:
5′-GATCCGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAACTTTTTTG-3′,
negative shRNA control antisense:
5′-AATTCAAAAAAGTTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAACG-3′.
The primers used for LAMP2A overexpression were as follows:
forward:5′-GATATGGCGGCCGCGCCACCATGGTGTGCTTCCGCCTCTTCC-3′, reverse:
5′-GTATGGGATCCCTAAAATTGCTCATATCCAGCATGATG-3′ and then the LAMP2A
sequences were cloned into the NotI and BamHI sites
of EF1α-LV5-EGFP vector (GenePharma). The LAMP2A shRNA and
overexpressing vector were used for lentivirus packing and then the
relative lentivirus suspension was obtained (all performed by
GenePharma). SW480 and HT29 cells (3×105 cells/well)
were infected with lentivirus-mediated LAMP2A shRNA and
overexpressing vector at a multiplicity of infection of 30 for 12 h
at 37°C, and then cells were stably selected with 1 µg/ml puromycin
(Invitrogen; Thermo Fisher Scientific, Inc.) after 3 days.
Western blotting
Western blotting analysis was conducted as described
previously (20). Anti-LAMP2A
antibody was from Abcam (1 mg/ml; cat. no. ab18528, 1:1,000), and
anti-β-actin antibody (500 µg/ml; cat. no. BM3873, 1:10,000) and
anti-GAPDH antibody (500 µg/ml; cat. no. BA2913, 1:10,000) were
from Wuhan Boster Biological Technology, Ltd. Briefly, the cells
were lysed in RIPA buffer (Beyotime Institute of Biotechnology) and
protein levels were tested using a BCA kit (Beyotime Institute of
Biotechnology). A total of 20 µg protein was loaded per lane, then
10% SDS-PAGE was performed and the proteins were transferred to
polyvinylidene difluoride (PVDF) membranes, blocked with blocking
buffer (Beyotime Institute of Biotechnology) at room temperature
25°C for 60 min and incubated with the corresponding primary
antibodies in primary antibody dilution buffer (Beyotime Institute
of Biotechnology) at 4°C overnight, washed and incubated with
secondary antibody (Wuhan Boster Biological Technology, Ltd.; 1
mg/ml; cat. no. BA1082, 1:5,000) in secondary antibody dilution
buffer at room temperature 25°C for 60 min and then detected by an
ECL kit (MilliporeSigma). Densitometry was analyzed using ImageJ
v1.53e software (National Institutes of Health).
Degradation of human recombinant
his-GAPDH by lysosomal association
Degradation of human recombinant his-GAPDH by
lysosomal association was conducted as described previously
(20). Briefly, 250 mg pellets of
colorectal cancer cells with different LAMP2A expression were
harvested. Intact lysosomes (100 µg protein) were isolated using a
Lysosome Enrichment Kit (Merck KGaA), incubated with protease
inhibitor on ice for 10 min, mixed with 6X energy regenerating
system (60 mM ATP, 12 mM phosphocreatine, 0.3 mg/ml creatine
phosphokinase, 60 mM MgCl2; pH 7.3), purified
recombinant protein of 50 µg his-GAPDH and 2 µg his-HSC70 (all
obtained from Clinical Biochemistry Laboratory, Army Medical
University) and then collected by centrifugation at 18,000 × g for
30 min at 4°C, washed and immunoblotted with GAPDH antibody
(incubated with primary antibody at 4°C overnight, and incubated
with secondary antibody at 25°C for 60 min).
Colony formation assay
Colony formation assays were performed as described
previously (20). Cells were seeded
(300 cells/well) onto 6-well plates and cultured for 14 days. Then,
4% paraformaldehyde was used to fix the colonies for 15 min and 1%
crystal violet was used to stain the cells for 30 min at room
temperature, 25°C. The colonies were counted under a light
microscope at room temperature, 25°C. A clone was defined as
containing at ≥50 cells.
MTT assay
For the cell growth curve, SW480 and HT29 cells
(1×103/well) were seeded onto a 96-well plate. The
number of viable cells was determined using the MTT assay on days
1, 2, 3, 4, 5, 6 and 7. Briefly, 20 µl of 5 mg/ml MTT
(MilliporeSigma) was added to each well and the cells were
incubated at 37°C for 5 h. DMSO (200 µl per well) was added to the
plates and the optical density (OD) at 490 nm was detected. For the
oxaliplatin, H2O2 or 2-deoxy-D-glucose (2-DG)
treatment assay, SW480 and HT29 cells (1×104/well) were
seeded onto a 96-well plate and treated with oxaliplatin (Selleck
Chemicals; 0, 1, 2 and 3 µmol/l), H2O2 (0,
0.2, 0.4 and 0.8 µM) or 5 mmol/l 2-DG for 36 h.
Wound healing assay
SW480 and HT29 cells (2×104/well) were
inoculated onto a 24-well plate and wounds were made using a 200 µl
pipette tip upon reaching 90-100% confluence. After washing with
PBS three times, the cells were cultured in serum-free medium and
the wound widths were recorded under a light microscope at 0 and 48
h, at a magnification of ×40. The wound healing rate (%) was equal
to (width at 48 h/width at 0 h) ×100%.
Transwell invasion assay
The extracellular matrix gel (Matrigel;
MilliporeSigma) was diluted with cold DMEM at a ratio of 1:2,
incubated at 37°C for 30 min to form a gel and inoculated into the
upper chamber of a Transwell chamber (MilliporeSigma) at 40 µl.
SW480 and HT29 cells (1×104/well) were resuspended in
serum-free medium and inoculated into the upper chamber for 48 h to
allow the cells to invade into the lower chamber. The cells in the
upper chamber were wiped off with a cotton swab, fixed with 4%
paraformaldehyde for 15 min, stained with 1% crystal violet for 30
min and then counted in five randomly selected areas under a light
microscope at room temperature 25°C, and magnification, ×400.
Extracellular acidification rate
(ECAR) assay
The ECAR assay was performed as described previously
(22). Cells
(0.8×104/well) were seeded onto XF 96-well plates,
incubated for 16 h, incubated with 200 µl of Seahorse basic culture
medium (Agilent Technologies, Inc.) and then placed in a
CO2-free incubator at 37°C for 60 min. After calibration
with the hydration plate, the XF 96-well plate with cells was
supplemented with 25 mmol/l glucose, 1 µmol/l oligomycin and 50
mmol/l 2-DG, all from Seahorse and then detected by a Seahorse
energy metabolism analyzer (Agilent Technologies, Inc.).
Lactate determination assay
The level of lactic acid in the culture medium of
colorectal cancer cells was detected by the L-Lactate Assay Kit
(Merck KGaA). The supernatant from cells (1×106/well)
was seeded onto a 6-well plate for 36 h and collected and
centrifuged at a speed of 3,350 × g for 10 min at room temperature,
25°C. A standard curve was prepared and the level of lactic acid
was detected according to the manufacturer's instructions.
Statistical analysis
Statistical analysis was conducted using the
statistical software SPSS 21.0 (IBM Corp.). The results of the
experiments were analyzed by unpaired Student's t-test and
P<0.05 was considered to indicate a statistically significant
difference.
Results
LAMP2A expression is elevated in colon
cancer
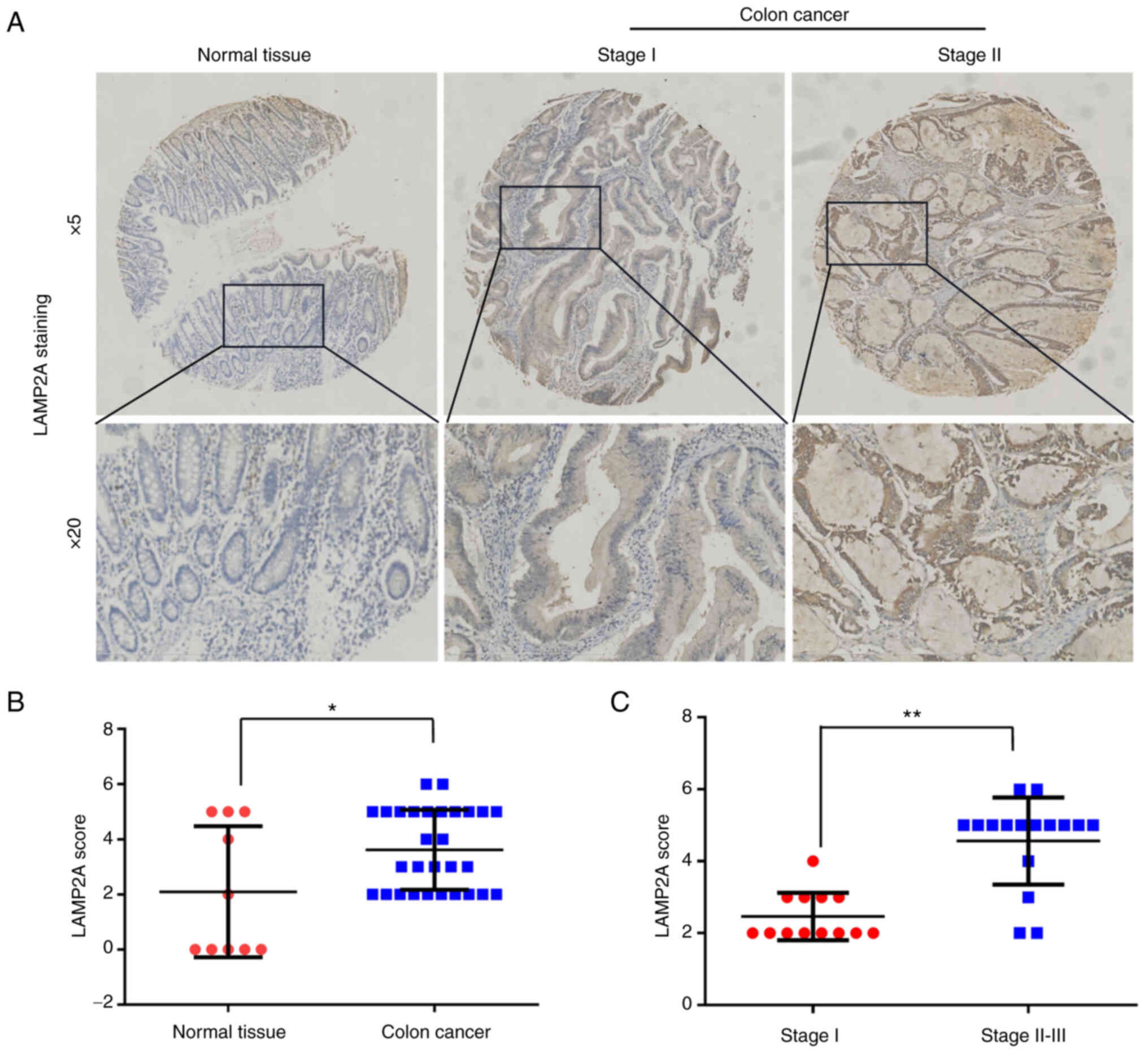
To understand the physiological significance of
LAMP2A in colon cancer, LAMP2A expression was assessed in 29 colon
cancer tissues and 10 normal tissues by tissue array-based
immunohistochemistry. The LAMP2A expression level and
immunoreactivity score (calculated as the sum of the LAMP2A
intensity and percentage scores) were higher in colon cancer
tissues than in normal tissues (Fig. 1A
and B), indirectly suggesting higher CMA activity in colon
cancer tissues. In addition, the LAMP2A expression level was
significantly higher in stage II–III colon cancer than in stage I
colon cancer, indicating that LAMP2A expression is positively
associated with the stage of colon cancer (Fig. 1C).
Elevated LAMP2A expression in
colorectal cancer cells is related to elevated CMA activity
The most efficient way to regulate CMA activity is
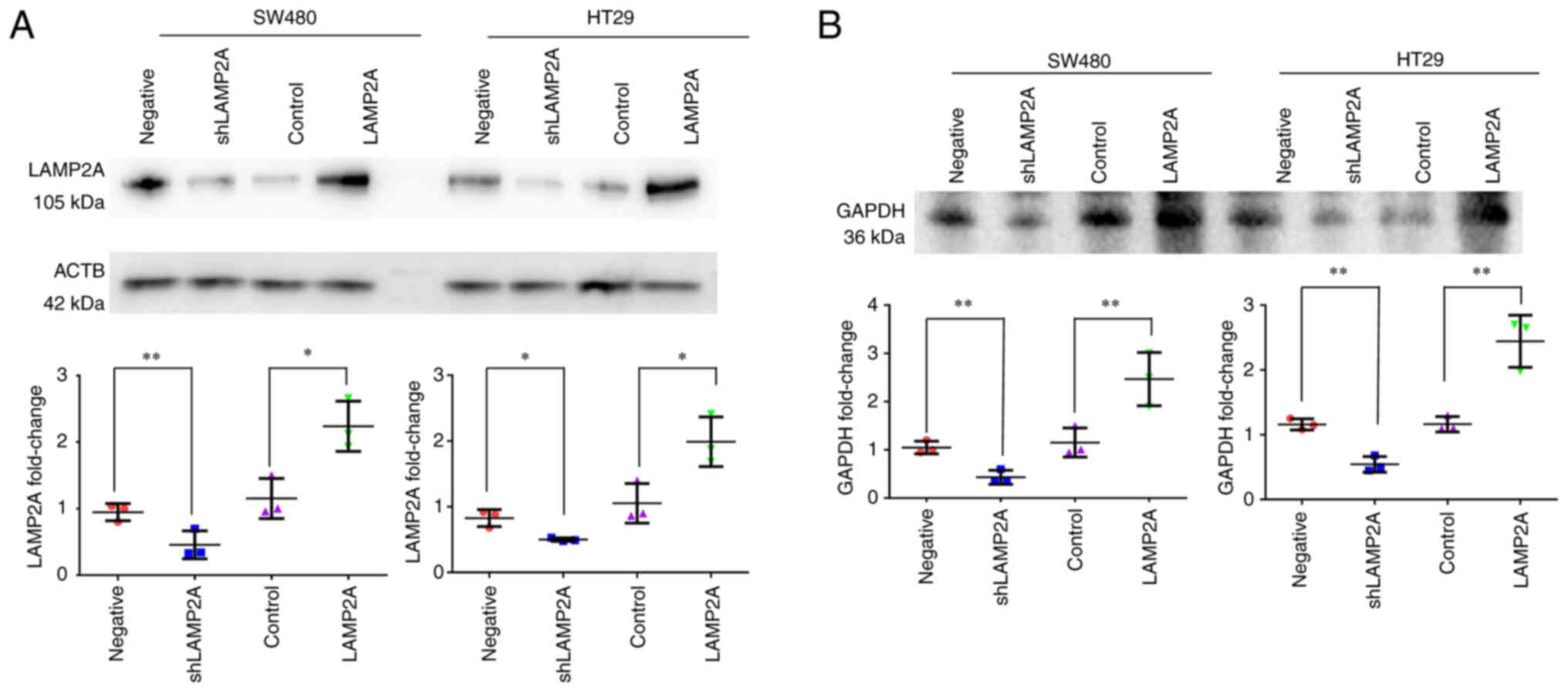
by modifying the expression level of LAMP2A (23). The present study manipulated CMA
activity in SW480 and HT29 cells by cell infection with LAMP2A
shRNA and overexpression vector to illustrate the role of CMA in
colorectal cancer more fully. The results showed that SW480 and
HT29 colorectal cancer cells with different levels of LAMP2A
expression were successfully constructed (Fig. 2A). A direct method was performed to
measure CMA activity by lysosome association of purified GAPDH with
active intact lysosomes under the condition of an energy
regenerating system, his-HSC70 and protease inhibitor (21). Lysosomes isolated from SW480 and
HT29 cells with LAMP2A knockdown showed decreased lysosomal
association of GAPDH, suggesting lower CMA activity, in these cells
compared with the control cells, while lysosomes from SW480 and
HT29 cells with LAMP2A overexpression demonstrated increased
lysosomal association of GAPDH, indicating higher CMA activity
(Fig. 2B).
LAMP2A promotes colorectal cancer cell
proliferation
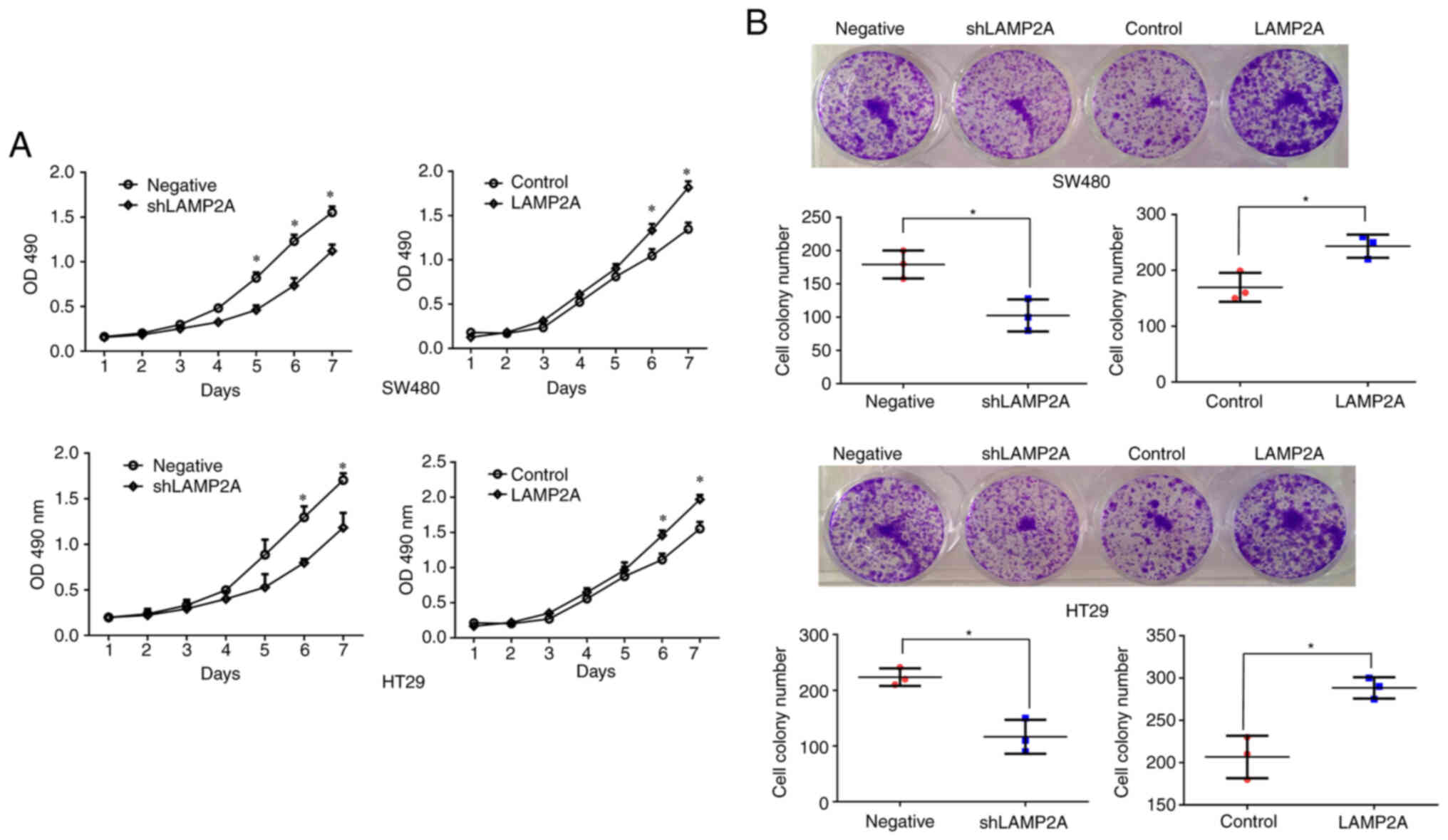
CMA has been demonstrated to be involved in tumor
proliferation and is required for tumor growth (17). The 7-day MTT growth curve was used
to observe the effect of LAMP2A on cell proliferation and cell
proliferation was inhibited after LAMP2A knockdown and promoted
after LAMP2A overexpression in SW480 and HT29 cells (Fig. 3A). Colony formation assays also
demonstrated that LAMP2A knockdown significantly suppressed the
proliferation of SW480 and HT29 cells and that LAMP2A
overexpression significantly promoted the proliferation of SW480
and HT29 cells (Fig. 3B).
LAMP2A protects colorectal cancer
cells from oxidative damage and promotes colorectal cancer cell
drug resistance
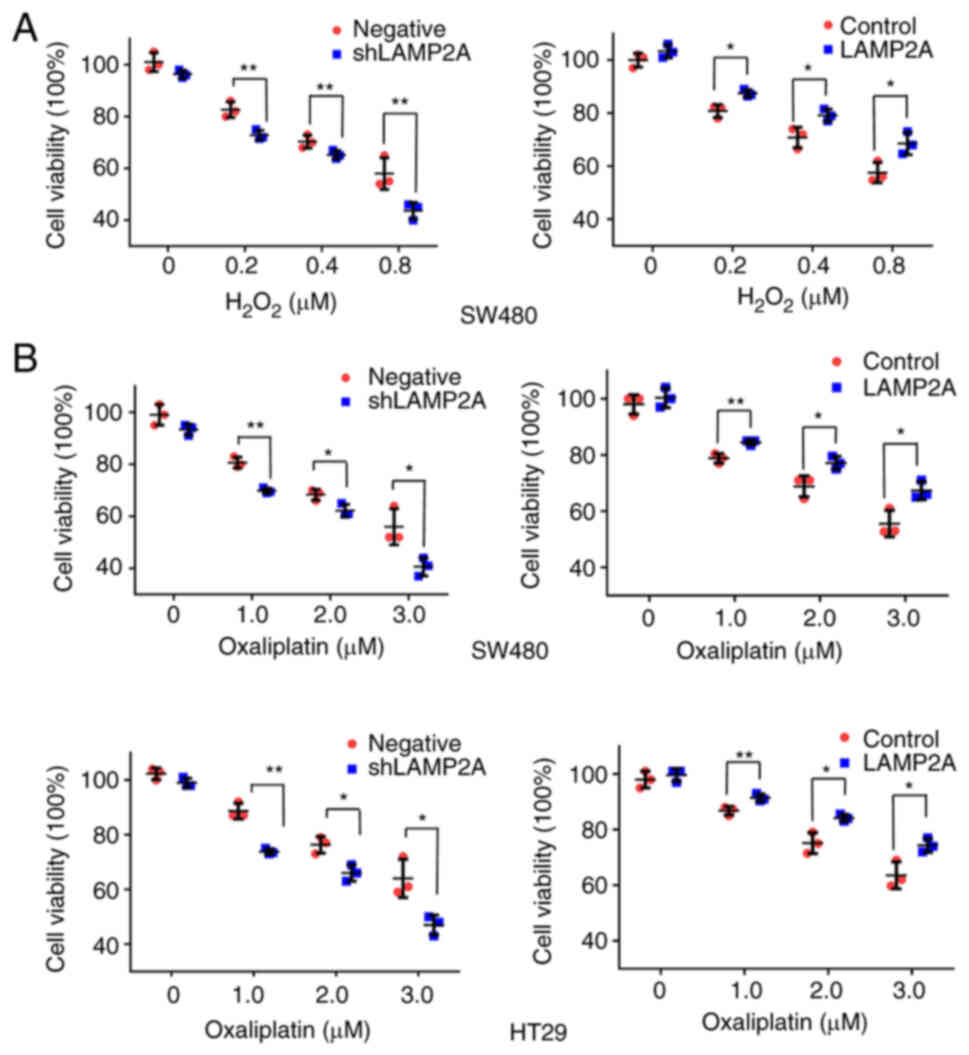
Tumors are always under the influence of oxidative
stress mediated by reactive oxygen species (ROS), which might be
one of the causes of cancer progression (21). To induce oxidative stress,
colorectal cancer cells were treated with different concentrations
of H2O2. The cell viability rate was
decreased after LAMP2A knockdown and increased after LAMP2A
overexpression (Fig. 4A).
Oxaliplatin is a chemotherapeutic drug that is widely used to treat
colon cancer (24). Colorectal
cancer cells were treated with different concentrations of
oxaliplatin and it was found that oxaliplatin resistance was
attenuated after LAMP2A knockdown and enhanced after LAMP2A
overexpression (Fig. 4B).
LAMP2A promotes colorectal cancer cell
migration and invasion
Metastasis is the most complex and deadly process
related to cancer and metastasis relies on tumor cell migration and
invasion (25). The effect of
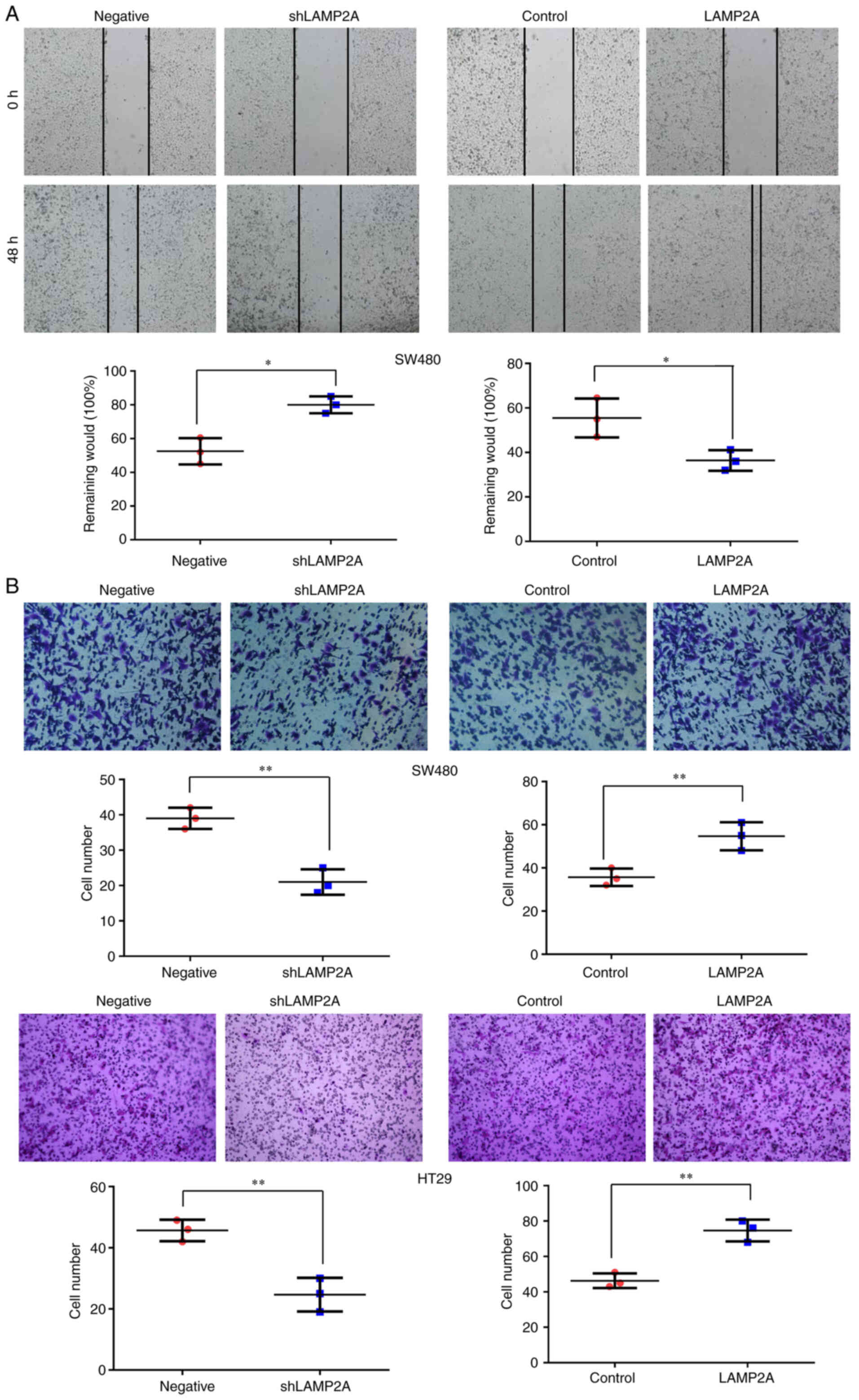
LAMP2A on colorectal cell migration was measured by wound healing
assay. LAMP2A knockdown significantly inhibited the migration of
SW480 cells, while LAMP2A overexpression significantly promoted the
migration of SW480 cells (Fig. 5A).
To further ascertain the role of CMA in colorectal cancer cell
invasion, a Transwell invasion assay was conducted. The number of
invasive cells was significantly reduced after LAMP2A knockdown and
increased after LAMP2A overexpression in SW480 and HT29 cells
(Fig. 5B).
LAMP2A promotes glycolysis in
colorectal cancer cells
Energy metabolism is required to maintain tumor
growth (26). The ECAR assay was
used to detect the effect of LAMP2A on the extracellular
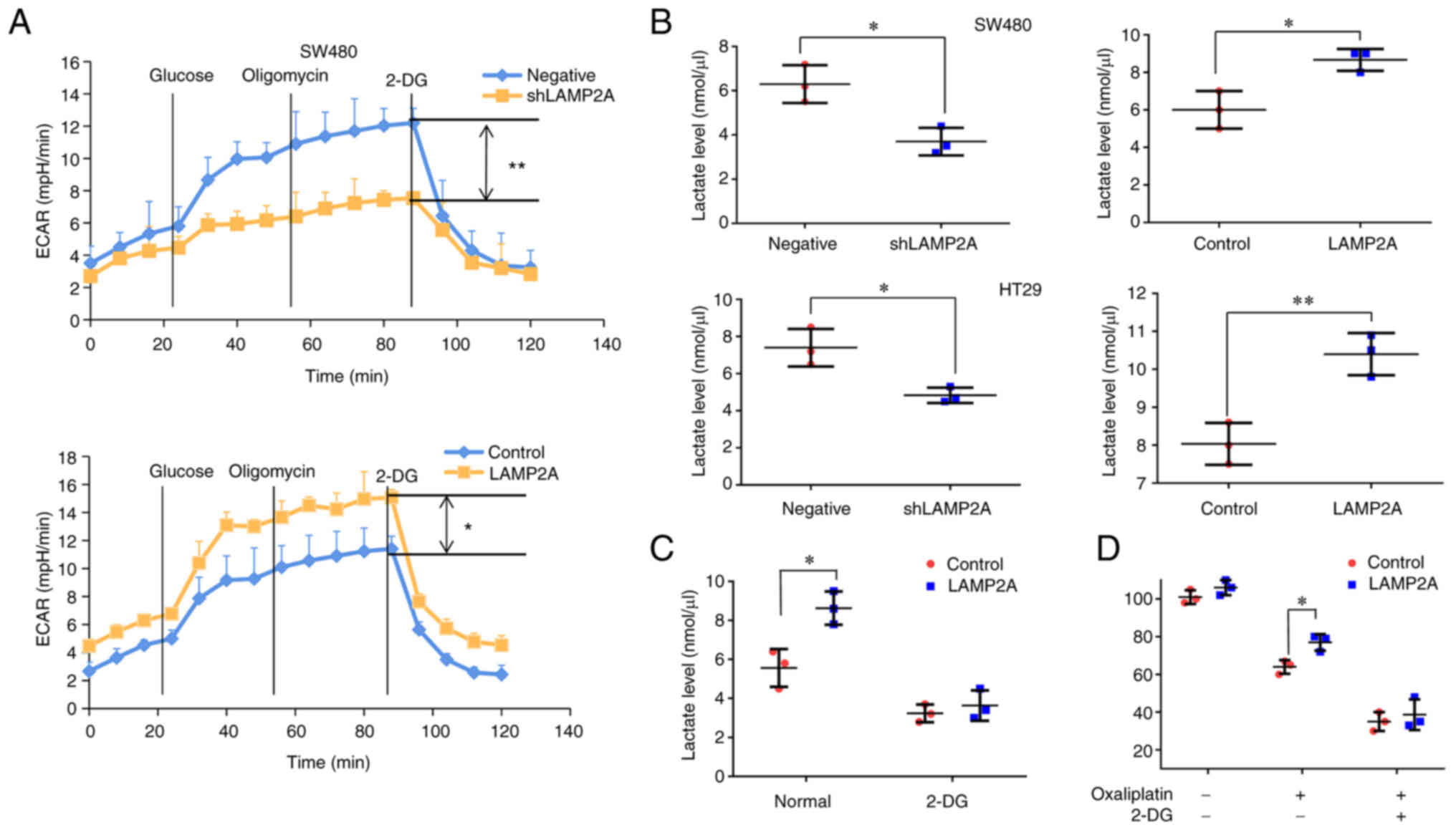
acidification rate of colorectal cancer cells. The results showed
that the maximum glycolytic capacity (MPH/min) was significantly
reduced after LAMP2A knockdown and the maximum glycolytic capacity
(MPH/min) was significantly increased after LAMP2A overexpression
(Fig. 6A). The extracellular
lactate level was decreased after LAMP2A knockdown and increased
after LAMP2A overexpression (Fig.
6B). To further explore the effect of glycolysis on
LAMP2A-induced oxaliplatin resistance in colorectal cancer cells,
SW480 cells were treated with 2-DG (a glycolysis inhibitor).
Lactate production was significantly suppressed in SW480 cells
overexpressing LAMP2A (Fig. 6C)
after 2-DG treatment. In addition, treatment with 2-DG also
markedly attenuated the effect of LAMP2A overexpression on the
viability of SW480 cells treated with oxaliplatin (Fig. 6D), which suggests that inhibition of
glycolysis could abate LAMP2A-induced chemoresistance in SW480
cells.
Discussion
Colon cancer is characterized by a high degree of
malignancy and rapid progression; 20-30% of colon cancer patients
present with metastasis at initial diagnosis (27) and ~50% of colon cancer patients will
experience recurrence within 3 years of initial surgery (28,29).
Combination chemotherapy is usually used in patients with colon
cancer, especially in advanced stages. The main conventional
chemotherapy drugs are 5-fluorouracil, oxaliplatin, irinotecan and
capecitabine (30) and combination
chemotherapy is initially effective for most patients. However,
with the emergence of drug resistance, ~50% of colon cancer
patients experience recurrence, leading to a decrease in the 5-year
survival rate of patients with distant metastasis to ~14% (31).
The latest cancer research indicates that CMA is
involved in the growth and proliferation of tumors and is expected
to become a new target for tumor treatment (32). LAMP2A, as the key protein of the CMA
pathway, has been confirmed to exhibit abnormal expression in
various tumor tissues and is generally considered to promote tumor
proliferation, metastasis and drug resistance (33). Previous studies have shown that CMA
can reduce apoptosis and promote the proliferation of colon
carcinoma cells and they monitored CMA activity by measurement of
changes in LAMP2A expression levels (16,34).
However, accurate measurement of CMA is only achieved by directly
measuring the turnover of CMA substrate proteins into lysosomes in
the presence of protease inhibitor. The present study performed a
direct method of measuring CMA activity by GAPDH degradation in
colon cancer cells to explore the effect of CMA on colorectal
cancer more fully. Lysosomal association of GAPDH was reduced in
LAMP2A knockdown cells and upregulated in cells with LAMP2A
overexpression, indicating the corresponding CMA activity had been
directly observed. Pathologic importance of CMA in cancer cell
proliferation has been extensively studied, whereas a link between
CMA and cell viability under the treatment of
H2O2 or oxaliplatin has not been thoroughly
explored, especially in colorectal cancer. ROS have been regarded
as a series of destructive molecules that have a detrimental effect
on cell homoeostasis and H2O2 appears to be
the main ROS (35). The cell
viability rate was decreased after LAMP2A knockdown and increased
after LAMP2A overexpression, indicating that LAMP2A can protect
colorectal cancer cells from oxidative damage. Oxaliplatin
resistance is a challenge in the treatment of colorectal cancer
patients (36). The present study
found that oxaliplatin resistance was significantly inhibited
following LAMP2A knockdown and enhanced following LAMP2A
overexpression. The data obtained from the previous studies
(16,34) demonstrated the significance of CMA
in colorectal cancer, which prompted the present study to further
explore the other effects of CMA on different colorectal cancer
cells due to the cell heterogeneity. Metastasis causes most
cancer-related deaths and is a multistep biological process that
involves the dissemination of cancer cells to anatomically distant
organs/sites (37). In present
study, the migration and invasion of colorectal cancer cells was
significantly inhibited after silencing LAMP2A and significantly
increased after upregulation of LAMP2A.
Abnormal glucose metabolism is considered an
important marker of cancer cells and this characteristic of tumor
cells has attracted widespread interest (38). Mitochondrial oxidative
phosphorylation is the main energy source for normal cells.
Conversely, aerobic glycolysis is usually the main energy source of
cancer cells. This phenomenon is called the ‘Warburg effect’
(39). Metabolic reprogramming has
become a new center of activity in cancer research and is closely
related to tumor proliferation, metastasis, drug resistance and a
poor prognosis (40). CMA has been
shown to regulate glycolysis in lung cancer, melanoma and breast
cancer (17,22). However, the modulation between CMA
and glycolysis in colorectal cancer cells remains to be elucidated.
The present study demonstrated that LAMP2A could promote the
production of lactic acid, increase the extracellular acidification
rate and increase glycolysis levels in colorectal cancer cells. In
addition, inhibition of glycolysis by 2-DG markedly attenuated
LAMP2A-induced chemoresistance in colorectal cancer cells, which
might provide a new strategy to treat colorectal cancer by
combination of downregulation of CMA activity and glycolysis
inhibitor. However, the exact mechanism by which LAMP2A regulates
chemoresistance and glycolysis in colorectal cancer requires
further study.
In summary, the present study demonstrates that CMA
could promote cell proliferation, cell metastasis, cell survival
during oxidative stress, oxaliplatin resistance and glycolysis in
colorectal cancer and thus could have remarkable diagnostic and
therapeutic implications for colorectal cancer patients.
Acknowledgements
The authors would like to thank Mr. Siyuan Liu for
language editing.
Funding
The present study was supported by National Natural Science
Foundation of China (grant nos. 82001997 and 82072946) and Doctoral
Program of University of Tibetan Medicine (grant nos. BSDJS-20-08
and BSDJS-23-06). The funders had no role in study design, data
collection and analysis, decision to publish, or preparation of the
manuscript.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
RC, SL and QH conceived and designed the
experiments, JW, CH, JX, XZ, MT, ZW, LP, YZ and MH performed the
experiments, ZW, JY and YG analyzed the data, SL and QH supervised
the study and wrote the manuscript. SL and QH confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from all
patients before collecting the samples. All the procedures were
approved by the Ethics Committee of General Hospital of Tibet Area
Military Command and the ethical approval number was 2023-KD004-01.
All experiments were performed in accordance with relevant
guidelines and regulations of Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lotfollahzadeh S, Recio-Boiles A and Cagir
B: Colon cancer. StatPearls [Internet] Treasure Island (FL):
StatPearls Publishing; 2023
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu IS and Cheung WY: Metastatic colorectal
cancer in the era of personalized medicine: A more tailored
approach to systemic therapy. Can J Gastroenterol Hepatol.
2018:94507542018.PubMed/NCBI
|
|
4
|
Das V, Kalita J and Pal M: Predictive and
prognostic biomarkers in colorectal cancer: A systematic review of
recent advances and challenges. Biomed Pharmacother. 87:8–19. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xie YH, Chen YX and Fang JY: Comprehensive
review of targeted therapy for colorectal cancer. Signal Transduct
Target Ther. 5:222020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moghimi-Dehkordi B and Safaee A: An
overview of colorectal cancer survival rates and prognosis in Asia.
World J Gastrointest Oncol. 4:71–75. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dice JF: Peptide sequences that target
cytosolic proteins for lysosomal proteolysis. Trends Biochem Sci.
15:305–309. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Olson TS, Terlecky SR and Dice JF:
Targeting specific proteins for lysosomal proteolysis. Biomed
Biochim Acta. 50:393–397. 1991.PubMed/NCBI
|
|
9
|
Chiang HL, Terlecky SR, Plant CP and Dice
JF: A role for a 70-kilodalton heat shock protein in lysosomal
degradation of intracellular proteins. Science. 246:382–385. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cuervo AM and Dice JF: A receptor for the
selective uptake and degradation of proteins by lysosomes. Science.
273:501–503. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cuervo AM and Dice JF: Unique properties
of lamp2a compared to other lamp2 isoforms. J Cell Sci.
113:4441–4450. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cuervo AM and Dice JF: Regulation of
lamp2a levels in the lysosomal membrane. Traffic. 1:570–583. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bandyopadhyay U, Sridhar S, Kaushik S,
Kiffin R and Cuervo AM: Identification of regulators of
chaperone-mediated autophagy. Mol Cell. 39:535–547. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kaushik S and Cuervo AM: The coming of age
of chaperone-mediated autophagy. Nat Rev Mol Cell Biol. 19:365–381.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang M, Luo S, Chen W, Zhao L and Wang X:
Chaperone-mediated autophagy: A potential target for metabolic
diseases. Curr Med Chem. 30:1887–1899. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Peng JQ, Han SM, Chen ZH, Yang J, Pei YQ,
Bao C, Qiao L, Chen WQ and Liu B: Chaperone-mediated autophagy
regulates apoptosis and the proliferation of colon carcinoma cells.
Biochem Biophys Res Commun. 522:348–354. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kon M, Kiffin R, Koga H, Chapochnick J,
Macian F, Varticovski L and Cuervo AM: Chaperone-mediated autophagy
is required for tumor growth. Sci Transl Med. 3:109ra1172011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding ZB, Fu XT, Shi YH, Zhou J, Peng YF,
Liu WR, Shi GM, Gao Q, Wang XY, Song K, et al: Lamp2a is required
for tumor growth and promotes tumor recurrence of hepatocellular
carcinoma. Int J Oncol. 49:2367–2376. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang T, Ren C, Qiao P, Han X, Wang L, Lv
S, Sun Y, Liu Z, Du Y and Yu Z: PIM2-mediated phosphorylation of
hexokinase 2 Is critical for tumor growth and paclitaxel resistance
in breast cancer. Oncogene. 37:5997–6009. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Han Q, Deng Y, Chen S, Chen R, Yang M,
Zhang Z, Sun X, Wang W, He Y, Wang F, et al: Downregulation of
ATG5-dependent macroautophagy by chaperone-mediated autophagy
promotes breast cancer cell metastasis. Sci Rep. 7:47592017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Saha T: LAMP2A overexpression in breast
tumors promotes cancer cell survival via chaperone-mediated
autophagy. Autophagy. 8:1643–1656. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen R, Li P, Fu Y, Wu Z, Xu L, Wang J,
Chen S, Yang M, Peng B, Yang Y, et al: Chaperone-mediated autophagy
promotes breast cancer angiogenesis via regulation of aerobic
glycolysis. PLoS One. 18:e2815772023.
|
|
23
|
Massey AC, Kaushik S, Sovak G, Kiffin R
and Cuervo AM: Consequences of the selective blockage of
chaperone-mediated autophagy. Proc Natl Acad Sci USA.
103:5805–5810. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
André T, Boni C, Mounedji-Boudiaf L,
Navarro M, Tabernero J, Hickish T, Topham C, Zaninelli M, Clingan
P, Bridgewater J, et al: Oxaliplatin, fluorouracil, and leucovorin
as adjuvant treatment for colon cancer. N Engl J Med.
350:2343–2351. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Majidpoor J and Mortezaee K: Steps in
metastasis: An updated review. Med Oncol. 38:32021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pavlova NN, Zhu J and Thompson CB: The
hallmarks of cancer metabolism: Still emerging. Cell Metab.
34:355–377. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van der Geest LG, Lam-Boer J, Koopman M,
Verhoef C, Elferink MA and de Wilt JH: Nationwide trends in
incidence, treatment and survival of colorectal cancer patients
with synchronous metastases. Clin Exp Metastasis. 32:457–465. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Desantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aghili M, Izadi S, Madani H and Mortazavi
H: Clinical and pathological evaluation of patients with early and
late recurrence of colorectal cancer. Asia Pac J Clin Oncol.
6:35–41. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alfaro Alfaro ÁE, Murillo Castillo B,
Cordero García E, Tascón J and Morales AI: Colon cancer
pharmacogenetics: A narrative review. Pharmacy (Basel). 10:952022.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Siegel RL, Miller KD, Goding Sauer A,
Fedewa SA, Butterly LF, Anderson JC, Cercek A, Smith RA and Jemal
A: Colorectal cancer statistics, 2020. CA Cancer J Clin.
70:145–164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hubert V, Weiss S, Rees AJ and Kain R:
Modulating chaperone-mediated autophagy and its clinical
applications in cancer. Cells. 11:25622022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Arias E and Cuervo AM: Pros and cons of
chaperone-mediated autophagy in cancer biology. Trends Endocrinol
Metab. 31:53–66. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xuan Y, Zhao S, Xiao X, Xiang L and Zheng
HC: Inhibition of chaperone-mediated autophagy reduces tumor growth
and metastasis and promotes drug sensitivity in colorectal cancer.
Mol Med Rep. 23:3602021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Landry WD and Cotter TG: ROS signalling,
NADPH oxidases and cancer. Biochem Soc Trans. 42:934–938. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ning T, Li J, He Y, Zhang H, Wang X, Deng
T, Liu R, Li H, Bai M, Fan Q, et al: Exosomal miR-208b related with
oxaliplatin resistance promotes Treg expansion in colorectal
cancer. Mol Ther. 29:2723–2736. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Farhadi P, Yarani R, Dokaneheifard S and
Mansouri K: The emerging role of targeting cancer metabolism for
cancer therapy. Tumour Biol. 42:10104283209652842020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Weng ML, Chen WK, Chen XY, Lu H, Sun ZR,
Yu Q, Sun PF, Xu YJ, Zhu MM, Jiang N, et al: Fasting inhibits
aerobic glycolysis and proliferation in colorectal cancer via the
Fdft1-mediated AKT/mTOR/HIF1α pathway suppression. Nat Commun.
11:18692020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y, Zhao T, Li Z, Wang L, Yuan S and
Sun L: The role of ASCT2 in cancer: A review. Eur J Pharmacol.
837:81–87. 2018. View Article : Google Scholar : PubMed/NCBI
|