Introduction
Chronic myeloid leukemia (CML) is a
myeloproliferative disorder driven by a translocation of
chromosomes 9 and 22, which encodes the oncogenic breakpoint
cluster region-Abelson (Bcr-Abl) tyrosine kinase (1). Approximately 5,000 new patients are
affected by CML each year in the US (2). Although considerable advance has been
made in the treatment of CML by the development of imatinib
mesylate (STI-571), a tyrosine kinase inhibitor which targets the
Bcr-Abl kinase (3), evidence
suggests that up to a third of CML patients require alternate
therapy due to intolerance or resistance to STI-571 and/or
progression of the advanced disease (4). Thus, further research is needed to
better understand and overcome drug intolerance and resistance
mechanisms as well as to identify new drugs or substances with
better therapeutic efficacy against CML.
Mistletoe extract has been used as complementary
therapy for several types of human solid tumors (5). All industrially produced mistletoe
extracts are made from the semi-parasitic plant mistletoe Viscum
album Loranthaceae, also called European mistletoe. Mistletoe
extract contains several biologically active components, such as
lectins, viscotoxins, flavonoides, and phytosterols (6,7).
Accumulating evidence indicates that mistletoe extract or its
components has antitumor effects by stimulating the immune system
(8), reducing tumor mass (9), and inhibiting growth and/or inducing
apoptosis of cancer cells (9–11).
Results from numerous in vitro and in vivo studies
suggest possible mechanisms underlying the antitumor effect of
mistletoe extract or its components, including inhibition of cell
cycle (11), reactive oxygen
species (ROS) and loss of the mitochondrial membrane potential
(12,13), activation of caspases (14–16),
degradation of cytoskeletal proteins (16), inhibition of protein synthesis
(17), and altered expression
and/or activity of the family of B-cell lymphoma-2 (Bcl-2),
telomerase, protein kinase B (PKB), and the family of
mitogen-activated protein kinase (MAPK) (18–20).
Abnobaviscum F® is a standardized
preparation of aqueous European mistletoe extract from the host
tree Fraxinus. It has been shown that Abnobaviscum F inhibits
growth of various established cancer cell lines and primary mammary
carcinoma cells isolated from surgical resections (21). The actions of Abnobaviscum F on
hematological malignancies, however, remain unknown. In this study,
we investigated the effect of Abnobaviscum F on the growth and
survival of K562 (human CML), RPMI-8226 (human plasmacytoma), and
L1210 (murine lymphocytic leukemia) cells.
Materials and methods
Materials
RPMI, FBS, and penicillin-streptomycin were
purchased from WelGene (Daegu, Korea). ECL western detection
reagents were obtained from Thermo Scientific (Waltham, MA, USA).
Bradford reagent was purchased from Bio-Rad (Hercules, CA, USA).
N-benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone (z-VAD-fmk),
SP600125, and protease inhibitor cocktail (100X) were bought from
Calbiochem (Madison, WI, USA). SB203580 was purchased from Biomol
(Plymouth Meeting, PA, USA). An antibody of caspase-9 was purchased
from Stressgen (Ann Arbor, MI, USA). Myeloid cell leukemia-1
(Mcl-1), protein kinase C-δ (PKC-δ), glucose-regulated protein 78
(GRP78), and activating transcription factor 6 (ATF6) was purchased
from Santa Cruz Biotechnology (Delaware, CA, USA). An antibody of
poly (ADP-ribose) polymerase (PARP) was purchased from Roche
(Basel, Switzerland). Antibodies of phospho-extracellular signal
regulated kinase-1/2 (p-ERK-1/2), total-ERK (T-ERK-1/2),
phospho-c-Jun N-terminal kinase-1/2 (p-JNK-1/2), T-JNK-1/2, p-p38
MAPK, T-p38 MAPK, p-PKB, T-PKB, phospho-eukaryotic translation
initiation factor 2α (p-eIF-2α) were purchased from Epitomics
(Burlingame, CA, USA). An antibody of T-eIF-2α was purchased from
Cell Signaling Tech (Beverly, MA, USA). Other reagents, including
anti-actin mouse monoclonal antibody and glutathione (GSH), were
purchased from Sigma (St. Louis, MO, USA).
Cell culture
K562, RPMI-8226 (ATCC; Manassas, VA, USA), and L1210
(KCLB; Seoul, Korea) cells were grown at 37°C in a humidified
condition of 95% air and 5% CO2 in RPMI-1640
supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin,
and 100 μg/ml streptomycin.
Cell count assay
K562, RPMI-8226, or L1210 cells were seeded in
24-well plates and treated without or with different concentrations
of Abnobaviscum F for 24 h. The number of surviving cells, which
cannot be stained with trypan blue dye, were counted using standard
light microscopy.
Measurement of DNA fragmentation
K562, RPMI-8226, or L1210 cells were seeded in a
60-mm dish and treated with Abnobaviscum F at the indicated
concentrations or times. The cells were harvested, washed, and
lysed in a buffer [50 mM Tris (pH 8.0), 0.5% sarkosyl, 0.5 mg/ml
proteinase K, and 1 mM EDTA] at 55°C for 3 h, followed by the
addition of RNase A (0.5 μg/ml) and incubation at 55°C for
18 h. The lysates were centrifuged at 10,000 × g for 20 min.
Genomic DNA was extracted with equal volume of neutral
phenol-chloroform-isoamyl alcohol mixture (25:24:1), and analyzed
by electrophoresis on a 1.8% agarose gel. The DNA was visualized
and photographed under UV illumination after staining with ethidium
bromide.
Preparation of whole cell lysates
Treated cells were washed with PBS and exposed to
cell lysis buffer [50 mM Tris-Cl (pH 7.4), 150 mM NaCl, 0.1% sodium
dodecyl sulfate, 0.25% sodium deoxycholate, 1% Triton X-100, 1%
Nonidet P-40, 1 mM EDTA, 1 mM EGTA, proteinase inhibitor cocktail
(1X)]. The cell lysates were collected in a 1.5 ml tube and
centrifuged for 20 min at 4°C at 12,000 rpm. The supernatant was
saved and protein concentrations were determined with Bradford
reagent.
Western blotting
Proteins (50 μg) were separated by SDS-PAGE
(10%) and transferred onto nitrocellulose membranes (Millipore;
Billerica, MA, USA). The membranes were washed with TBS (10 mM
Tris, 150 mM NaCl) supplemented with 0.05% (vol/vol) Tween-20
(TBST) followed by blocking with TBST containing 5% (wt/vol)
non-fat dried milk. The membranes were incubated overnight with
antibodies specific for the protein of interest at 4°C. The
membranes were exposed to secondary antibodies coupled to
horseradish peroxidase at room temperature for 2 h. The membranes
were washed, and immunoreactivities were detected by ECL
reagents.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total-RNA was isolated using the RNAzol-B
(Tel-Test). Total-RNA (3 μg) was reverse transcribed using a
random hexadeoxynucleotide primer and reverse transcriptase. Single
stranded cDNA was amplified by PCR with the following primers:
Mcl-1 sense, 5′-ATCTCTCGGTACCT TCGGGAG-3′ and anti-sense,
5′-ACCAGCTCCTACT CCAGCAAC-3′; death receptor-4 (DR4) sense,
5′-CTGAGCAACGCAGACTCGC TGTCCAC-3′ and anti-sense,
5′-AAGGACACGGCAGAGC CTGTGCCAT-3′; DR5 sense, 5′-AGCCGCTCATGAGGA
AGTTGG-3′ and anti-sense, 5′-GGCAAGTCTCTCTCCCAG CGTCTC-3′; C/EBP
homologous protein (CHOP) sense, 5′-GTCCCTAGCTTGGCTGACAGA′ and
anti-sense, 5′-TGG AGAGCGAGGGCTTTG-3′; actin sense, 5′-GGTGAAGGT
CGGTGTGAACG-3′ and anti-sense, 5′-GGTAGGAACAC GGAAGGCCA-3′. The PCR
conditions applied were: Mcl-1, 25 cycles of denaturation at 95°C
for 45 sec, annealing at 56°C for 45 sec, and extension at 72°C for
45 sec; DR4 and DR5, 35 cycles of denaturation at 95°C for 30 sec,
annealing at 63°C for 30 sec, and extension at 72°C for 30 sec;
CHOP, 35 cycles of denaturation at 94°C for 30 sec, annealing at
55°C for 1 min, and extension at 72°C for 30 sec; actin, 25 cycles
of denaturation at 95°C for 30 sec, annealing at 56°C for 30 sec,
and extension at 72°C for 30 sec.
Measurement of GSH content
Cellular GSH content was measured using a GSH Assay
kit (Calbiochem). Briefly, K562 cells were grown in a 60-mm dish
and treated with Abnobaviscum F for 0.5 or 2 h. At each time point,
the culture medium was removed and cells were sonicated in 5%
metaphosphoric acid. The homogenate was centrifuged at 3,000 × g
for 10 min, and its supernatant was used for GSH measurement
according to the manufacturer’s instructions.
Statistical analysis
Cell count analysis was performed in triplicates and
repeated three times. Data are expressed as mean ± standard error.
Significance (p<0.05) was determined by one-way ANOVA.
Results
Abnobaviscum F reduces survival and
induces apoptosis of leukemic cells
Initially, we investigated whether Abnobaviscum F
affects the survival of K562, RPMI-8226, or L1210 cells by cell
count analysis. Treatment with Abnobaviscum F for 24 h reduced
survival of K562 (Fig. 1A),
RPMI-8226 (Fig. 1C), or L1210
(Fig. 1E) cells in a
concentration-dependent manner. We next analyzed whether
Abnobaviscum F induces apoptosis of K562, RPMI-8226, or L1210 cells
by measuring nuclear DNA fragmentation, a strong apoptotic index.
Treatment with Abnobaviscum F for 24 h similarly induced apoptosis
of K562 (Fig. 1B), RPMI-8226
(Fig. 1D), or L1210 (Fig. 1F) cells in a concentration-dependent
manner. The 20 μg/ml dose of Abnobaviscum F presented the
strongest anti-survival and pro-apoptotic effects and was therefore
used in further studies.
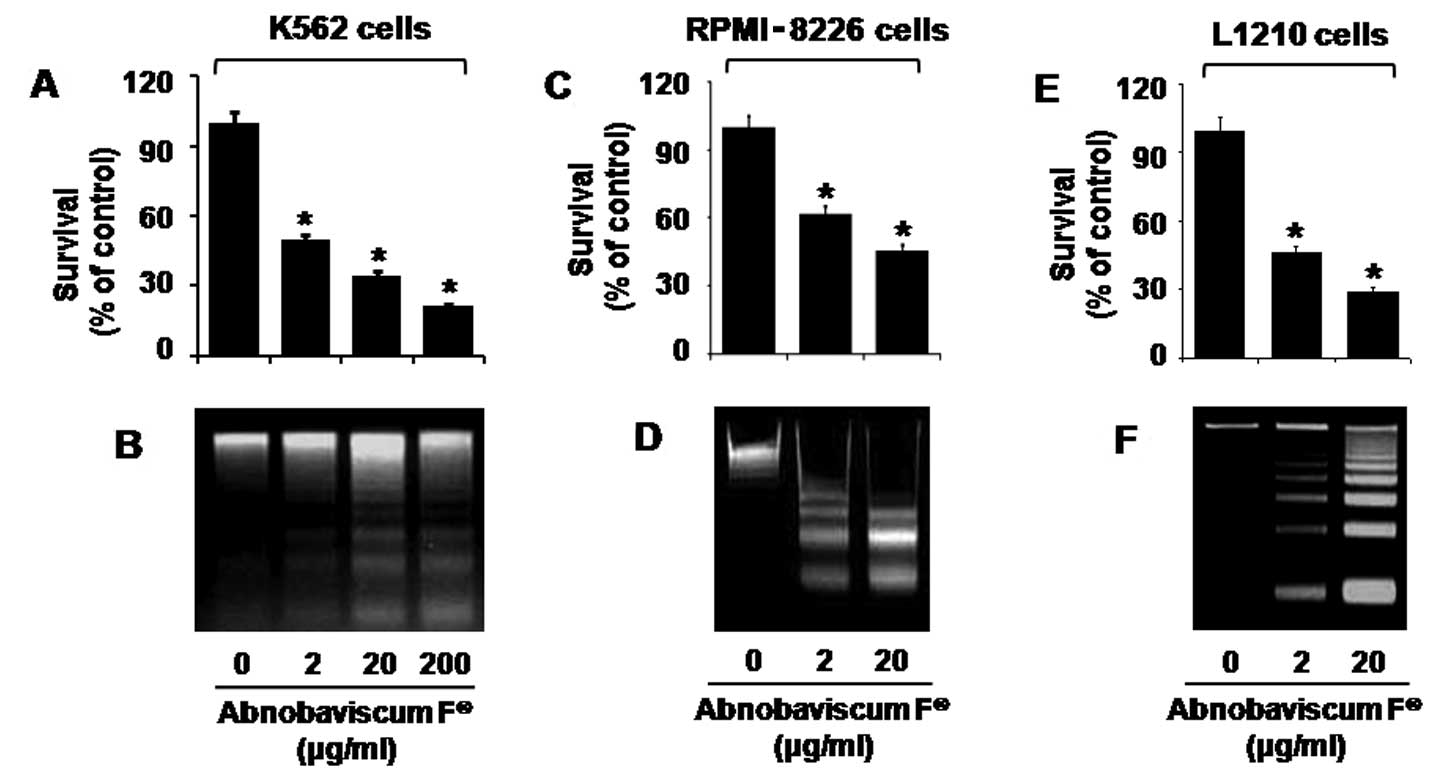 | Figure 1Effect of Abnobaviscum F®
on the growth of various leukemic cells. (A, C and E) K562,
RPMI-8226, or L1210 cells, respectively, were treated with the
indicated concentrations of Abnobaviscum F for 24 h. The number of
surviving cells was counted under microscope and normalized as
percentage of drug-free control. Data are mean ± SE of three
independent experiments. *p<0.05 compared to the
value in the absence of Abnobaviscum F. (B, D and F) K562,
RPMI-8226, or L1210 cells, respectively, were treated with
Abnobaviscum F at the indicated doses for 24 h. Extra-nuclear
fragmented DNA from the conditioned cells was extracted and
analyzed on a 1.7% agarose gel. The image is representative of
three independent experiments. |
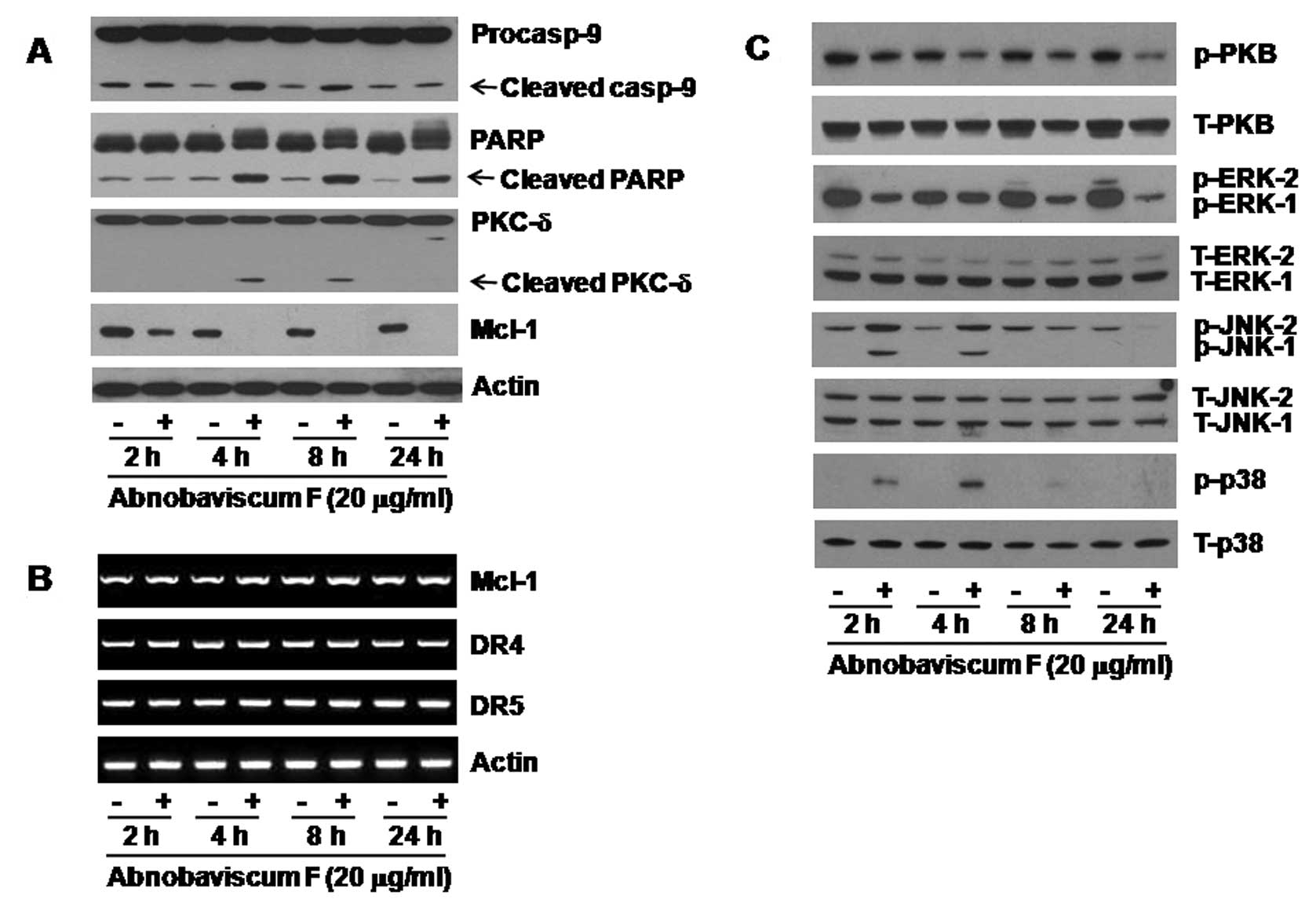
Abnobaviscum F induces activation of the
intrinsic caspase pathway and the downregulation of Mcl-1 in K562
cells
To understand the mechanisms underlying Abnobaviscum
F-induced apoptosis, we next treated K562 cells with Abnobaviscum F
for the indicated times and measured expression of growth-related
proteins. In Fig. 2A, Abnobaviscum
F treatment, particularly at 4 h, decreased the expression of
procaspase-9 but increased that of cleaved (active) caspase-9. High
levels of cleaved PARP and PKC-δ were also observed at 4 h.
Moreover, there was a substantial reduction of Mcl-1 at 2 h,
followed by a complete loss at 4 h. However, mRNA levels of Mcl-1
and DR4 and DR5 were not changed in K562 cells treated with
Abnobaviscum F at any time point measured (Fig. 2B).
Abnobaviscum F modulates expression
levels and/or activities of signaling proteins in K562 cells
Treatment with Abnobaviscum F time-dependently
reduced the levels of both phosphorylated PKB and total PKB
(Fig. 2C). Abnobaviscum F treatment
also reduced the levels of phosphorylated ERK-1/2 but did not
affect expression of total ERK-1/2. Of note, Abnobaviscum F
treatment at 2 or 4 h stimulated phosphorylation of JNK-1/2 and p38
MAPK. Expression levels of total JNK-1/2 and p38 MAPK remained
unchanged by Abnobaviscum F treatment for all the times tested.
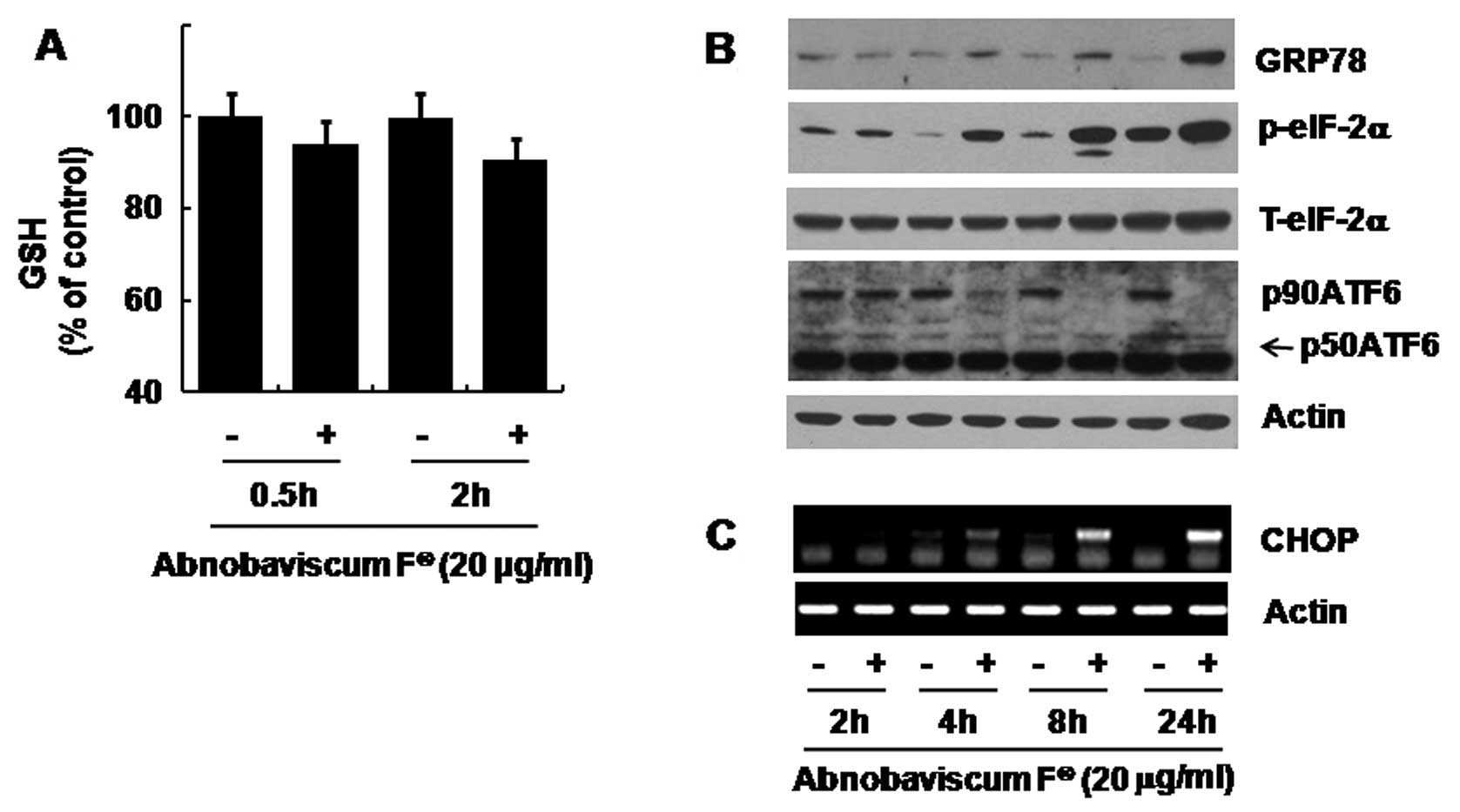
Abnobaviscum F decreases cellular GSH
content but induces ER stress in K562 cells
We next determined whether Abnobaviscum F induces
oxidative stress (by measuring cellular GSH levels) in K562 cells.
In Fig. 3A, early treatments of
Abnobaviscum F decreased cellular GSH levels. We also analyzed
whether Abnobaviscum F triggers ER stress (by measuring expressions
of ER stress-related proteins) in K562 cells. In Fig. 3B, treatment with Abnobaviscum F
time-dependently increased the expression of GRP78 and
phosphorylation of eIF-2α. However, levels of p90ATF6 were largely
decreased at 4 h, followed by complete loss at 8 h. Although only
mildly increased, p50ATF6 was also detected at 24 h. As shown in
Fig. 3C, there was also a
time-dependent induction of CHOP mRNA following treatment with
Abnobaviscum F.
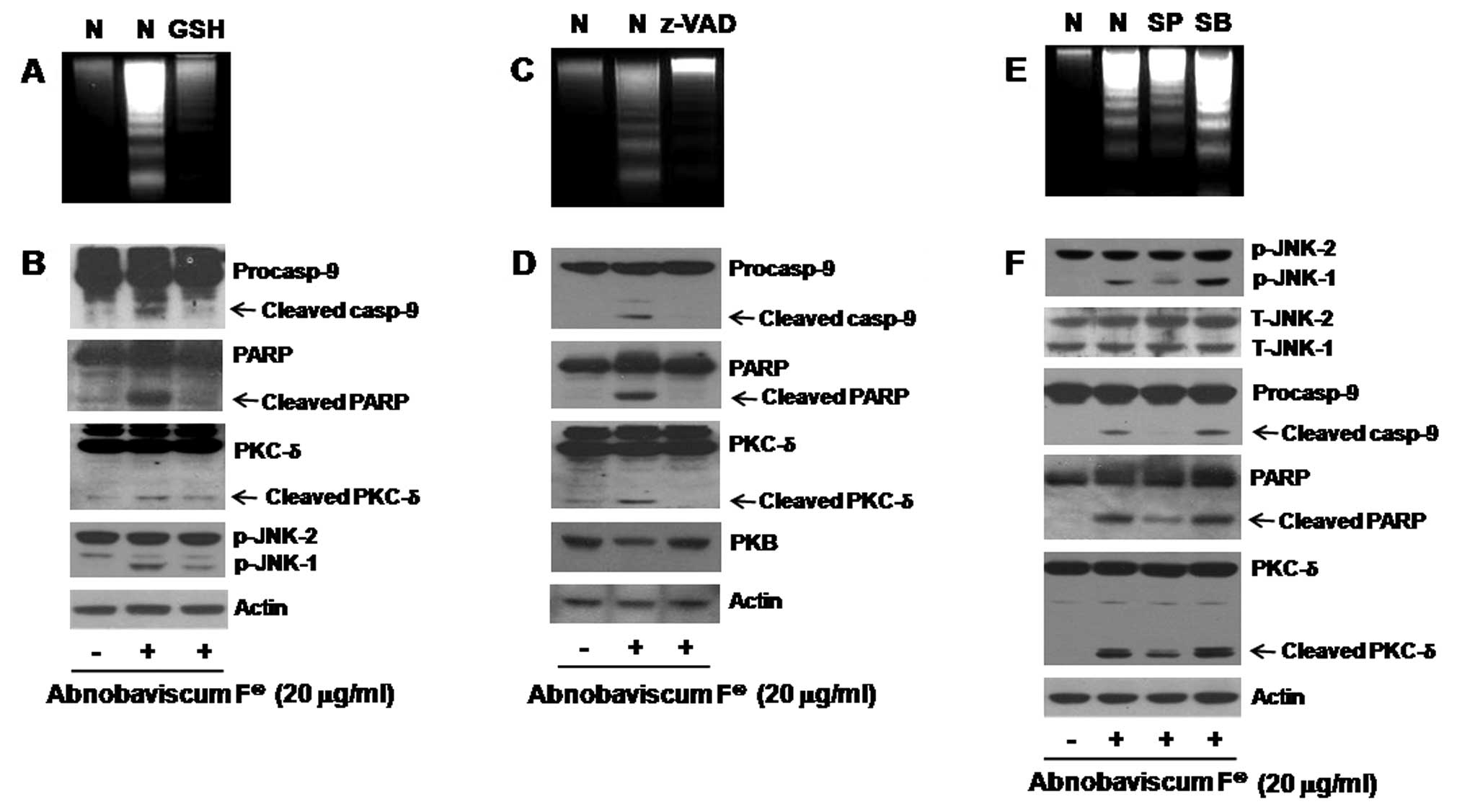
Role of GSH reduction, activation of
caspases, and JNK-1 phosphorylation in Abnobaviscum F-induced
apoptosis in K562 cells
We next evaluated the role of GSH depletion,
activation of caspases, and phosphorylation of JNK-1/2 or p38 MAPK
in the Abnobaviscum F-induced apoptosis of K562 cells. Abnobaviscum
F-induced apoptosis of K562 cells was blocked by pretreatment with
either GSH (a reducing agent; Fig.
4A), z-VAD-fmk (a pan-caspase inhibitor; Fig. 4C), or SP600125 (an inhibitor of
JNK-1/2) but not SB203580 (a p38 MAPK inhibitor; Fig. 4E). Pretreatment with GSH also
blocked the activation of caspase-9, cleavage of PARP and PKC-δ,
and phosphorylation of JNK-1 by Abnobaviscum F (Fig. 4B). The caspase inhibitor was also
able to suppress the ability of Abnobaviscum F to cleave PARP and
PKC-δ and to downregulate total PKB (Fig. 4D). SP600125, which only repressed
phosphorylation of JNK-1, effectively inhibited the Abnobaviscum
F-induced activation of caspase-9 and cleavage of PARP and
PKC-δ.
Discussion
Herein we demonstrated for the first time that
Abnobaviscum F, an aqueous European mistletoe extract from
Fraxinus, induces apoptosis of K562, RPMI-8226 and L1210 leukemic
cells. Moreover, our data suggest that at least in K562 cells
Abnobaviscum F induces apoptosis through the modulation of cellular
GSH levels, the regulation of expression levels and activities of a
number of intracellular cell survival signaling proteins, and via
the induction of ER stress.
Previous studies have suggested a similar
antileukemic effect of mistletoe extract or its components. For
instance, Iscador®, a fermented European mistletoe
extract from oak, inhibits growth of Molt 4 human T-lymphoblastoid
leukemic cells (22), and mistletoe
lectins or agglutinins suppress growth and induce apoptosis of U937
human myeloleukemic cells (12,15,20)
and HL-60 human acute lymphoblastic leukemia cells (14). In the present study, we demonstrated
that Abnobaviscum F reduces survival and induces apoptosis of K562
cells. Apoptosis is closely related to two pathways; the intrinsic
(mitochondrial) and the extrinsic (DR) pathways and several
proteins are involved in both these pathways to mediate apoptosis.
Central to both apoptosis pathways are the caspases, a group of the
essential proteases required for the execution of cell death by
apoptotic stimuli (23). In resting
cells, caspases are synthesized as zymogens (inactive precursors),
but upon exposure to apoptotic stimuli, they become processed via
partial proteolytic cleavage and activated in cells. Active
caspases then cleave many target proteins, including PARP, PKC-δ,
and other vital proteins (23,24).
It has previously been demonstrated that
Iscador-induced apoptosis of human lung and breast cancer cells is
mediated via the mitochondrial intrinsic caspase pathway (11). The present findings that
Abnobaviscum F induces activation of caspase-9 (Fig. 2A) without affecting the expression
of DRs (Fig. 2B) and z-VAD-fmk, a
pan-caspase inhibitor, blocks Abnobaviscum F-induced apoptosis
(Fig. 4C) in K562 cells also
suggest that activation of caspases through the intrinsic pathway
is important for Abnobaviscum F-induced apoptosis of K562 cells.
Mcl-1 is an anti-apoptotic protein that is involved in apoptosis
initiation and caspase activation by regulating the mitochondrial
membrane integrity (25,26). There is further evidence suggesting
Mcl-1 as a critical survival factor for multiple myeloma (27). Considering that Abnobaviscum F
rapidly reduces expression of Mcl-1 (2 h) at the protein but not
mRNA levels, which precedes processing and activation of caspase-9
(4 h) in K562 cells (Fig. 2A and
B), the present study suggests that Abnobaviscum F targets
Mcl-1 at post-transcriptional stage and loss of Mcl-1 may
contribute to activation of the intrinsic caspase pathway and/or
apoptosis of K562 cells exposed to Abnobaviscum F.
PKB is a survival protein kinase (28), and a high expression and/or activity
of PKB often correlates strongly with the growth of cancer cells
(29,30). Moreover, PKB dephosphorylation has
been shown to play a role in the apoptosis of human head and neck
cancer cells induced by mistletoe lectin (18). Herein Abnobaviscum F treatment led
to PKB dephosphorylation in K562 cells (Fig. 2C), and to the caspase-dependent
reduction of total PKB protein (Fig. 5D). ERK-1/2 are frequently
activated and have a pro-survival function in AML and their
pharmacological inhibitors are promising agents in the treatment of
AML (31,32). On the contrary, JNK-1/2 and p38 MAPK
are often activated in cells under stressful conditions (33), and their activities are linked to
apoptotic death of K562 or B lymphoma cells by anticancer drugs
and/or agents (34,35). Previous reports showed that
mistletoe lectin-II-induced apoptosis of U937 cells is mediated by
activating JNK-1/2 (19) or p38
MAPK (20). However, our results
show that Abnobaviscum F-induced apoptosis of K562 cells is
associated with the ability of Abnobaviscum F to inhibit PKB and
ERK-1/2 but activate JNK-1 (Figs.
2C and 4E).
Oxidative stress is also involved in apoptosis
induction (36). Both ROS-dependent
mechanisms of mistletoe lectin-induced apoptosis (12,13)
and ROS-independent mechanisms of Viscum album
agglutinin-I-induced apoptosis (16) have been proposed. In our
experimental conditions, we showed that oxidative stress,
associated with a reduction of cellular GSH, is critical for the
killing effect of Abnobaviscum F on K562 cells (Figs. 3A and 4A). Our data also suggest a crosstalk
between reduction of cellular GSH and several other cellular events
triggered by Abnobaviscum F, in which Abnobaviscum F may primarily
lower cellular GSH that leads to activation of JNK-1, JNK-1
mediates the activation of caspases, and caspases induce the
proteolysis of PARP, PKC-δ, and PKB (Fig. 4B, D, and F).
ER stress appears to be a common process in cancer
cell death by anticancer drugs and/or agents (37,38).
Cells undergoing ER stress are characterized by upregulation of
molecular chaperones (e.g., GRP78) (39) and transcription factors (e.g., CHOP)
(40), phosphorylation of eIF-2α
and inhibition of global translation (41), and reduction of p90ATF6 (42). In line with these studies, our
present findings of Abnobaviscum F-mediated upregulation of GRP78
and CHOP, enhancement of eIF-2α phosphorylation, and reduction of
p90ATF6 in K562 cells (Fig. 3B and
C) indicate that Abnobaviscum F induces ER stress, which may
contribute to apoptosis.
Cancer cells differ in their cell of origin and the
molecular alterations causing them to transform. Thus, drugs or
agents that inhibit growth and induce apoptosis in different cancer
cells may represent better anticancer strategies. In this context,
the present study demonstrates the ability of Abnobaviscum F to
inhibit growth and induce apoptosis in not only K562 (Fig. 1B) but also RPMI-8226 (Fig. 1D) and L1210 (Fig. 1F) leukemic cells. Collectively, the
findings presented herein may shed light on the possibility of
applying Abnobaviscum F to the treatment of human leukemia, as a
single and/or combinatorial regimen with other antileukemic
therapies.
Acknowledgements
The authors thank Dr David Bishop-Bailey (William
Harvey Research Institute, Queen Mary University of London, UK) for
carefully reading and correcting the manuscript, and Abnoba KOREA
(Seoul, Korea) for kindly providing Abnobaviscum F® for
this study.
References
|
1
|
Borgaonkar DS: Philadelphia-chromosome
translocation and chronic myeloid leukaemia. Lancet. 1:12501973.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics. CA Cancer J Clin. 59:225–249. 2009.
|
|
3
|
Druker BJ, Talpaz M, Resta DJ, et al:
Efficacy and safety of a specific inhibitor of the BCR-ABL tyrosine
kinase in chronic myeloid leukemia. N Engl J Med. 344:1031–1037.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Druker BJ, Guilhot F, O’Brien SG, et al:
Five-year follow-up of patients receiving imatinib for chronic
myeloid leukemia. N Engl J Med. 355:2408–2417. 2006.PubMed/NCBI
|
|
5
|
Ernst E, Schmidt K and Steuer-Vogt MK:
Mistletoe for cancer? A systematic review of randomised clinical
trials. Int J Cancer. 107:262–267. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khwaja TA, Dias CB and Pentecost S: Recent
studies on the anticancer activities of mistletoe Viscum
album and its alkaloids. Oncology. 1:42–50. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Urech K, Schaller G and Jaggy C:
Viscotoxins, mistletoe lectins and their isoforms in mistletoe
Viscum album L. extracts Iscador. Arzneimittelforschung.
56:428–434. 2006.PubMed/NCBI
|
|
8
|
Heinzerling L, von Baehr V, Liebenthal C,
et al: Immunologic effector mechanisms of a standardized mistletoe
extract on the function of human monocytes and lymphocytes in
vitro, ex vivo, and in vivo. J Clin Immunol. 26:347–359. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Burger AM, Mengs U, Schüler JB, et al:
Antiproliferative activity of an aqueous mistletoe extract in human
tumor cell lines and xenografts in vitro. Arzneimittelforschung.
51:748–757. 2001.PubMed/NCBI
|
|
10
|
Büssing A, Suzart K and Schweizer K:
Differences in the apoptosis-inducing properties of Viscum
album L. extracts. Anticancer Drugs. 8:S9–S14. 1997.
|
|
11
|
Harmsma M, Ummelen M, Dignef W, et al:
Effects of mistletoe Viscum album L. extracts Iscador on
cell cycle and survival of tumor cells. Arzneimittelforschung.
56:474–482. 2006.
|
|
12
|
Kim MS, Lee J, Lee KM, et al: Involvement
of hydrogen peroxide in mistletoe lectin-II-induced apoptosis of
myeloleukemic U937 cells. Life Sci. 73:1231–1243. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim WH, Park WB, Gao B, et al: Critical
role of reactive oxygen species and mitochondrial membrane
potential in Korean mistletoe lectin-induced apoptosis in human
hepatocarcinoma cells. Mol Pharmacol. 66:1383–1396. 2004.
View Article : Google Scholar
|
|
14
|
Lyu SY, Park WB, Choi KH, et al:
Involvement of caspase-3 in apoptosis induced by Viscum
album varcoloratum agglutinin in HL-60 cells. Biosci
Biotechnol Biochem. 65:534–541. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim MS, So HS, Lee KM, et al: Activation
of caspase cascades in Korean mistletoe Viscum album
var.coloratum lectin-II-induced apoptosis of human
myeloleukemic U937 cells. Gen Pharmacol. 34:349–355.
2000.PubMed/NCBI
|
|
16
|
Lavastre V, Chiasson S, Cavalli H and
Girard D: Viscum album agglutinin-I (VAA-I) induces
apoptosis and degradation of cytoskeletal proteins in human
leukemia PLB-985 and X-CGD cells via caspases: lamin B1 is a novel
target of VAA-I. Leuk Res. 29:1443–1453. 2005. View Article : Google Scholar
|
|
17
|
Endo Y, Tsurugi K and Franz H: The site of
action of the A-chain of mistletoe lectin I on eukaryotic
ribosomes. The RNA N-glycosidase activity of the protein. FEBS
Lett. 231:378–380. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi SH, Lyu SY and Park WB: Mistletoe
lectin induces apoptosis and telomerase inhibition in human A253
cancer cells through dephosphorylation of Akt. Arch Pharm Res.
27:68–76. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park R, Kim MS, So HS, et al: Activation
of c-Jun N-terminal kinase 1 (JNK1) in mistletoe lectin II-induced
apoptosis of human myeloleukemic U937 cells. Biochem Pharmacol.
60:1685–1691. 2000. View Article : Google Scholar
|
|
20
|
Pae HO, Oh GS, Kim NY, et al: Roles of
extracellular signal-regulated kinase and p38 mitogen-activated
protein kinase in apoptosis of human monoblastic leukemia U937
cells by lectin-II isolated from Korean mistletoe. In Vitr Mol
Toxicol. 14:99–106. 2001. View Article : Google Scholar
|
|
21
|
Knöpfl-Sidler F, Viviani A, Rist L and
Hensel A: Human cancer cells exhibit in vitro individual
receptiveness towards different mistletoe extracts. Pharmazie.
60:448–454. 2005.PubMed/NCBI
|
|
22
|
Ribéreau-Gayon G, Jung ML, Di Scala D and
Beck JP: Comparison of the effects of fermented and unfermented
mistletoe preparations on cultured tumor cells. Oncology. 43:35–41.
1986.PubMed/NCBI
|
|
23
|
Cohen GM: Caspase: the executioners of
apoptosis. Biochem J. 326:1–16. 1997.
|
|
24
|
Lazebnik YA, Kaufmann SH, Desnoyers S, et
al: Cleavage of poly(ADP-ribose) polymerase by a proteinase with
properties like ICE. Nature. 371:346–347. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Adams JM and Cory S: The Bcl-2 protein
family: arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Festjens N, Van Gurp M, Van Loo G, et al:
Bcl-2 family members as sentinels of cellular intergrity and role
of mitochondrial intermembrane space proteins in apoptotic cell
death. Acta Haematol. 111:7–27. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang B, Gojo I and Fenton RG: Myeloid
cell factor-1 is a critical survival factor for multiple myeloma.
Blood. 99:1885–1893. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yoeli-Lerner M and Toker A: Akt/PKB
signaling in cancer: a function in cell motility and invasion. Cell
Cycle. 5:603–605. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakayama H, Ikebe T, Beppu M, et al: High
expression levels of nuclear factor kappaB, IkappaB kinase alpha
and Akt kinase in squamous cell carcinoma of the oral cavity.
Cancer. 92:3037–3044. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kallergi G, Agelaki S, Kalykaki A, et al:
Phosphorylated EGFR and PI3K/Akt signaling kinases are expressed in
circulating tumor cells of breast cancer patients. Breast Cancer
Res. 10:R802008. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Milella M, Kornblau SM, Estrov Z, et al:
Therapeutic targeting of the MEK/MAPK signal transduction module in
acute myeloid leukemia. J Clin Invest. 108:851–859. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lunghi P, Tabilio A, Dall’Aglio PP, et al:
Downmodulation of ERK activity inhibits the proliferation and
induces the apoptosis of primary acute myelogenous leukemia blasts.
Leukemia. 17:1783–1793. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tibbles LA and Woodgett JR: The
stress-activated protein kinase pathways. Cell Mol Life Sci.
55:1230–1254. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shim MJ, Kim HJ, Yang SJ, et al: Arsenic
trioxide induces apoptosis in chronic myelogenous leukemia K562
cells: possible involvement of p38 MAP kinase. J Biochem Mol Biol.
35:377–383. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Takada E, Hata K and Mizuguchi J:
c-Jun-NH2-terminal kinase potentiates apoptotic cell death in
response to carboplatin in B lymphoma cells. Cancer Chemother
Pharmacol. 62:569–576. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Simon HU, Haj-Yehia A and Levi-Schaffer F:
Role of reactive oxygen species (ROS) in apoptosis induction.
Apoptosis. 5:415–418. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nieto-Miguel T, Fonteriz RI, Vay L, et al:
Endoplasmic reticulum stress in the proapoptotic action of
edelfosine in solid tumor cells. Cancer Res. 67:10368–10378. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang X, Zhang Z, Jia L, et al:
Endoplasmic reticulum stress contributes to vitamin E
succinate-induced apoptosis in human gastric cancer SGC-7901 cells.
Cancer Lett. 296:123–131. 2010. View Article : Google Scholar
|
|
39
|
Lee AS: The glucose-regulated proteins:
stress induction and clinical applications. Trends Biochem Sci.
26:504–510. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma Y, Brewer JW, Diehl JA, et al: Two
distinct stress signaling pathways converge upon the CHOP promoter
during the mammalian unfolded protein response. J Mol Biol.
318:1351–1365. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
De Haro C, Méndez R and Santoyo J: The
eIF-2alpha kinases and the control of protein synthesis. FASEB J.
10:1378–1387. 1996.PubMed/NCBI
|
|
42
|
Haze K, Yoshida H, Yanagi T, et al:
Mammalian transcription factor ATF6 is synthesized as a
transmembrane protein and activated by proteolysis in response to
endoplasmic reticulum stress. Mol Biol Cell. 10:3787–3799. 1999.
View Article : Google Scholar
|