Introduction
Glioma is the most common type of primary tumour
found in the central nervous system, where it accounts for 45–55%
of all primary tumours and is a leading cause of mortality in
patients with intracranial tumours (1,2).
Currently, conventional treatments for glioma include surgery,
radiotherapy, and chemotherapy (3,4);
however, the efficacy of these treatments remains poor. In
addition, the molecular mechanisms that result in the occurrence
and development of glioma are not yet well understood. Therefore,
the discovery of molecules with roles in the occurrence and
development of glioma is required to understand the malignant
biological behaviour of glioma. These novel molecules may provide
valuable, reliable molecular targets for future targeted
therapies.
The Ezh2 gene is an important member of the
polycomb-group (PcG) family. PcG genes regulate the transcription
process and thereby play an important role in the regulation of
cell proliferation and the cell cycle. The Ezh2 gene is often
expressed at low levels in normal cells but at high levels in a
variety of stem cells (5–7). In addition, expression of the Ezh2
gene has been found in many tumour cells, including hepatic cancer
(8), breast cancer (9), renal cell carcinoma (10), prostate cancer (11), and lymphoma cells (12). In neuroblastoma, increased
expression of the Ezh2 gene has been shown to enhance the
proliferation, migration, and angiogenesis capacity of the tumour
cells, and these phenotypes could be reduced by the downregulation
of Ezh2 gene expression (13).
These results indicate that the Ezh2 gene plays a key role in
maintaining the growth and invasion of neuroblastoma. Studies have
also shown that the apoptosis of glioma cells is closely related to
the mitochondrial pathway (14,15).
In this study, we showed that the silencing of the Ezh2 gene leads
to changes in the levels of Bax and Bcl-2. The translocations of
Bax and Bcl-2 have been shown to alter the mitochondrial membrane
potential, cause the release of cytochrome c, activate the
caspase family, and eventually lead to apoptosis (16).
We investigated the effects of the downregulation of
the Ezh2 gene on the proliferation, apoptosis, and cell cycle of
human glioma cells. We also explored the signal transduction
pathway induced by the silencing of the Ezh2 gene that results in
apoptosis.
Materials and methods
Cells, antibodies, and reagents
The U87 and U251 human glioma cells were purchased
from the American Type Culture Collection (ATCC; USA).
Lipofectamine™ 2000 was purchased from Invitrogen (USA), the siRNAs
were synthesised by GenePharma (China), the RT-PCR kit [Takara RNA
PCR kit (AMV) Ver. 3.0] was purchased from Takara Biotechnology
(Japan), and propidium iodide was purchased from Sigma (USA). The
antibodies against Ezh2, Bax, Bcl-2, caspase 3 and 9, CDK4, CDK6,
and cyclin D1 were purchased from Cell Signaling Technology, Inc.,
USA.
Cell culture
The U87 and U251 human glioma cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) containing 10% foetal
bovine serum, 100 U/ml penicillin, and 100 μg/ml streptomycin at
37°C with 5% CO2. All of the experiments were performed
with logarithmically growing cells.
Detection of cellular viability
To measure the cellular viability using the MTT
assay, the cells were seeded in 96-well culture plates at a density
of 1×105 cells/ml, and Ezh2 siRNA was added when the
cells were determined to be logarithmically growing. There were 5
groups of 6 wells of cells for each time point (24, 48, 72, 96 and
120 h) within each experiment. After 20 μl of a 5-mg/ml solution of
5-diphenyltetrazolium bromide (MTT) was added to each well, the
cells were cultured for an additional 4 h. Then, the culture medium
was removed, and 150 μl of dimethyl sulphoxide (DMSO) was added to
each well. The cells were subsequently vortexed at room temperature
for 10 min, and the OD value (570 nm) of each well was detected
with a microplate reader.
siRNA transfection
For the siRNA transfections, the cells were seeded
in 6-well plates at a density of 5×105 cells/ml and
cultured in DMEM without antibiotics. The transfection was
performed when the confluence of the cells was ≥60%, and 2
individual groups of cells were either transfected with an Ezh2
siRNA or a non-targeting oligonucleotide control using
Lipofectamine (Invitrogen) according to the manufacturer’s
instructions. The media were changed prior to the transfection with
serum-free DMEM without antibiotics and replaced 4–6 h after the
transfection with serum-containing DMEM medium. The sequence of the
Ezh2 siRNA was 5′-AAG ACT CTG AAT GCA GTT GCT-3′, and a
non-targeting siRNA served as a negative control. The efficiency of
the siRNA was determined using reverse transcription polymerase
chain reaction (RT-PCR) and western blotting.
RNA extraction and RT-PCR
The extraction of the total-RNA was performed using
an RNAiso™ Plus kit (Takara) according to the manufacturer’s
instructions. After calculating the RNA concentration, the RT-PCR
was performed using an RT-PCR kit (Takara) according to the product
manual. The primers for the Ezh2 and β-actin RT-PCR reaction were
synthesised by Invitrogen and were as follows: Ezh2, forward primer
5′-GCC AGA CTG GGA AGA AAT CTG-3′ and reverse primer 5′-TGT GCT GGA
AAA TCC AAG TCA-3′; β-actin, forward primer 5′-CTG GGA CGA CAT GGA
GAA AA-3′ and reverse primer 5′-AAG GAA GGC TGG AAG AGT GC-3′. The
PCR reaction was then performed in a 50-μl volume with the
following reaction conditions: an initial denaturing step at 94°C
for 2 min and 30 cycles of denaturing at 94°C for 30 sec, annealing
at 60°C for 30 sec, and extension at 72°C for 30 sec. The PCR
products were separated on a 1.0% agarose gel by electrophoresis
and analysed with a gel imaging scanning system.
Detection of apoptosis by flow
cytometry
The cells were collected by trypsinisation and
washed twice in ice-cold PBS. Next, the cells were resuspended in
PBS, and a single-cell suspension was prepared by pipetting the
medium up and down. An Annexin-V and PI staining solution was then
added according to the manufacturer’s instructions (Annexin-V-FITC
kit, Biosea Biotechnology, China). The cells were stained for 15
min in the dark at room temperature, and the apoptotic cells were
detected using a flow cytometer (Becton-Dickinson, USA).
Detection of mitochondrial membrane
potential
JC-1 staining and flow cytometry were used to detect
changes in the mitochondrial membrane potential according to a
previously published protocol (17). The fluorescence signals of the JC-1
monomer and polymer were detected by the FL1 and FL2 detectors,
respectively. FL1-H and FL2-H represent the green and red
fluorescence intensities, respectively. CellQuest software was used
for the quantification of the results.
Cell cycle analysis by flow
cytometry
For the cell cycle analysis, the cells were
trypsinised with 0.25% trypsin, collected, washed twice in PBS, and
fixed with a 70% ethanol solution at 4°C overnight. The following
day, the ethanol was discarded. Subsequently, the cells were washed
twice with PBS, stained with 1 ml of PI dye that contained 10 μg of
RNaseA and 5 μl of Triton X-100 for 30 min at 4°C in the dark, and
analysed by flow cytometry.
Western blotting
For the western blotting, the transfected U87 cells
from each group were collected and washed twice in PBS. Then, 2 ml
of lysis buffer (50 mM Tris-HCl pH 7.4, 137 mM NaCl, 10% glycerol,
100 mM sodium vanadate, 1 mM PMSF, 10 mg/ml aprotinin, 10 mg/ml
leupeptin, 1% NP-40, and 5 mM cocktail) was added to the cells. The
protein concentration was determined using the BCA method, and the
proteins were stained with bromophenol blue. Equal amounts of
protein were loaded and separated on a 10% polyacrylamide gel by
SDS-PAGE electrophoresis, and the proteins were transferred onto a
PVDF membrane using the semi-dry method. The membrane was then
blocked with 5% non-fat dry milk overnight. The following day, the
membrane was washed with TBST, incubated for 2 h with the primary
antibodies, washed with TBST, and incubated for an additional 2 h
with the secondary antibody. After adding the chemiluminescence
reagent, X-ray autoradiography was performed, the bands were
scanned, and the gray scale images were analysed.
Statistical analysis
SPSS 16.0 statistical software was used for the
statistical analysis. The values are shown as the mean ± SD.
Statistical analysis was performed using the Student’s t-test, and
the differences between the groups were considered to be
statistically significant at p<0.05.
Results
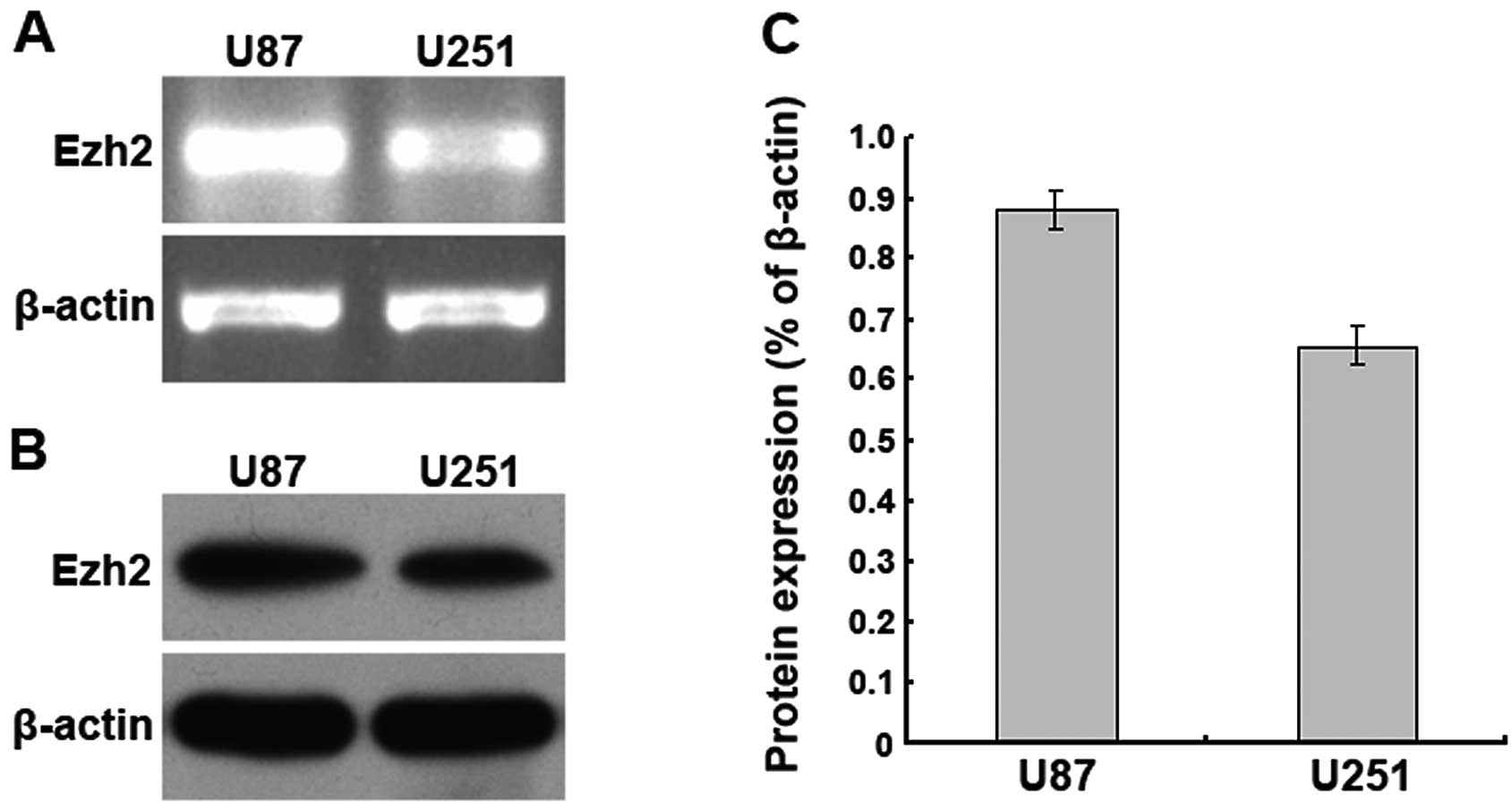
The Ezh2 gene is highly expressed in U87
human glioma cells
The Ezh2 mRNA and protein expression levels in U87
and U251 human glioma cells were examined using RT-PCR and western
blotting, respectively. We found that Ezh2 mRNA was highly
expressed in the U87 and U251 cells, and a higher level was present
in the U87 cells. Additionally, western blot analysis demonstrated
that the expression levels of the Ezh2 protein in the U87 and U251
cells correlated with the mRNA expression levels. These results
suggest that Ezh2 is highly expressed in the U87 and U251 human
glioma cells (Fig. 1).
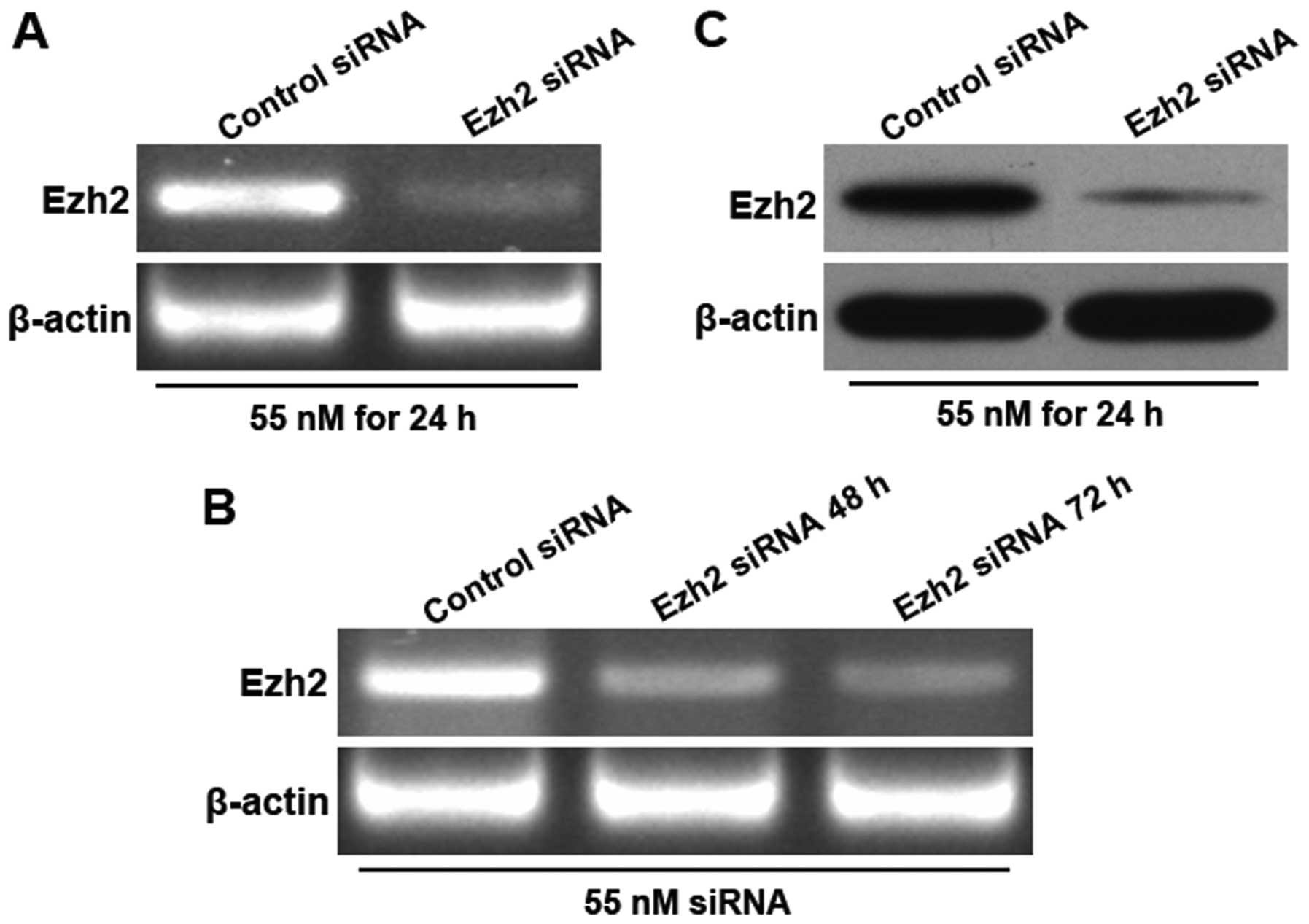
Downregulation of Ezh2 expression in U87
human glioma cells by RNA interference
An siRNA targeting Ezh2 and a non-targeting
oligonucleotide were individually transfected into U87 cells, and
the changes in the Ezh2 mRNA and protein expression levels after
transfection were analysed by RT-PCR and western blotting,
respectively. The results demonstrated that at 24 h after the
transfection of the Ezh2 siRNA (55 nM), the Ezh2 mRNA expression
level was significantly reduced. The silencing of Ezh2 lasted for
at least 72 h after the siRNA transfection (Fig. 2). These results suggest that after
the transfection of the siRNA targeting Ezh2 at 55 nM for 24 h, the
Ezh2 mRNA and protein expression levels are effectively
downregulated.
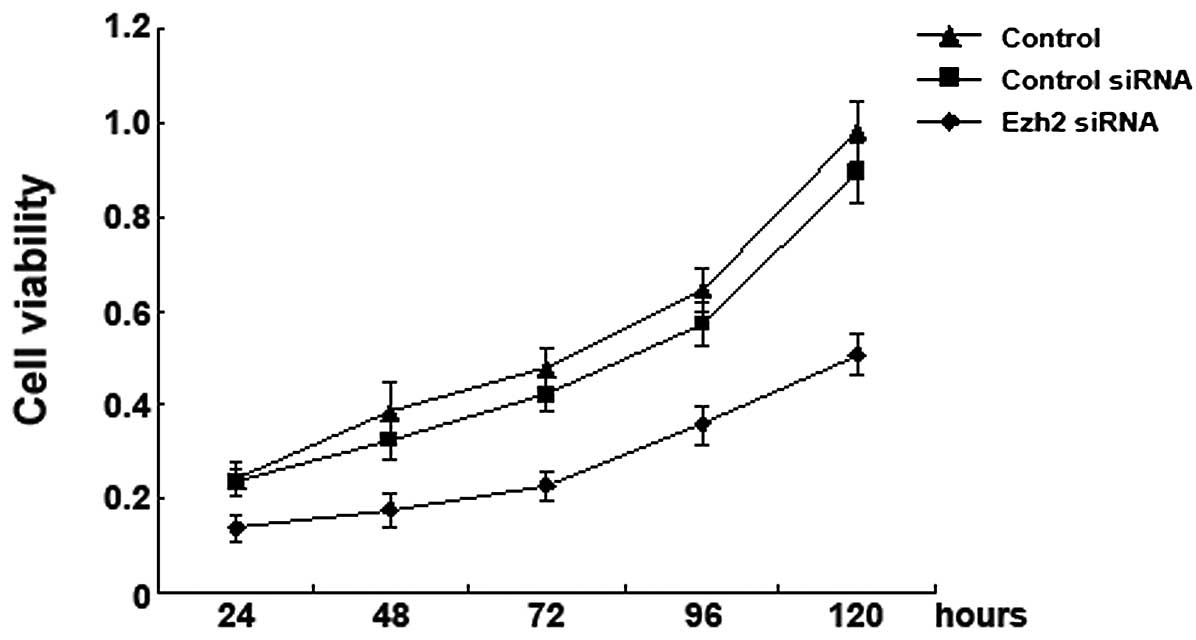
Downregulation of Ezh2 expression
inhibits the proliferation of U87 human glioma cells
MTT assay was used to determine the effects of the
downregulation of Ezh2 expression on the proliferation of U87
cells. We found that compared to the mock-transfected cells or the
cells transfected with the non-targeting siRNA, the proliferation
of the U87 cells transfected with the Ezh2 siRNA was significantly
reduced at 24, 48, 72, 96, or 120 h after the transfection. These
results suggest that the silencing of the Ezh2 gene inhibits the
proliferation of U87 cells (Fig.
3).
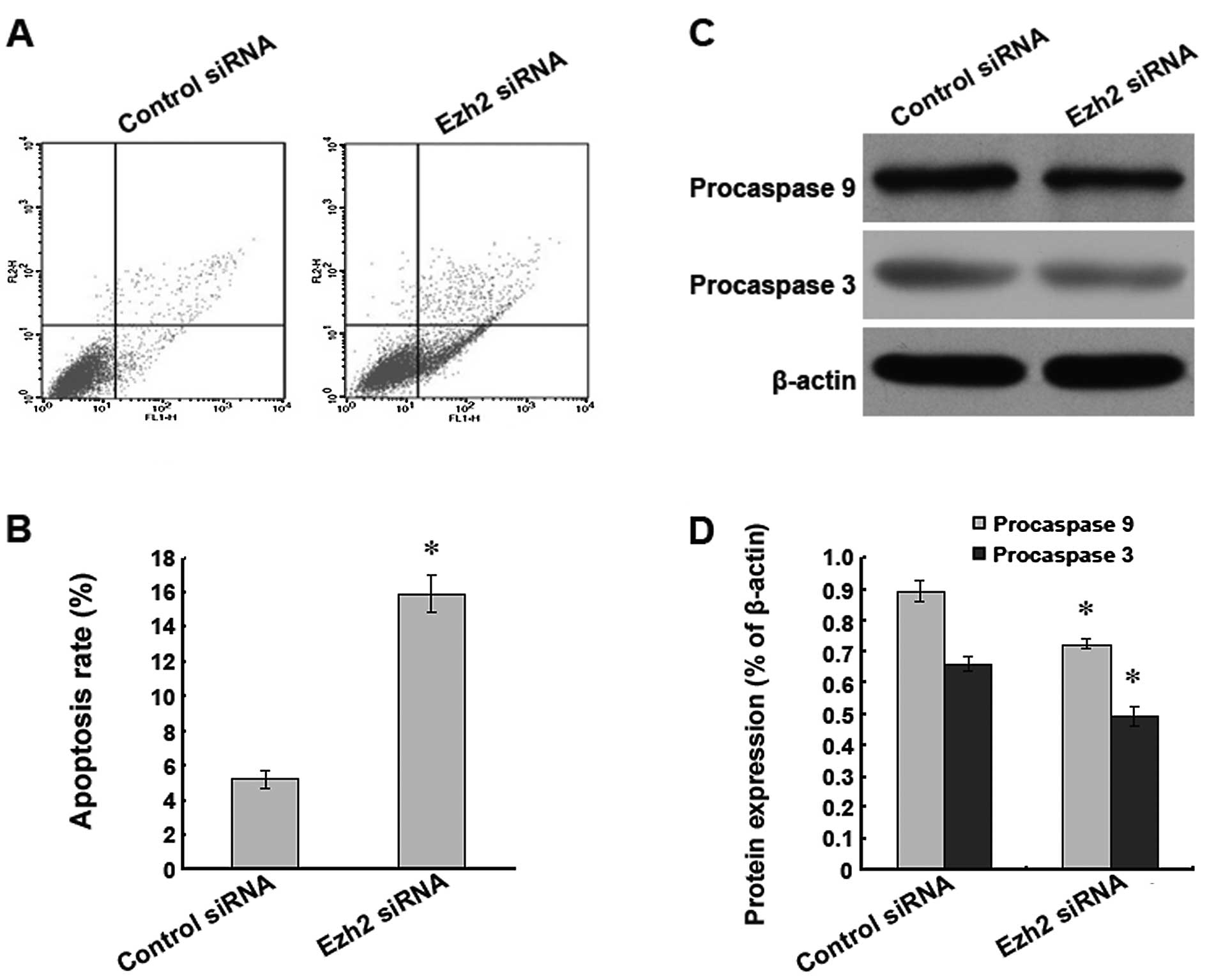
Downregulation of Ezh2 expression induces
apoptosis in U87 human glioma cells
To investigate whether the level of Ezh2 expression
was related to the level of apoptosis found in the U87 cells, the
expression of the Ezh2 gene was downregulated by RNA interference,
and the indicators of apoptosis were analysed. We found that after
the silencing of the Ezh2 gene, a significantly higher rate of
apoptosis was present in the U87 cells as detected by flow
cytometry (Fig. 4A and B). In
addition, western blot analysis indicated that the protein
expression levels of procaspase 9 and 3 were significantly reduced
after the downregulation of Ezh2 expression (Fig. 4C and D). These results suggest that
the silencing of the Ezh2 gene leads to apoptosis in U87 cells.
Downregulation of Ezh2 expression induces
apoptosis in the U87 human glioma cells through the mitochondrial
pathway
To verify whether the apoptosis induced by the
silencing of the Ezh2 gene in the U87 cells was related to the
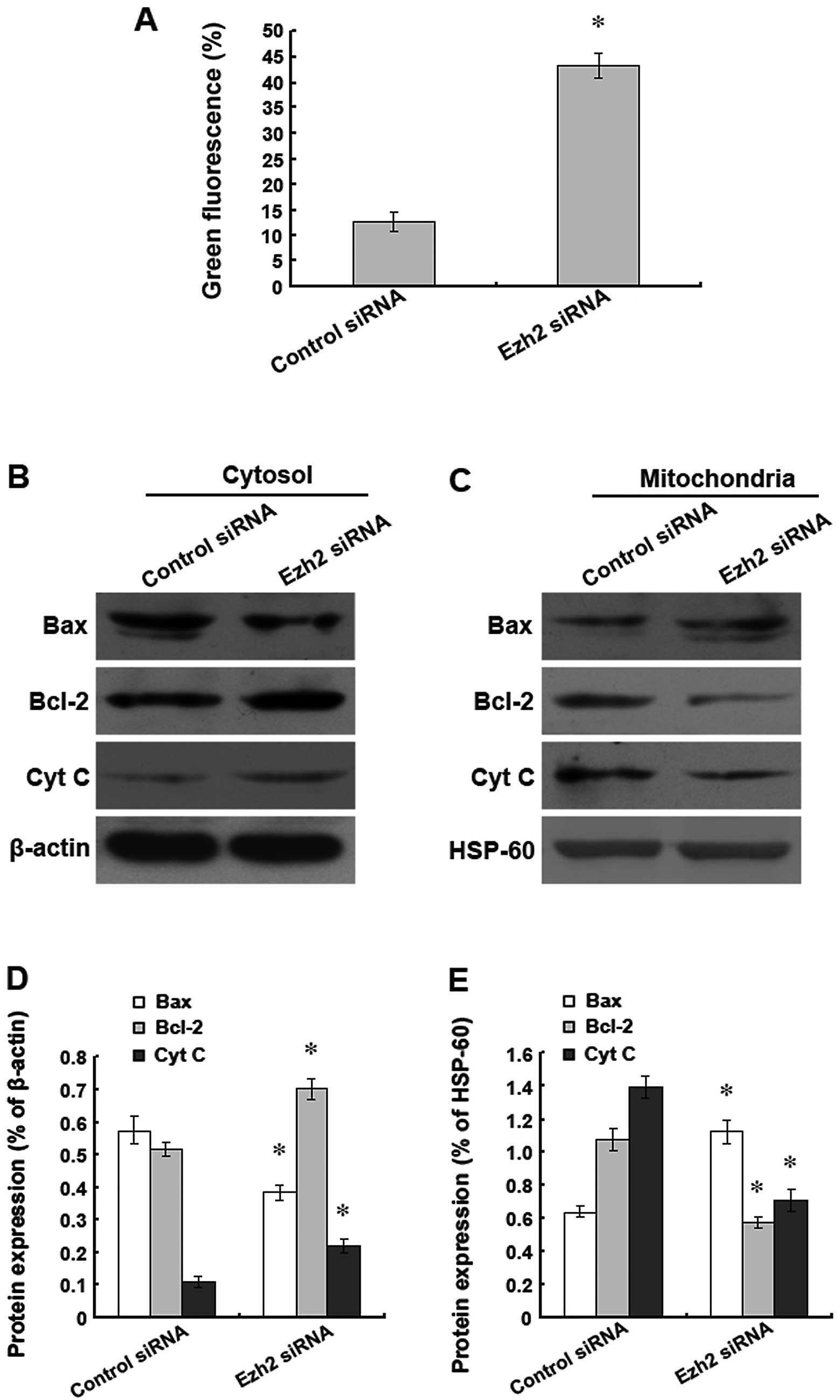
mitochondrial pathway, the changes in the mitochondrial membrane
potential were detected using JC-1 staining. The results
demonstrated that the downregulation of Ezh2 expression
significantly reduced the mitochondrial membrane potential
(Fig. 5A). In addition, the levels
of Bax, Bcl-2, and cytochrome c were analysed by western
blotting. The silencing of Ezh2 expression decreased the level of
Bax and increased the levels of Bcl-2 and cytochrome c in
the cytoplasm. However, the changes in the levels of Bax, Bcl-2,
and cytochrome c in the mitochondria were the opposite of
the changes found in the cytoplasm (Fig. 5B–E). Therefore, our data indicate
that the downregulation of Ezh2 expression promotes the
translocation of Bax and Bcl-2, causes the release of mitochondrial
cytochrome c, reduces the mitochondrial membrane potential,
and results in the apoptosis of human glioma cells.
Downregulation of Ezh2 expression causes
a cell cycle arrest in the G0/G1 phase in U87 human glioma
cells
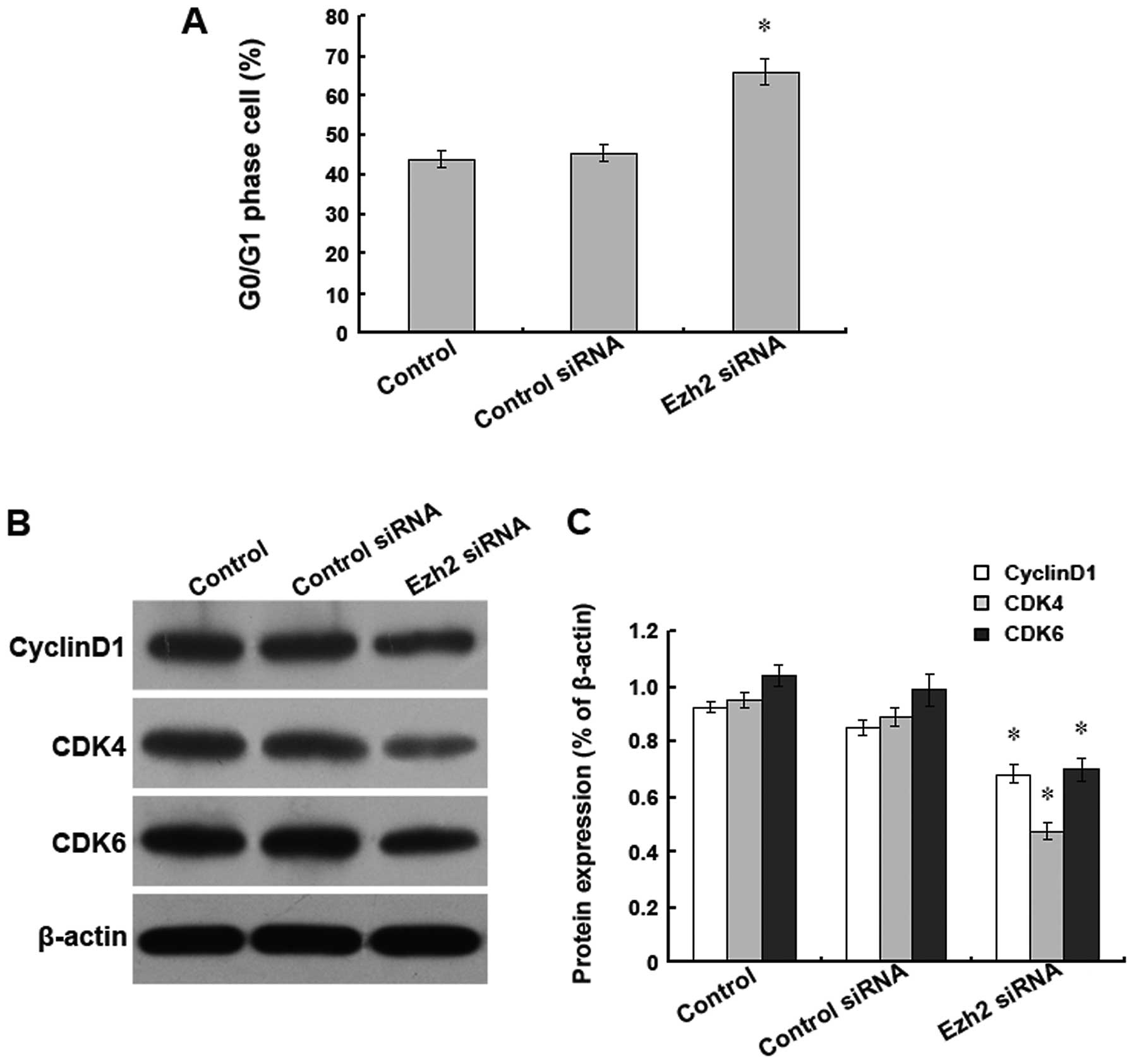
To further investigate the inhibition of
proliferation caused by Ezh2 siRNA treatment, U87 cells were
individually transfected with 55 nM Ezh2 siRNA or a non-targeting
siRNA as a negative control, and the cell cycle was analysed by
flow cytometry. We found that upon Ezh2 siRNA treatment, the
percentage of cells in the G0/G1 phase was significantly higher
than the percentage found in the mock-transfected cells or the
cells treated with the non-targeting siRNA (Fig. 6A). Additionally, we found that the
downregulation of Ezh2 expression altered the levels of several
cell cycle proteins, including cyclin D1, CDK4, and CDK6, all of
which have roles in the G0/G1 phase. Western blot analysis revealed
that compared with the mock-transfected cells or the cells
transfected with the non-targeting siRNA, the protein expression
levels of cyclin D1, CDK4, and CDK6 were significantly reduced in
the cells transfected with the Ezh2 siRNA (Fig. 6B and C). These results suggest that
the inhibition of proliferation caused by the downregulation of
Ezh2 expression in the U87 cells may be mediated by the reduced
expression levels of cyclin D1, CDK4, and CDK6, which result in an
arrest of the cell cycle in the G0/G1 phase.
Discussion
In the present study, we explored the role of the
Ezh2 gene for the treatment of human glioma. We demonstrated that
Ezh2 was highly expressed both at the mRNA and protein level in
human glioma cells, which suggests that the Ezh2 gene is closely
related to the occurrence and development of glioma. Additionally,
we hypothesised that the Ezh2 gene may serve as a potential
biomarker and therapeutic target for the treatment of glioma. To
further elucidate the detailed mechanism of the function of Ezh2 in
human glioma, we used RNA interference to downregulate the
expression of Ezh2 and observe the resulting changes in human
glioma cells. We found that the Ezh2 mRNA and protein expression
levels were significantly reduced with the siRNA treatment, and the
downregulation of Ezh2 expression led to a reduction in the
proliferation of the glioma cells. These data suggest that the Ezh2
gene plays a role in promoting the proliferation of glioma cells.
Consistent with our findings, previous studies have also shown that
the Ezh2 gene promotes the proliferation of lung (18), breast (9), thyroid (19), colon (20), ovarian (21), and pancreatic cancer cells (22). Therefore, we concluded that the Ezh2
gene plays an important role in the regulation of the proliferation
of glioma cells.
To clarify how the downregulation of Ezh2 expression
inhibited the proliferation of glioma cells, we used flow cytometry
to analyse the cell cycle and determine the level of apoptosis. Our
results demonstrated that the silencing of Ezh2 expression led to
an induction of apoptosis in the glioma cells. To date, research
regarding the role of Ezh2 in the induction of apoptosis is very
limited. One study demonstrated that the silencing of the Ezh2 gene
led to increased levels of apoptosis in renal cancer cells
(23). In addition, the suppression
of Ezh2 expression has been reported to cause apoptosis by
downregulating the expression levels of Bax and caspase 3 (24). Consistent with these studies, we
also found that the reduced proliferation caused by Ezh2 siRNA
treatment correlated with an increased level of apoptosis in the
glioma cells. We also found that the downregulation of Ezh2
expression caused an arrest of the cell cycle in the G0/G1 phase
and a reduction in the protein levels of the cell cycle regulatory
proteins, cyclin D1, CDK4, and CDK6. These results indicate that
the Ezh2 gene is essential for cellular proliferation, and the
downregulation of Ezh2 expression leads to a reduced number of
dividing glioma cells, which is consistent with previous reports
(25,26).
Tumour cells can undergo apoptosis through the
mitochondrial pathway or the death receptor pathway (27). We found that in glioma cells, the
silencing of Ezh2 expression led to the translocation of Bax and
Bcl-2, which was followed by a decrease in the mitochondrial
membrane potential and the release of cytochrome c. These
results suggest that the apoptosis of glioma cells induced by the
downregulation of Ezh2 expression is dependent on the mitochondrial
pathway. Bcl-2 and Bax play key regulatory roles in the
mitochondrial apoptosis pathway (28). When Bax translocates from the
cytoplasm to the mitochondrial membrane, it changes the
permeability of the mitochondrial membrane and promotes the release
of cytochrome c from the mitochondria into the cytoplasm
(29) to initiate the events that
lead to cellular apoptosis. The activation of the caspase family is
necessary to induce apoptosis. Herein, we analysed the protein
levels of procaspase 9 and procaspase 3 after Ezh2 siRNA treatment
and found that the levels of these proteins were significantly
reduced. Cytochrome c released into the cytoplasm activates
caspase 9 and caspase 3, which play key roles in the apoptosis
pathway (30). Therefore, these
results suggest that the downregulation of Ezh2 expression induces
apoptosis through the mitochondrial pathway in glioma cells.
Collectively, we demonstrated that the Ezh2 gene is
highly expressed in human glioma cells, and the downregulation of
Ezh2 expression induces apoptosis through the mitochondrial pathway
by regulating the Bcl-2/Bax family in human glioma cells.
Furthermore, this study provides a new approach for the clinical
treatment of glioma.
Acknowledgements
We thank Professor Luo Yinan from the First Hospital
of Jilin University and Professor Li Shulei from the College of
Medicine of Jilin University for their guidance of this study.
References
|
1
|
Johannesen TB, Langmark F and Lote K:
Cause of death and long-term survival in patients with
neuro-epithelial brain tumours: a population-based study. Eur J
Cancer. 39:2355–2363. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hess KR, Broglio KR and Bondy ML: Adult
glioma incidence trends in the United States, 1977–2000. Cancer.
101:2293–2299. 2004.PubMed/NCBI
|
|
3
|
Stummer W and Kamp MA: The importance of
surgical resection in malignant glioma. Curr Opin Neurol.
22:645–649. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Norden AD and Wen PY: Glioma therapy in
adults. Neurologist. 12:279–292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
De Haan G and Gerrits A: Epigenetic
control of hematopoietic stem cell aging the case of Ezh2. Ann NY
Acad Sci. 1106:233–239. 2007.PubMed/NCBI
|
|
6
|
Venneti S, Le P, Martinez D, Xie SX,
Sullivan LM, Rorke-Adams LB, Pawel B and Judkins AR: Malignant
rhabdoid tumors express stem cell factors, which relate to the
expression of EZH2 and Id proteins. Am J Surg Pathol. 35:1463–1472.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Juan AH, Derfoul A, Feng X, Ryall JG,
Dell’Orso S, Pasut A, Zare H, Simone JM, Rudnicki MA and Sartorelli
V: Polycomb EZH2 controls self-renewal and safeguards the
transcriptional identity of skeletal muscle stem cells. Genes Dev.
25:789–794. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yonemitsu Y, Imazeki F, Chiba T, Fukai K,
Nagai Y, Miyagi S, Arai M, Aoki R, Miyazaki M, Nakatani Y, et al:
Distinct expression of polycomb group proteins EZH2 and BMI1 in
hepatocellular carcinoma. Hum Pathol. 40:1304–1311. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Reijm EA, Jansen MP, Ruigrok-Ritstier K,
van Staveren IL, Look MP, van Gelder ME, Sieuwerts AM, Sleijfer S,
Foekens JA and Berns EM: Decreased expression of EZH2 is associated
with upregulation of ER and favorable outcome to tamoxifen in
advanced breast cancer. Breast Cancer Res Treat. 125:387–394. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hinz S, Weikert S, Magheli A, Hoffmann M,
Engers R, Miller K and Kempkensteffen C: Expression profile of the
polycomb group protein enhancer of Zeste homologue 2 and its
prognostic relevance in renal cell carcinoma. J Urol.
182:2920–2925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Varambally S, Dhanasekaran SM, Zhou M,
Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt
RG, Otte AP, et al: The polycomb group protein EZH2 is involved in
progression of prostate cancer. Nature. 419:624–629. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Visser HP, Gunster MJ, Kluin-Nelemans HC,
Manders EM, Raaphorst FM, Meijer CJ, Willemze R and Otte AP: The
Polycomb group protein EZH2 is upregulated in proliferating,
cultured human mantle cell lymphoma. Br J Haematol. 112:950–958.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smits M, Nilsson J, Mir SE, van der Stoop
PM, Hulleman E, Niers JM, de Witt Hamer PC, Marquez VE, Cloos J,
Krichevsky AM, et al: miR-101 is down-regulated in glioblastoma
resulting in EZH2-induced proliferation, migration, and
angiogenesis. Oncotarget. 1:710–720. 2010.PubMed/NCBI
|
|
14
|
Tomiyama A, Tachibana K, Suzuki K, Seino
S, Sunayama J, Matsuda KI, Sato A, Matsumoto Y, Nomiya T, Nemoto K,
et al: MEK-ERK-dependent multiple caspase activation by
mitochondrial proapoptotic Bcl-2 family proteins is essential for
heavy ion irradiation-induced glioma cell death. Cell Death Dis.
1:e602010. View Article : Google Scholar
|
|
15
|
Qiao S, Murakami K, Zhao Q, Wang B, Seo H,
Yamashita H, Li X, Iwamoto T, Ichihara M and Yoshino M:
Mimosine-induced apoptosis in C6 glioma cells requires the release
of mitochondria-derived reactive oxygen species and p38, JNK
activation. Neurochem Res. 37:417–427. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu HF, Chie YJ, Yang MS, Lee CS, Fu JJ,
Yang JS, Tan TW, Wu SH, Ma YS, Ip SW and Chung JG: Apigenin induces
caspase-dependent apoptosis in human lung cancer A549 cells through
Bax- and Bcl-2-triggered mitochondrial pathway. Int J Oncol.
36:1477–1484. 2010.PubMed/NCBI
|
|
17
|
Tang B, Zhang Y, Liang R, Yuan P, Du J,
Wang H and Wang L: Activation of the δ-opioid receptor inhibits
serum deprivation-induced apoptosis of human liver cells via the
activation of PKC and the mitochondrial pathway. Int J Mol Med.
28:1077–1085. 2011.
|
|
18
|
Dang X, Ma A, Yang L, Hu H, Zhu B, Shang
D, Chen T and Luo Y: MicroRNA-26a regulates tumorigenic properties
of EZH2 in human lung carcinoma cells. Cancer Genet. 205:113–123.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Esposito F, Tornincasa M, Pallante P,
Federico A, Borbone E, Pierantoni GM and Fusco A: Down-regulation
of the miR-25 and miR-30d contributes to the development of
anaplastic thyroid carcinoma targeting the polycomb protein EZH2. J
Clin Endocrinol Metab. 97:E710–E718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fussbroich B, Wagener N, Macher-Goeppinger
S, Benner A, Fälth M, Sültmann H, Holzer A, Hoppe-Seyler K and
Hoppe-Seyler F: EZH2 depletion blocks the proliferation of colon
cancer cells. PLoS One. 6:e216512011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Cai Q, Godwin AK and Zhang R:
Enhancer of zeste homolog 2 promotes the proliferation and invasion
of epithelial ovarian cancer cells. Mol Cancer Res. 8:1610–1618.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ougolkov AV, Bilim VN and Billadeau DD:
Regulation of pancreatic tumor cell proliferation and
chemoresistance by the histone methyltransferase enhancer of zeste
homologue 2. Clin Cancer Res. 14:6790–6796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wagener N, Holland D, Bulkescher J,
Crnković-Mertens I, Hoppe-Seyler K, Zentgraf H, Pritsch M, Buse S,
Pfitzenmaier J, Haferkamp A, et al: The enhancer of zeste homolog 2
gene contributes to cell proliferation and apoptosis resistance in
renal cell carcinoma cells. Int J Cancer. 123:1545–1550. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu ZL, Zheng SS, Li ZM, Qiao YY, Aau MY
and Yu Q: Polycomb protein EZH2 regulates E2F1-dependent apoptosis
through epigenetically modulating Bim expression. Cell Death
Differ. 17:801–810. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu SC and Zhang Y: Cyclin-dependent kinase
1 (CDK1)-mediated phosphorylation of enhancer of zeste 2 (Ezh2)
regulates its stability. J Biol Chem. 286:28511–28519. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen S, Bohrer LR, Rai AN, Pan Y, Gan L,
Zhou X, Bagchi A, Simon JA and Huang H: Cyclin-dependent kinases
regulate epigenetic gene silencing through phosphorylation of EZH2.
Nat Cell Biol. 12:1108–1114. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
von Haefen C, Wendt J, Semini G, Sifringer
M, Belka C, Radetzki S, Reutter W, Daniel PT and Danker K:
Synthetic glycosidated phospholipids induce apoptosis through
activation of FADD, caspase-8 and the mitochondrial death pathway.
Apoptosis. 16:636–651. 2011.
|
|
28
|
Mattson MP and Kroemer G: Mitochondria in
cell death: novel targets for neuroprotection and cardioprotection.
Trends Mol Med. 9:196–205. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saito M, Korsmeyer SJ and Schlesinger PH:
BAX-dependent transport of cytochrome c reconstituted in pure
liposomes. Nat Cell Biol. 2:553–555. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|