Introduction
Mesenchymal stem cells (MSCs) are marrow-derived
non-hematopoietic precursor cells that contribute to the
maintenance and regeneration of connective tissues through
engraftment (1). In vivo
engraftment is not only an intrinsic function of MSCs but also
depends on appropriate external signals produced by the tissue
microenvironment (2). Over the
years, the correlation between bone marrow-derived stem cells and
cancers has been increasingly emphasized. Cancer is increasingly
being viewed as a stem cell disease (3), and a large body of convincing evidence
has shown MSCs can home to the tumor site and play an important
role in tumor progression (1,4–6). The
mechanisms of action of MSCs are related to tumor angiogenesis
(7) and immunosuppression (8).
Hepatocellular carcinoma (HCC) is a lethal
malignancy with an extremely poor prognosis due to a high rate of
tumor recurrence after surgery and intra-hepatic metastases
(9,10). It has been reported that bone marrow
stem cells are a source of liver oval cells (11), and that transplanted bone marrow can
regenerate liver cells (12,13).
In our previous study, we found that MSC injection inhibited the
pulmonary metastasis of HCC (14).
However, the distribution and mechanisms of action of MSCs after
injection have not been well documented and the majority of studies
have concentrated on benign disease.
Cell fusion is a highly regulated and dramatic
cellular event that is required for development and homeostasis. It
has been reported that cell fusion events involving bone
marrow-derived cells (BMDCs) commonly occur after different types
of tissue damage and play a crucial role in tissue restoration
(15,16). A number of studies have suggested
that cell fusion is the main mechanism of action after stem cell
transplantation other than transdifferentiation. Moreover, the
broad differentiation potential of bone marrow cells in most cases
is a consequence of cell fusion (15,17).
Therefore, taking into account the above data, we
hypothesized that MSCs may affect HCC progression by fusing with
cancer cells. In this study, we injected labeled MSCs into mice
with HCC, in order to observe the in vivo distribution of
MSCs and the cell fusion between MSCs and HCC cells.
Materials and methods
Cell lines
The MSC cell line was obtained from ScienCell
Research Laboratories (Carlsbad, CA, USA), which was isolated from
human bone marrow, and characterized by immunofluorescence with
CD44 and CD90 antibodies, and lipid staining after differentiation.
The fifth passage MSCs did not express the surface marker, CD34;
they expressed low levels of fetal liver kinase-1 (Flk-1) and
higher levels of CD29 and CD105. Quantitative RT-PCR showed that
the MSCs expressed octamer-binding transcription factor-4 (OCT-4)
and Flk-1. They were cultured in Alpha Minimum Essential Medium
(α-MEM, Gibco) supplemented with 10% fetal bovine serum (FBS,
Gibco) and 100 U/ml penicillin/streptomycin solution. The fifth to
eighth passage cells were used in the following experiments.
MHCC97-H is human HCC cell line with a higher
metastatic potential (18,19). These cells were cultured in high
glucose Dulbecco’s modified Eagle’s medium (H-DMEM, Gibco),
supplemented with 10% FBS at 37°C in a humidified incubator
containing 5% CO2.
Labeling of MSCs and liver cancer
cells
MHCC97-H cells were labeled with green fluorescence
protein (GFP) by transfection with the plasma vector, pEGFP-N1
(Clontech). Lipofectamine 2000 which mediated the highest
transfection rates was used as a transfection agent. After 2 weeks
of selection with G418 (800 μg/ml), the individual G418-resistant
clones were picked up and subcultured. Finally, a stable eukaryotic
cell line with the highest fraction of EGFP expression was obtained
(GFP-MHCC97-H).
MSCs were labeled with red fluorescence protein
(RFP) and GFP by transfection with the plasma vector, pERFP-N1
(Clontech), and after 2 weeks of selection with G418 (400 μg/ml),
stable RFP-MSCs and GFP-MSCs were then acquired, respectively.
MSCs were labeled with 4′,6-diamidino-2-phenylindole
(DAPI, Vector Laboratories) according to the manufacturer’s
instructions.
MSCs were also labeled with 5-bromodeoxyuridine
(BrdU, Sigma) according to the manufacturer’s intuductions. For
BrdU labeling, MSCs were cultured for 15 min in the presence of 2
mM BrdU, washed and expanded for 2 additional passages before being
injected into the mice. This procedure labeled 70–80% of the
MSCs.
Co-culture of MSCs and liver cancer
cells
To evaluate the in vitro cell fusion between
MSCs and HCC cells, the GFP-MHCC97-H cells were co-cultured with
DAPI-MSCs at a ratio of 5:1 in a 6-well plate. After 4 days, the
culture was observed under a fluorescence microscope. Similarly,
GFP-MHCC97-H cells and RFP-MSCs were also co-cultured at a ratio of
5:1 and observed under a microscope.
An Axioplan Epifluorescence microscope (Carl Zeiss,
Oberkochen, Germany) was used and images were obtained using a
DC300 digital video camera (Leica). Optical images were acquired
using a DMR microscope connected to a DC300 video camera
(Leica).
Co-culture of GFP-MSCs and RFP-MSCs
RFP-MSCs and GFP-MSCs were co-cultured at a ratio of
1:1 in a 6-well plate. The culture was observed under a
fluorescence microscope to evaluate the in vitro cell fusion
of the MSCs.
Cytoimmunochemistry
MSCs (2×105) were plated and cultured in
6-well plate. When the cells had reached 60% confluency, they were
fixed with 100% methanol, permeabilized with 0.5% Triton X-100, and
sequentially incubated with primary anti-matrix metalloproteinase
(MMP)2 or primary anti-MMP9 monoclonal antibodies and anti-mouse
immunoglobulin (Ig) coupled with horseradish peroxidase (HRP). The
cells were then stained with 3,3′-diaminobenzidine (DAB) and
counterstained with hematoxylin.
Transwell assay for in vitro migration of
MSCs
In vitro invasion assay was performed as
follows: briefly, 80 μl of serum-free α-MEM-diluted Matrigel (0.8
mg/ml) was added to the Transwell filters (8.0μm pore size) of a
Boyden chamber (Costar, MA, USA) and incubated at 37°C for 2 h to
form matrix gel. MSCs (1×106) were cultured in FBS-free
α-MEM for 24 h, and subsequently the cells were collected and
counted. Cells (2×105) were re-suspended with α-MEM and
seeded in the upper well of Transwell chamber, a mixture of 600 μl
of α-MEM with 10% fetal calf serum (FCS) was added to the lower
chamber, serving as the chemoattractant. After incubation at 37°C
for 48 h, the cells that had invaded across the Matrigel and passed
through the Transwell filter were stained and observed under a
light microscope.
Gelatin zymography detection of MMP2 and
MMP9 activities
Equal amounts of protein from the MSCs, MHCC97-H and
MHCC97-L cells (HCC cells with lower metastasis) were mixed with
SDS buffer and incubated for 20 min at 37°C. After incubation,
samples (30 μg/lane) were added onto a 4.5% (w/v) stacking
polyacrylamide gel and separated on a 7.5% (w/v) polyacrylamide gel
containing 1 mg/ml gelatin for the detection of MMP2 and MMP9
activities. After electrophoresis, the gels were soaked in 2.5%
Triton X-100 for 1 h to remove SDS and incubated for 16 h at 37°C
in 50 mM Tris-HCl (pH 7.6) containing 150 mM NaCl, 10 mM
CaCl2 and 0.02% NaN3. Finally, the gels were
stained for 1 h in 45% methanol/10% acetic acid containing 0.5%
Coomassie brilliant blue G250. Proteolytic activity was detected as
clear bands on a blue background of the Coomassie blue staining
gel.
In vivo visualization of MSCs
To detect the distribution of MSCs in vivo,
we subcutaneously injected 6×106 GFP- MHCC97-H cells
into nude mice (n=4). When the nodular tumors were formulated, we
injected 5×105 of human DAPI-MSCs into the tail veins of
the mice. Four days after the injection, the mice were sacrificed,
the subcutaneous tumor tissues, livers and lungs were removed and
embedded with 1:4 dilution of optimum cutting temperature (OCT)
compound in phosphate-buffered saline (PBS). Fresh-frozen tumor
sections (5-μm thick) were mounted on glass slides, and the
distribution of DAPI-MSCs was observed under a fluorescence
microscope. These experiments were approved by the Shanghai Medical
Experimental Animal Care Commission.
We also orthotopically implanted tissues of
subcutaneous tumor into the livers of 10 nude mice, and 15 days
after the implantation, 5 out of the 10 mice were intravenously
injected with 5×105 of human BrdU-MSCs 3 times per week.
The other 5 mice were injected with PBS as the controls. After 20
days, the tumors, livers and lungs were removed and fixed in
paraformaldehyde and embedded in paraffin wax. Paraffin sections
(5-μm-thick) were mounted on glass slides.
The tumor and liver slides were deparaffinized and
rehydrated over 10 min through a graded alcohol series to deionized
water. Subsequently, 1% Antigen Unmasking Solution (Vector
Laboratories) was added and the slides were microwaved to enhance
antigen retrieval, followed by immunostaining with primary mouse
anti-human antibody against BrdU (Sigma). Goat anti-mouse
IgG-peroxidase (A9917, Sigma) or Cy3-Goat anti-mouse IgG were used
as the secondary antibodies.
Analysis of pulmonary metastasis
Lung samples were sliced into 20 sections of 5 μm
thickness, and a 50-μm interval between 2 successive sections.
After staining with hematoxylin and eosin (H&E), the sections
were independently observed under a microscope by 2 pathologists to
evaluate pulmonary metastasis.
Statistical analysis
The data were analyzed using SPSS 11.5 software
(SPSS Corp., Chicago, IL). The Student’s t-test was used to analyze
the differences in tumor weight. Fisher’s exact test was used for
the comparison of the ratio involved. All statistical tests were
two-sided and P<0.05 was considered to indicate a statistically
significant difference.
Results
MSCs fuse with HCC cells in vitro at a
low frequency
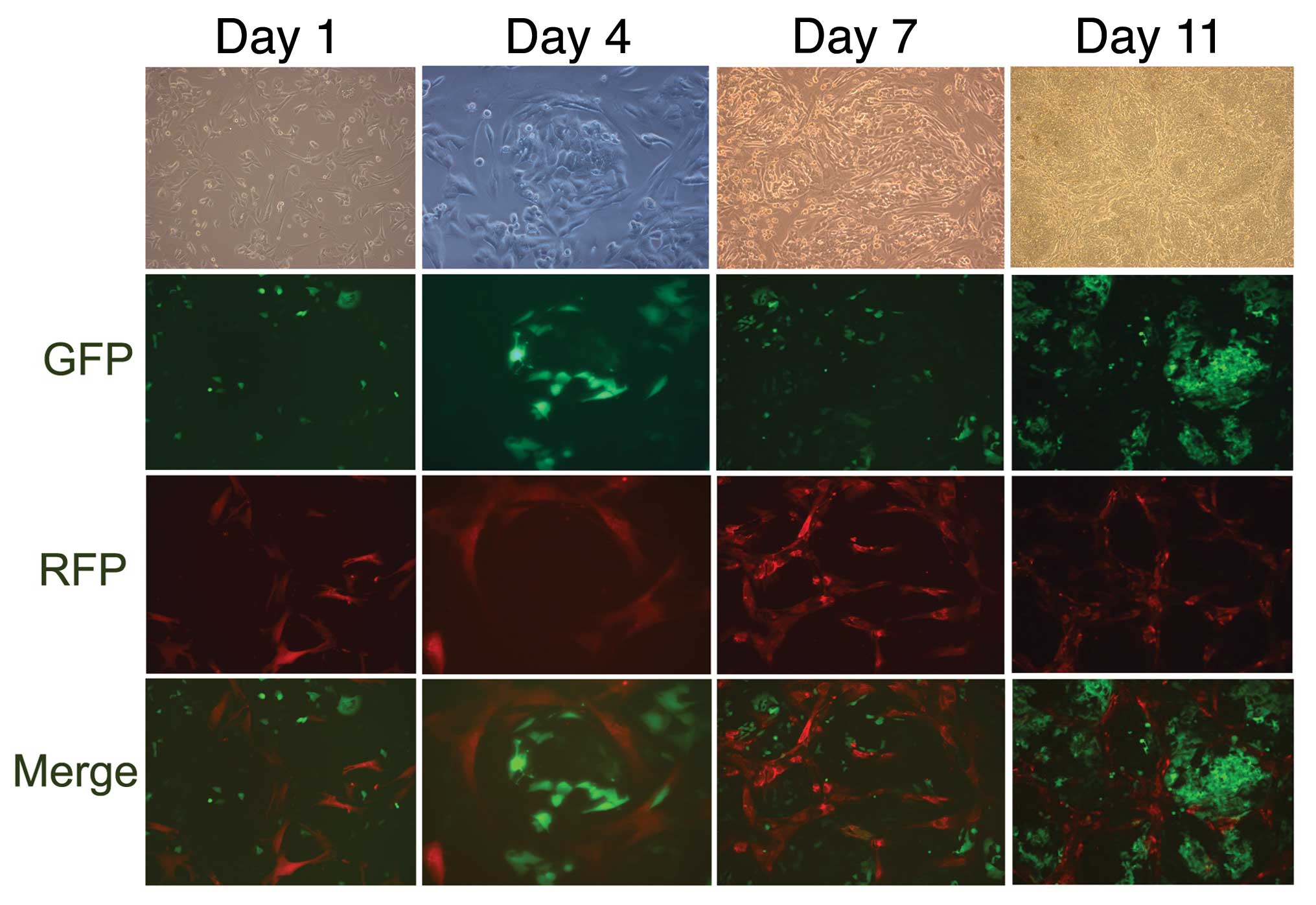
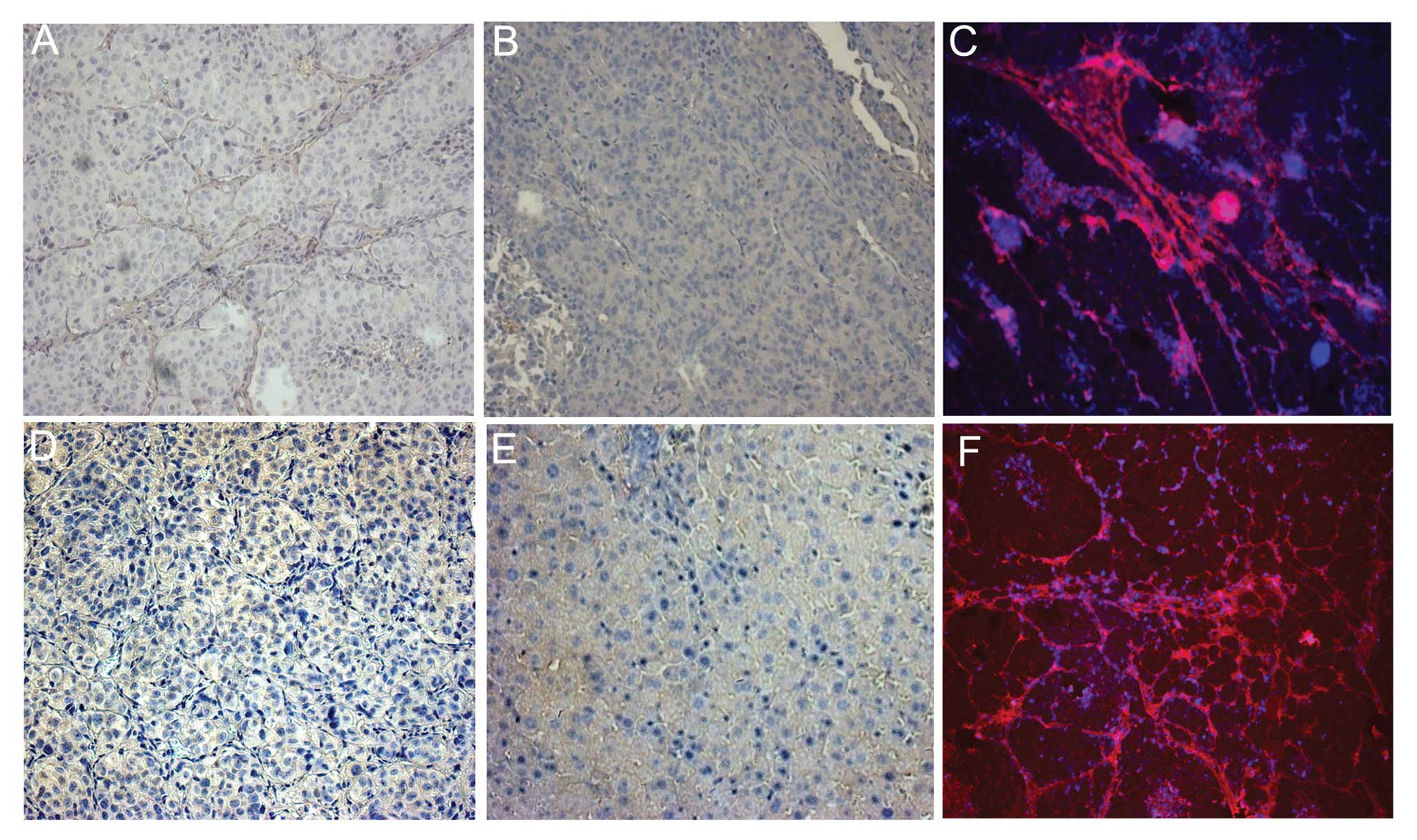
After the co-culture of GFP-MHCC97-H with RFP-MSCs,
and the co-culture of GFP-MHCC97-H with DAPI-MSCs, the culture
formation mimicked the in vivo tumor structures. MSCs
gradually circled and partitioned cancer cells (Fig. 1). Cell fusion was observed in the
cultures on the 4th day, and binucleated or yellow fluorescent
hybrid cells had formed. However, the cells fused at a very low
frequency (approximately 4–5 GFP+ RFP+ cells
and 4–5 binucleated cells were observed per microscopic sight;
×20), and the cell fusion did not obviously increase in the
following days (Fig. 2A and B).
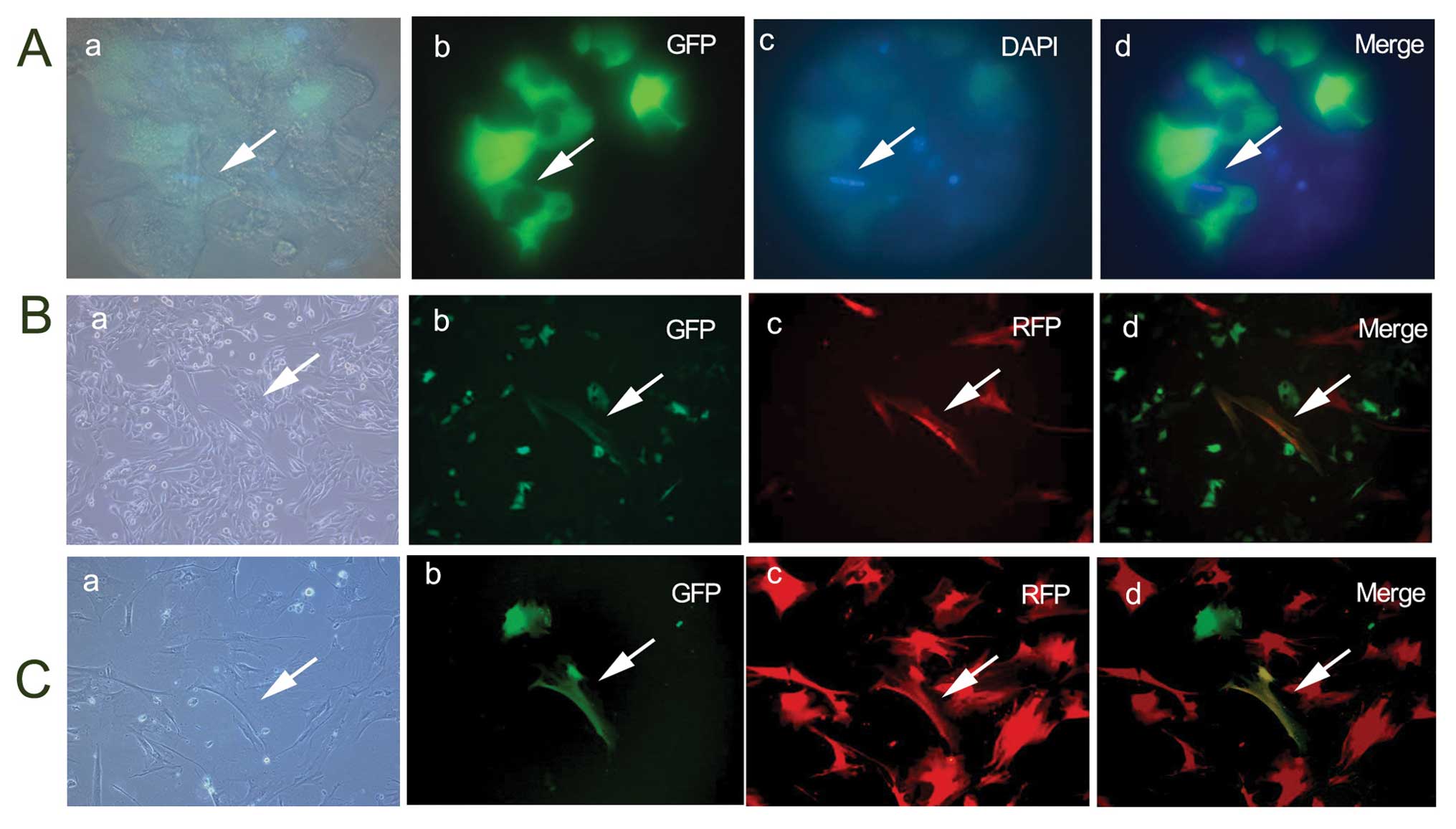
MSCs spontaneously fuse with each other
in vitro
When GFP-MSCs and RFP-MSCs were co-cultured for 4
days, cell fusion was also observed in the culture; however, the
cell fusion was also generated at a low frequency and did not
increase in the following time-periods (Fig. 2C).
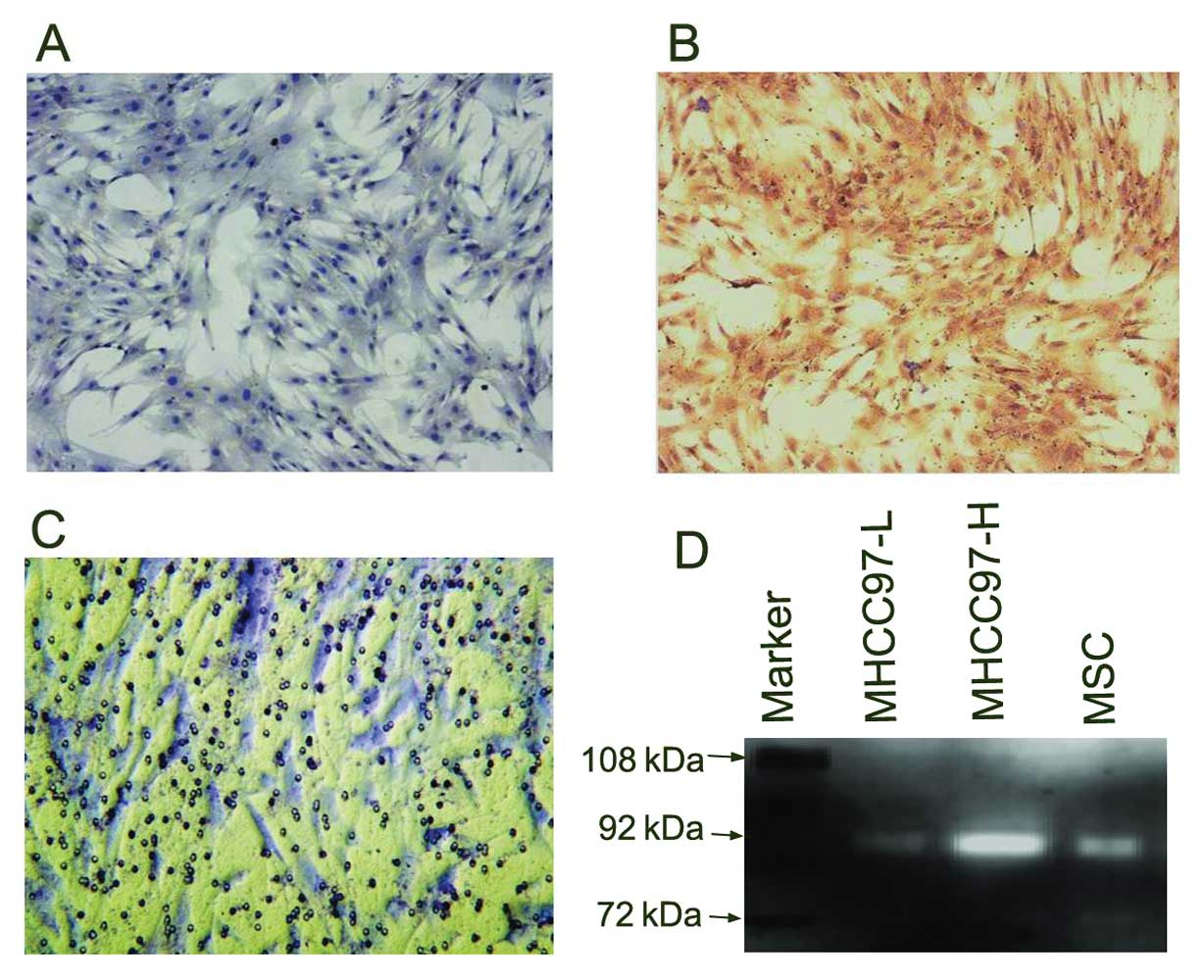
Detection of MMP expression and invasive
capability of MSCs
We found that MSCs expressed MMP2 at a low level
(Fig. 3A), but highly expressed
MMP9 (Fig. 3B), as shown by
immunocytochemistry staining with MMP2 and MMP9 monoclonal
antibodies. MSCs infiltrated through the Matrigel, as shown by
in vitro Transwell assay (Fig.
3C). The zymographic pattern showed that the MSCs had a higher
activity of MMP9 than the MHCC97-L cells (HCC cells with a lower
metastatic potential) (Fig. 3D)
(17,22).
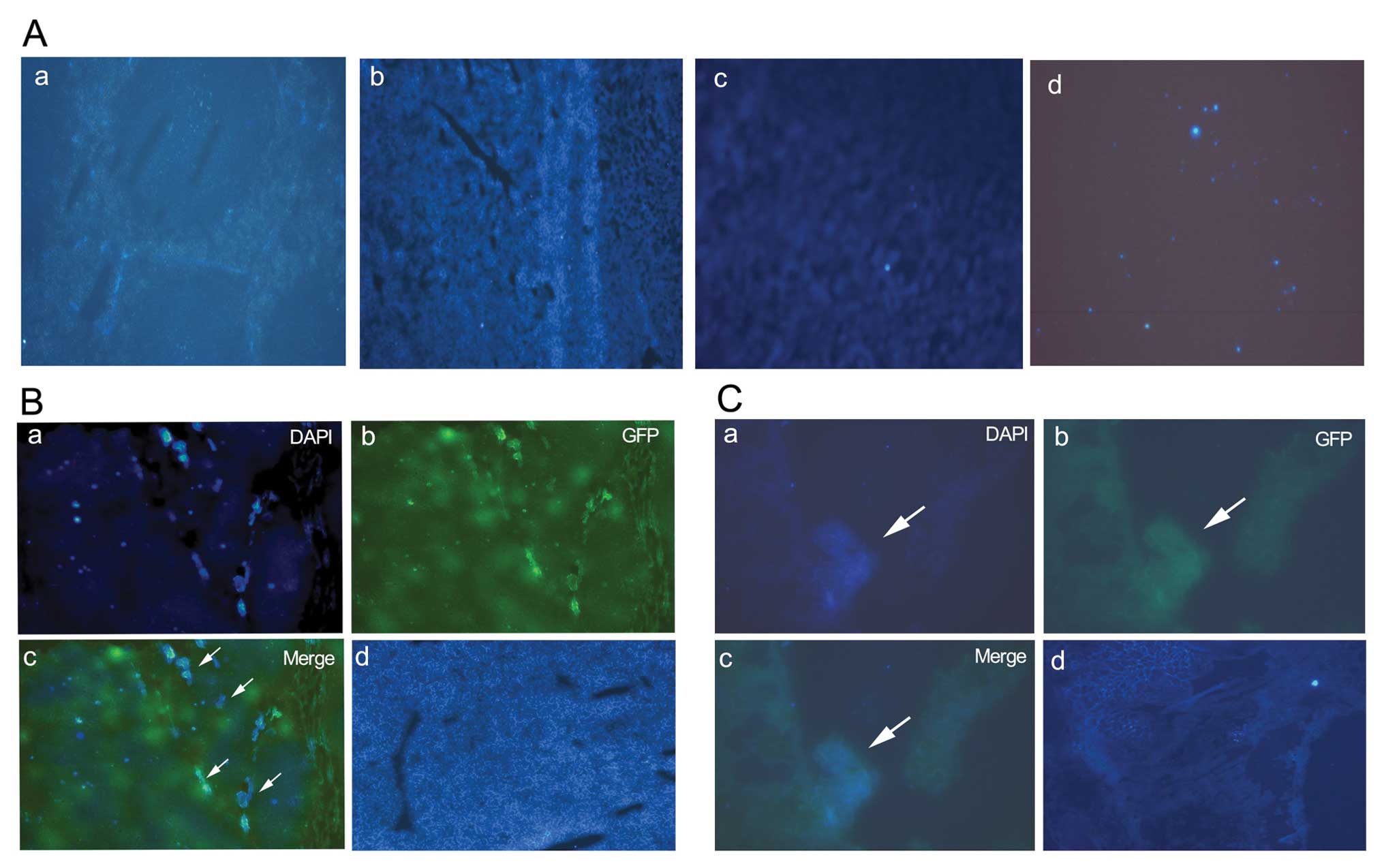
DAPI-MSCs home to tumor site
Four days after injecting DAPI-MSCs into the mice,
we found that fewer MSCs had engrafted the heart, spleen, kidneys
and bone marrow, (Fig. 4A) while,
more DAPI-MSCs were present in the tumor site compared with the
normal liver tissues (Fig. 4B).
More DAPI-MSCs were present in the site of pulmonary metastasis
compared with the normal lung tissues (Fig. 4C).
MSCs distributed in tumor stroma
Twenty days after the BrdU-MSC injection, we did not
find any more binucleated cells in the tumor tissuses and the cells
were mainly distributed in the tumor stroma. Moreover, we found
that the components of the tumor stroma had increased compared to
the tumors not injected with MSCs, as shown by immunostaining with
Brdu (Fig. 5A–D).
Injection of MSCs affects the invasion of
HCC cells
After the in vivo injection of MSCs, we also
found that the invasive capability of the HCC cells significantly
decreased. The rate of pulmonary metastasis was 20% in the group
injected with MSCs and 100% in the group not injected with MSCs
(P=0.01). The total number of metastatic cells, as well as the
total number of metastatic lesions in the lungs had decreased
(Table I).
 | Table IEffect of MSCs on tumor growth and
invasion of HCC. |
Table I
Effect of MSCs on tumor growth and
invasion of HCC.
| MSC injection | No. of animals | (Mean ± SE) | P-value |
|---|
| Tumor weight | Yes | 5 | 2.63±0.29 | 0.163 |
| No | 5 | 3.10±0.21 | |
| Rate (%) | Yes | 5 | 20% | 0.010 |
| No | 5 | 100% | |
| No. of lesions | Yes | 5 | 0.60±0.60 | 0.008 |
| No | 5 | 3.00±0.71 | |
| No. of cells | Yes | 5 | 7.60±7.60 | 0.029 |
| No | 5 | 58.00±17.36 | |
Discussion
Although cell fusion has been emphasized in other
studies, few hybrid cells were observed in our experiment both
in vitro and in vivo, and cell fusion mainly occurred
4–5 days after in vitro co-culture; even between the MSCs
themselves, cell fusion spontaneously occurred at a low level
(Fig. 2C). Our results suggested
that cell fusion was not a major mechanism by which MSCs influence
HCC. These results were consistent with the report that engrafted
MSCs do not fuse with somatic cells in rats, as shown by
fluorescence in situ hybridization (20). In another study (21), binucleated heterokaryons were not
observed in the regenerated liver after MSC transplantation. The
efficiency of in vivo somatic fusion is possibly influenced
by a number of factors. BMDCs have been found to regenerate liver
tissue by fusing with existing hepatocytes at a low frequency
(12,22,23).
Moreover, it is haematopoietic stem cells (HSCs), rather than MSCs,
that are more often associated with the phenomena of cell fusion
(11,22–25).
MMPs are important enzymes that mediate endothelial
cell invasion and the homing of stem cells (25,26).
It has been reported that MMP9 is one of the elements required to
break down basement membranes and to provide a road map for the
homing of HSCs (25). In this
study, MSCs expressed a high level of MMP9, suggesting that the
homing of MSCs also correlates with MMP9. We also observed a higher
invasive capability of MSCs in vitro by Transwell assay and
a higher number of MSCs distributed in the tumor site rather than
the normal liver tissues. These data suggest that tumor
microenvironment may provide a special niche for the homing of
engrafted MSCs.
Our results demonstrated that MSCs were mainly
distributed in the tumor stroma and differentiated into stromal
cells; which is in disagreement with the hepatocellular
differentiation of MSCs, and which reflects a limited
differentiation of MSCs in the microenvironment of HCC. The views
regarding the differentiation capability of MSCs are controversial.
The self-renewal and proliferative capacity of adult stem cells is
very limited (27) and only a
limited proportion of adult stem cells isolated from post-natal
tissues are capable of differentiating into the hepatic lineage
(28,29). The hepatic differentiation of MSCs
in vitro may be due to the ideal milieu created by humans
MSCs which are chemically defined, either by the use of serum-free
or synthetic serum replacements (30), with the possible supplementation of
specific recombinant cytokines, growth factors and extracellular
matrix (ECM) substratum.
The tumor stroma is composed of myofibroblasts and
fibroblasts which produce the extracellular matrix supporting the
tumor structure and influences invasiveness (31–33).
The tumor stroma has been regarded as a more dynamic component of
tumors. In our study, we found that MSCs were mainly distributed in
the tumor stroma. Therefore, the engrafted MSCs may be capable of
changing the tumor microenvironment, influencing tumor progression
in a complex manner associated with the secretion of growth
factors, chemokines and cytokines (6,34,35).
In conclusion, our results suggest that after
transplanting MSCs in HCC tumors, cell fusion is not the major
mechanism of action of the MSCs; MSCs mainly engraft into the tumor
tissues and differentiate into tumor stromal cells and thus
regulate the formation of the stroma. The data presented in this
study may provide novel potential methods for the application of
anticancer therapies.
Acknowledgements
The present study was supported in part by the China
National Key Projects for Infectious Disease (2008ZX10002-021), the
China National Natural Science Foundation for Distinguished Young
Scholars (30325041) and the China National High-tech Research and
Development Program (863 Program) (2006AA02Z473), and the Program
of Shanghai Subject Chief Scientist (08XD1400800). We would like to
thank Dr Qiong Xue, Dr Dongmei Gao and Dr Jun Chen for assistance
with the animal experiments, Dr Ruixia Sun and Dr Jie Chen for
helpful suggestions for the cell culture experiments and Dr Haiying
Zeng and Dr Tengfang Zhu for assistance with the pathological
experiments.
References
|
1
|
Studeny M, Marini FC, Dembinski JL,
Zompetta C, Cabreira-Hansen M and Bekele BN: Mesenchymal stem
cells: potential precursors for tumor stroma and targeted-delivery
vehicles for anticancer agents. J Natl Cancer Inst. 96:1593–1603.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Horwitz EM, Prockop DJ, Fitzpatrick LA,
Koo WW, Gordon PL and Neel M: Transplantability and therapeutic
effects of bone marrow-derived mesenchymal cells in children with
osteogenesis imperfecta. Nat Med. 5:309–313. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Beachy PA, Karhadkar SS and Berman DM:
Tissue repair and stem cell renewal in carcinogenesis. Nature.
432:324–331. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fierro FA, Sierralta WD, Epuñan MJ and
Minguell JJ: Marrow-derived mesenchymal stem cells: role in
epithelial tumor cell determination. Clin Exp Metastas. 21:313–319.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu W, Xu W, Jiang R, Qian H, Chen M, Hu
J, Cao W, Han C and Chen Y: Mesenchymal stem cells derived from
bone marrow favor tumor cell growth in vivo. Exp Mol Pathol.
80:267–274. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Karnoub AE, Dash AB, Vo AP, Sullivan A,
Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R and Weinberg
RA: Mesenchymal stem cells within tumour stroma promote breast
cancer metastasis. Nature. 449:557–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reyes M, Dudek A, Jahagirdan B, Koodie L,
Marker PH and Verfaillie CM: Origin of endothelial progenitors in
human postnatal bone marrow. J Clin Invest. 109:337–346. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Djouad F, Plence P, Bony C, Tropel P,
Apparailly F, Sany J, Noël D and Jorgensen C: Immunosuppressive
effect of mesenchymal stem cells favors tumor growth in allogeneic
animals. Blood. 102:3837–3844. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Portolani N, Coniglio A, Ghidoni S,
GiovanellMi A, Benetti G and Tiberio A: Early and late recurrence
after liver resection for hepatocellular carcinoma: prognostic and
therapeutic implications. Ann Surg. 243:229–235. 2006. View Article : Google Scholar
|
|
10
|
Ye QH, Qin LX, Forgues M, He P, Kim JW and
Peng AC: Predicting hepatitis B virus-positive metastatic
hepatocellular carcinomas using gene expression profiling and
supervised machine learning. Nat Med. 9:416–423. 2003. View Article : Google Scholar
|
|
11
|
Alison MR and Lovell MJ: Liver cancer: the
role of stem cells. Cell Prolif. 38:407–421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vassilopoulos G, Wang PR and Russell DW:
Transplanted bone marrow regenerates liver by cell fusion. Nature.
422:901–904. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang X, Willenbring H, Akkari Y, Torimaru
Y, Foster M, Al-Dhalimy M, Lagasse E, Finegold M, Olson S and
Grompe M: Cell fusion is the principal source of
bone-marrow-derived hepatocytes. Nature. 422:897–901.
2003.PubMed/NCBI
|
|
14
|
Li GC, Ye QH, Xue YH, Sun HJ, Zhou HJ, Ren
N, Jia HL, Shi J, Wu JC, Dai C, Dong QZ and Qin LX: Human
mesenchymal stem cells inhibit metastasis of a hepatocellular
carcinoma model using the MHCC97-H cell line. Cancer Sci.
101:2546–2553. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nygren JM, Jovinge S, Breitbach M, Säwén
P, Röll W, Hescheler J, Taneera J, Fleischmann BK and Jacobsen SE:
Bone marrow-derived hematopoietic cells generate cardiomyocytes at
a low frequency through cell fusion, but not transdifferentiation.
Nat Med. 10:494–501. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen EH, Grote E, Mohler W and Vignery A:
Cell-cell fusion. FEBS Lett. 581:2181–2193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ying QL, Nichols J, Chambers I and Smith
A: BMP induction of Id proteins suppresses differentiation and
sustains embryonic stem cell self-renewal in collaboration with
STAT3. Cell. 115:281–292. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Tang Y, Ye L, Liu B, Liu K and Chen
J: Establishment of a hepatocellular carcinoma cell line with
unique metastatic characteristics through in vivo selection and
screening for metastasis-related genes through cDNA microarray. J
Cancer Res Clin Oncol. 129:43–51. 2003.
|
|
19
|
Li Y, Tang ZY, Ye SL, Liu YK, Chen J and
Xue Q: Establishment of cell clones with different metastatic
potential from the metastatic hepatocellular carcinoma cell line
MHCC97. World J Gastroenterol. 7:630–636. 2001.PubMed/NCBI
|
|
20
|
Sato Y, Araki H, Kato J, Nakamura K,
Kawano Y, Kobune M, Sato T, Miyanishi K, Matsuura A, Hamada H and
Niitsu Y: Human mesenchymal stem cells xenografted directly to rat
liver are differentiated into human hepatocytes without fusion.
Blood. 106:756–763. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kozorovitskiy Y and Gould E: Stem cell
fusion in the brain. Nat Cell Biol. 5:952–954. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, Willenbring H and Akkari Y: Cell
fusion is the principal source of bone-marrow-derived hepatocytes.
Nature. 42:2897–2901. 2003.
|
|
23
|
Alvarez-Dolado M, Pardal R, Garcia-Verdugo
JM, Fike JR, Lee HO, Pfeffer K, Lois C, Morrison SJ and
Alvarez-Buylla A: Fusion of bone-marrow-derived cells with Purkinje
neurons, cardiomyocytes and hepatocytes. Nature. 425:968–973. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Menthena A, Deb N, Oertel M, Grozdanov PN,
Sandhu J, Shah S, Guha C, Shafritz DA and Dabeva MD: Bone marrow
progenitors are not the source of expanding oval cells in injured
liver. Stem Cells. 22:1049–1061. 2004.PubMed/NCBI
|
|
25
|
Kaplan RN, Riba RD, Zacharoulis S, Bramley
AH, Vincent L, Costa C, Daniel DM, Ruggero D and Shmelkov SV:
VEGFR1-positive haematopoietic bone marrow progenitors initiate the
pre-metastatic niche. Nature. 438:820–827. 2005. View Article : Google Scholar
|
|
26
|
Ball SG, Shuttleworth AC and Kielty CM:
Circulating fibrocytes: collagen-secreting cells of the peripheral
blood. Int J Biochem Cell Biol. 36:598–606. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iakova P, Awad SS and Timchenko NA: Aging
reduces proliferative capacities of liver by switching pathways of
C/EBPalpha growth arrest. Cell. 113:495–506. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wagemaker G, Neelis KJ and Wognum AW:
Surface markers and growth factor receptors of immature hemopoie
tic stem cell subsets. Stem Cells. 13(Suppl 1): 165–171.
1995.PubMed/NCBI
|
|
29
|
Zheng YW and Taniguchi H: Diversity of
hepatic stem cells in the fetal and adult liver. Semin Liver Dis.
23:337–348. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goldsborough MD, Tilkins ML, Price PJ,
Lobo-Alfonso J, Morrison JR and Stevens ME: Serum-free culture of
murine embryonic stem (ES) cells. Focus. 20:8–12. 1998.
|
|
31
|
Desmouliere A, Guyot C and Gabbiani G: The
stroma reaction myofibroblast: a key player in the control of tumor
cell behavior. Int J Dev Biol. 48:509–517. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
De Wever O and Mareel M: Role of tissue
stroma in cancer cell invasion. J Pathol. 200:429–447.
2003.PubMed/NCBI
|
|
33
|
Direkze NC and Alison MR: Bone marrow and
tumour stroma: an intimate relationship. Hematol Oncol. 24:189–195.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Littlepage LE, Egeblad M and Werb Z:
Coevolution of cancer and stromal cellular responses. Cancer Cell.
7:499–500. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Welm AL: TGFbeta primes breast tumor cells
for metastasis. Cell. 33:27–28. 2008. View Article : Google Scholar : PubMed/NCBI
|