Introduction
Multiple myeloma (MM) is an incurable lymphoid
cancer of terminally differentiated B-cell lineage or plasma cells
that accounts for 1% of all cancers and slightly more than 10% of
all hematological malignancies (1).
Current treatment for MM, including proteasome inhibitors,
immunomodulatory drugs, high-dose chemotherapy and stem cell
transplantation, have significantly prolonged the 10-year overall
survival (2,3). However, patients presenting with
relapse are numerous due to drug-resistant disease and minimal
residual disease (4), necessitating
the development of new therapeutic approaches.
Tumor necrosis factor (TNF)-related
apoptosis-inducing ligand (TRAIL, Apo-2L) is one of the members of
the TNF gene superfamily that selectively induces apoptosis of
cancer cells but not of normal cells in various tissue types and is
currently in clinical trials for the treatment of human cancers
(5,6). Since MM is characterized as a tumor
composed of long-surviving rather than fast-growing malignant
plasma cells, potentiating apoptosis has been envisaged as a
potential therapeutic approach (7).
Previous studies found that exogenous TRAIL induced apoptosis in MM
cells from patients, one of which had chemotherapy-refractory
disease and in many MM cell lines, including cells sensitive or
resistant to dexamethasone (Dex), doxorubicin (Dox), melphalan and
mitoxantrone (8,9). This indicates that TRAIL appears
effective at inducing apoptosis in chemo-resistant MM cells.
Moreover, it has been shown that chemotherapeutic drugs such as Dox
can sensitize MM cells to TRAIL through upregulation of death
receptor 4 (DR4) and death receptor 5 (DR5) (10). Notably, a recent study revealed that
TRAIL, used in combination with Dox, eliminated tumor-initiating
cells resulting in complete and lasting eradication of MM tumors
in vivo (11). Taken
together, these observations indicate that TRAIL-based therapy
represents a novel and effective approach for the treatment of
human MM.
Cancer targeting gene-virotherapy is a novel
therapeutic platform for cancer (12–14).
This strategy involves the use of oncolytic adenoviruses (OAs)
engineered to express a therapeutic gene inserted into the E1 or E3
region of the viruses, in which the input amount of transgene is
amplified by selective replication of the viruses, resulting in
longer transgene expression within tumor cells and potent activity
against tumors. For example, it was reported that TRAIL expression
following gene transfer of OAs in human cancer cell lines was
4-fold higher than that obtained with a replication-defective
adenovirus (14). We previously
constructed the oncolytic adenoviral vector ZD55, an E1B-55K
deleted OA, carrying TRAIL (ZD55-TRAIL) (15) and demonstrated that the combination
of TRAIL and viral oncolysis yielded an additive cytotoxic effect
on cancer cells in vitro. This virus was more effective than
the unarmed control at killing tumors in vivo and prolonging
survival in murine models (15,16).
These findings prompted us to study the effect of ZD55-TRAIL on MM
cell lines. In the present study, we found that ZD55-TRAIL was
effective against MM cell lines including a drug-resistant cell
line and inhibited levels of insulin-like growth factor-1 receptor
(IGF-1R). However, the activation of AKT and mTOR was observed upon
virus infection. Based on this finding, we investigated the
synergistic effects of ZD55-TRAIL combined with the PI3K inhibitor
LY294002. This combination therapy significantly improved the
sensitivity of MM cells to ZD55-TRAIL. Furthermore, treatment of MM
cells with ZD55-TRAIL in combination with MG132, a proteasome
inhibitor, resulted in enhanced apoptosis, which was related, at
least in part, to upregulation of DR5. Together, the findings of
the present study suggest that combination therapy with ZD55-TRAIL
and a PI3K inhibitor or MG132 may be particularly appealing in the
context of drug-resistant MM.
Materials and methods
Cell culture and reagents
Cell culture medium RPMI-1640 and fetal bovine serum
(FBS) were purchased from HyClone Laboratories (Logan, UT, USA).
Human MM cell line RPMI-8226 was purchased from the American Type
Culture Collection (Rockville, MD, USA). Dex-sensitive (MM.1S) and
Dex-resistant (MM1.R) cell lines were kindly provided by Dr Steven
Rosen (Northwestern University, Chicago, IL, USA). L-02, a normal
human liver cell line, was obtained from the Shanghai Cell
Collection (Shanghai, China). The reagents used in the present
study were purchased from the following vendors: interleukin-6
(IL-6) and IGF-1 (both from Peprotech, Inc., Rocky Hill, NJ, USA);
LY294002 and MG132 (Sigma, St. Louis, MO, USA). Oncolytic viruses
including ZD55-TRAIL, ZD55-EGFP and ZD55 were generous gifts from
Professor Xinyuan Liu (Xinyuan Institute of Medicine and
Biotechnology, School of Life Sciences, Zhejiang University of
Technology and Sciences, Hangzhou, China).
Cell viability (MTT) assay
Cell viability was determined by the MTT-based
cytotoxicity assay (Sigma), as previously described (16). Briefly, cells were treated with
various concentrations of the oncolytic virus ZD55 or ZD55-TRAIL
for 48 h. On termination, the medium was replaced with fresh medium
containing 0.5 mg/ml MTT. The cells were incubated for an
additional 4 h at 37°C. Following removal of the medium and MTT,
DMSO (200 μl) was added to each well, and the absorbance at 570 nm
was detected.
Colony forming cell assay
MM colony formation assay was performed as described
elsewhere (17). Briefly, RPMI-8226
cells (2×103 cells) were cultured with IMDM medium
containing FBS (200 μl) and 2.7% methylcellulose (300 μl; Sigma) at
37°C in a humidified atmosphere with 5% CO2. After 7
days, colonies that contained >50 cells were scored manually
under a light microscope (Olympus, Tokyo, Japan).
Hoechst DNA staining
Nuclei were stained with Hoechst 33258 (Sigma) to
detect chromatin condensation and nuclear fragmentation, which are
phenotypic characteristics of apoptosis. Treated RPMI-8226 cells
were fixed with 4% paraformaldehyde and then stained with Hoechst
33258 (1 μg/ml) for 15 min. Stained slides were analyzed under a
fluorescence microscope (Olympus).
Western blot analysis
Western blotting was carried out as described
previously (17). Briefly, equal
amounts of total cell lysates were separated on sodium
dodecyl-sulfate (SDS)-polyacrylamide gels containing 8–12%
acrylamide gradients and then transferred to polyvinylidene
difluoride membranes (Millipore, Bedford, MA, USA). Non-specific
binding was blocked with 10 mM Tris-HCL buffered saline plus 0.05%
Tween-20 containing 5% skimmed milk powder for 1 h at room
temperature. Membranes were incubated with the primary antibody
overnight at 4°C. The primary antibodies used here were as follows:
poly[adenosine diphosphate (ADP)-ribose] polymerase (PARP),
caspase-8, -9 and -3, Bcl-xL, cytochrome c (cyto c),
p110α, IGF-1R, AKT, p-AKT (Ser473), mTOR, p-mTOR (Ser2448), p-mTOR
(Ser2481), p70S6K, p-p70S6K (Thr389), 4E-BP1, p-4E-BP1 (Thr70) and
DR5 purchased from Cell Signaling Technology, Inc. (Beverly, CA,
USA); DR4 was provided by Bioworld (Louis Park, MN, USA); β-actin
was obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). TRAIL and p110δ were purchased from BioLegend (San Diego, CA,
USA) and Epitomics (Burlingame, CA, USA), respectively.
Preparation of cDNA and real-time
PCR
Preparation of total RNA was conducted using an
RNeasy Plus kit (Takara Shuzo, Kyoto, Japan). Each cDNA template
was made from total RNA with the Superscript II reverse
transcriptase kit according to the manufacturer’s instructions
(Invitrogen Life Technologies, Carlsbad, CA, USA). For IGF-1R we
used 5′-TTAAAATGGC CAGAACCTGAG-3′ as the forward primer, and
5′-ATTATA ACCAAGCCTCCCAC-3′ as the reverse primer. For GAPDH we
used 5′-GTAACCCGTTGAACCCCATT-3′ as the forward primer, and
5′-CCATCCAATCGGTAGTAGCG-3′ as the reverse primer. Amplification
reactions were performed using SYBR® Premix Ex Taq™
(Takara Shuzo) in the iQ5 Multicolor Real-time PCR detection system
(Bio-Rad, Hercules, CA, USA). Thermal cycling conditions were as
follows: 30 sec at 95°C for initial denaturing, 5 sec at 95°C for
denaturing and 30 sec at 60°C for annealing and extension for a
total of 40 cycles. The fold-change in mRNA was calculated by the
2−ΔΔCt method.
Statistical analysis
Statistical analysis of the experimental results was
conducted by the Student’s t-test. A P-value <0.05 was
considered to indicate a statistically significant result.
Synergisms in the combination treatments were analyzed using
CalcuSyn software (Biosoft, Cambridge, UK).
Results
ZD55 effectively infects MM cells and
inhibits the growth of myeloma cell lines
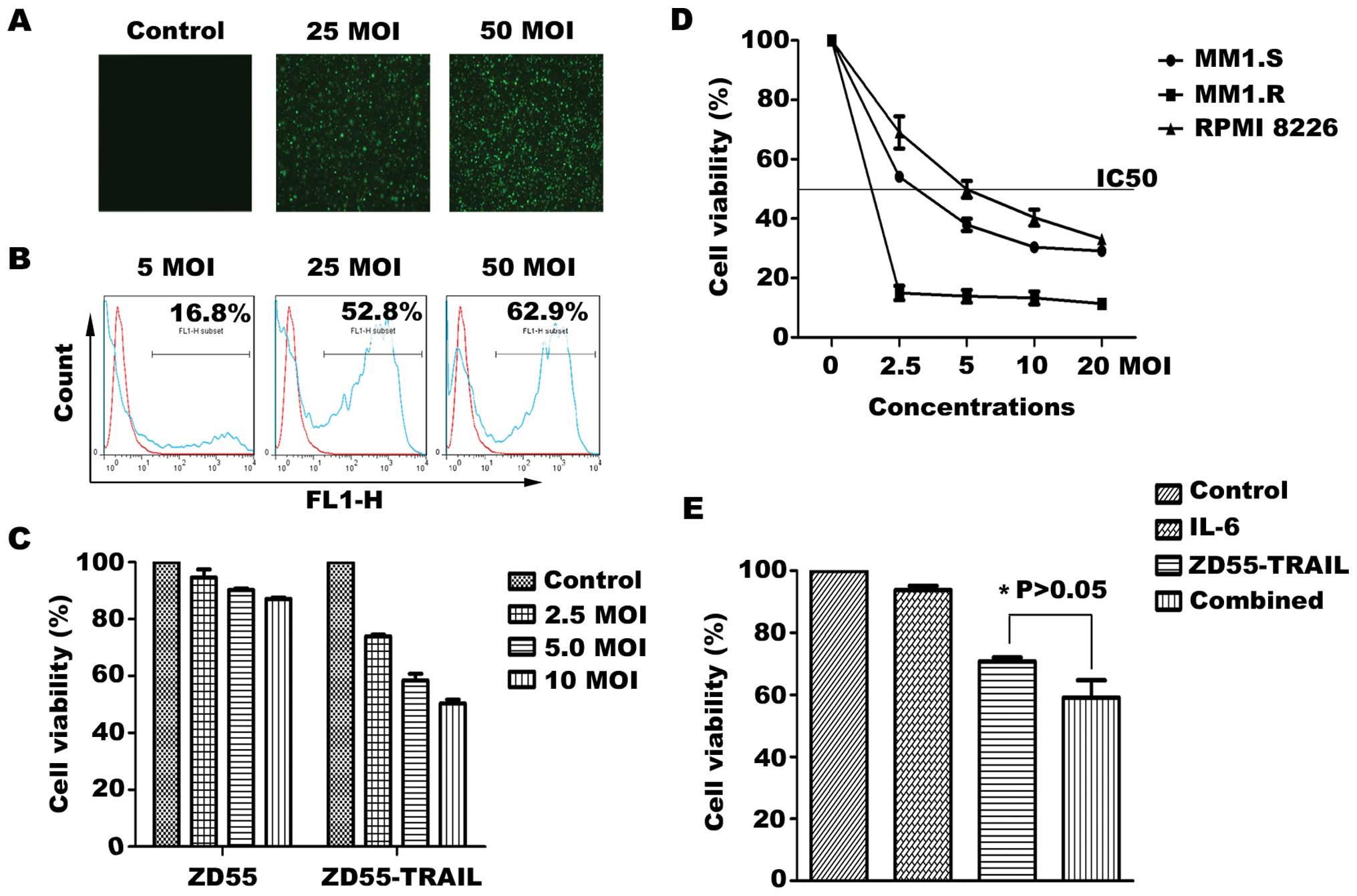
To determine the infectivity of ZD55 in MM cells,
RPMI-8226 cells were treated with the ZD55-EGFP vector at the
indicated MOIs for 24 h, and then GFP-positive cells were observed
under a fluorescence microscopy. As shown in Fig. 1A, RPMI-8226 cells in culture were
effectively infected with ZD55-EGFP in a dose-dependent manner. As
determined by flow cytometric analysis, 52% GFP-positive cells were
noted in the group treated with ZD55-EGFP at an MOI of 25 for 48 h,
while 62% GFP-positive cells were observed in the group treated
with this virus at an MOI of 50 (Fig.
1B). To test whether ZD55-TRAIL has enhanced cytotoxicity, a
cell viability assay was performed using RPMI-8226 cells infected
with ZD55-TRAIL or ZD55, respectively, at the indicated MOIs for 48
h. The viability of RPMI-8226 cells was significantly reduced by
ZD55-TRAIL in a dose-dependent manner when compared with the
viability of the cells with no viral treatment, whereas ZD55 had a
slight cytotoxic effect (Fig. 1C).
Importantly, dose-dependent growth inhibition was also observed in
the MM1.S and the drug-resistant cell line MM1.R (Fig. 1D). Since IL-6 mediates growth and
survival of MM cells, we next examined the effect of ZD55-TRAIL on
RPMI-8226 cells in the presence of exogenous IL-6. We found that
IL-6 (10 ng/ml) did not provide protection against
ZD55-TRAIL-induced cell death (Fig.
1E), suggesting that ZD55-TRAIL, unlike Dex, is able to inhibit
cytokine-induced MM cell growth.
Inhibition of myeloma cell proliferation
by ZD55-TRAIL is due to apoptosis induction
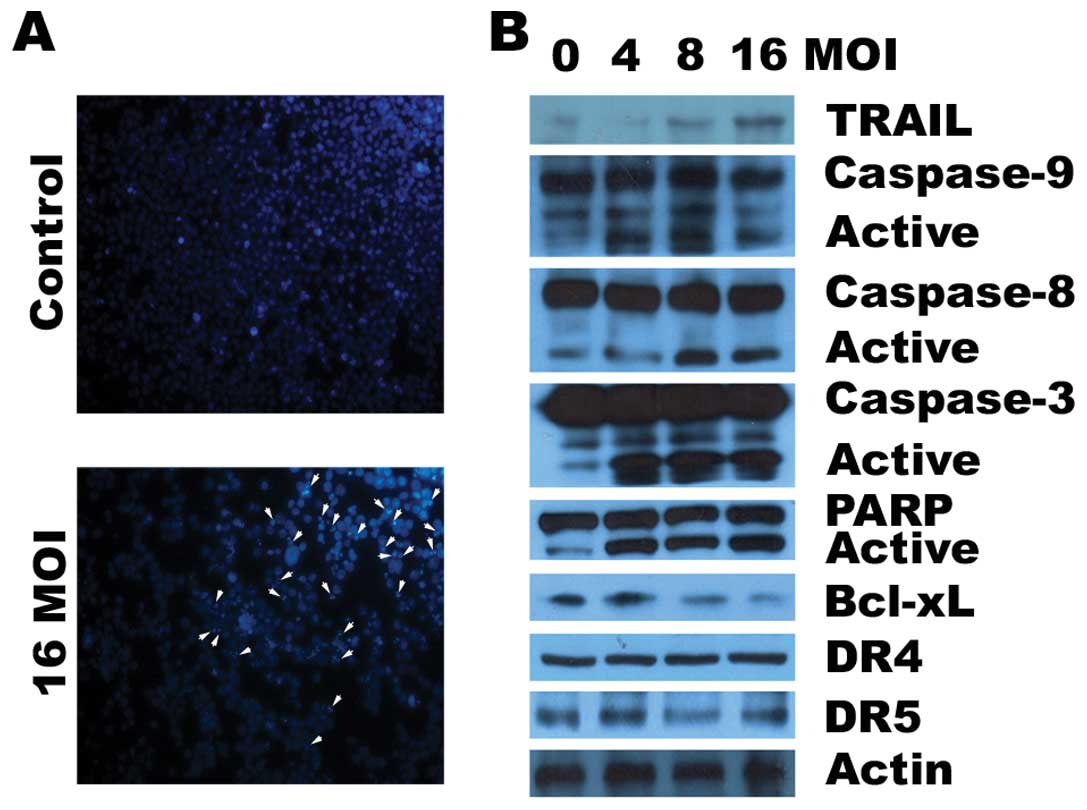
We next determined the effects of ZD55-TRAIL on
induction of apoptosis by the quantification of apoptotic nuclei
following Hoechst 33258 staining. As shown in Fig. 2A, ZD55-TRAIL significantly increased
DNA fragmentation in RPMI-8226 cells, indicating that MM cells
underwent apoptosis following ZD55-TRAIL infection. We further used
immunoblotting to assess the expression of TRAIL and activation of
caspases in the RPMI-8226 cells treated with ZD55-TRAIL. ZD55-TRAIL
at the tested concentration range (4–16 MOIs) effectively induced
cleavage of caspase-8, -9 and -3, followed by PARP cleavage, while
the levels of TRAIL expression increased in a dose-dependent manner
(Fig. 2B). However, during
ZD55-TRAIL treatment, the expression levels of DR4 and DR5 were not
affected. Previous studies suggest that Bcl-2 or Bcl-xL confers
resistance to chemotherapy in MM cells (18). We found that ZD55-TRAIL decreased
the expression of Bcl-xL in a dose-dependent fashion.
ZD55-TRAIL inhibits IGF-1R and may
overcome drug resistance
It was recently reported that signaling through the
IGF-1/IGF-1R axis contributes to acquired bortezomib resistance
(19), and that inhibition of
IGF-1R enhances cell death of MM cells induced by an IKK2 inhibitor
(20). In the present study, we
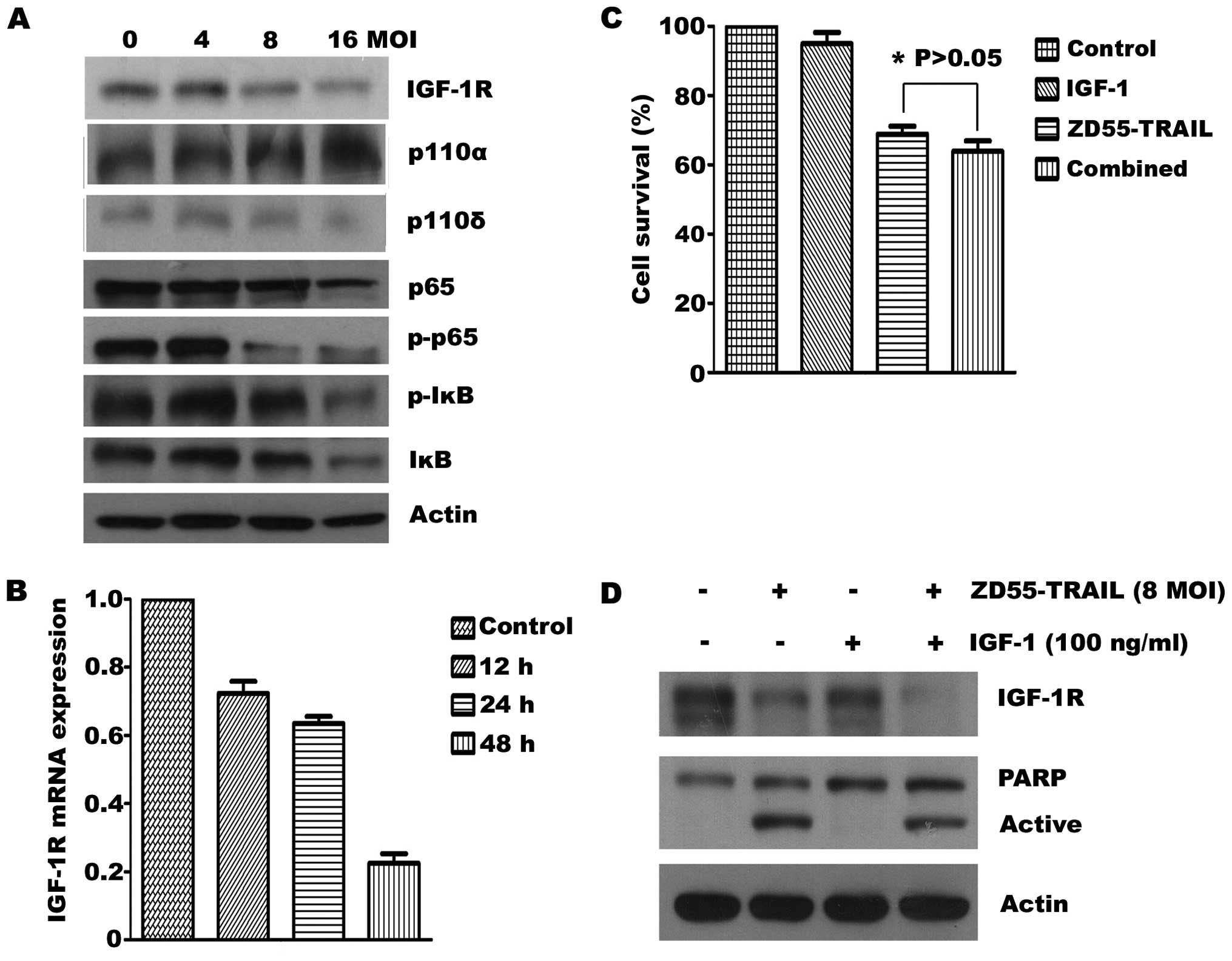
aimed to ascertain whether ZD55-TRAIL suppresses IGF-1R expression
in RPMI-8226 cells. As shown in Fig.
3A, the virus decreased the level of IGF-1R as well as NFκB
pathway-related proteins, which included p-p65 and p-IκB, although
the precise mechanism by which it does so is unknown. Moreover,
quantitative PCR analysis showed that the mRNA expression of IGF-1R
was also significantly inhibited by ZD55-TRAIL in a time-dependent
manner (Fig. 3B). We next examined
the effects of IGF-1 on ZD55-TRAIL-induced cell death. Exposure of
MM cells to IGF-1 (100 ng/ml) did not reverse the cellular growth
inhibition and apoptosis induced by ZD55-TRAIL (Fig. 3C and D) indicating that IGF-1R
inhibition, by ZD55-TRAIL, may contribute to the enhanced cytotoxic
action of the virus.
LY294002 has a synergistic effect with
ZD55-TRAIL to inhibit cell proliferation in myeloma cells
Given the fact that the PI3K/AKT/mTOR pathway is
constitutively activated in human MM cell lines and in primary
plasmocytes from patients with MM, we next examined the effects of
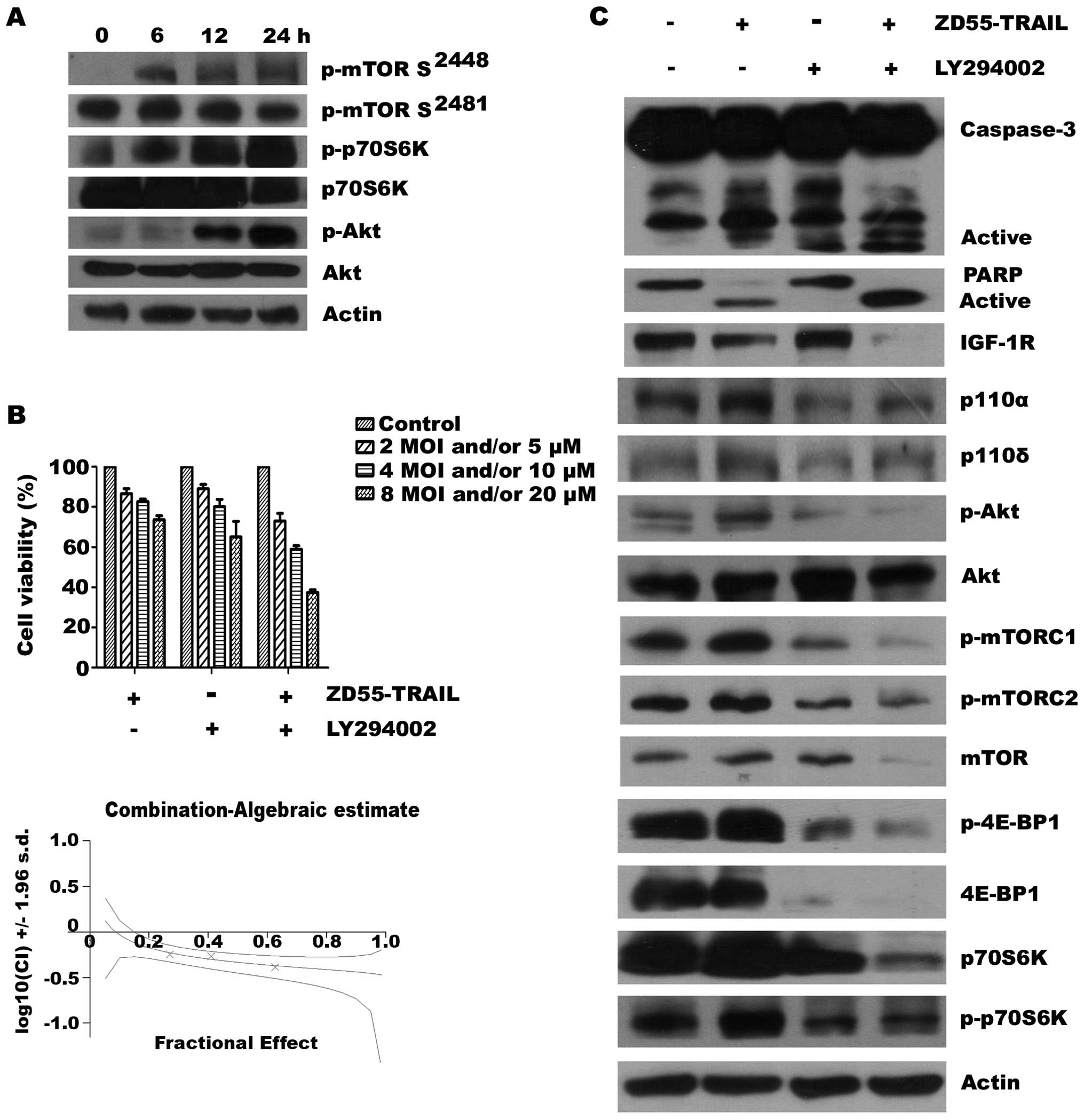
ZD55-TRAIL on several key signaling molecules in RPMI-8226 cells.
We found that ZD55-TRAIL activated AKT and mTOR as evidenced by the
upregulation of phosphor-mTOR (Ser2448) and phosphorylated AKT at
Ser473 (Fig. 4A), which may
stimulate MM cell growth and promote cell survival and migration
(21,22). To improve the anti-myeloma efficacy
of ZD55-TRAIL, RPMI-8226 cells were treated with a series of doses
of ZD55-TRAIL or/and LY294002, a PI3K inhibitor. As revealed in
Fig. 4B, treatment of MM cells with
ZD55-TRAIL plus LY294002 resulted in significantly lower cell
viability than that following either ZD55-TRAIL or LY294002
treatment. By combination index analysis, we observed synergistic
cytotoxic effects when the two agents were combined, as evidenced
by a CI <0 (Fig. 4B).
Furthermore, western blot analysis showed that addition of the PI3K
inhibitor LY294002 led to increased levels of cleaved forms of
caspase-3 and PARP (Fig. 4C),
suggesting enhanced apoptosis upon co-treatment. To understand the
mechanisms involved in the apoptotic synergy between ZD55-TRAIL and
LY294002, we examined the effects of the combination therapy on
mTOR, a member of the PI3K-related kinase protein family. Western
blot analysis indicated that LY294002 suppressed mTOR expression
and phosphorylation of mTOR protein on Ser2448, a marker for the
presence of mammalian target of rapamycin complex 1 (mTORC1)
(23), as well as phosphorylation
of mTOR on Ser2481, a marker for mTORC2 activity (23,24).
As a result, phosphorylation of 4E-BP1 and p70S6K, downstream
targets of mTORC1, as well as phospho-AKT (Ser473), a downstream
target of mTORC2 were also inhibited by LY294002. Notably, enhanced
inhibition of key components of the mTOR pathway was observed in
the cells treated with ZD55-TRAIL combined with LY294002 (Fig. 4C).
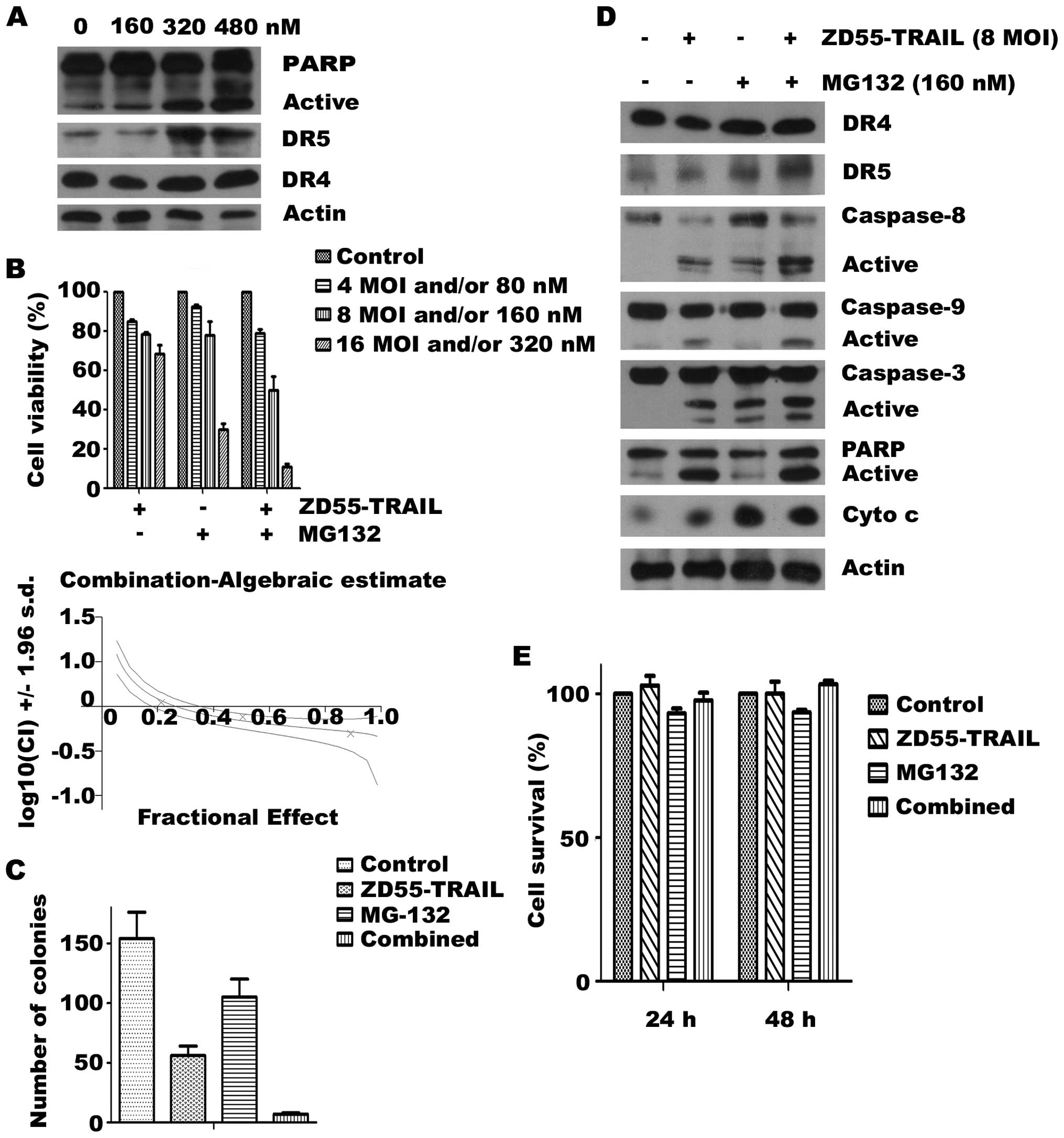
MG132 upregulates DR5 and potentiates
ZD55-TRAIL-mediated cell death in RPMI-8226 cells
Since proteasome inhibitor MG132 is shown to
sensitize tumor cells to TRAIL via upregulation of DR4 and DR5
(25,26), we therefore examined the synergistic
effects of ZD55-TRAIL in combination with MG132. For this purpose,
we treated RPMI-8226 cells with different concentrations of MG132
and ZD55-TRAIL either alone or in combination for 24 h and assessed
cell viability using an MTT assay. We found that a low-dose of
MG132 (160–320 nM) markedly enhanced the cell death of low-dose
ZD55-TRAIL (8–16 MOIs)-infected RPMI-8226 cells (Fig. 5A). The combination index values
indicated a strong synergy between MG132 and ZD55-TRAIL (Fig. 5B). Furthermore, ZD55-TRAIL and MG132
reduced the colony formation ability of MM cells in vitro,
which was significantly increased when the two treatments were
combined (Fig. 5C). Analysis of
RPMI-8226 nuclei after Hoechst staining revealed that the
synergistic cytotoxic effect of the combination therapy was due to
enhanced apoptosis (data not shown), which was confirmed by western
blotting that showed an enhanced caspase cascade activation in the
cells treated with the two agents as compared with the ZD55-TRAIL-
or MG132-treated cells (Fig. 5D).
To assess the effects of ZD55-TRAIL combined with MG132 on normal
cells, we treated L02, a normal human liver cell line, with
ZD55-TRAIL, MG132, or ZD55-TRAIL plus MG132 and then determined the
cell viability using an MTT assay. As shown in Fig. 5E, ZD55-TRAIL, MG132, or the combined
treatment at the indicated dosage did not induce cytotoxicity
against normal cells.
Discussion
The oncolytic adenovirus is a potent member of a
potentially new class of antitumor agents that has been previously
demonstrated to have activity in MM (27–29).
Among the viruses used for virotherapy of MM, different serotypes
of adenoviruses have been studied, with the goal of finding a
translational application for the treatment of myeloma progressing
on prior therapy (30,31). In this regard, Senac et al
showed that adenovirus serotype 5 (Ad5) infected most MM cell lines
and primary myeloma cells from patients and that oncolytic Ad5
appeared to infect CD138+ MM cells 10- to 70-fold more
efficiently than normal marrow cells (30), suggesting the possibility of
translating adenovirus-based virotherapies to MM patients. In the
present study, we show that ZD55, an Ad5 oncolytic virus,
efficiently infected the MM cell line RPMI-8226, as demonstrated by
fluorescence microscopy and flow cytometric analysis. Using
ZD55-TRAIL as a gene virotherapy, we found that the virus was able
to effectively kill a panel of myeloma cell lines, which included a
drug-resistant cell line MM1.R. Consistent with previous reports on
hepatocellular carcinoma cell lines (15,16),
treatment of RPMI-8226 cells with ZD55-TRAIL resulted in induction
of apoptosis. This effect was also confirmed by our observed
cleavage of caspase-3 and PARP. In addition, we found that IL-6
failed to protect against ZD55-TRAIL-induced growth inhibition.
Therefore, ZD55-TRAIL, which kills MM cells via both oncolysis and
apoptosis, may be particularly useful to overcome drug
resistance.
The proliferation and survival of MM cells have been
shown to be related to the activation of several signaling pathways
such as PI3K/AKT, JAK/STAT3, MAPK/ERK and NFκB (32–34),
which can be stimulated by several cytokines including IL-6 and
IGF-1 (35). In MM, IGFs and IGF-1R
signaling was correlated with stimulation of proliferation,
survival and drug-resistance (36,37).
Recent studies suggest that the inhibition of IGF-1R decreases cell
proliferation. Furthermore, combinations of multiple drugs such as
the IKK2 inhibitor AS602868 and bortezomib with agents intervening
in the IGF-1R pathway were found to result in enhanced apoptosis
and reversal of drug-resistance (19,20,38),
suggesting that targeting the IGF-1/IGF-1R axis is a promising
approach for the treatment of MM. Our results confirm the
inhibitory effect of ZD55-TRAIL on the expression of IGF-1R in
RPMI-8226 cells. Moreover, we found that the virus significantly
inhibited the NFκB pathway in MM cells, as assessed by
ZD55-TRAIL-induced downregulation of phosphorylation and expression
of the p65 subunit of NFκB and inhibitory κB (IκB). It is
noteworthy that RPMI-8226 cells were more sensitive to
ZD55-TRAIL-induced cell death than MM1.S and MM1.R cells. This
result is consistent with a previous finding that RPMI-8226 cells
express more IGF-1R than MM.1S and U266 cells, and that the
cytotoxic effect of anti-IGF-1R is more effective on MM cells with
a high level of IGF-1R (20). Our
data also showed that ZD55-TRAIL-mediated apoptosis and cell growth
inhibition were not reversed by IGF. Thus, ZD55-TRAIL can be
considered as an efficient anticancer agent acting through diverse
mechanisms of action in the killing of MM cells by downregulation
of IGF-1R.
Prior studies have shown that adenoviral infection
triggers the activation of PI3K and its downstream molecules such
as AKT, ERK1/2, JNK and MAPK by viral fiber binding with the
Coxsackie-adenovirus receptor (39–41),
which may benefit tumor cell survival. In the present study,
mechanistic analyses in RPMI-8226 cells showed a high level of
phosphorylation of mTOR on Ser2448, which represents the activity
of mTORC1, and downstream activation of AKT and p70S6K in
ZD55-TRAIL-treated cells, which increased in a dose-dependent
manner. These data suggest that the virus elicited the activation
of the AKT/mTOR pathway. Based on these results, we investigated
the effect of ZD55-TRAIL combined with the AKT inhibitor LY294002
on MM cells. Our results revealed that the combination of LY294002
together with ZD55-TRAIL synergistically inhibited cell
proliferation and induced apoptosis via the inhibition of the
AKT/mTOR pathway. To our knowledge, this is the first report
demonstrating that an oncolytic adenovirus synergizes with an AKT
inhibitor to induce death of MM cells.
TRAIL, which induces apoptosis via interactions with
its death receptors DR4 and DR5, represents a promising candidate
agent for the treatment of MM (11,42,43).
Recent studies suggest that there is a correlation between TRAIL
sensitivity and death receptor expression in cancer cells (44), thus the upregulation of death
receptor expression renders MM cells more sensitive to the action
of TRAIL. In the present study, we showed that increased expression
of DR5, but not DR4, induced by MG132 was associated with
ZD55-TRAIL sensitivity in RPMI-8226 cells. This is consistent with
a previous report showing that overexpression of DR5 in
TRAIL-resistant cancer cells restored TRAIL sensitivity (45). In addition, we observed that MG132
and ZD55-TRAIL combination therapy significantly inhibited the
clonogenic survival of MM cells, which indicates the importance of
the combination therapy in the elimination of cancer progenitor
cells.
In summary, our results showed that ZD55-TRAIL
induced significant cytotoxicity in drug-sensitive and
drug-resistant myeloma cell lines in vitro. In addition to
an oncolytic effect, ZD55-TRAIL induced apoptosis through
activation of the caspase pathway. Furthermore, we showed for the
first time that ZD55-TRAIL markedly inhibited the expression of
IGF-1R and downstream NFκB. The combination of ZD55-TRAIL and
LY294002 in RPMI-8226 cells overcame the activation of mTOR and AKT
induced by the virus and consequently promoted cell death.
Moreover, increased expression of DR5 enhanced the apoptotic
response to the combined treatment of MG132 and ZD55-TRAIL. These
findings provide the rationale for further pre-clinical evaluation
of TRAIL-based adenoviral therapy in combination with a PI3K
inhibitor or a proteosome inhibitor for MM.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China grants (nos. 81370646, 81370645
and 81302036), the Doctoral Fund of the Ministry of Education of
China (no. 20120101110010), the Funds of the Science Technology
Department of Zhejiang Province (no. 2012C13021-2), the Fund of the
Health Bureau of Zhejiang Province (no. 2010SSA006), the Natural
Science Foundation of Zhejiang Province (no. Y2090455), and the
Foundation of the Zhejiang Municipal Commission of Population and
Family Planning (no. 2009-74).
References
|
1
|
Kyle RA: Multiple myeloma: an odyssey of
discovery. Br J Haematol. 111:1035–1044. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brenner H, Gondos A and Pulte D: Expected
long-term survival of patients diagnosed with multiple myeloma in
2006–2010. Haematologica. 94:270–275. 2009.
|
|
3
|
Barlogie B, Attal M, Crowley J, et al:
Long-term follow-up of autotransplantation trials for multiple
myeloma: update of protocols conducted by the Intergroupe
Francophone du Myelome, Southwest Oncology Group, and University of
Arkansas for Medical Sciences. J Clin Oncol. 28:1209–1214. 2010.
View Article : Google Scholar
|
|
4
|
Hideshima T and Anderson KC: Molecular
mechanisms of novel therapeutic approaches for multiple myeloma.
Nat Rev Cancer. 2:927–937. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hellwig CT and Rehm M: TRAIL signaling and
synergy mechanisms used in TRAIL-based combination therapies. Mol
Cancer Ther. 11:3–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Herbst RS, Eckhardt SG, Kurzrock R, et al:
Phase I dose-escalation study of recombinant human Apo2L/TRAIL, a
dual proapoptotic receptor agonist, in patients with advanced
cancer. J Clin Oncol. 28:2839–2846. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gajate C and Mollinedo F: Edelfosine and
perifosine induce selective apoptosis in multiple myeloma by
recruitment of death receptors and downstream signaling molecules
into lipid rafts. Blood. 109:711–719. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mitsiades CS, Treon SP, Mitsiades N, et
al: TRAIL/Apo2L ligand selectively induces apoptosis and overcomes
drug resistance in multiple myeloma: therapeutic applications.
Blood. 98:795–804. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gazitt Y: TRAIL is a potent inducer of
apoptosis in myeloma cells derived from multiple myeloma patients
and is not cytotoxic to hematopoietic stem cells. Leukemia.
13:1817–1824. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jazirehi AR, Ng CP, Gan XH, Schiller G and
Bonavida B: Adriamycin sensitizes the adriamycin-resistant
8226/Dox40 human multiple myeloma cells to Apo2L/tumor necrosis
factor-related apoptosis-inducing ligand-mediated (TRAIL)
apoptosis. Clin Cancer Res. 7:3874–3883. 2001.
|
|
11
|
Vitovski S, Chantry AD, Lawson MA and
Croucher PI: Targeting tumour-initiating cells with TRAIL based
combination therapy ensures complete and lasting eradication of
multiple myeloma tumours in vivo. PLoS One. 7:e358302012.
View Article : Google Scholar
|
|
12
|
Hermiston TW and Kuhn I: Armed therapeutic
viruses: strategies and challenges to arming oncolytic viruses with
therapeutic genes. Cancer Gene Ther. 9:1022–1035. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang ZL, Zou WG, Luo CX, et al: An armed
oncolytic adenovirus system, ZD55-gene, demonstrating potent
antitumoral efficacy. Cell Res. 13:481–489. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong F, Wang L, Davis JJ, et al:
Eliminating established tumor in nu/nu nude mice by a tumor
necrosis factor-α-related apoptosis-inducing ligand-armed oncolytic
adenovirus. Clin Cancer Res. 12:5224–5230. 2006.PubMed/NCBI
|
|
15
|
Pei Z, Chu L, Zou W, et al: An oncolytic
adenoviral vector of Smac increases antitumor activity of TRAIL
against HCC in human cells and in mice. Hepatology. 39:1371–1381.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang SB, Tan Y, Lei W, et al: Complete
eradication of xenograft hepatoma by oncolytic adenovirus ZD55
harboring TRAIL-IETD-Smac gene with broad antitumor effect. Hum
Gene Ther. 23:992–1002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng H, Jin Y, Liu H, You L, Yang C, Yang
X and Qian W: SNS-032 inhibits mTORC1/mTORC2 activity in acute
myeloid leukemia cells and has synergistic activity with perifosine
against Akt. J Hematol Oncol. 6:182013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chauhan D, Velankar M, Brahmandam M, et
al: A novel Bcl-2/Bcl-XL/Bcl-w inhibitor ABT-737 as
therapy in multiple myeloma. Oncogene. 26:2374–2380.
2007.PubMed/NCBI
|
|
19
|
Kuhn DJ, Berkova Z, Jones RJ, et al:
Targeting the insulin-like growth factor-1 receptor to overcome
bortezomib resistance in preclinical models of multiple myeloma.
Blood. 120:3260–3270. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tagoug I, Sauty De Chalon A and Dumontet
C: Inhibition of IGF-1 signalling enhances the apoptotic effect of
AS602868, an IKK2 inhibitor, in multiple myeloma cell lines. PLoS
One. 6:e226412011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ikeda H, Hideshima T, Fulciniti M, et al:
PI3K/p110δ is a novel therapeutic target in multiple myeloma.
Blood. 116:1460–1468. 2010.PubMed/NCBI
|
|
22
|
Maiso P, Liu Y, Morgan B, et al: Defining
the role of TORC1/2 in multiple myeloma. Blood. 118:6860–6870.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park S, Chapuis N, Tamburini J, et al:
Role of the PI3K/AKT and mTOR signaling pathways in acute myeloid
leukemia. Haematologica. 95:819–828. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oh WJ and Jacinto E: mTOR complex 2
signaling and functions. Cell Cycle. 10:2305–2316. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hougardy BM, Maduro JH, van der Zee AG, de
Groot DJ, van den Heuvel FA, de Vries EG and de Jong S: Proteasome
inhibitor MG132 sensitizes HPV-positive human cervical cancer cells
to rhTRAIL-induced apoptosis. Int J Cancer. 118:1892–1900. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seol DW: p53-Independent up-regulation of
a TRAIL receptor DR5 by proteasome inhibitors: a mechanism for
proteasome inhibitor-enhanced TRAIL-induced apoptosis. Biochem
Biophys Res Commun. 416:222–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bais S, Bartee E, Rahman MM, McFadden G
and Cogle CR: Oncolytic virotherapy for hematological malignancies.
Adv Virol. 2012:1865122012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Raus S, Coin S and Monsurrò V: Adenovirus
as a new agent for multiple myeloma therapies: opportunities and
restrictions. Korean J Hematol. 46:229–238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Naik S, Nace R, Federspiel MJ, Barber GN,
Peng KW and Russell SJ: Curative one-shot systemic virotherapy in
murine myeloma. Leukemia. 26:1870–1878. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Senac JS, Doronin K, Russell SJ, Jelinek
DF, Greipp PR and Barry MA: Infection and killing of multiple
myeloma by adenoviruses. Hum Gene Ther. 21:179–190. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen CY, Senac JS, Weaver EA, et al:
Species D adenoviruses as oncolytics against B-cell cancers. Clin
Cancer Res. 17:6712–6722. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Catlett-Falcone R, Landowski TH, Oshiro
MM, et al: Constitutive activation of Stat3 signaling confers
resistance to apoptosis in human U266 myeloma cells. Immunity.
10:105–115. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tu Y, Gardner A and Lichtenstein A: The
phosphatidylinositol 3-kinase/AKT kinase pathway in multiple
myeloma plasma cells: roles in cytokine-dependent survival and
proliferative responses. Cancer Res. 60:6763–6770. 2000.PubMed/NCBI
|
|
34
|
Hsu J, Shi Y, Krajewski S, et al: The AKT
kinase is activated in multiple myeloma tumor cells. Blood.
98:2853–2855. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bruno B, Giaccone L, Rotta M, Anderson K
and Boccadoro M: Novel targeted drugs for the treatment of multiple
myeloma: from bench to bedside. Leukemia. 19:1729–1738. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sprynski AC, Hose D, Caillot L, et al: The
role of IGF-1 as a major growth factor for myeloma cell lines and
the prognostic relevance of the expression of its receptor. Blood.
113:4614–4626. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huo J, Xu S, Lin B, Chng WJ and Lam KP:
Fas apoptosis inhibitory molecule is upregulated by IGF-1 signaling
and modulates Akt activation and IRF4 expression in multiple
myeloma. Leukemia. 27:1165–1171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Strömberg T, Ekman S, Girnita L, et al:
IGF-1 receptor tyrosine kinase inhibition by the cyclolignan PPP
induces G2/M-phase accumulation and apoptosis in
multiple myeloma cells. Blood. 107:669–678. 2006.PubMed/NCBI
|
|
39
|
Thaci B, Ulasov IV, Wainwright DA and
Lesniak MS: The challenge for gene therapy: innate immune response
to adenoviruses. Oncotarget. 2:113–121. 2011.PubMed/NCBI
|
|
40
|
Tamanini A, Nicolis E, Bonizzato A,
Bezzerri V, Melotti P, Assael BM and Cabrini G: Interaction of
adenovirus type 5 fiber with the coxsackievirus and adenovirus
receptor activates inflammatory response in human respiratory
cells. J Virol. 80:11241–11254. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Verdino P, Witherden DA, Havran WL and
Wilson IA: The molecular interaction of CAR and JAML recruits the
central cell signal transducer PI3K. Science. 329:1210–1214. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
David E, Sinha R, Chen J, Sun SY, Kaufman
JL and Lonial S: Perifosine synergistically enhances TRAIL-induced
myeloma cell apoptosis via up-regulation of death receptors. Clin
Cancer Res. 14:5090–5098. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Surget S, Chiron D, Gomez-Bougie P, et al:
Cell death via DR5, but not DR4, is regulated by p53 in myeloma
cells. Cancer Res. 72:4562–4573. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Voortman J, Resende TP, Abou El Hassan MA,
Giaccone G and Kruyt FA: TRAIL therapy in non-small cell lung
cancer cells: sensitization to death receptor-mediated apoptosis by
proteasome inhibitor bortezomib. Mol Cancer Ther. 6:2103–2112.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mitsiades N, Poulaki V, Mitsiades C and
Tsokos M: Ewing’s sarcoma family tumors are sensitive to tumor
necrosis factor-related apoptosis-inducing ligand and express death
receptor 4 and death receptor 5. Cancer Res. 61:2704–2712.
2001.
|